
Identification and Cloning of a CC-NBS-NBS-LRR Gene as a Candidate of Pm40 by Integrated Analysis of Both the Available Transcriptional Data and Published Linkage Mapping

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Identification of Pm40 Effectiveness against Bgt Infection
2.2. Functional Annotation of the Genes within the Region Flanked by BF478514 and BF291338
2.3. Polymorphic EST Sequences in the Pm40 Region
2.4. Differential Expression Level of Genes within Pm40 Candidate Region
2.5. Screening and Cloning of TraesCS7B01G164000 and TraesCS7B01G164600
2.6. Validation of the Candidate Gene by Cosegregation with Powdery Mildew Resistance
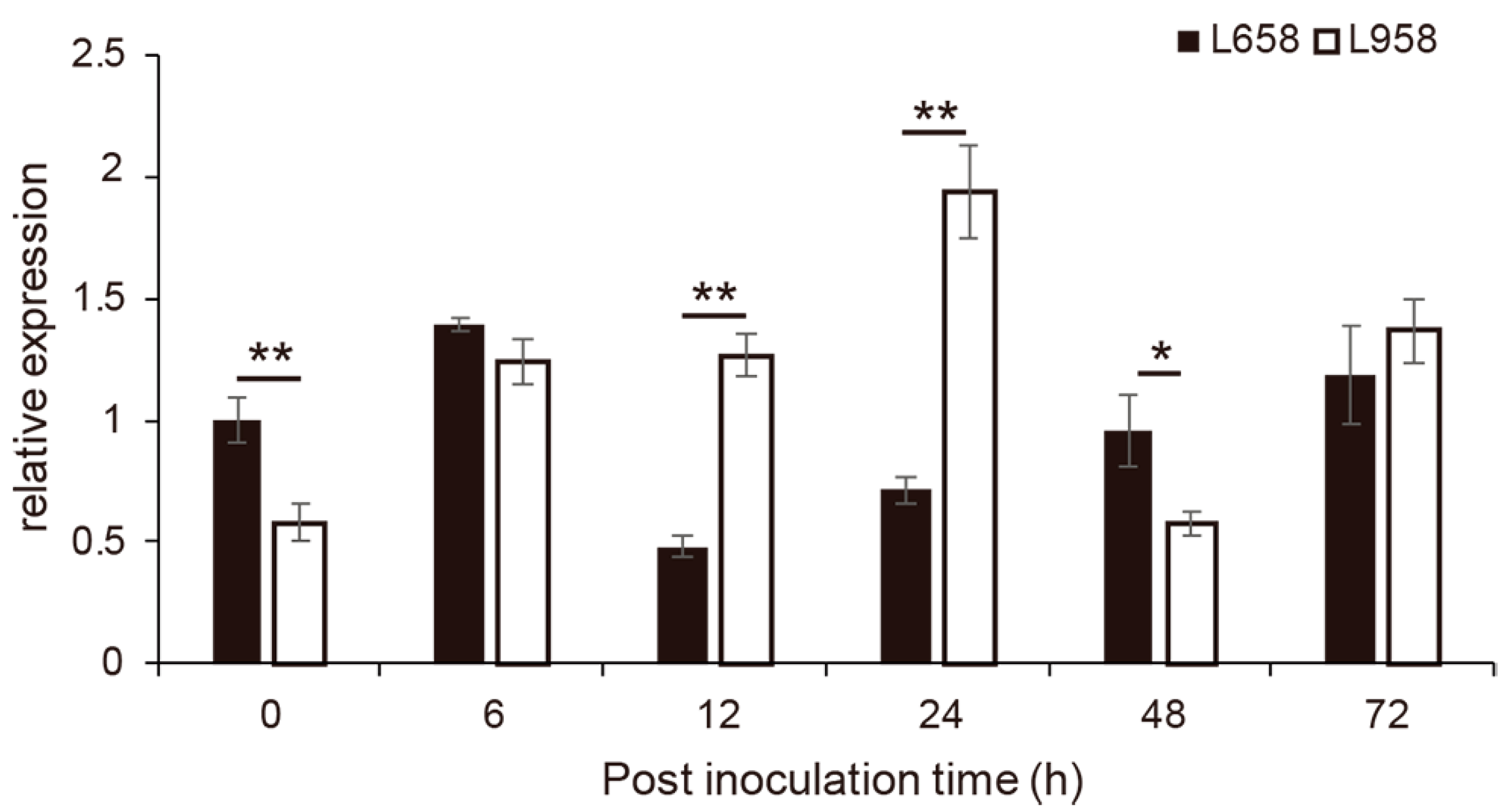
2.7. Expression Analysis of TraesCS7B01G164000 after Bgt Inoculation by qRT-PCR
3. Discussion
3.1. The Broad Spectrum and Putatively Durable Resistance of Pm40
3.2. The Possible Chromosomal Regions Containing the Candidate Pm40 Gene
3.3. TraesCS7B01G164000 as the Putative Candidate Gene of Pm40
3.4. Validation of TraesCS7B01G164000 as the Candidate Gene of Pm40
4. Materials and Methods
4.1. Plant Materials, the Resistance Evaluations, and the Cytological Observations of Cell Death
4.2. Annotation of the Genes within the Physical Region of Pm40
4.3. Acquisition of RNA Sequencing Data and Detection of Deferentially Expressed Gene
4.4. Variation Extraction, Filtering and Annotation
4.5. Gene Cloning and Sequence Analysis
4.6. SNP Genotyping by AS-PCR
4.7. Gene Expression Analysis by qRT-PCR
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Juroszek, P.; Von Tiedemann, A. Climate change and potential future risks through wheat diseases: A review. Eur. J. Plant. Pathol. 2013, 136, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.X.; Zhong, S.F.; Liu, N.; Chen, W.Q.; Liu, T.G.; Li, X.; Zhang, M.; Ren, Z.L.; Yang, J.Z.; Luo, P.G. Gene expression profile and physiological and biochemical characterization of hexaploid wheat inoculated with Blumeria graminis f. sp. tritici. Physiol. Mol. Plant. Pathol. 2015, 90, 39–48. [Google Scholar] [CrossRef]
