
Reassessment of SST4 Somatostatin Receptor Expression Using SST4-eGFP Knockin Mice and the Novel Rabbit Monoclonal Anti-Human SST4 Antibody 7H49L61

 ,
,
Abstract
:1. Introduction
2. Results
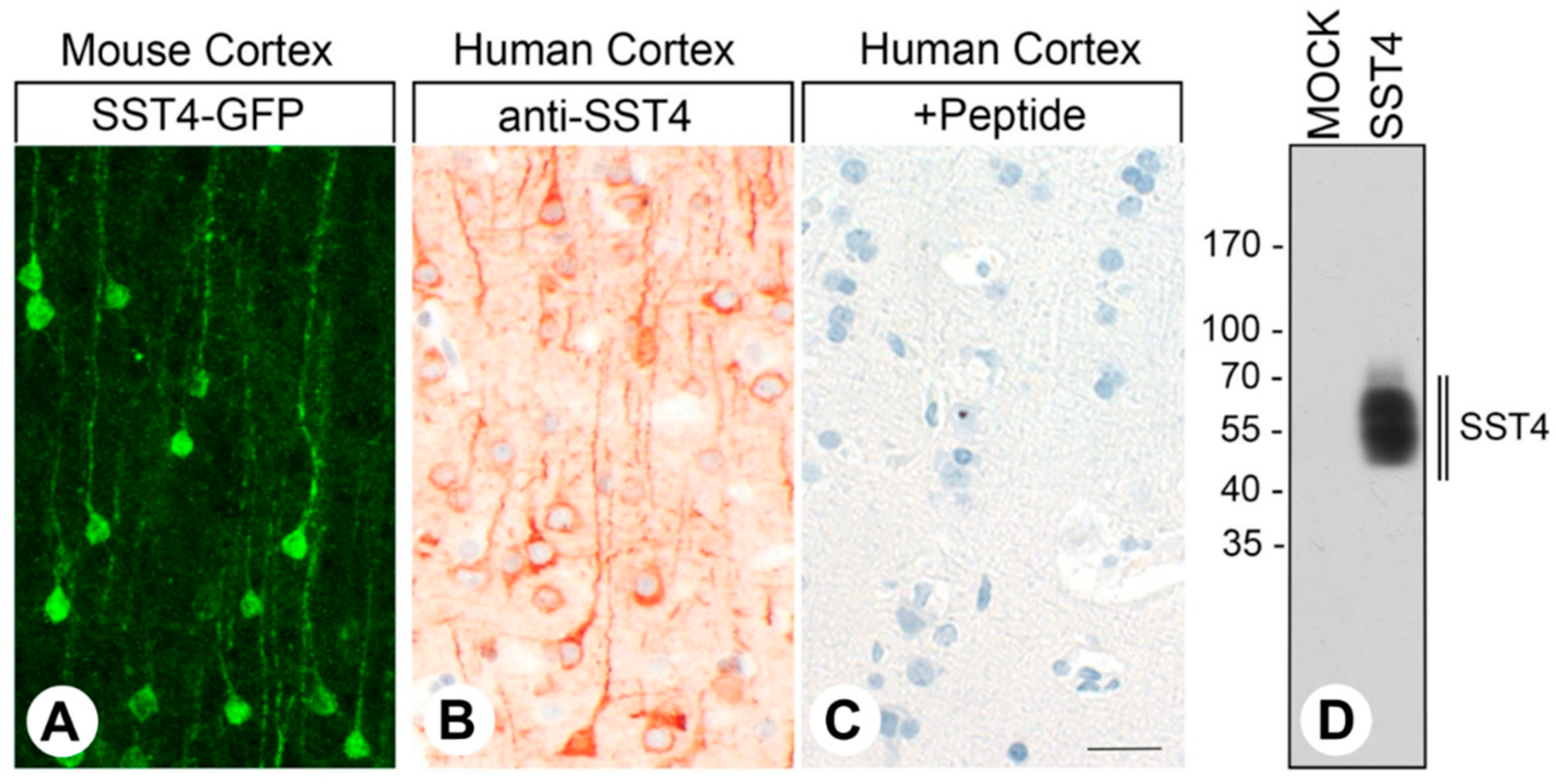
2.1. SST4-eGFP Expression in the Knockin Mouse Line
2.2. Specificity of the Monoclonal Rabbit Anti-Human SST4 Antibody 7H49L61
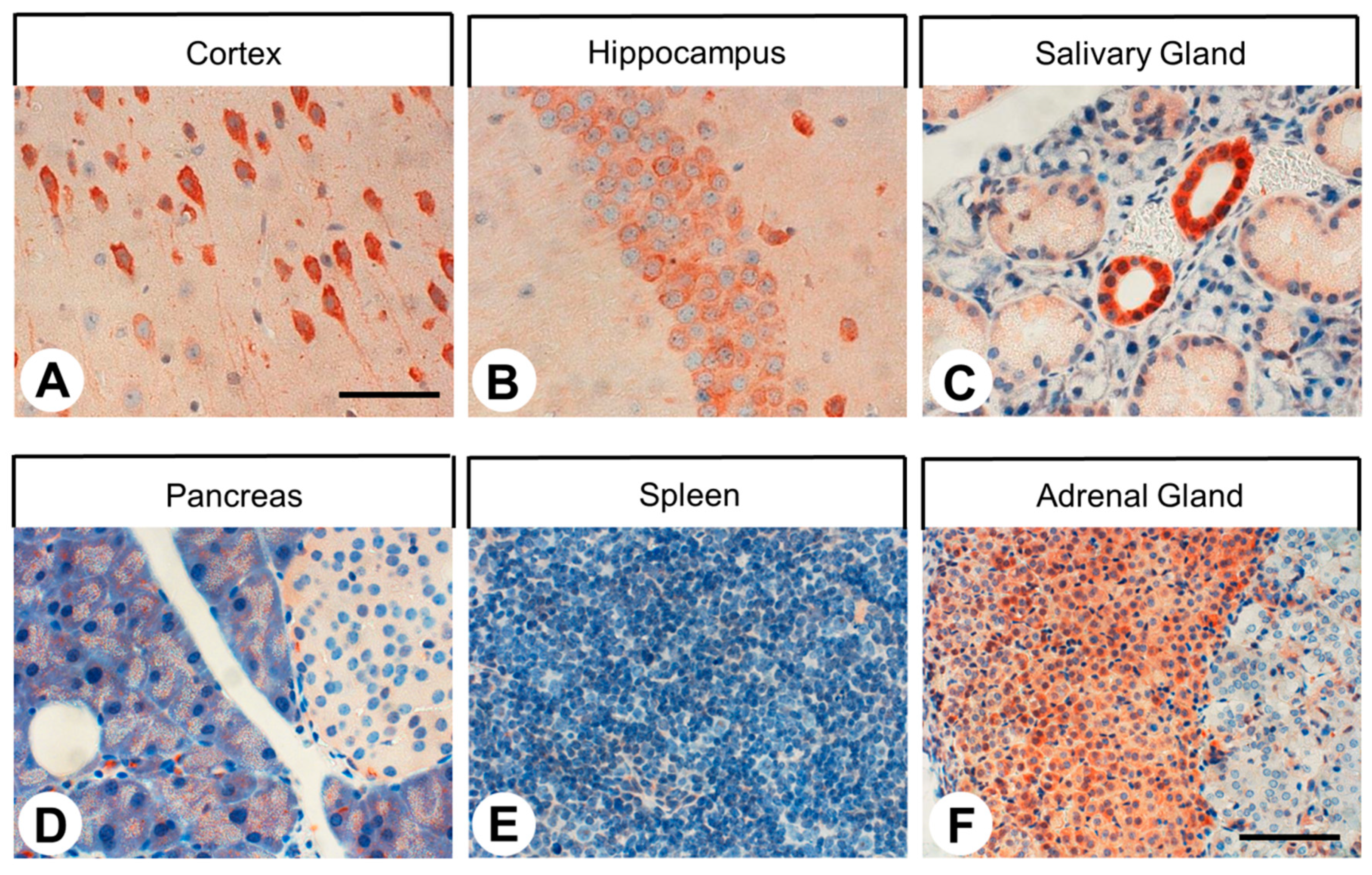
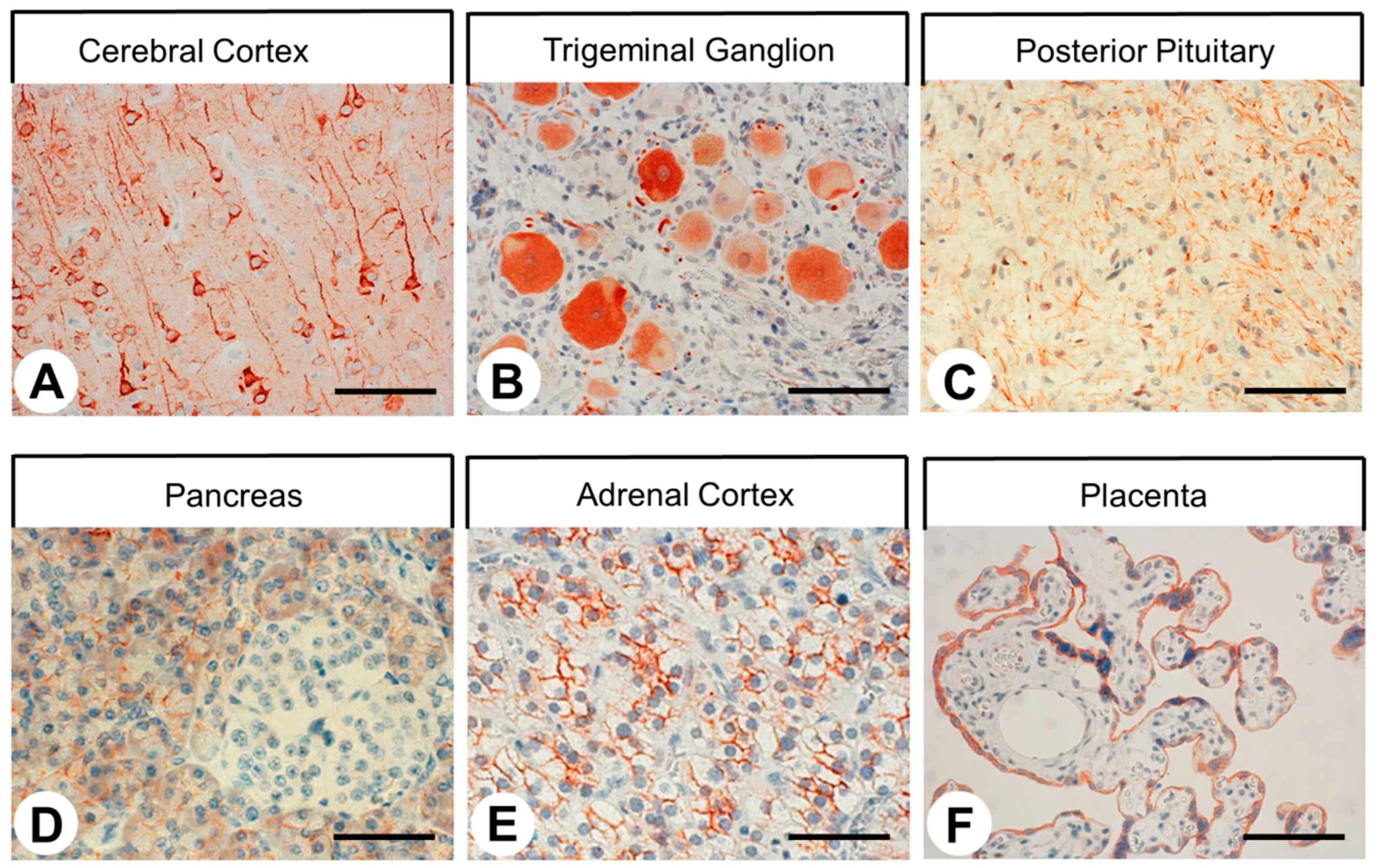
2.3. SST4 Expression in Normal Human Tissues
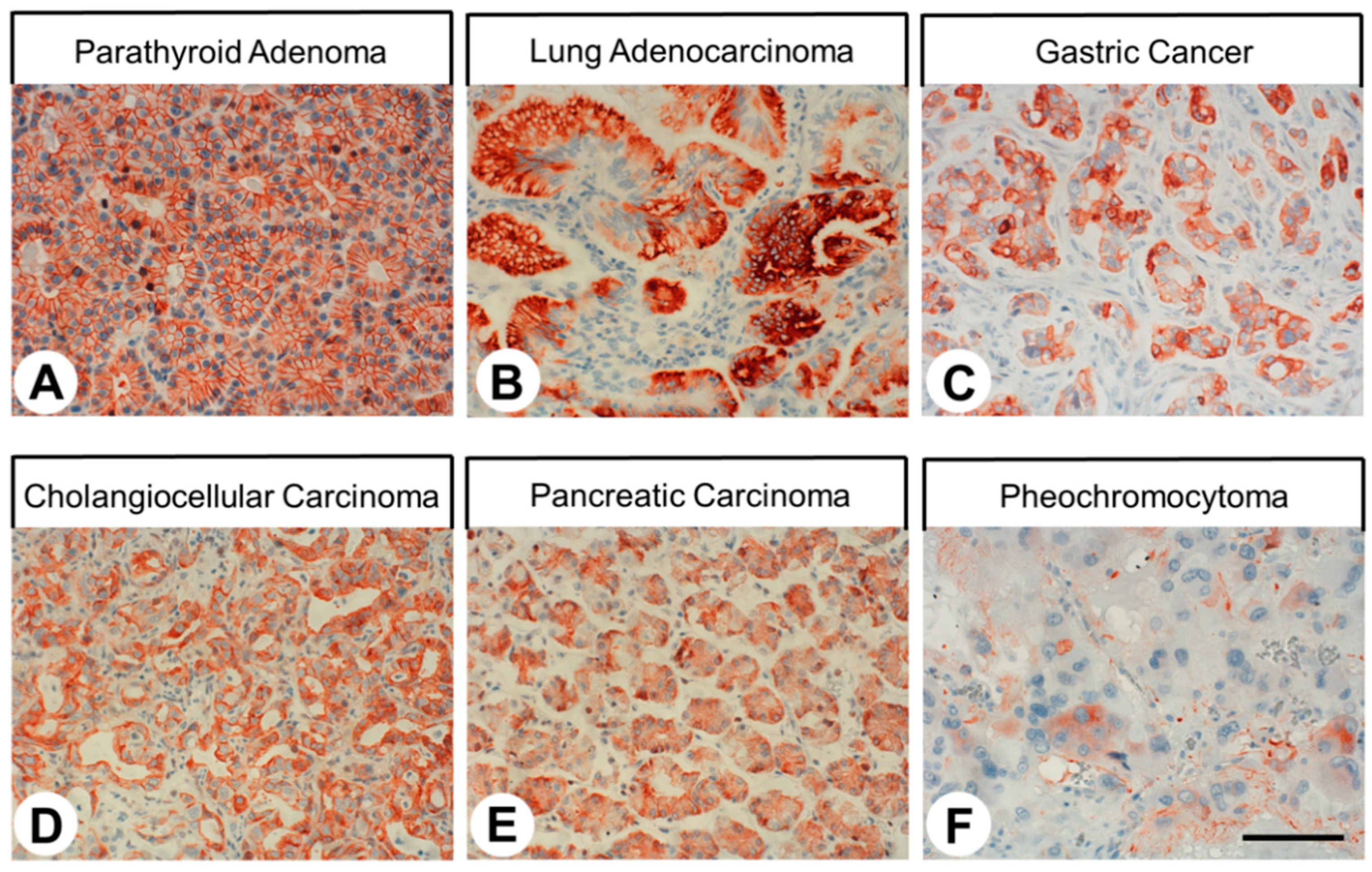
2.4. SST4 Expression in Human Tumors
3. Discussion
3.1. SST4-eGFP Expression in the Knockin Mouse Line
3.2. Specificity of the Monoclonal Rabbit Anti-human SST4 Antibody 7H49L61
3.3. SST4 Expression in Normal Human Tissues
3.4. SST4 Expression in Human Tumors
4. Materials and Methods
4.1. Monoclonal Antibody
4.2. Immunoblot Analysis
4.3. SST4-eGFP Knockin Mice
4.4. Immunohistochemistry on SST4-eGFP Knockin Mouse Tissues
4.5. In Situ Hybridization Experiments
4.6. Human Tissue Samples
4.7. Immunohistochemistry on Human Tissue Samples
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| eGFP | enhanced green fluorescent protein |
| IRS | immunoreactivity score |
| SST | somatostatin receptor |
References
- Günther, T.; Tulipano, G.; Dournaud, P.; Bousquet, C.; Csaba, Z.; Kreienkamp, H.J.; Lupp, A.; Korbonits, M.; Castano, J.P.; Wester, H.J.; et al. International Union of Basic and Clinical Pharmacology. CV. Somatostatin receptors: Structure, function, ligands, and new nomenclature. Pharmacol. Rev. 2018, 70, 763–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeyda, T.; Hochgeschwender, U. Null mutant mouse models of somatostatin and cortistatin, and their receptors. Mol. Cell. Endocrinol. 2008, 286, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Theodoropoulou, M.; Stalla, G.K. Somatostatin receptors: From signaling to clinical practice. Front. Neuroendocrinol. 2013, 34, 228–252. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Zeyda, T.; Johnson, B.; Hochgeschwender, U.; de Lecea, L.; Tallent, M.K. Somatostatin receptor subtype 4 couples to the M-current to regulate seizures. J. Neurosci. 2008, 28, 3567–3576. [Google Scholar] [CrossRef]
- Farrell, S.R.; Raymond, I.D.; Foote, M.; Brecha, N.C.; Barnes, S. Modulation of voltage-gated ion channels in rat retinal ganglion cells mediated by somatostatin receptor subtype 4. J. Neurophysiol. 2010, 104, 1347–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrell, S.R.; Rankin, D.R.; Brecha, N.C.; Barnes, S. Somatostatin receptor subtype 4 modulates L-type calcium channels via Gβγ and PKC signaling in rat retinal ganglion cells. Channels 2014, 8, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Gorham, L.; Just, S.; Doods, H. Somatostatin 4 receptor activation modulates G-protein coupled inward rectifying potassium channels and voltage stimulated calcium signals in dorsal root ganglion neurons. Eur. J. Pharmacol. 2014, 736, 101–106. [Google Scholar] [CrossRef]
