
Effects of Aluminium Contamination on the Nervous System of Freshwater Aquatic Vertebrates: A Review


1
University Lille, CNRS, UMR 8576-UGSF-Unité de Glycobiologie Structurale et Fonctionnelle, F-59000 Lille, France
2
Normandie University, UNILEHAVRE, CNRS, UMR 3730 SCALE, Environmental Stress and Aquatic Biomonitoring (SEBIO), F-76600 Le Havre, France
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(1), 31; https://doi.org/10.3390/ijms23010031
Submission received: 13 October 2021
/
Revised: 7 December 2021
/
Accepted: 10 December 2021
/
Published: 21 December 2021
(This article belongs to the Special Issue Reproductive and Developmental Toxicology 2.0)
Abstract
:Aluminium (Al) is the most common natural metallic element in the Earth’s crust. It is released into the environment through natural processes and human activities and accumulates in aquatic environments. This review compiles scientific data on the neurotoxicity of aluminium contamination on the nervous system of aquatic organisms. More precisely, it helps identify biomarkers of aluminium exposure for aquatic environment biomonitoring in freshwater aquatic vertebrates. Al is neurotoxic and accumulates in the nervous system of aquatic vertebrates, which is why it could be responsible for oxidative stress. In addition, it activates and inhibits antioxidant enzymes and leads to changes in acetylcholinesterase activity, neurotransmitter levels, and in the expression of several neural genes and nerve cell components. It also causes histological changes in nerve tissue, modifications of organism behaviour, and cognitive deficit. However, impacts of aluminium exposure on the early stages of aquatic vertebrate development are poorly described. Lastly, this review also poses the question of how accurate aquatic vertebrates (fishes and amphibians) could be used as model organisms to complement biological data relating to the developmental aspect. This “challenge” is very relevant since freshwater pollution with heavy metals has increased in the last few decades.
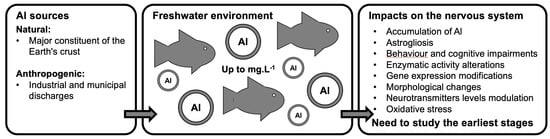
1. Introduction
Since the nineteenth century, anthropogenic activities have significantly altered ecosystems and triggered the sixth biodiversity crisis [1]. One of the main causes of this biodiversity erosion is the release of micropollutants of diverse nature and origins [2,3,4]. Once emitted, contaminants can reach non-target areas via several kinds of transport (e.g., runoffs, wet and dry deposition, long range transports), where they can have hazardous impacts on biodiversity. For example, the pollution of surface waters causes significant environmental and health issues [5,6,7].
Toxic metals, including aluminium (Al), negatively affect aquatic organisms [6,8,9]. Al is the third most common mineral and the most prevalent natural metallic element in the Earth’s crust, accounting for 8.1% of the Earth’s mass [10]. It naturally occurs exclusively in the +3-oxidation state (Al3+) in combination with other elements such as oxygen, silicon, and fluorine [11,12,13]. Al3+ is the major component of a large number of minerals, including mica, feldspars, and clays [12], and is naturally released into the environment through the weathering of rocks or minerals or through volcanic activities [13]. Produced by electrolysis from bauxite, Al is commercially manufactured under various forms, including particles in paints, pigments, and coatings, and it is used as a catalyst in the chemical and paper industries or textile dyeing. [13]. It has many industrial applications, particularly in electrical engineering, transportation, construction, and in the manufacture of household utensils, appliances, and packaging materials [11,13]. Aluminium sulphate (Al2(SO4)3) is widely used to improve the clarity of drinking water [14], and various Al compounds are used in processing, packaging, and the preservation of food [15]. In addition, Al has cosmetic and medical applications. It is found in antiperspirants, antacids and adjuvants for vaccines, toxoids, or used in patients with kidney failure to prevent hyperphosphatemia [16,17,18].
Due to the large number of natural and anthropogenic sources, Al is abundant in the environment. It has incompatible properties with fundamental life processes [12,19] and displays harmful effects in living organisms. In fact, Al is responsible for oxidative stress, cytotoxicity, genotoxicity, pro-inflammatory effects, immunological alterations, peptide denaturation or transformation, enzymatic dysfunctions, metabolic derangements, membrane disruption, microtubule perturbation, iron dyshomeostasis, amyloidogenesis, apoptosis, necrosis, and dysplasia [20]. Studies on animals have also shown that Al is neurotoxic and targets the central nervous system [11,19,21,22,23,24] by crossing the blood–brain barrier or by being transported through olfactory nerves [25]. In a rodent model, Al causes neurodegeneration, nerve cell death, changes in acetylcholinesterase (AChE) and neurotransmitter levels, histopathological changes (such as neuronal vacuolisation), and impaired cognitive and locomotor performances [12,24,25,26,27,28]. In humans, it is known to be associated with many pathologies of the nervous system, such as Alzheimer’s and Parkinson’s diseases, dementia, and autism [20].
While Al exposure is recognised to reduced survival, reproduction, and growth rates in fish and amphibians [10,29,30,31,32,33,34,35,36,37,38,39,40], only a few studies have addressed the neurotoxicity impacts in aquatic vertebrates. However, damages to the nervous system could alter the relational functions of organisms, threatening their survival, reproduction, and, ultimately, the population dynamics. Therefore, it appears of major importance to characterise the effects and action mechanisms of this contaminant on the nervous system of aquatic vertebrates. This literature review reports the effects of Al on the nervous system of freshwater aquatic vertebrates. It also poses the question of accurate aquatic vertebrates as model organisms that could complement the biological data relating to the developmental aspect.
2. Aluminium in Surface Freshwater
As a major constituent of the Earth’s crust, Al’s natural release into the environment exceeds those resulting from human activities [41]. However, its concentration in surface waters is increased by human activities, such as industrial and municipal discharges and Al2(SO4)3 is also found in drinking water [21,42,43].
Properties of Al in soil and water, such as persistence, mobility, chemical reactivity, and sorption dynamics, are governed by physicochemical and geological parameters, such as pH, temperature, organic matter, and suspended matters [44,45,46], which also directly affect its bioavailability. Dissolved Al concentrations in surface waters are highly variable and strongly influenced by the pH and the amount of dissolved organic matter (DOM) [11,44]. Al and its derivatives are poorly soluble in water at pH comprised between 6 and 8, which is the case for most natural surface waters [12]. Nevertheless, recent environmental monitoring campaigns revealed its occurrence at concentrations exceeding the World Health Organization and United States Environmental Protection Agency standards (0.2 mg·L−1). Indeed, in 2010, Al was found in rivers and lakes sampled all around the world at 1.2 ± 0.8 mg·L−1 (n = 9) and at 1.6 ± 1 mg·L−1 (n = 8), and it could reach a mean concentration of 3.1 ± 1.9 mg·L−1 in waterbodies (n = 5) sampled in Asia [47]. Usually, high concentrations in natural waters are only observed when the water pH is below 5. Therefore, concentrations in most surface freshwaters (e.g., ponds, lakes, and streams) with a pH greater than 5.5 are less than 0.1 mg·L−1 [13,21]. However, acidification of freshwater ecosystems leads to Al mobilization. Strong pH depressions have an anthropogenic origin, resulting from acidifying mine drainage, rain, and fertilisers [12,48], but can also be natural with snowmelt in spring or erosion caused by storms [28,49]. In sulphide-rich regions, water is strongly acidic (pH less than 3.5) and soluble Al concentrations are close to 50 mg·L−1 [21] and can reach 90 mg·L−1 due to acid mine drainage and discharge [46]. Additionally, in urban and industrial areas, high concentrations are regularly quantified [13].