- Liang, Y.; Xia, Y.; Chang, X.; Gong, G.; Yang, J.; Hu, Y.; Cahill, M.; Luo, L.; Li, T.; He, L. Comparative proteomic analysis of wheat carrying Pm40 response to Blumeria graminis f. sp. tritici using two-dimensional electrophoresis. Int. J. Mol. Sci. 2019, 20, 933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Xiao, J.; Xu, J.; Wan, W.; Qin, B.; Cao, A.; Chen, W.; Xing, L.; Du, C.; Gao, X. Two members of TaRLK family confer powdery mildew resistance in common wheat. BMC Plant. Biol. 2016, 16, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, P.G.; Hu, X.Y.; Chang, Z.J.; Zhang, M.; Zhang, H.Q.; Ren, Z.L. A new stripe rust resistance gene transferred from Thinopyrum intermedium to hexaploid wheat (Triticum aestivum). Phytoprotection 2009, 90, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Liang, Y.; Zhang, M.; Tan, F.; Zhong, S.; Li, X.; Gong, G.; Chang, X.; Shang, J.; Tang, S. Comparative transcriptome profiling of Blumeria graminis f. sp. tritici during compatible and incompatible interactions with sister wheat lines carrying and lacking Pm40. PLoS ONE 2018, 13, e0198891. [Google Scholar] [CrossRef]
- Tang, S.; Hu, Y.; Zhong, S.; Luo, P. The potential role of powdery mildew-resistance gene Pm40 in Chinese wheat-breeding programs in the post-Pm21 Era. Engineering 2018, 4, 500–506. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, W.J.; Morris, C.; Xia, X.C. Catalogue of Gene Symbols for Wheat: 2017 Supplement.KOMUGI Wheat Genetic Resource Database. Available online: http://shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2017.pdf (accessed on 5 March 2021).