- Gorham, L.; Just, S.; Doods, H. Somatostatin 4 receptor activation modulates TRPV1 currents in dorsal root ganglion neurons. Neurosci. Lett. 2014, 573, 35–39. [Google Scholar] [CrossRef]
- Schuelert, N.; Just, S.; Kuelzer, R.; Corradini, L.; Gorham, L.C.J.; Doods, H. The somatostatin receptor 4 agonist J-2156 reduces mechanosensitivity of peripheral nerve afferents and spinal neurons in an inflammatory pain model. Eur. J. Pharmacol. 2015, 746, 274–281. [Google Scholar] [CrossRef]
- Dasgupta, P.; Günther, T.; Schulz, S. Pharmacological characterization of veldoreotide as a somatostatin receptor 4 agonist. Life 2021, 11, 1075. [Google Scholar] [CrossRef]
- Yamamoto, J.; Ohnuma, K.; Hatano, R.; Okamoto, T.; Komiya, E.; Yamazaki, H.; Iwata, S.; Dang, N.H.; Aoe, K.; Kishimoto, T.; et al. Regulation of somatostatin receptor 4-mediated cytostatic effects by CD26 in malignant pleural mesothelioma. Br. J. Cancer 2014, 110, 2232–2245. [Google Scholar] [CrossRef] [Green Version]
- Alderton, F.; Humphrey, P.P.A.; Sellers, L.A. High-intensity p38 kinase activity is critical for p21(cip1) induction and the antiproliferative function of G(i) protein-coupled receptors. Mol. Pharmacol. 2001, 59, 1119–1128. [Google Scholar] [CrossRef]
- Smalley, K.S.M.; Koenig, J.A.; Feniuk, W.; Humphrey, P.P.A. Ligand internalization and recycling by human recombinant somatostatin type 4 (h sst(4)) receptors expressed in CHO-K1 cells. Br. J. Pharmacol. 2001, 132, 1102–1110. [Google Scholar] [CrossRef] [Green Version]
- Kreienkamp, H.-J.; Roth, A.; Richter, D. Rat somatostatin receptor subtype 4 can be made sensitive to agonist-induced internalization by mutation of a single threonine (residue 331). DNA Cell Biol. 1998, 17, 869–878. [Google Scholar] [CrossRef]
- Schreff, M.; Schulz, S.; Händel, M.; Keilhoff, G.; Braun, H.; Pereira, G.; Klutzny, M.; Schmidt, H.; Wolf, G.; Höllt, V. Distribution, targeting, and internalization of the sst4 somatostatin receptor in rat brain. J. Neurosci. 2000, 20, 3785–3797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheich, B.; Gaszner, B.; Kormos, V.; László, K.; Ádori, C.; Borbély, É.; Hajna, Z.; Tékus, V.; Bölcskei, K.; Ábrahám, I.; et al. Somatostatin receptor subtype 4 activation is involved in anxiety and depression-like behavior in mouse models. Neuropharmacology 2016, 101, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Scheich, B.; Csekö, K.; Borbély, É.; Ábrahám, I.; Csernus, V.; Gaszner, B.; Helyes, Z. Higher susceptibility of somatostatin 4 receptor gene-deleted mice to chronic stress-induced behavioral and neuroendocrine alterations. Neuroscience 2017, 346, 320–336. [Google Scholar] [CrossRef]