Variations in Al toxicity are also observed according to pH or DOM fluctuation. When the pH is below 5.5, exposure to low concentration of Al (0.0125 mg·L−1) causes severe physiological alterations in Oncorhynchus mykiss [29]. In Danio rerio, exposure to 0.05 mg·L−1 of aluminium sulphate increases AChE activity in the brain at pH 5.8 but not at pH 6.8 [50]. Similarly, waters with high contents of DOM, such as humic or fulvic acid, promote the dissolution of Al and its derivatives (aluminium oxide and aluminium salts) [21]. Basically, DOM increases Al solubility while decreasing its toxicity [44]. High levels of suspended particles, which can be caused by storms, also strongly modify Al concentrations in surface waters by making new sites of adsorption available [51].
The behaviour of Al in aquatic ecosystems is strongly influenced by its binding chemistry. It can be suspended or dissolved as a monomer or a polymer, in the form of a free ion, and complexed to water molecules or bound to organic or inorganic ligands and negatively charged functional groups on humic materials and clay [46]. Different salts of Al can be found: aluminium oxide, chlorohydrate, hydroxide, fluoride, chloride, sulfate, lactate, phosphate, and nitrate [44,46]. Aluminium hydroxide and aluminium fluoride are the most important inorganic species in natural waters, although aluminium phosphate may be important for aluminium-treated wastewater [52]. Except for aluminium phosphide, anionic components (e.g., fluoride, chloride, and nitrate) do not affect the toxicity, although they affect the bioavailability [21]. The toxicity is decreased in ligands—complexed forms such as organic acids, fluoride, sulphate and silicate—and solely the monomeric inorganic form contributes to acute toxicity [45].
Aluminium speciation depends on several factors, including concentrations of dissolved organic carbon, fluoride, sulphate and phosphate, suspended particles, and water temperature and pH [44,53]. All parameters significantly alter its bioavailability and toxicity [53]. As previously specified, the toxicity increases at low pH (5.5) due to changes in speciation [45]. In water, for acidic pH values below 4, the dominant speciation corresponds to the oxidation state Al3+ and is generally in the form of a hydrated complex, Al(H2O)63+. For a pH between 5 and 6, the Al2(OH)24+ and Al(OH)52− species dominate, and Al may complex with phosphate and no longer be available. The insoluble form Al(OH)3 is a predominant form in the pH range between 5.2 and 8.8. Above pH 9, the soluble species Al(OH)4− is dominant and is the only one present at pH levels above 10 [21,45]. At basic pH and under non-equilibrium conditions, Al polymerises and forms Al2(OH)2(H2O)84+ and Al13(OH)327+ polycations [46]. These structures become large enough to precipitate and carry Al, reducing its mobility. In general, monomeric Al compounds are more reactive and labile than polymeric compounds. However, the above considerations are only valid when the organic matter and silica contents remain low [13]. In the presence of large amounts of DOM, particularly fulvic acid, Al binds to these substances and becomes a dissolved organic complex [21,46].
Al bioconcentration in aquatic organisms, studied in fish and amphibians [33,39,54,55,56,57,58], also depends on several parameters, including pH and organic carbon content. For example, Salvelinus fontinalis accumulates more Al at pH 5.3 than at pH 7.2 [33]. In freshwater ecosystems, toxic metals, including metalloids, are widely sorbed on surface sediments and suspended particles that modulate their speciation, dispersion, and ecotoxicology [53,59]. Since many freshwater organisms are in contact with dissolved and particulate matter fractions, they accumulate Al from both water and solid phases [53], despite the bioaccumulation potential appearing low [12].
3. Effects of Aluminium on the Nervous System of Freshwater Aquatic Vertebrates
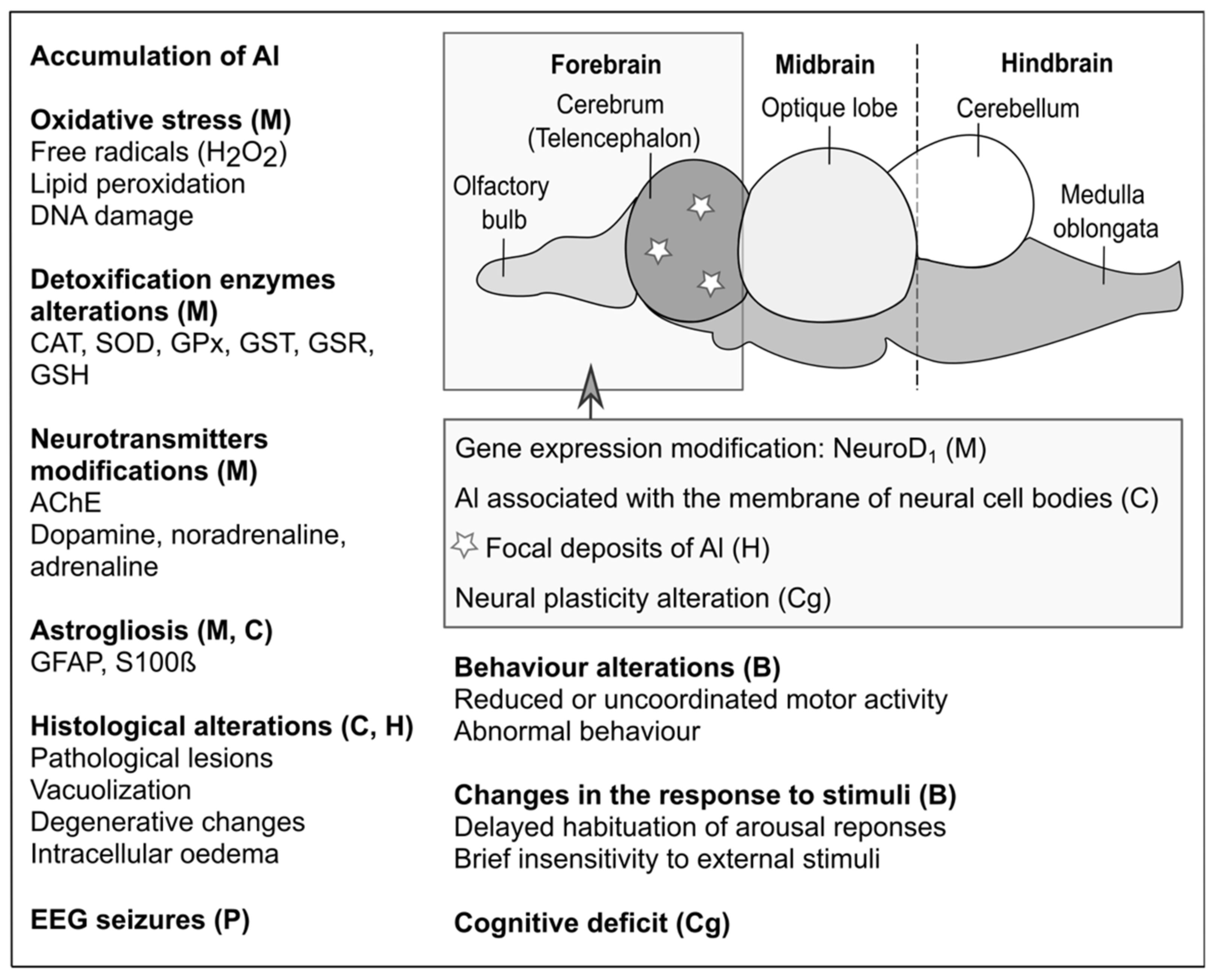
The neurotoxic action of Al impacts motor and cognitive capabilities. At the cellular level, several important mechanisms are affected: axonal transport, neurotransmitter synthesis, synaptic transmission, calcium homeostasis, energy metabolism, inflammatory responses, cell death, and glial cell activation [27]. At the molecular level, serious modifications occur in protein phosphorylation/dephosphorylation and degradation, gene expression, DNA repair, formation of reactive oxygen species, antioxidant enzyme activity, NF-kB and JNK pathways, and DNA binding [27]. However, these changes are essentially observed in mammals, and only a few studies have addressed the effects produced on the nervous system of aquatic vertebrates. Table 1 and Figure 1 report the effects of Al on the nervous system of several aquatic vertebrates.