- Li, H.; Dong, Z.; Ma, C.; Xia, Q.; Tian, X.; Sehgal, S.; Koo, D.H.; Friebe, B.; Ma, P.; Liu, W. A spontaneous wheat-Aegilops longissima translocation carrying Pm66 confers resistance to powdery mildew. Theor. Appl. Genet. 2020, 133, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Liu, R.; Ma, P.; Du, H.; Zhang, H.; Wu, Q.; Yang, L.; Gong, S.; Liu, T.; Huo, N.; et al. Characterization of Pm68, a new powdery mildew resistance gene on chromosome 2BS of Greek durum wheat TRI 1796. Theor. Appl. Genet. 2021, 134, 53–62. [Google Scholar] [CrossRef]
- Green, A.J.; Berger, G.; Griffey, C.A.; Pitman, R.; Thomason, W.; Balota, M. Genetic resistance to and effect of leaf rust and powdery mildew on yield and its components in 50 soft red winter wheat cultivars. Crop. Prot. 2014, 64, 177–186. [Google Scholar] [CrossRef]
- Summers, R.W.; Brown, J.K.M. Constraints on breeding for disease resistance in commercially competitive wheat cultivars. Plant. Pathol. 2013, 62, 115–121. [Google Scholar] [CrossRef]
- Xu, H.; Yi, Y.; Ma, P.; Qie, Y.; Fu, X.; Xu, Y.; Zhang, X.; An, D. Molecular tagging of a new broad-spectrum powdery mildew resistance allele Pm2c in Chinese wheat landrace Niaomai. Theor. Appl. Genet. 2015, 128, 2077–2084. [Google Scholar] [CrossRef]
- He, R.; Chang, Z.; Yang, Z.; Yuan, Z.; Zhan, H.; Zhang, X.; Liu, J. Inheritance and mapping of powdery mildew resistance gene Pm43 introgressed from Thinopyrum intermedium into wheat. Theor. Appl. Genet. 2009, 118, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, J.; Zhang, X.; Zhang, Q.; Huang, J.; Chen, J.; Hartl, D.L.; Tian, D. Rapidly evolving R genes in diverse grass species confer resistance to rice blast disease. Proc. Natl. Acad. Sci. USA 2013, 110, 18572–18577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, A.; Xing, L.; Wang, X.; Yang, X.; Wang, W.; Sun, Y.; Qian, C.; Ni, J.; Chen, Y.; Liu, D. Serine/threonine kinase gene Stpk-V, a key member of powdery mildew resistance gene Pm21, confers powdery mildew resistance in wheat. Proc. Natl. Acad. Sci. USA 2011, 108, 7727–7732. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Wang, B.; Li, Q.; Wu, X.; Wang, F.; Liu, H.; Liu, Q. Analysis on the virulent genes of Erysiphe graminis f. sp. tritici and the resistance genes of wheat commercial cultivars in Shaanxi Province. J. Triticeae Crops 2009, 29, 706–711. [Google Scholar]
- Yang, L.; Xiang, L.; Zeng, F.; Wang, H.; Shi, W.; Yu, D. Virulence gene structure analysis of Blumeria graminis f. sp. tritici in Hubei. Plant. Prot. 2009, 35, 76–79. [Google Scholar]
- Luo, P.G.; Luo, H.Y.; Chang, Z.J.; Zhang, H.Y.; Zhang, M.; Ren, Z.L. Characterization and chromosomal location of Pm40 in common wheat: A new gene for resistance to powdery mildew derived from Elytrigia intermedium. Theor. Appl. Genet. 2009, 118, 1059–1064. [Google Scholar] [CrossRef]
- Zhong, S.; Ma, L.; Fatima, S.A.; Yang, J.; Chen, W.; Liu, T.; Hu, Y.; Li, Q.; Guo, J.; Zhang, M. Collinearity analysis and high-density genetic mapping of the wheat powdery mildew resistance gene Pm40 in PI 672538. PLoS ONE 2016, 11, e0164815. [Google Scholar] [CrossRef]