- Szentes, N.; Tékus, V.; Mohos, V.; Borbély, E.; Helyes, Z. Exploratory and locomotor activity, learning and memory functions in somatostatin receptor subtype 4 gene-deficient mice in relation to aging and sex. GeroScience 2019, 41, 631–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helyes, Z.; Pinter, E.; Sandor, K.; Elekes, K.; Banvolgyi, A.; Keszthelyi, D.; Szoke, E.; Toth, D.M.; Sandor, Z.; Kereskai, L.; et al. Impaired defense mechanism against inflammation, hyperalgesia, and airway hyperreactivity in somatostatin 4 receptor gene-deleted mice. Proc. Natl. Acad. Sci. USA 2009, 106, 13088–13093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sándor, K.; Elekes, K.; Szabó, A.; Pintér, E.; Engström, M.; Wurster, S.; Szolcsányi, J.; Helyes, Z. Analgesic effects of the somatostatin sst4 receptor selective agonist J-2156 in acute and chronic pain models. Eur. J. Pharmacol. 2006, 539, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Elekes, K.; Helyes, Z.; Kereskai, L.; Sándor, K.; Pintér, E.; Pozsgai, G.; Tékus, V.; Bánvölgyi, A.; Németh, J.; Szüts, T.; et al. Inhibitory effects of synthetic somatostatin receptor subtype 4 agonists on acute and chronic airway inflammation and hyperreactivity in the mouse. Eur. J. Pharmacol. 2008, 578, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, P.A.; Kuo, A.; Khan, N.; Gorham, L.; Nicholson, J.R.; Corradini, L.; Vetter, I.; Smith, M.T. The somatostatin receptor-4 agonist J-2156 alleviates mechanical hypersensitivity in a rat model of breast cancer induced bone pain. Front. Pharmacol. 2018, 9, 495. [Google Scholar] [CrossRef] [Green Version]
- Kecskes, A.; Pohoczky, K.; Kecskes, M.; Varga, Z.V.; Kormos, V.; Szoke, E.; Henn-Mike, N.; Feher, M.; Kun, J.; Gyenesei, A.; et al. Characterization of neurons expressing the novel analgesic drug target somatostatin receptor 4 in mouse and human brains. Int. J. Mol. Sci. 2020, 21, 7788. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, K.E.; Farr, S.A.; Banks, W.A.; Crider, A.M.; Morley, J.E.; Witt, K.A. Somatostatin receptor subtype-4 agonist NNC 26–9100 decreases extracellular and intracellular Ab1242 trimers. Eur. J. Pharmacol. 2012, 683, 116–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval, K.E.; Farr, S.A.; Banks, W.A.; Crider, A.M.; Morley, J.E.; Witt, K.A. Somatostatin receptor subtype-4 agonist NNC 26-9100 mitigates the effect of soluble Ab(42) oligomers via a metalloproteinase-dependent mechanism. Brain Res. 2013, 1520, 145–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehlmann, D.; Langenegger, D.; Schuepbach, E.; Siehler, S.; Feuerbach, D.; Hoyer, D. Distribution and characterisation of somatostatin receptor mRNA and binding sites in the brain and periphery. J. Physiol. Paris 2000, 94, 265–281. [Google Scholar] [CrossRef]