The nervous system of aquatic vertebrates can accumulate Al, as proven in various species. For example, accumulation was observed in the brain of Cirrhinus mrigala, Ctenopharyngodon idella, and Oncorhynchus mykiss exposed to Al2(SO4)3 [55,58,66]. For C. mrigala, this observation could be due to a dysfunction of the liver, and thus, of a detoxification process, where Al was also detected [58]. In O. mykiss exposed to environmental concentrations, small deposits on the apical surface of the cerebrovascular endothelium and in the telencephalon indicated that Al crossed the blood–brain barrier. In the telencephalon, it was intimately associated with the membrane of neuronal cell bodies in the form of diffuse deposits surrounding the brain capillaries. In addition, cell bodies contained several distinct types of neural debris [55]. Accumulation is regulated by absorption and excretion rates, toxicant concentration, and exposure duration [54,58]. For instance, in C. mrigala, the rate of absorption and the biomagnification factor was higher, while the rate of excretion was lower in chronic compared to acute exposures [58]. In contrary to the previous studies, Anandhan and Hemalatha [54] did not detect Al accumulation in the brain of D. rerio exposed to 5.69 and 17.08 ppm of AlCl3, while accumulation occurred in the liver, gills, and muscles.
As shown in Table 1, most studies focused on the assessment of oxidative stress (which results in high production of free radicals) of Al on the nervous system. Al replaces iron in various biomolecules and increases intracellular iron concentrations, promoting a Fenton oxidation reaction [71,72]. Additionally, it disrupts the electron transport chain in mitochondria [66] and generates oxidising radicals. Oxidative stress is deleterious to organisms because it leads to protein and enzyme inactivation, lipid peroxidation, and DNA damages. Fish nervous tissue is particularly sensitive to Al-induced oxidative actions because of its richness in polyunsaturated fatty acids and high consumption of oxygen (about 1/5 of the total consumption) [62]. Therefore, oxidative damages of the nervous tissue are one of the main mechanisms leading to the toxic effects of Al [62]. In Lepomis gibbosus, oxidative stress occurred in nerve tissue [70]. In D. rerio, Lepomis macrochirus, Rutilus rutilus, Carassius carassius, and Neogobius fluviatilis, a significant increase in the level of brain lipid peroxidation was seen after exposure to AlCl3 [70,73]. This accumulation was also observed in C. idella exposed to Al2(SO4)3 [66] and in Oreochromis mossambicus after exposure to Al oxide nanoparticles (Al2O3NPs) [67].
Al-induced oxidative stress alters the activity of antioxidant enzymes. The enzyme activity, initially increased to compensate for the oxidative stress, is depleted by extended exposure, leading to protein and DNA damages [66]. A significant decrease in the brain catalase (CAT) activity was observed in Channa punctatus, C. idella, and O. mossambicus exposed to AlCl3, Al2(SO4)3, and Al2O3NPs, respectively [61,66,67]. The decrease was linked to the production of glutathione peroxidase (GPx), an antioxidant enzyme, in competition with CAT for the common hydrogen peroxide (H2O2) substrate [74] and with the establishment of non-enzymatic mechanisms as, for example, the sequestration of oxidant radicals by metallothioneins [75]. Another explanation is the inhibition of CAT by an Al ion capable of binding the enzyme thiol groups [66]. Additionally, the decrease in CAT activity could be explained by a decrease in gene expression [76]. Finally, antioxidant enzymes may themselves undergo oxidative changes [67,77]. In D. rerio, CAT activity increased significantly in the brain after long-term exposure to AlCl3. This increase reflects the need for a greater amount of antioxidant enzymes to eliminate free radicals produced during Al long-term exposure [61], as CAT. The activity of the superoxide dismutase (SOD) which neutralises oxidising radicals and converts superoxide ions [78] into H2O2 [66], is altered by aluminium exposure. On one hand, a significant increase in SOD activity was detected in the brain of L. macrochirus, R. rutilus, C. carassius, and N. fluviatilis after exposure to AlCl3 [62] and C. idella exposed to Al2(SO4)3 [66]. On the other hand, in O. mossambicus exposed to Al2O3NPs, a significant decrease in the brain of SOD, GPx, and glutathione S-transferase (GST) activities and an increase in the level of H2O2 were observed by Vidya and Chitra [67]. The decrease in SOD activity may result from the generation of an excess of oxidising radicals following the exposure to nanoparticles, which could then lead to inactivation of the enzyme. The oxidant radicals would further decrease the activity of other antioxidant enzymes, such as CAT, GPx, or GSR, decreasing the neutralization potential of oxidant radicals and increasing lipid peroxidation [67,77]. Finally, GST, another antioxidant enzyme also involved in tissue protection from oxidative stress and damages [79], increases its activity in response to a rise in free radicals [67]. It results in a GSH decrease, which normally acts as a GST cofactor to neutralise oxidising radicals [80]. A significant rise in GST activity and a decrease in reduced glutathione (GSH) content were generated in O. mossambicus exposed to Al2O3NPs [67].
Another harmful effect of Al on the nervous system of aquatic vertebrates is the alteration of AChE activity, a key nervous system hydrolase that catalyses the hydrolytic metabolism of the neurotransmitter acetylcholine (ACh) into choline and acetate [67]. AChE is usually used as a biomarker of effects for the central nervous system [81]. In fish, AChE activity is essential for muscle function and behaviour [82]. A significant increase in the enzyme activity was observed in D. rerio following exposure to AlCl3 [50,61] and in Oreochromis niloticus after exposure to AlCl3 and Al2(SO4)3 [65]. According to Maheswari et al. [61], AChE increased in activity in D. rerio after short exposure times and in quantity for longer exposure times. The increased activity could be due to an allosteric interaction between the anionic peripheral site of AChE and Al3+ ions [83], an increase in the production of free radicals [61,77], or a conformational change consecutively to the peroxidation of the membrane lipids of the brain cells [84]. In contrast, a significant decrease in AChE activity was observed in O. mossambicus following exposure to Al2O3NPs [67]. Al neurotoxicity also results in altered levels of brain neurotransmitters. In C. idella exposed to Al2O3NPs, a significant increase in dopamine and noradrenaline content was observed by Fernández-Dávila et al. [66], while the adrenaline content significantly decreased. The observed changes in these neurotransmitter levels could be related to their synthesis. These three neurotransmitters are derived from tyrosine. Dopamine is converted into noradrenaline, which is further converted into adrenaline. Enzymes catalysing these transformations are probably affected by the binding of Al to the thiol groups [66]. As mentioned previously, the induction of oxidising radicals may never be responsible for direct damages on the enzymes or indirect actions on the corresponding genes (in C. idella brain, [85]). The synthesis of catecholamines, which include dopamine, noradrenaline, and adrenaline, is sequential, and inhibition of the final stages probably increases the content of noradrenaline and dopamine, as seen in C. idella [66].
At the genetic level, a decrease in the production of NeuroD1 mRNA, involved in the regulation and the control of nerve differentiation, was observed in Salmo salar exposed to AlCl3 and was probably due to an increased level of stress [63,86]. Additionally, chromatin and DNA are particularly vulnerable to Al3+ [87]. Al ions strongly bind DNA, RNA, and mononucleotides [12,88,89]. In L. macrochirus, R. rutilus, C. carassius, and N. fluviatilis, exposure to AlCl3 induced an overexpression of glial fibrillary acidic protein (GFAP), a subunit of the cytoskeleton intermediate filaments, and S100β, a calcium-binding protein mainly present in astrocytes [62,90]. This overexpression correlated with an increase in the content of the lysed forms of GFAP and S100β fragments. This indicates that Al ions could activate intracellular proteases which alter intermediate filaments in astrocytes [62], as in D. rerio exposed to AlCl3 [37]. The overexpression of GFAP and S100β may be responsible for astrogliosis in N. fluviatilis exposed to AlCl3 [62] and in L. gibbosus [70]. Astrogliosis are changes characterised by an overexpression of GFAP, that occur in astrocytes in response to central nervous tissue injuries and damages induced by toxic substances in the brain of many vertebrates [62]. This glial cell reactivity is commonly used as a biomarker to detect nerve tissue disorders [90].