- Hewitt, T.; Müller, M.C.; Molnár, I.; Mascher, M.; Holušová, K.; Šimková, H.; Kunz, L.; Zhang, J.; Li, J.; Bhatt, D. A highly differentiated region of wheat chromosome 7AL encodes a Pm1a immune receptor that recognizes its corresponding AvrPm1a effector from Blumeria graminis. New Phytol. 2021, 229, 2812–2826. [Google Scholar] [CrossRef]
- Sánchez-Martín, J.; Steuernagel, B.; Ghosh, S.; Herren, G.; Hurni, S.; Adamski, N.; Vrána, J.; Kubaláková, M.; Krattinger, S.G.; Wicker, T. Rapid gene isolation in barley and wheat by mutant chromosome sequencing. Genome Biol. 2016, 17, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Yahiaoui, N.; Srichumpa, P.; Dudler, R.; Keller, B. Genome analysis at different ploidy levels allows cloning of the powdery mildew resistance gene Pm3b from hexaploid wheat. Plant J. 2004, 37, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Guo, G.; Wang, Y.; Hu, T.; Wang, L.; Li, J.; Qiu, D.; Li, Y.; Wu, Q.; Lu, P. A rare single nucleotide variant in Pm5e confers powdery mildew resistance in common wheat. New Phytol. 2020, 228, 1011–1026. [Google Scholar] [CrossRef] [PubMed]
- Hurni, S.; Brunner, S.; Buchmann, G.; Herren, G.; Jordan, T.; Krukowski, P.; Wicker, T.; Yahiaoui, N.; Mago, R.; Keller, B. Rye Pm8 and wheat Pm3 are orthologous genes and show evolutionary conservation of resistance function against powdery mildew. Plant J. 2013, 76, 957–969. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Zhu, S.; Zhao, R.; Jiang, Z.; Ji, Y.; Ji, J.; Qiu, D.; Li, H.; Bie, T. Pm21, encoding a typical CC-NBS-LRR protein, confers broad-spectrum resistance to wheat powdery mildew disease. Mol. Plant 2018, 11, 879–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, P.; Guo, L.; Wang, Z.; Li, B.; Li, J.; Li, Y.; Qiu, D.; Shi, W.; Yang, L.; Wang, N. A rare gain of function mutation in a wheat tandem kinase confers resistance to powdery mildew. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krattinger, S.G.; Lagudah, E.S.; Spielmeyer, W.; Singh, R.P.; Huerta-Espino, J.; McFadden, H.; Bossolini, E.; Selter, L.L.; Keller, B. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 2009, 323, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Dong, L.; Li, B.; Wang, Z.; Xie, J.; Qiu, D.; Li, Y.; Shi, W.; Yang, L.; Wu, Q. A CNL protein in wild emmer wheat confers powdery mildew resistance. New Phytol. 2020, 228, 1027–1037. [Google Scholar] [CrossRef]
- Moore, J.W.; Herrera-Foessel, S.; Lan, C.; Schnippenkoetter, W.; Ayliffe, M.; Huerta-Espino, J.; Lillemo, M.; Viccars, L.; Milne, R.; Periyannan, S. A recently evolved hexose transporter variant confers resistance to multiple pathogens in wheat. Nat. Genet. 2015, 47, 1494–1498. [Google Scholar] [CrossRef]
- Zou, S.; Wang, H.; Li, Y.; Kong, Z.; Tang, D. The NB-LRR gene Pm60 confers powdery mildew resistance in wheat. New Phytol. 2018, 218, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; Poland, J. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar]