- Schulz, S.; Händel, M.; Schreff, M.; Schmidt, H.; Höllt, V. Localization of five somatostatin receptors in the rat central nervous system using subtype-specific antibodies. J. Physiol. Paris 2000, 94, 259–264. [Google Scholar] [CrossRef]
- Stumm, R.K.; Zhou, C.; Schulz, S.; Endres, M.; Kronenberg, G.; Allen, J.P.; Tulipano, G.; Höllt, V. Somatostatin receptor 2 is activated in cortical neurons and contributes to neurodegeneration after focal ischemia. J. Neurosci. 2004, 24, 11404–11415. [Google Scholar] [CrossRef] [PubMed]
- Bär, K.J.; Schurigt, U.; Scholze, A.; Segond Von Banchet, G.; Stopfel, N.; Bräuer, R.; Halbhuber, K.J.; Schaible, H.G. The expression and localization of somatostatin receptors in dorsal root ganglion neurons of normal and monoarthritic rats. Neuroscience 2004, 127, 197–206. [Google Scholar] [CrossRef]
- Ludvigsen, E.; Carlsson, C.; Tiensuu Janson, E.; Sandler, S.; Stridsberg, M. Somatostatin receptor 1-5; expression profiles during rat development. Ups. J. Med. Sci. 2015, 120, 157–168. [Google Scholar] [CrossRef]
- Selmer, I.-S.; Schindler, M.; Humphrey, P.P.A.; Waldvogel, H.J.; Faull, R.L.M.; Emson, P.C. First localisation of somatostatin sst(4) receptor protein in selected human brain areas: An immunohistochemical study. Brain Res. Mol. Brain Res. 2000, 82, 114–125. [Google Scholar] [CrossRef]
- Taniyama, Y.; Suzuki, T.; Mikami, Y.; Moriya, T.; Satomi, S.; Sasano, H. Systemic distribution of somatostatin receptor subtypes in human: An immunohistochemical study. Endocr. J. 2005, 52, 605–611. [Google Scholar] [CrossRef] [Green Version]
- Remes, S.M.; Leijon, H.L.; Vesterinen, T.J.; Arola, J.T.; Haglund, C.H. Immunohistochemical expression of somatostatin receptor subtypes in a panel of neuroendocrine neoplasias. J. Histochem. Cytochem. 2019, 67, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Nemes, B.; Bölcskei, K.; Kecskés, A.; Kormos, V.; Gaszner, B.; Aczél, T.; Hegedüs, D.; Pintér, E.; Helyes, Z.; Sándor, Z. Human somatostatin SST4 receptor transgenic mice: Construction and brain expression pattern characterization. Int. J. Mol. Sci. 2021, 22, 3758. [Google Scholar] [CrossRef] [PubMed]
- Lichtenauer-Kaligis, E.G.; van Hagen, P.M.; Lamberts, S.W.; Hofland, L.J. Somatostatin receptor subtypes in human immune cells. Eur. J. Endocrinol. 2000, 143 (Suppl. 1), S21–S25. [Google Scholar] [CrossRef] [Green Version]