Morphological changes in tissues are considered as signs of various pathologies. In aquatic ecosystems, chronic exposure to contaminants at sublethal concentrations can alter the structural architecture of tissues without killing fish. Such structural tissue changes were observed by Vidya and Chitra [68] in the brain of O. mossambicus exposed to 4 mg−1 of Al2O3NPs (sublethal concentration). After 96 h of exposure, moderate degenerative changes occurred in all cerebral regions associated with a slight vacuolisation in the neural cells. After 60 days of treatment, severe degenerative changes and intracellular oedema were noted. As previously mentioned, Al2O3NPs can cross the blood–brain barrier, accumulate in nerve tissue, and induce damages to the brain [68]. These results are therefore in agreement with results obtained in C. idella by Sivakumar et al. [58] and in O. mykiss by Exley [55]. Vidya and Chitra [67,68], showing that deleterious effects of Al2O3NPs in O. mossambicus are persistent after cessation of exposure, indicating the irreversible neurotoxic properties of Al nanoparticles.
Finally, several studies have highlighted the impacts of Al on the behaviour of aquatic vertebrates in connection with an alteration of the nervous system. In D. rerio exposed to AlCl3, a significant decrease in the locomotor activity was demonstrated. A decrease in the distance travelled, a reduction of the maximum speed, and an increase in the absolute angle of rotation were mentioned [50]. The involvement of the cholinergic system in the locomotor activity, the response to new stimuli, and the performance of spatial memory tasks was fully established [91]. This implies that the induction of AChE activity in the brain observed in D. rerio may be responsible for the behavioural and neurotoxic effects of Al on the central nervous system [50]. Fish activity may also be limited by their compromised ability to extract oxygen from water. Al is believed to interfere with oxygen supply to tissues by causing osmoregulatory and ion-regulatory dysfunction and changing the haematological status [92,93]. In D. rerio larvae exposed to AlCl3, Capriello et al. [60] observed a significant decrease in the average of moved distance, velocity, time of movement, and number of heading at low concentrations (below 100 µM), with a recovery at high concentrations (100 and 200 µM). The impairment of the swimming ability of D. rerio larvae was probably caused by a reduction of the number of neural stem cell—limiting neuroblast differentiation [94]—and/or alteration of the glucose metabolism [95]. In S. salar exposed to AlCl3, Grassie et al. [63] observed an increased number of errors made by individuals in a maze, indicating a decrease in their spatial learning capabilities. Cognitive deficits are associated with a decrease in neuronal plasticity of the forebrain, and in NeuroD1, by an mRNA expression in the telencephalon [63]. Laming et al. [69] showed that a topical application of Al(OH)3 on the telencephalon of R. rutilus induces unusual and gentle lateral undulations of the body and a sporadic, violent, uncoordinated motor activity. These effects were associated with a delayed habituation of arousal responses to repeated presentations of two stimuli and the presence of electroencephalographic seizures in which the EEG amplitude was elevated from 4–20 times compared to a normal level. Even though R. rutilus lacks a cerebral cortex and has a relatively undifferentiated telencephalon, observed seizures are an expression of the malfunction of a fundamental mechanism, as in other vertebrate brains. Seizures correlate with over-activity of the brain, which normally operates during arousal [69]. The same topical application of Al(OH)3 in Carassius carassius induced a delayed habituation of cardiac arousal to a moving shadow stimulus [69]. Finally, Andrén et al. [96] showed that the swimming behaviour of the moor frog Rana arvalis is disturbed by environmentally relevant concentrations of AlCl3, while the behaviour of the common frog Rana temporaria and the agile frog Rana dalmatina exposed to the same concentrations are not. Altogether, aquatic vertebrates’ behaviour changes have serious consequences: limited survival in the wild [64] and affected swimming activity, predation, migration, and reproductive success [92].
4. Perspectives: Interests of Biological Models to Study the Effects of Aluminium on the Nervous System of Aquatic Vertebrates
Due to the permeable properties of the blood–brain barrier, the central nervous system is one of the major targets of Al in freshwater species. Several questions remain concerning the doses, the exposure times, and the sensitivity of the embryo developmental stages required to trigger a toxic effect. To date, no data exist on Al accumulation and cumulative/additive effects during specific periods of the neural system development. Using aquatic vertebrate models to perform dose–response tests, time-lapse exposures, and behavioural assessments in early developmental stages could provide precious pieces of information on Al toxicity. Studies on critical exposure phases could be precisely determined for the development of the central nervous system. For instance, the different phases of the neural embryonic development ranging from the early neural plaque induction and tube folding to the late formation of the neurogenic territories of the brain regions could have different sensitivity and accumulation rates. Moreover, these developmental parameters could generate important data and lead to the determination of sensitive toxicity periods and specific markers. There are advantages to using organisms such as D. rerio or Xenopus sp. in environmental toxicology studies [97,98,99]. Both models share a short life cycle [100,101], which can be studied from the oogenesis period to the late development in controlled conditions. The developmental stages, molecular signalling, genetic compositions, and the neurodevelopmental processes of both models are well characterised [102,103]. Both embryo nervous systems are visible by transparency and easily accessible for various in vivo recordings and studies [104]. Live-imaging at high resolution with structural and dynamical details and quantification of neuronal properties are also possible [105]. The neurotoxicity endpoints can be assessed during the neural development with proteomic and genomic large-scale screenings [106]. Their cellular and molecular neuronal parameters can be analysed in relation to the behavioural abnormalities, including locomotion, foraging, and avoidance [105,107]. Given the environmental concerns related to Al, its underestimated neurotoxic impacts on freshwater organisms, and also the interesting possibilities offered by the methods widely used on well-known fish and amphibian models, additional studies would allow a better understanding of the action of Al on the neural system and, more globally, its effect at the population level.
