
The Role of Glycosyltransferases in Colorectal Cancer

 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Glycosyltransferase Gene Expression Profile in Colorectal Cancer

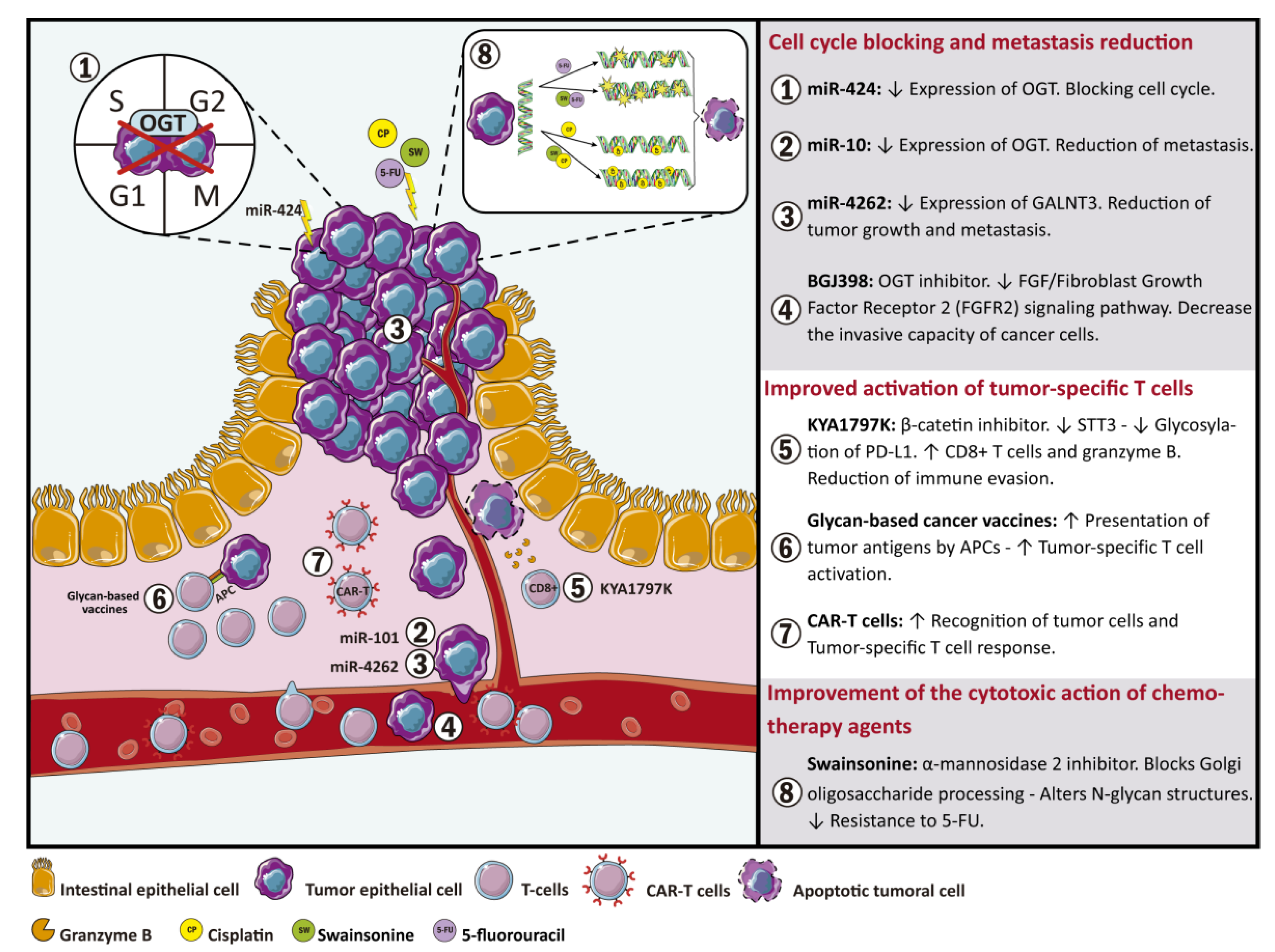{kind=link}
| A. Upregulated glycosyltransferase genes in CRC. Cellular effects induced by the upregulation of genes encoding glycosyltransferases are described. | |||
| Glycosyltransferases | Gene | Effects on Cancer Cells | References |
| Beta 3-glycosyltransferases | B3GNT3 | Cell migration. Cell invasion. Maintenance of CSCs. | Ashkani 2016 [22]. Barkeer 2018 [55]. |
| B3GNT8 | Cell migration. Cell invasion. Resistance to 5-FU. | Ashkani 2016 [22]. Ishida 2005 [27]. Ni 2014 [33]. Shen 2014 [56]. | |
| C1GALT1 | Induction of stem-like cell properties. Cell survival. Cell migration. Cell invasion. Radioresistance. | Hung 2014 [26]. Zhang 2018 [37]. | |
| Beta 4-glycosyltransferases | B4GALNT3 | Cell migration. Cell invasion. Maintenance of CSCs. | Che 2014 [23]. |
| Fucosyltransferases | FUT1 | Cell proliferation. Cell migration. Cell invasion. Metastasis. EMT. Maintenance of CSCs. | Ashkani 2016 [22]. Lai 2019 [43]. Petretti 2000 [57]. |
| FUT2 | Cell proliferation. Cell adhesion to extracellular matrix. Cell migration. Cell invasion. Metastasis. EMT. Maintenance of CSCs. | Ashkani 2016 [22]. Lai 2019 [43]. | |
| FUT3 | TGF-β-induced EMT. Cell adhesion to endothelium. Cell migration. | Ashkani 2016 [22]. Meng 2017 [58]. Padró 2011 [44]. Hirakawa 2014 [42]. | |
| FUT4 | MDR. | Ashkani 2016 [22]. Cheng 2013 [41]. Petretti 2000 [57]. | |
| FUT5 | Cell adhesion to endothelium. Cell migration. | Ashkani 2016 [22]. Padró 2011 [44]. | |
| FUT6 | TGF-β-induced EMT. MDR. Tumor progression. Metastasis. | Ashkani 2016 [22]. Cheng 2013 [41]. Hirakawa 2014 [42]. Sethi 2014 [31]. | |
| FUT8 | Tumor progression. Cell migration. Cell invasion. Metastasis. TGF-β-induced EMT. Tumor immune evasion. EGF-mediated cellular growth. | Sethi 2014 [31]. Tu 2017 [45]. Bastian 2021 [40]. | |