- Kebede, A.Z.; Johnston, A.; Schneiderman, D.; Bosnich, W.; Harris, L.J. Transcriptome profiling of two maize inbreds with distinct responses to Gibberella ear rot disease to identify candidate resistance genes. BMC Genom. 2018, 19, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chittem, K.; Yajima, W.R.; Goswami, R.S.; Del Río Mendoza, L.E. Transcriptome analysis of the plant pathogen Sclerotinia sclerotiorum interaction with resistant and susceptible canola (Brassica napus) lines. PLoS ONE 2020, 15, e0229844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, L.; Li, Y.; Ma, C.; Tong, L.; Du, F.; Xu, M. Combined genome-wide association study and transcriptome analysis reveal candidate genes for resistance to Fusarium ear rot in maize. J. Integr. Plant Biol. 2020, 62, 1535–1551. [Google Scholar] [CrossRef] [PubMed]
- Klymiuk, V.; Yaniv, E.; Huang, L.; Raats, D.; Fatiukha, A.; Chen, S.; Feng, L.; Frenkel, Z.; Krugman, T.; Lidzbarsky, G. Cloning of the wheat Yr15 resistance gene sheds light on the plant tandem kinase-pseudokinase family. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kourelis, J.; Van Der Hoorn, R.A. Defended to the nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Luo, P.G.; Reng, T.H.; Jiang, H.R.; Yang, Z.J. Genetic analysis and chromosomal location of two new genes for resistance to powdery mildew in wheat (Triticum aestivum L.). Acta. Agron. Sin. 2007, 33, 1–8. [Google Scholar]
- Tang, Z.; Li, M.; Chen, L.; Wang, Y.; Ren, Z.; Fu, S. New types of wheat chromosomal structural variations in derivatives of wheat-rye hybrids. PLoS ONE 2014, 9, e110282. [Google Scholar] [CrossRef] [Green Version]
- Bento, M.; Gustafson, P.; Viegas, W.; Silva, M. Genome merger: From sequence rearrangements in triticale to their elimination in wheat–rye addition lines. Theor. Appl. Genet. 2010, 121, 489–497. [Google Scholar] [CrossRef]
- Beaulieu, J.; Jean, M.; Belzile, F. The allotetraploid Arabidopsis thaliana–Arabidopsis lyrata subsp. petraea as an alternative model system for the study of polyploidy in plants. Mol. Genet. Genom. 2009, 281, 421–435. [Google Scholar] [CrossRef]
- Li, X.; Liu, T.; Chen, W.; Zhong, S.; Zhang, H.; Tang, Z.; Chang, Z.; Wang, L.; Zhang, M.; Li, L. Wheat WCBP1 encodes a putative copper-binding protein involved in stripe rust resistance and inhibition of leaf senescence. BMC Plant. Biol. 2015, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhong, S.; Zhang, M.; Liang, Y.; Gong, G.; Chang, X.; Tan, F.; Yang, H.; Qiu, X.; Luo, L. Potential Role of Photosynthesis in the Regulation of Reactive Oxygen Species and Defence Responses to Blumeria graminis f. sp. tritici in Wheat. Int. J. Mol. Sci. 2020, 21, 5767. [Google Scholar] [CrossRef]
- Keller, B.; Wicker, T.; Krattinger, S.G. Advances in wheat and pathogen genomics: Implications for disease control. Annu. Rev. Phytopathol. 2018, 56, 67–87. [Google Scholar] [CrossRef] [Green Version]
- Dhokane, D.; Karre, S.; Kushalappa, A.C.; McCartney, C. Integrated metabolo-transcriptomics reveals Fusarium head blight candidate resistance genes in wheat QTL-Fhb2. PLoS ONE 2016, 11, e0155851. [Google Scholar]