5. Conclusions
Al is responsible for various toxic effects. This metal is well-known for its neurotoxicity in mammalian models, but only a few studies have been conducted on aquatic organisms. However, due to the large number of natural and anthropogenic sources, Al is abundant in the environment and can be found in aquatic ecosystems. Previous works have shown that Al accumulates in the nervous system of freshwater vertebrates, where it can trigger oxidative stress, alter enzymatic activities, and neurotransmitters levels but also affect gene expression, cause astrogliosis and morphological changes, and impair behaviour and cognitive abilities. These effects were primarily studied in adult organisms without considering early stages of development, which are critical windows of exposure. In conclusion, further studies are needed to better characterise Al neurotoxic effects during whole developmental processes with the determination of the critical periods of time, duration, and the quantities that threaten freshwater life. Thus, Xenopus and Zebrafish could be valuable model organisms since their development are external and easily accessible. Sequential and additive exposures could be undertaken to understand the toxic mechanisms of the action of aluminium on the embryonic development of the nervous system and propose molecular signatures associated with functional states of media contaminated by this metal.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Acknowledgments
The authors are thankful to the Research Federation FRABio (University Lille, CNRS, FR 3688, FRABio, Biochimie Structurale et Fonctionnelle des Assemblages Biomoléculaires).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Monastersky, R. Biodiversity: Life—A status report. Nat. News 2014, 516, 158. [Google Scholar] [CrossRef] [PubMed]
- Beketov, M.A.; Kefford, B.J.; Schäfer, R.B.; Liess, M. Pesticides reduce regional biodiversity of stream invertebrates. Proc. Natl. Acad. Sci. USA 2013, 110, 11039–11043. [Google Scholar] [CrossRef] [Green Version]
- Dudgeon, D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr. Biol. Acad. 2019, 29, R960–R967. [Google Scholar] [CrossRef] [PubMed]
- Dudley, N.; Alexander, S. Agriculture and biodiversity: A review. Biodiversity 2017, 18, 45–49. [Google Scholar] [CrossRef]
- Adjagodo, A.; Tchibozo MA, D.; Kelome, N.C.; Lawani, R. Flux des polluants liés aux activités anthropiques, risques sur les ressources en eau de surface et la chaine trophique à travers le monde: Synthèse bibliographique. Int. J. Biol. Chem. Sci. 2016, 10, 1459–1472. [Google Scholar] [CrossRef]
- Scott, G.R.; Sloman, K.A. The effects of environmental pollutants on complex fish behaviour: Integrating behavioural and physiological indicators of toxicity. Aquat. Toxicol. 2004, 68, 369–392. [Google Scholar] [CrossRef] [PubMed]
- Vijayaraman, S.; Mondal, P.; Nandan, A.; Siddiqui, N.A. Presence of microplastic in water bodies and its impact on human health. In Advances in Air Pollution Profiling and Control; Springer: Singapore, 2020; pp. 57–65. [Google Scholar]
- Correia, T.G.; Narcizo, A.D.M.; Bianchini, A.; Moreira, R.G. Aluminium as an endocrine disruptor in female Nile tilapia (Oreochromis niloticus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 151, 461–466. [Google Scholar] [CrossRef]
- Sfakianakis, D.G.; Renieri, E.; Kentouri, M.; Tsatsakis, A.M. Effect of heavy metals on fish larvae deformities: A review. Environ. Res. 2015, 137, 246–255. [Google Scholar] [CrossRef]
- Sparling, D.W.; Lowe, T.P. Environmental hazards of aluminium to plants, invertebrates, fish, and wildlife. Rev. Environ. Contam. Toxicol. 1996, 145, 1–127. [Google Scholar]
- Jones, K.C.; Bennett, B.G. Exposure of man to environmental aluminium—an exposure commitment assessment. Sci. Total Environ. 1986, 52, 65–82. [Google Scholar] [CrossRef]
- Ganrot, P.O. Metabolism and possible health effects of aluminium. Environ. Health Perspect. 1986, 65, 363–441. [Google Scholar]
- INERIS. Aluminium et Dérivés; Institut National de l’Environnement Industriel et des Risques—Fiche de Données Toxicologiques et Environnementales des Substances Chimiques, INERIS: Creil, France, 2005. [Google Scholar]
- Pernitsky, D.J.; Edzwald, J.K. Selection of alum and polyaluminum coagulants: Principles and applications. J. Water Supply Res. Technol. AQUA 2006, 55, 121–141. [Google Scholar] [CrossRef]
- Stahl, T.; Taschan, H.; Brunn, H. Aluminium content of selected foods and food products. Environ. Sci. Eur. 2011, 23, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Exley, C. Aluminium and medicine. Molecular and Supramolecular. Bioinorg. Chem. 2008, 6, 1–24. [Google Scholar]
- Shaman, A.M.; Kowalski, S.R. Hyperphosphatemia Management in Patients with Chronic Kidney Disease. Saudi Pharm. J. 2016, 24, 494–505. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Zhu, H.; Xia, X.; Liang, Z.; Ma, X.; Sun, B. Vaccine adjuvants: Understanding the structure and mechanism of adjuvanticity. Vaccine 2019, 37, 3167–3178. [Google Scholar] [CrossRef]
- Singla, N.; Dhawan, D.K. Zinc modulates aluminium-induced oxidative stress and cellular injury in rat brain. Metallomics 2014, 6, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Igbokwe, I.O.; Igwenagu, E.; Igbokwe, N.A. Aluminium toxicosis: A review of toxic actions and effects. Interdiscip. Toxicol. 2019, 12, 45–70. [Google Scholar] [CrossRef] [Green Version]
- ATSDR. Toxicological Profiles for Aluminium; US Department of Health and Human Services, Public Health Services, Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2008.
- Turner, M.; Mutter, S.T.; Kennedy-Britten, O.D.; Platts, J.A. Molecular dynamics simulation of aluminium binding to amyloid-β and its effect on peptide structure. PLoS ONE 2019, 14, 1–14. [Google Scholar] [CrossRef]
- Wang, L.; Hu, J.; Zhao, Y.; Lu, X.; Zhang, Q.; Niu, Q. Effects of aluminium on β-amyloid (1-42) and secretases (APP-cleaving enzymes) in rat brain. Neurochem. Res. 2014, 39, 1338–1345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, W.; Xu, F.; Cao, Z.; Jia, F.; Li, Y. Iron Dyshomeostasis Participated in Rat Hippocampus Toxicity Caused by Aluminum Chloride. Biol. Trace Elem. Res. 2019, 197, 580–590. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Zhang, Q.; Li, H.; Wang, L.; Lu, X. The immunotoxicity and neurotoxicity of aluminium. Environ. Occup. Health Ser. 2018, 1708, 1–122. [Google Scholar]
- Hao, S.; Li, Y.F.; Hu, C.W.; Yue, S.; Li, G. Effects of sub-chronic aluminium intoxication on apoptosis of cerebrum neurocytes in chickens: Preliminary study on effect of aluminium accumulation on nervous system in chickens. In Proceedings of the International Conference on Bioinformatics and Biomedical Engineering, Chengdu, China, 18–20 June 2010. [Google Scholar]
- Muhammad, M.S.; Ayo, J.O.; Danjuma, N.M.; Abdul Wahab, A.; Isa, A.S.; Maina, M.B. Molecular mechanisms of aluminium neurotoxicity in animal models of Alzheimer’s disease. J. Afr. Assoc. Physiol. Sci. 2019, 7, 70–79. [Google Scholar]
- Rosseland, B.O.; Eldhuset, T.D.; Staurnes, M.J.E.G. Environmental effects of aluminium. Environ. Geochem. Health 1990, 12, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Allin, C.J.; Wilson, R.W. Effects of pre-acclimation to aluminium on the physiology and swimming behaviour of juvenile rainbow trout (Oncorhynchus mykiss) during a pulsed exposure. Aquat. Toxicol. 2000, 51, 213–224. [Google Scholar] [CrossRef]
- Beattie, R.C.; Tyler-Jones, R. The effects of low pH and aluminium on breeding success in the frog Rana temporaria. J. Herpetol. 1992, 26, 353–360. [Google Scholar] [CrossRef]
- Bradford, D.F.; Swanson, C.; Gordon, M.S. Effects of low pH and aluminium on two declining species of amphibians in the Sierra Nevada, California. J. Herpetol. 1992, 26, 369–377. [Google Scholar] [CrossRef]
- Calevro, F.; Campani, S.; Ragghianti, M.; Bucci, S.; Mancino, G. Tests of toxicity and teratogenicity in biphasic vertebrates treated with heavy metals (Cr3+, A13+, Cd2+). Chemosphere 1998, 37, 3011–3017. [Google Scholar] [CrossRef]