- Papotti, M.; Kumar, U.; Volante, M.; Pecchioni, C.; Patel, Y.C. Immunohistochemical detection of somatostatin receptor types 1–5 in medullary carcinoma of the thyroid. Clin. Endocrinol. 2001, 54, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Hansson, J.; Bjartell, A.; Gadaleanu, V.; Dizeyi, N.; Abrahamsson, P.A. Expression of somatostatin receptor subtypes 2 and 4 in human benign prostatic hyperplasia and prostatic cancer. Prostate 2002, 53, 50–59. [Google Scholar] [CrossRef]
- Schulz, S.; Schmitt, J.; Quednow, C.; Roessner, A.; Weise, W. Immunohistochemical detection of somatostatin receptors in human ovarian tumors. Gynecol. Oncol. 2002, 84, 235–240. [Google Scholar] [CrossRef]
- Mundschenk, J.; Unger, N.; Schulz, S.; Höllt, V.; Schulz, S.; Steinke, R.; Lehnert, H. Somatostatin receptor subtypes in human pheochromocytoma: Subcellular expression pattern and functional relevance for octreotide scintigraphy. J. Clin. Endocrinol. Metab. 2003, 88, 5150–5157. [Google Scholar] [CrossRef] [Green Version]
- Schulz, S.; Schmitt, J.; Weise, W. Frequent expression of immunoreactive somatostatin receptors in cervical and endometrial cancer. Gynecol. Oncol. 2003, 89, 385–390. [Google Scholar] [CrossRef]
- Klagge, A.; Krause, K.; Schierle, K.; Steinert, F.; Dralle, H.; Fuhrer, D. Somatostatin receptor subtype expression in human thyroid tumours. Horm. Metab. Res. 2010, 42, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Müssig, K.; Wehrmann, T.; Dittmann, H.; Wehrmann, M.; Ueberberg, B.; Schulz, S.; Bares, R.; Petersenn, S. Expression of the proliferation marker Ki-67 associates with tumour staging and clinical outcome in differentiated thyroid carcinomas. Clin. Endocrinol. 2012, 77, 139–145. [Google Scholar] [CrossRef]
- Pazaitou-Panayiotou, K.; Tiensuu Janson, E.; Koletsa, T.; Kotoula, V.; Stridsberg, M.; Karkavelas, G.; Karayannopoulou, G. Somatostatin receptor expression in non-medullary thyroid carcinomas. Hormones 2012, 11, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Tsuta, K.; Wistuba, I.I.; Moran, C.A. Differential expression of somatostatin receptors 1-5 in neuroendocrine carcinoma of the lung. Pathol. Res. Pract. 2012, 208, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Unger, N.; Ueberberg, B.; Schulz, S.; Saeger, W.; Mann, K.; Petersenn, S. Differential expression of somatostatin receptor subtype 1-5 proteins in numerous human normal tissues. Exp. Clin. Endocrinol. Diabetes 2012, 120, 482–489. [Google Scholar] [CrossRef]
- Arne, G.; Nilsson, B.; Dalmo, J.; Kristiansson, E.; Arvidsson, Y.; Forssell-Aronsson, E.; Nilsson, O.; Ahlman, H. Gastrointestinal stromal tumors (GISTs) express somatostatin receptors and bind radiolabeled somatostatin analogs. Acta Oncol. 2013, 52, 783–792. [Google Scholar] [CrossRef]
- Frati, A.; Rouzier, R.; Lesieur, B.; Werkoff, G.; Antoine, M.; Rodenas, A.; Darai, E.; Chereau, E. Expression of somatostatin type-2 and -4 receptor and correlation with histological type in breast cancer. Anticancer Res. 2014, 34, 3997–4003. [Google Scholar]