- Cheng, B.; Ding, Y.Q.; Gao, X.; Cao, N.; Xin, Z.H.; Zhang, L.Y. The diversity of powdery mildew resistance gene loci among wheat germplasm in Southwest China. Cereal. Res. Commun. 2020, 48, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.J.; Shi, J.R.; Singh, S.; Fickus, E.W.; Costa, J.M.; Lewis, J.; Gill, B.S.; Ward, R.; Cregan, P.B. Development and mapping of microsatellite (SSR) markers in wheat. Theor. Appl. Genet 2005, 110, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Periyannan, S.; Moore, J.; Ayliffe, M.; Bansal, U.; Wang, X.; Huang, L.; Deal, K.; Luo, M.; Kong, X.; Bariana, H. The gene Sr33, an ortholog of barley Mla genes, encodes resistance to wheat stem rust race Ug99. Science 2013, 341, 786–788. [Google Scholar] [CrossRef]
- Lee, H.; Yeom, S. Plant NB-LRR proteins: Tightly regulated sensors in a complex manner. Brief Funct. Genom. 2015, 14, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Jach, G.; Görnhardt, B.; Mundy, J.; Logemann, J.; Pinsdorf, E.; Leah, R.; Schell, J.; Maas, C. Enhanced quantitative resistance against fungal disease by combinatorial expression of different barley antifungal proteins in transgenic tobacco. Plant J. 1995, 8, 97–109. [Google Scholar] [CrossRef]
- Xiao, Y.; Heu, S.; Yi, J.; Lu, Y.; Hutcheson, S.W. Identification of a putative alternate sigma factor and characterization of a multicomponent regulatory cascade controlling the expression of Pseudomonas syringae pv. syringae Pss61 hrp and hrmA genes. J. Bacteriol. 1994, 176, 1025–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockrum, C.; Kaneshiro, K.R.; Rechtsteiner, A.; Tabuchi, T.M.; Strome, S. A primer for generating and using transcriptome data and gene sets. Development 2020, 147, dev193854. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Ye, Z.; Stanton, R. Misuse of RPKM or TPM normalization when comparing across samples and sequencing protocols. RNA 2020, 26, 903–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devonshire, A.S.; Elaswarapu, R.; Foy, C.A. Applicability of RNA standards for evaluating RT-qPCR assays and platforms. BMC Genom. 2011, 12, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bai, S.; Liu, J.; Chang, C.; Zhang, L.; Maekawa, T.; Wang, Q.; Xiao, W.; Liu, Y.; Chai, J.; Takken, F.L. Structure-function analysis of barley NLR immune receptor MLA10 reveals its cell compartment specific activity in cell death and disease resistance. PLoS Pathog. 2012, 8, e1002752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Césari, S.; Kanzaki, H.; Fujiwara, T.; Bernoux, M.; Chalvon, V.; Kawano, Y.; Shimamoto, K.; Dodds, P.; Terauchi, R.; Kroj, T. The NB-LRR proteins RGA 4 and RGA 5 interact functionally and physically to confer disease resistance. EMBO J. 2014, 33, 1941–1959. [Google Scholar] [CrossRef]
- Casey, L.W.; Lavrencic, P.; Bentham, A.R.; Cesari, S.; Ericsson, D.J.; Croll, T.; Turk, D.; Anderson, P.A.; Mark, A.E.; Dodds, P.N. The CC domain structure from the wheat stem rust resistance protein Sr33 challenges paradigms for dimerization in plant NLR proteins. Proc. Natl. Acad. Sci. USA 2016, 113, 12856–12861. [Google Scholar] [CrossRef] [Green Version]