- Cleveland, L.; Buckler, D.R.; Brumbaugh, W.G. Residue dynamics and effects of aluminum on growth and mortality in brook trout. Environ. Toxicol. Chem. Int. J. 1991, 10, 243–248. [Google Scholar] [CrossRef]
- Cleveland, L.; Little, E.E.; Ingersoll, C.G.; Wiedmeyer, R.H.; Hunn, J.B. Sensitivity of brook trout to low pH, low and elevated aluminum concentrations during laboratory pulse exposures. Aquat. Toxicol. 1991, 19, 303–317. [Google Scholar] [CrossRef]
- Cummins, C.P. Effects of aluminium and low pH on growth and development in Rana temporaria tadpoles. Oecologia 1986, 69, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Herkovits, J.; Castañaga, L.A.; D’Eramo, J.L.; Jourani, V.P. Living organisms influence on environmental conditions: pH modulation by amphibian embryos versus aluminum toxicity. Chemosphere 2015, 139, 210–215. [Google Scholar] [CrossRef]
- Monaco, A.; Grimaldi, M.C.; Ferrandino, I. Aluminium chloride-induced toxicity in zebrafish larvae. J. Fish Dis. 2017, 40, 629–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayer MD, J.; Reader, J.P.; Morris, R. Embryonic and larval development of brown trout, Salmo trutta L.: Exposure to aluminium, copper, lead or zinc in soft, acid water. J. Fish Biol. 1991, 38, 431–455. [Google Scholar] [CrossRef]
- Slaninova, A.; Machova, J.; Svobodova, Z. Fish kill caused by aluminium and iron contamination in a natural pond used for fish rearing: A case report. Vet. Med. 2014, 59, 573–581. [Google Scholar] [CrossRef] [Green Version]
- Stephens, F.J.; Ingram, M. Two cases of fish mortality in low pH, aluminium rich water. J. Fish Dis. 2006, 29, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Cave, M.; Zhang, C. Identifying geogenic and anthropogenic controls on different spatial distribution patterns of aluminium, calcium and lead in urban topsoil of Greater London Authority area. Chemosphere 2020, 238, 124541. [Google Scholar] [CrossRef] [PubMed]
- Eisenreich, S.J. Atmospheric input of trace metals to Lake Michigan. Water Air Soil Pollut. 1980, 13, 287–301. [Google Scholar] [CrossRef]
- His, E.; Beiras, R.; Seaman, M.N.; Pagano, G.; Trieff, N.M. Sublethal and lethal toxicity of aluminium industry effluents to early developmental stages of the Crassostrea gigas oyster. Arch. Environ. Contam. Toxicol. 1996, 30, 335–339. [Google Scholar] [CrossRef]
- Gensemer, R.W.; Playle, R.C. The Bioavailability and Toxicity of Aluminum in Aquatic Environments. Crit. Rev. Environ. Sci. Technol. 1999, 29, 315–450. [Google Scholar] [CrossRef]
- Witters, H.E. Chemical speciation dynamics and toxicity assessment in aquatic systems. Ecotoxicol. Environ. Saf. 1998, 41, 90–95. [Google Scholar] [CrossRef]
- World Health Organization. Environmental Health Criteria. 194: Aluminium. Print. Finl. 1997, 97, 1–282. [Google Scholar]
- Zhou, Q.; Yang, N.; Li, Y.; Ren, B.; Ding, X.; Bian, H.; Yao, X. Total concentrations and sources of heavy metal pollution in global river and lake water bodies from 1972 to 2017. Glob. Ecol. Conserv. 2020, 22, e00925. [Google Scholar] [CrossRef]
- Akcil, A.; Koldas, S. Acid Mine Drainage (AMD): Causes, treatment and case studies. J. Clean. Prod. 2006, 14, 1139–1145. [Google Scholar] [CrossRef]
- Nelson, W.O.; Campbell, P.G. The effects of acidification on the geochemistry of Al, Cd, Pb and Hg in freshwater environments: A literature review. Environ. Pollut. 1991, 71, 91–130. [Google Scholar] [CrossRef]
- Senger, M.R.; Seibt, K.J.; Ghisleni, G.C.; Dias, R.D.; Bogo, M.R.; Bonan, C.D. Aluminium exposure alters behavioral parameters and increases acetylcholinesterase activity in zebrafish (Danio rerio) brain. Cell Biol. Toxicol. 2011, 27, 199–205. [Google Scholar] [CrossRef]
- Goenaga, X.; Williams, D.J. Aluminium speciation in surface waters from a Welsh upland area. Environ. Pollut. 1988, 52, 131–149. [Google Scholar] [CrossRef]
- La Zerte, B.D.; van Loon, G.; Anderson, B. Aluminum in water. Res. Issues Alum. Toxic. 1997, 17–45. [Google Scholar]
- Huang, P.M. An overview of dynamics and biotoxicity of metals in the freshwater environment. Water Qual. Res. J. 1993, 28, 1–6. [Google Scholar] [CrossRef]
- Anandhan, R.; Hemalatha, S. Bioaccumulation of aluminium in selected tissues of Zebra fish Brachydanio rerio (Ham). Nat. Environ. Pollut. Technol. 2009, 8, 751–753. [Google Scholar]
- Exley, C. Aluminium in the brain and heart of the rainbow trout. J. Fish Biol. 1996, 48, 706–713. [Google Scholar] [CrossRef]
- Monette, M.Y.; McCormick, S.D. Impacts of short-term acid and aluminum exposure on Atlantic salmon (Salmo salar) physiology: A direct comparison of parr and smolts. Aquat. Toxicol. 2008, 86, 216–226. [Google Scholar] [CrossRef]
- Oberholster, P.J.; Myburgh, J.G.; Ashton, P.J.; Coetzee, J.J.; Botha, A.M. Bioaccumulation of aluminium and iron in the food chain of Lake Loskop, South Africa. Ecotoxicol. Environ. Saf. 2012, 75, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Sivakumar, S.; Khatiwada, C.P.; Sivasubramanian, J. Bioaccumulations of aluminium and the effects of chelating agents on different organs of Cirrhinus mrigala. Environ. Toxicol. Pharmacol. 2012, 34, 791–800. [Google Scholar] [CrossRef]
- Baudo, R. Sediments: Chemistry and Toxicity of In-Place Pollutants; CRC Press: Boca Raton, FL, USA, 1990. [Google Scholar]
- Capriello, T.; Grimaldi, M.C.; Cofone, R.; D’Aniello, S.; Ferrandino, I. Effects of aluminium and cadmium on hatching and swimming ability in developing zebrafish. Chemosphere 2019, 222, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Maheswari, S.L.; Venkatakrishna Murali, R.; Balaji, R. Aluminium induced cholinotoxicity in zebra fish brain—A sequel of oxidative stress. Int. J. Adv. Res. 2014, 2, 322–335. [Google Scholar]
- Sukharenko, E.V.; Samoylova, I.V.; Nedzvetsky, V.S. Molecular mechanisms of aluminium ions neurotoxicity in brain cells of fish from various pelagic areas. Regul. Mech. Biosyst. 2017, 3, 461–466. [Google Scholar] [CrossRef]
- Grassie, C.; Braithwaite, V.A.; Nilsson, J.; Nilsen, T.O.; Teien, H.C.; Handeland, S.O.; Stefansson, S.O.; Tronci, V.; Gorisson, M.; Filk GEbbesson, L.O. Aluminium exposure impacts brain plasticity and behavior in Atlantic salmon (Salmo salar). J. Exp. Biol. 2013, 216, 3148–3155. [Google Scholar] [PubMed] [Green Version]
- Brodeur, J.C.; Økland, F.; Finstad, B.; Dixon, D.G.; McKinley, R.S. Effects of subchronic exposure to aluminium in acidic water on bioenergetics of Atlantic salmon (Salmo salar). Ecotoxicol. Environ. Saf. 2001, 49, 226–234. [Google Scholar] [CrossRef]
- Oliveira, V.M.; Assis, C.R.D.; Costa, H.M.S.; Silva, R.P.F.; Santos, J.F.; Carvalho, L.B., Jr.; Bezerra, R.S. Aluminium sulfate exposure: A set of effects on hydrolases from brain, muscle and digestive tract of juvenile Nile tilapia (Oreochromis niloticus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 191, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Dávila, M.L.; Razo-Estrada, A.C.; García-Medina, S.; Gómez-Oliván, L.M.; Piñón-López, M.J.; Ibarra, R.G.; Galar-Martínez, M. Aluminium-induced oxidative stress and neurotoxicity in grass carp (Cyprinidae—Ctenopharingodon idella). Ecotoxicol. Environ. Saf. 2012, 76, 87–92. [Google Scholar] [CrossRef]
- Jena, B.S.; Nayak, S.B.; Patnaik, B.K. Age-related effect of aluminium on the catalase activities of the brains of two species of poikilothermic vertebrates. Gerontology 2002, 48, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Vidya, P.V.; Chitra, K.C. Aluminium oxide nanoparticles induced irrevocable damages in gill, liver and brain tissues of the freshwater Fish, Oreochromis mossambicus (Peters, 1852). Int. J. Fish. Aquat. Res. 2018, 3, 13–17. [Google Scholar]
- Laming, P.R.; Rooney, D.J.; Ferguson, J. Epileptogenesis is associated with heightened arousal responses in fish. Physiol. Behav. 1987, 40, 617–624. [Google Scholar] [CrossRef]
- Novitskiy, R.A.; Sukharenko, Y.V.; Nedzvetskiy, V.S. Molecular Biomarkers of Al 3+ Effects on Induction of Oxidative Stress and Cellular Reactivation in Organism of Lepomis gibbosus (Pisces: Centrarchidae). Hydrobiol. J. 2014, 50. [Google Scholar] [CrossRef]