- Kaemmerer, D.; Specht, E.; Sänger, J.; Wirtz, R.M.; Sayeg, M.; Schulz, S.; Lupp, A. Somatostatin receptors in bronchopulmonary neuroendocrine neoplasms: New diagnostic, prognostic, and therapeutic markers. J. Clin. Endocrinol. Metab. 2015, 100, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Stollberg, S.; Kaemmerer, D.; Neubauer, E.; Schulz, S.; Simonitsch-Klupp, I.; Kiesewetter, B.; Raderer, M.; Lupp, A. Differential somatostatin and CXCR4 chemokine receptor expression in MALT-type lymphoma of gastric and extragastric origin. J. Cancer Res. Clin. Oncol. 2016, 142, 2239–2247. [Google Scholar] [CrossRef]
- Kaemmerer, D.; Schindler, R.; Mussbach, F.; Dahmen, U.; Altendorf-Hofmann, A.; Dirsch, O.; Sänger, J.; Schulz, S.; Lupp, A. Somatostatin and CXCR4 chemokine receptor expression in hepatocellular and cholangiocellular carcinomas: Tumor capillaries as promising targets. BMC Cancer 2017, 17, 896. [Google Scholar] [CrossRef] [Green Version]
- Stumpf, C.; Kaemmerer, D.; Neubauer, E.; Sänger, J.; Schulz, S.; Lupp, A. Somatostatin and CXCR4 expression patterns in adenocarcinoma and squamous cell carcinoma of the lung relative to small cell lung cancer. J. Cancer Res. Clin. Oncol. 2018, 144, 1921–1932. [Google Scholar] [CrossRef] [PubMed]
- Kajtazi, Y.; Kaemmerer, D.; Sänger, J.; Schulz, S.; Lupp, A. Somatostatin and chemokine CXCR4 receptor expression in pancreatic adenocarcinoma relative to pancreatic neuroendocrine tumours. J. Cancer Res. Clin. Oncol. 2019, 145, 2481–2493. [Google Scholar] [CrossRef] [PubMed]
- Storvall, S.; Leijon, H.; Ryhanen, E.; Louhimo, J.; Haglund, C.; Schalin-Jantti, C.; Arola, J. Somatostatin receptor expression in parathyroid neoplasms. Endocr. Connect. 2019, 8, 1213–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Tan, H.; Zhao, Y.; Zhou, Y.; Cao, L. Expression and selective activation of somatostatin receptor subtypes induces cell cycle arrest in cancer cells. Oncol. Lett. 2019, 17, 1723–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maas, M.; Mayer, L.; Hennenlotter, J.; Stuhler, V.; Walz, S.; Scharpf, M.; Kuhs, U.; Neumann, T.; Stenzl, A.; Todenhofer, T. Prognostic impact of somatostatin receptor expression in advanced bladder cancer. Urol. Oncol. 2020, 38, 935. [Google Scholar] [CrossRef]
- Panetta, R.; Patel, Y.C. Expression of mRNA for all five human somatostatin receptors (hSSTR1-5) in pituitary tumors. Life Sci. 1995, 56, 333–342. [Google Scholar] [CrossRef]
- Makhlouf, G.M.; Schubert, M.L. Gastric somatostatin: A paracrine regulator of acid secretion. Metabolism 1990, 39, 138–142. [Google Scholar] [CrossRef]