- Takken, F.L.; Goverse, A. How to build a pathogen detector: Structural basis of NB-LRR function. Curr. Opin. Plant. Biol. 2012, 15, 375–384. [Google Scholar] [CrossRef]
- Burch-Smith, T.M.; Schiff, M.; Caplan, J.L.; Tsao, J.; Czymmek, K.; Dinesh-Kumar, S.P. Correction: A Novel Role for the TIR Domain in Association with Pathogen-Derived Elicitors. PLoS Biol. 2016, 14, e1002374. [Google Scholar] [CrossRef] [Green Version]
- Kwok, P.; Chen, X. Detection of single nucleotide polymorphisms. Curr. Issues Mol. Biol. 2003, 5, 43–60. [Google Scholar] [PubMed]
- Liu, Z.; Xu, M.; Xiang, Z.; Li, X.; Chen, W.; Luo, P. Registration of the novel wheat lines L658, L693, L696, and L699, with resistance to Fusarium Head blight, stripe rust, and powdery mildew. J. Plant. Regist. 2015, 9, 121–124. [Google Scholar] [CrossRef]
- Zhao, Z.; Sun, H.; Song, W.; Lu, M.; Huang, J.; Wu, L.; Wang, X.; Li, H. Genetic analysis and detection of the gene MlLX99 on chromosome 2BL conferring resistance to powdery mildew in the wheat cultivar Liangxing 99. Theor. Appl. Genet. 2013, 126, 3081–3089. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.L.; Luo, L.Y.; Liang, Y.P.; Hu, Y.T.; Luo, P.G.; Gong, G.S.; Chen, H.B.; Khaskheli, M.I.; Liu, T.G.; Chen, W.Q. Papilla formation, defense gene expression and HR contribute to the powdery mildew resistance of the novel wheat line L699 carrying Pm40 gene. Physiol. Mol. Plant. Pathol. 2019, 106, 208–216. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEf. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Kala, N.; Babu, S.; Manjeu, J.; Aadivalavan, A.; Khan, R. Allele-specific polymerase chain reaction for the detection of single nucleotide polymorphism in amlodipine-induced gingival enlargement. J. Clin. Pharm. Ther. 2017, 43, 110–113. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Faheem, M.; Li, Y.; Arshad, M.; Jiangyue, C.; Jia, Z.; Wang, Z.; Xiao, J.; Wang, H.; Cao, A.; Xing, L. A disulphide isomerase gene (PDI-V) from Haynaldia villosa contributes to powdery mildew resistance in common wheat. Sci. Rep. 2016, 6, 1–14. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Pfam Annotation | Swissprot Annotation | Eggnog Annotation | NR Annotation |
|---|---|---|---|---|
| TraesCS7B01G146500 | Protein tyrosine kinase | Serine/threonine-protein kinase | Signal transduction mechanisms | Serine/threonine-protein kinase |
| TraesCS7B01G148500 | Protein tyrosine kinase | Wall-associated receptor kinase | Signal transduction mechanisms | Wall-associated receptor kinase |
| TraesCS7B01G148600 | Protein tyrosine kinase | Wall-associated receptor kinase | Signal transduction mechanisms | Wall-associated receptor kinase |
| TraesCS7B01G148700 | Protein tyrosine kinase | Wall-associated receptor kinase | Signal transduction mechanisms | Wall-associated receptor kinase |
| TraesCS7B01G148800 | Protein tyrosine kinase | Wall-associated receptor kinase | Signal transduction mechanisms | Wall-associated receptor kinase |
| TraesCS7B01G162500 | Protein tyrosine kinase | Chitin elicitor receptor kinase | Signal transduction mechanisms | predicted protein |