- Amador, F.C.; Santos, M.S.; Oliveira, C.R. Lipid peroxidation and aluminium effects on the cholinergic system in nerve terminals. Neurotox. Res. 2001, 3, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.Y.; Guo-Ross, S.X.; Bondy, S.C. The stabilization of ferrous iron by a toxic β-amyloid fragment and by an aluminium salt. Brain Res. 1999, 839, 221–226. [Google Scholar] [CrossRef]
- Vidya, P.V.; Chitra, K.C. Aluminium oxide nanoparticles induced irreversible alterations in the antioxidant defense system of the fish, Oreochromis mossambicus (Peters, 1852). Eur. J. Biomed. Pharm. Sci. 2018, 5, 1162–1170. [Google Scholar]
- Atli, G.; Alptekin, Ö.; Tükel, S.; Canli, M. Response of catalase activity to Ag+, Cd2+, Cr6+, Cu2+ and Zn2+ in five tissues of freshwater fish Oreochromis niloticus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 143, 218–224. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Semsei, I.; Rao, G.; Richardson, A. Expression of superoxide dismutase and catalase in rat brain as a function of age. Mech. Ageing Dev. 1991, 58, 13–19. [Google Scholar] [CrossRef]
- Sohal, R.S. Effect of hydrogen peroxide administration on life span, superoxide dismutase, catalase, and glutathione in the adult housefly, Musca domestica. Exp. Gerontol. 1988, 23, 211–216. [Google Scholar] [CrossRef]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Vontas, J.G.; Small, G.J.; Hemingway, J. Glutathione S-transferases as antioxidant defence agents confer pyrethroid resistance in Nilaparvata lugens. Biochem. J. 2001, 357, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Browne, R.W.; Armstrong, D. Reduced glutathione and glutathione disulfide. In Free Radical and Antioxidant Protocols; Humana Press: Totowa, NJ, USA, 1998; pp. 347–352. [Google Scholar]
- Ucán-Marín, F.; Ernst, W.; O’Dor, R.K.; Sherry, J. Effects of food borne ivermectin on juvenile Atlantic salmon (Salmo salar L.): Survival, growth, behavior, and physiology. Aquaculture 2012, 334, 169–175. [Google Scholar] [CrossRef]
- Payne, J.F.; Mathieu, A.; Melvin, W.; Fancey, L.L. Acetylcholinesterase, an old biomarker with a new future? Field trials in association with two urban rivers and a paper mill in Newfoundland. Mar. Pollut. Bull. 1996, 32, 225–231. [Google Scholar] [CrossRef]
- Gulya, K.; Rakonczay, Z.; Kasa, P. Cholinotoxic effects of aluminium in rat brain. J. Neurochem. 1990, 54, 1020–1026. [Google Scholar] [CrossRef]
- Kaizer, R.R.; Corrêa, M.C.; Spanevello, R.M.; Morsch, V.M.; Mazzanti, C.M.; Gonçalves, J.F.; Schetinger, M.R. Acetylcholinesterase activation and enhanced lipid peroxidation after long-term exposure to low levels of aluminium on different mouse brain regions. J. Inorg. Biochem. 2005, 99, 1865–1870. [Google Scholar] [CrossRef] [PubMed]
- Borg, D.C.; Schaich, K.M. Cytotoxicity from Coupled Redox Cycling of Autoxidizing Xenobiotics and Metals: A Selective Critical Review and Commentary on Work-in-Progress. Isr. J. Chem. 1984, 24, 38–53. [Google Scholar] [CrossRef]
- Lee, J.E.; Hollenberg, S.M.; Snider, L.; Turner, D.L.; Lipnick, N.; Weintraub, H. Conversion of Xenopus ectoderm into neurons by NeuroD, a basic helix-loop-helix protein. Science 1995, 268, 836–844. [Google Scholar] [CrossRef]
- Pereira, S.; Cavalie, I.; Camilleri, V.; Gilbin, R.; Adam-Guillermin, C. Comparative genotoxicity of aluminium and cadmium in embryonic zebrafish cells. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2013, 750, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Lari, M.; Biver, T.; Busto, N.; Lozano, H.J.; Leal, J.M.; Secco, F.; García, B. Binding of Al(iii) to synthetic RNA and metal-mediated strand aggregation. Dalton Trans. 2017, 46, 16671–16681. [Google Scholar] [CrossRef]
- Wu, J.; Du, F.; Zhang, P.; Khan, I.A.; Chen, J.; Liang, Y. Thermodynamics of the interaction of aluminum ions with DNA: Implications for the biological function of aluminum. J. Inorg. Biochem. 2005, 99, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Tykhomyrov, A.A.; Pavlova, A.S.; Nedzvetsky, V.S. Glial fibrillary acidic protein (GFAP): On the 45th anniversary of its discovery. Neurophysiology 2016, 48, 54–71. [Google Scholar] [CrossRef]
- Pepeu, G.; Giovannini, M.G. Changes in acetylcholine extracellular levels during cognitive processes. Learn. Mem. 2004, 11, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Allin, C.J.; Wilson, R.W. Behavioural and metabolic effects of chronic exposure to sublethal aluminum in acidic soft water in juvenile rainbow trout (Oncorhynchus mykiss). Can. J. Fish. Aquat. Sci. 1999, 56, 670–678. [Google Scholar] [CrossRef]
- Camargo, M.M.; Fernandes, M.N.; Martinez, C.B. How aluminium exposure promotes osmoregulatory disturbances in the neotropical freshwater fish Prochilus lineatus. Aquat. Toxicol. 2009, 94, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.M.; Kim, J.W.; Yoo, D.Y.; Kim, W.; Jung, H.Y.; Choi, J.H.; Yoon, Y.S. Effects of aluminum on the reduction of neural stem cells, proliferating cells, and differentiating neuroblasts in the dentate gyrus of D-galactose-treated mice via increasing oxidative stress. J. Vet. Sci. 2016, 17, 127–136. [Google Scholar] [CrossRef]
- Wei, X.; Wei, H.; Yang, D.; Li, D.; Yang, X.; He, M.; Wu, B. Effect of aluminum exposure on glucose metabolism and its mechanism in rats. Biol. Trace Elem. Res. 2018, 186, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Andrén, C.; Henrikson, L.; Olsson, M.; Nilson, G. Effects of pH and aluminium on embryonic and early larval stages of Swedish brown frogs Rana arvalis, R. temporaria and R. dalmatina. Ecography 1988, 11, 127–135. [Google Scholar] [CrossRef]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, G.N.; Brändli, A.W. Simple vertebrate models for chemical genetics and drug discovery screens: Lessons from zebrafish and Xenopus. Dev. Dyn. 2009, 238, 1287–1308. [Google Scholar] [CrossRef]
- Slaby, S.; Marin, M.; Marchand, G.; Lemiere, S. Exposures to chemical contaminants: What can we learn from reproduction and development endpoints in the amphibian toxicology literature? Environ. Pollut. 2019, 248, 478–495. [Google Scholar] [CrossRef]
- Harland, R.M.; Grainger, R.M. Xenopus research: Metamorphosed by genetics and genomics. Trends Genet. 2011, 27, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Gurdon, J.B.; Hopwood, N. The introduction of Xenopus laevis into developmental biology: Of empire, pregnancy testing and ribosomal genes. Int. J. Dev. Biol. 2003, 44, 43–50. [Google Scholar]
- Borodinsky, L.N. Xenopus laevis as a model organism for the study of spinal cord formation, development, function and regeneration. Front. Neural Circuits 2017, 11, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, Y.; Murakami, S.; Ashikawa, Y.; Sasagawa, S.; Umemoto, N.; Shimada, Y.; Tanaka, T. Zebrafish as a systems toxicology model for developmental neurotoxicity testing. Congenit. Anom. 2015, 55, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, B.; Ebbert, P.T.; Lowery, L.A. Using Xenopus laevis retinal and spinal neurons to study mechanisms of axon guidance in vivo and in vitro. Semin. Cell Dev. Biol. 2016, 51, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Pratt, K.G.; Khakhalin, A.S. Modeling human neurodevelopmental disorders in the Xenopus tadpole: From mechanisms to therapeutic targets. Dis. Models Mech. 2013, 6, 1057–1065. [Google Scholar] [CrossRef] [Green Version]
- Guo, S. Using zebrafish to assess the impact of drugs on neural development and function. Expert Opin. Drug Discov. 2009, 4, 715–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Amora, M.; Giordani, S. The utility of zebrafish as a model for screening developmental neurotoxicity. Front. Neurosci. 2018, 12, 976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Molecular targets and alterations produced by aluminium in freshwater aquatic vertebrates. M: molecular effects. C: cellular effects. H: histological effects. P: physiological effects. B: behavioural effects. Cg: cognitive effects.