- Odes, H.S.; Muallem, R.; Reimer, R.; Ioffe, S.; Beil, W.; Schwenk, M.; Sewing, K.F. Effect of somatostatin-14 on duodenal mucosal bicarbonate secretion in guinea pigs. Dig. Dis. Sci. 1995, 40, 678–684. [Google Scholar] [CrossRef]
- Magnusson, I.; Einarsson, K.; Angelin, B.; Nyberg, B.; Bergström, K.; Thulin, L. Effects of somatostatin on hepatic bile formation. Gastroenterology 1989, 96, 206–212. [Google Scholar] [CrossRef]
- Marteau, P.; Chrétien, Y.; Calmus, Y.; Parc, R.; Poupon, R. Pharmacological effect of somatostatin on bile secretion in man. Digestion 1989, 42, 16–21. [Google Scholar] [CrossRef]
- Gong, A.-Y.; Tietz, P.S.; Muff, M.A.; Splinter, P.L.; Huebert, R.C.; Strowski, M.Z.; Chen, X.-M.; LaRusso, N.F. Somatostatin stimulates ductal bile absorption and inhibits ductal bile secretion in mice via SSTR2 on cholangiocytes. Am. J. Physiol. Cell Physiol. 2003, 284, C1205–C1214. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, S.; Watson, N.; Long, E.; Sharpe, S.; Zhong, W.; Xu, S.Z.; Atkin, S.L. Expression of somatostatin and somatostatin receptor subtypes 1–5 in human normal and diseased kidney. J. Histochem. Cytochem. 2008, 56, 733–743. [Google Scholar] [CrossRef] [Green Version]
- Ray, C.; Carney, S.; Morgan, T.; Gillies, A. Somatostatin as a modulator of distal nephron water permeability. Clin. Sci. 1993, 84, 455–460. [Google Scholar] [CrossRef]
- Caron, P.; Buscail, L.; Beckers, A.; Esteve, J.P.; Igout, A.; Hennen, G.; Susini, C. Expression of somatostatin receptor SST4 in human placenta and absence of octreotide effect on human placental growth hormone concentration during pregnancy. J. Clin. Endocrinol. Metab. 1997, 82, 3771–3776. [Google Scholar] [PubMed] [Green Version]
- Werner, Y.; Mass, E.; Kumar, P.A.; Ulas, T.; Händlers, K.; Horne, A.; Klee, K.; Lupp, A.; Schütz, D.; Saaber, F.; et al. Cxcr4 distinguishes HSC-derived monocytes from microglia and reveals monocyte immune responses to experimental stroke. Nat. Neurosci. 2020, 23, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Memi, F.; Abe, P.; Cariboni, A.; MacKay, F.; Parnavelas, J.G.; Stumm, R. CXC chemokine receptor 7 (CXCR7) affects the migration of GnRH neurons by regulating CXCL12 availability. J. Neurosci. 2013, 33, 17527–17537. [Google Scholar] [CrossRef] [Green Version]
- Saaber, F.; Schutz, D.; Miess, E.; Abe, P.; Desikan, S.; Ashok Kumar, P.; Balk, S.; Huang, K.; Beaulieu, J.M.; Schulz, S.; et al. ACKR3 regulation of neuronal migration requires ACKR3 phosphorylation, but not beta-arrestin. Cell Rep. 2019, 26, 1473–1488. [Google Scholar] [CrossRef] [Green Version]
- Kaemmerer, D.; Sänger, J.; Arsenic, R.; D’Haese, J.G.; Neumann, J.; Schmitt-Graeff, A.; Wirtz, R.M.; Schulz, S.; Lupp, A. Evaluation of somatostatin, CXCR4 chemokine and endothelin A receptor expression in a large set of paragangliomas. Oncotarget 2017, 8, 89958–89969. [Google Scholar] [CrossRef] [Green Version]