| TraesCS7B01G164000 | NB-ARC domain | Putative disease resistance protein | Signal transduction mechanisms | resistance protein RGA2 |
| TraesCS7B01G164600 | Protein tyrosine kinase | PTI1-like tyrosine-protein kinase | Signal transduction mechanisms | PTI1-like tyrosine-protein kinase |
| Gene ID | Variants_Impact_HIGH | Variants_Impact_MODERATE | |||
|---|---|---|---|---|---|
| Frameshift | Splice Acceptor | Splice Donor | Stop Gained | Missense Variant | |
| TraesCS7B01G146300 | 0 | 0 | 0 | 0 | 3 |
| TraesCS7B01G149500 | 0 | 3 | 0 | 0 | 2 |
| TraesCS7B01G150600 | 0 | 0 | 6 | 0 | 2 |
| TraesCS7B01G150900 | 2 | 0 | 0 | 0 | 0 |
| TraesCS7B01G151800 | 0 | 0 | 0 | 0 | 3 |
| TraesCS7B01G152800 | 0 | 0 | 0 | 2 | 0 |
| TraesCS7B01G153200 | 0 | 0 | 0 | 0 | 1 |
| TraesCS7B01G155400 | 0 | 0 | 0 | 0 | 3 |
| TraesCS7B01G155600 | 0 | 0 | 0 | 0 | 1 |
| TraesCS7B01G157800 | 0 | 0 | 0 | 0 | 2 |
| TraesCS7B01G159900 | 0 | 0 | 0 | 0 | 1 |
| TraesCS7B01G160100 | 0 | 0 | 0 | 0 | 2 |
| TraesCS7B01G160200 | 0 | 0 | 0 | 0 | 2 |
| TraesCS7B01G160700 | 0 | 0 | 0 | 0 | 1 |
| TraesCS7B01G160800 | 1 | 0 | 0 | 0 | 3 |
| TraesCS7B01G160900 | 0 | 0 | 0 | 0 | 1 |
| TraesCS7B01G161200 | 0 | 0 | 0 | 0 | 5 |
| TraesCS7B01G161300 | 0 | 0 | 0 | 0 | 4 |
| TraesCS7B01G161400 | 1 | 0 | 0 | 0 | 2 |
| TraesCS7B01G161500 | 0 | 0 | 0 | 0 | 2 |
| TraesCS7B01G161600 | 0 | 0 | 0 | 0 | 2 |
| TraesCS7B01G162300 | 2 | 0 | 0 | 0 | 1 |
| TraesCS7B01G162500 | 0 | 0 | 0 | 0 | 3 |
| TraesCS7B01G162600 | 0 | 0 | 0 | 0 | 1 |
| TraesCS7B01G162700 | 0 | 0 | 0 | 0 | 5 |
| TraesCS7B01G162900 | 0 | 0 | 0 | 0 | 1 |
| TraesCS7B01G163000 | 0 | 0 | 0 | 0 | 1 |
| TraesCS7B01G163400 | 0 | 0 | 0 | 1 | 0 |
| TraesCS7B01G163500 | 0 | 0 | 0 | 0 | 1 |
| TraesCS7B01G163800 | 0 | 0 | 0 | 0 | 2 |
| TraesCS7B01G164000 | 0 | 0 | 0 | 0 | 2 |
| TraesCS7B01G164100 | 0 | 1 | 0 | 0 | 0 |
| TraesCS7B01G164200 | 0 | 0 | 0 | 0 | 1 |
| TraesCS7B01G164500 | 0 | 0 | 0 | 0 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Zhong, S.; Chen, C.; Yang, H.; Chen, W.; Tan, F.; Zhang, M.; Chen, W.; Ren, T.; Li, Z.; et al. Identification and Cloning of a CC-NBS-NBS-LRR Gene as a Candidate of Pm40 by Integrated Analysis of Both the Available Transcriptional Data and Published Linkage Mapping. Int. J. Mol. Sci. 2021, 22, 10239. https://doi.org/10.3390/ijms221910239
Yang H, Zhong S, Chen C, Yang H, Chen W, Tan F, Zhang M, Chen W, Ren T, Li Z, et al. Identification and Cloning of a CC-NBS-NBS-LRR Gene as a Candidate of Pm40 by Integrated Analysis of Both the Available Transcriptional Data and Published Linkage Mapping. International Journal of Molecular Sciences. 2021; 22(19):10239. https://doi.org/10.3390/ijms221910239
Chicago/Turabian StyleYang, Huai, Shengfu Zhong, Chen Chen, Hao Yang, Wei Chen, Feiquan Tan, Min Zhang, Wanquan Chen, Tianheng Ren, Zhi Li, and et al. 2021. "Identification and Cloning of a CC-NBS-NBS-LRR Gene as a Candidate of Pm40 by Integrated Analysis of Both the Available Transcriptional Data and Published Linkage Mapping" International Journal of Molecular Sciences 22, no. 19: 10239. https://doi.org/10.3390/ijms221910239