Figure 1.
Molecular targets and alterations produced by aluminium in freshwater aquatic vertebrates. M: molecular effects. C: cellular effects. H: histological effects. P: physiological effects. B: behavioural effects. Cg: cognitive effects.


{kind=link}
{kind=link}
Table 1.
Effects of aluminium on the nervous system of freshwater aquatic vertebrates reported in the literature.
Table 1.
Effects of aluminium on the nervous system of freshwater aquatic vertebrates reported in the literature.
| Al Form | Species | Exposure Conditions | Effects | Ref. |
|---|---|---|---|---|
| AlCl3 | Danio rerio | In vivo, embryos (6 hpf) to larvae (78 hpf) Conc.: 50, 100, 200 mM (sublethal conc.) Duration: 72 h |
| [60] |
| AlCl3 | Danio rerio | In vitro, embryos (4 hpf) to larvae (48 hpf) Conc.: 100 µM 44 h |
| [37] |
| AlCl3 | Danio rerio | In vivo, adults Conc.: 150 ppm Duration: 7, 14, 21 d | In brain,
| [61] |
| AlCl3 | Danio rerio | In vivo, adults (6–8 months) Conc.: 50 µg·L−1 pH: 5.8, 6.8 Duration: 24 h (acute exposure), 96 h (chronic exposure) |
| [50] |
| AlCl3 | Danio rerio | In vitro, brain homogenate of adults (6–8 months) Conc.: 50, 100, 250 µM Duration: 10 min |
| [50] |
| AlCl3 | Danio rerio | In vivo, adults (6–8 months) Conc.: 50 µg·L1 pH: 5.8 Duration: 96 h |
| [50] |
| AlCl3 | Danio rerio | In vivo, adults Conc.: 5.69, 17.08 ppm of Al (sublethal conc.) Duration: 7, 14, 21, 28 d |
| [54] |
| AlCl3 | Carassius carassius Lepomis macrochirus Neogobius fluviatilis Rutilus rutilus | In vivo, adults (3–5 years old) Conc.: 10 mg·L−1 Duration: 45 d All 4 species were used for the experiments, except for the assessment of S100ß protein content and S100ß polypeptide fragments content in the brain (L. macrochirus and C. carassius only) |
| [62] |
| AlCl3 | Salmo salar | In vivo, pre-smolt Conc.: 0.37 ± 0.04 μmol.L−1 Al pH: 5.7 Duration: 2 weeks |
| [63] |
| AlCl3 | Channa punctatus | In vitro, brain homogenate of young, middle-aged, and old individuals Conc.: 666 µM Duration: 10 min |
| [61] |
| AlCl3 | Rana arvalis Rana temporaria Rana dalmatina | In vivo, embryonic and young larvae Conc.: 100, 200, 400, 800 μg·L−1 of Al pH: 4, 5, 6 (± 0.1) Usual pH and conc. values of acidified areas in southern Sweden Duration: until a week after hatching |
| [64] |
| Al2(SO4)3 | Oreochromis niloticus | In vivo, juveniles Conc.: 1, 3 μg·mL−1 (water treatment conc.) Duration: 14 d |
| [65] |
| Al2(SO4)3 | Oreochromis niloticus | In vitro, juveniles Conc.: 1, 3 μg·mL−1 (water treatment conc.) Duration: 1 h |
| [65] |
| Al2(SO4)3 | Ctenopharyngodon idella | In vivo, adults Conc.: 0.1 mg·L−1 of Al (maximum conc. in water to protect aquatic life; not lethal for C. idella) Duration: 12, 24, 48, 72, 96 h | In brain,
| [66] |
| Al2(SO4)3 | Cirrhinus mrigala | In vivo, adults Conc.: 5.2 (chronic exposure), 17.3 ppm (acute exposure) Duration: 15, 30, 60, 90 d (chronic exposure), 14 d (acute exposure) | In brain,
| [58] |
| Al2O3NPs | Oreochromis mossambicus | In vivo, adults (6 ± 1.5 g, 6.5 ± 1 cm) Conc.: 4 mg·L−1 (sublethal conc.) Duration: 24, 72, 96 h, and 15, 30, 60 d | In brain,
| [67] |
| Al2O3NPs | Oreochromis mossambicus | In vivo, adults (6 ± 1.5 g, 6.5 ± 1 cm) Conc.: 4 mg·L−1 (1/10th of LC50-96 h) Duration: 96 h, 60 d | In brain,
| [68] |
| Al(OH)3 | Rutilus rutilus Carassius carassius | In vivo, adults Topical application of AlOH3 gel in the midline on the surface of the posterior telencephalon in living fish Conc.: NA Duration: 6 d | In R. rutilus,
| [69] |
| Al3+ (form not specified) | Lepomis gibbosus | In vivo, adults Conc.: 10 mg·L−1 Duration: NA |
| [70] |
| Al3+ (form not specified) | Oncorhynchus mykiss | In vivo, adults (≈800 g) from an aquaculture farm in South West Scotland Conc.: 8–9 mol·L−1 (mean conc. occurring in farm water) Duration: 2 years |
| [55] |
↘ for decrease, ↗ for increase, d for day, hpf for hours after fertilization.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Closset, M.; Cailliau, K.; Slaby, S.; Marin, M. Effects of Aluminium Contamination on the Nervous System of Freshwater Aquatic Vertebrates: A Review. Int. J. Mol. Sci. 2022, 23, 31. https://doi.org/10.3390/ijms23010031
AMA Style
Closset M, Cailliau K, Slaby S, Marin M. Effects of Aluminium Contamination on the Nervous System of Freshwater Aquatic Vertebrates: A Review. International Journal of Molecular Sciences. 2022; 23(1):31. https://doi.org/10.3390/ijms23010031
Chicago/Turabian StyleClosset, Marie, Katia Cailliau, Sylvain Slaby, and Matthieu Marin. 2022. "Effects of Aluminium Contamination on the Nervous System of Freshwater Aquatic Vertebrates: A Review" International Journal of Molecular Sciences 23, no. 1: 31. https://doi.org/10.3390/ijms23010031
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.