- Remmele, W.; Stegner, H.E. Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue. Pathologe 1987, 8, 138–140. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor Type (Total Number of Samples) | SST4 Positive Tumors (n) | Immunoreactivity Score (IRS) | ||
|---|---|---|---|---|
| Mean | Min | Max | ||
| Glioblastoma (9) | 9 | 6.78 | 4.5 | 9 |
| Thyroid carcinoma (36) | 18 | 2,95 | 0 | 9 |
| - papillary (11) | 5 | 2.05 | 0 | 5 |
| - follicular (10) | 3 | 2.30 | 0 | 7.5 |
| - medullary (7) | 6 | 5.18 | 0 | 9 |
| - anaplastic (8) | 4 | 3.06 | 0 | 8 |
| Parathyroid adenoma (10) | 10 | 9.35 | 3 | 12 |
| Lung cancer (31) | 10 | 2,19 | 0 | 9 |
| - Adenocarcinoma (12) | 9 | 4.50 | 0 | 9 |
| - Squamous cell carcinoma (10) | 0 | 0.68 | 0 | 2 |
| - Small cell lung cancer (9) | 1 | 0.78 | 0 | 5 |
| Gastric cancer (9) | 9 | 8.06 | 4.5 | 10 |
| Colon carcinoma (9) | 6 | 3.70 | 1 | 9 |
| Gastrointestinal stromal tumor (10) | 8 | 5.03 | 0 | 10 |
| Hepatocellular carcinoma (11) | 3 | 1.55 | 0 | 6 |
| Cholangiocellular carcinoma (9) | 3 | 3.70 | 0 | 12 |
| Pancreatic adenocarcinoma (11) | 10 | 6.27 | 2 | 9 |
| Renal clear cell carcinoma (8) | 0 | 0.63 | 0 | 2 |
| Pheochromocytoma (7) | 7 | 6.50 | 4 | 10.5 |
| Bladder Cancer (7) | 6 | 4.68 | 2 | 7.5 |
| Prostate adenocarcinoma (12) | 5 | 2.20 | 0 | 7.5 |
| Testicular cancer (12) | 8 | 3.60 | 0 | 7.5 |
| Breast carcinoma (9) | 6 | 3.47 | 0 | 6.25 |
| Endometrial cancer (10) | 2 | 1.53 | 0 | 4.5 |
| Cervical cancer (9) | 3 | 1.40 | 0 | 4.5 |
| Ovarian cancer (10) | 4 | 2.44 | 0 | 5 |
| Lymphoma (12) | 10 | 6.02 | 1.5 | 10 |
| Melanoma (5) | 3 | 3.20 | 1 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lupp, A.; Ehms, B.; Stumm, R.; Göckeritz, J.; Mawrin, C.; Schulz, S. Reassessment of SST4 Somatostatin Receptor Expression Using SST4-eGFP Knockin Mice and the Novel Rabbit Monoclonal Anti-Human SST4 Antibody 7H49L61. Int. J. Mol. Sci. 2021, 22, 12981. https://doi.org/10.3390/ijms222312981
Lupp A, Ehms B, Stumm R, Göckeritz J, Mawrin C, Schulz S. Reassessment of SST4 Somatostatin Receptor Expression Using SST4-eGFP Knockin Mice and the Novel Rabbit Monoclonal Anti-Human SST4 Antibody 7H49L61. International Journal of Molecular Sciences. 2021; 22(23):12981. https://doi.org/10.3390/ijms222312981
Chicago/Turabian StyleLupp, Amelie, Blanca Ehms, Ralf Stumm, Johannes Göckeritz, Christian Mawrin, and Stefan Schulz. 2021. "Reassessment of SST4 Somatostatin Receptor Expression Using SST4-eGFP Knockin Mice and the Novel Rabbit Monoclonal Anti-Human SST4 Antibody 7H49L61" International Journal of Molecular Sciences 22, no. 23: 12981. https://doi.org/10.3390/ijms222312981