| Mannosyl-glycoprotein N-acetylglucosaminyl transferases | MGAT4B | Tumor progression. Metastasis. | Ashkani 2016 [22]. |
| MGAT5A | Tumor progression. Metastasis. Cell invasion. ↓ Anti-VEGF effectivity. Increase CCSC population. | Murata 2000 [29]. Guo 2014 [59]. Kim 2008 [38]. Croci 2014 [39]. Petretti 2000 [57]. | |
| O-linked N-acetylglucosaminyl transferases | OGT | Cell proliferation. Cell migration. Cell invasion. | Xu 2019 [32]. |
| Sialyltransferases | ST6GAL1 | Cell migration Cell invasion. Cell survival. Induction of stem-like cell properties. Chemotherapy resistance. | Ashkani 2016 [22]. Schultz 2016 [50]. Swindall 2011 [51]. Park 2012 [49]. Sethi 2014 [31]. |
| ST6GALNAC1 | ↑Sialyl-Tn expression. Maintenance of CSCs. Resistance to 5-FU. | Ashkani 2016 [22]. Marcos 2004 [60]. Ogawa 2017 [48]. | |
| B. Downregulated glycosyltransferase genes in CRC. Cellular effects induced by the downregulation of genes encoding glycosyltransferases are described. | |||
| Glycosyltransferases | Gene | Effects on Cancer cells | References |
| Beta 3-glycosyltransferases | B3GNT1 | Ashkani 2016 [22]. | |
| B3GNT6 | Cell migration. Cell invasion. Metastasis. EMT. | Iwai 2005 [28]. Gupta 2020 [61]. | |
| Polypeptide N-acetylgalactosaminyl transferases | GALNT6 | Poor differentiation. Cell migration. Cell invasion. Chemoresistance to 5-FU. ↑Tn-antigen expression. | Noda 2018 [30]. |
| Glucosaminyl (N-acetyl)transferases/ xylosyltransferases | GCNT3 | Cell proliferation. Cell adhesion. Cell migration. Cell invasion. Cell survival. Tumor growth. Chemoresistance to 5-FU. | Huang 2006 [25]. González-Vallinas 2015 [24]. Fernández 2018 [16]. |
| Fucosyltransferases | FUT9 | Cell migration. Metastasis. | Ashkani 2016 [22]. Auslander 2017 [46]. |
| Mannosyl-glycoprotein N-acetylglucosaminyl transferases | MGAT3 | Tumor progression. Metastasis. | Ashkani 2016 [22]. |
| MGAT5b | Tumor progression. Metastasis. | Ashkani 2016 [22]. | |
| Sialyltransferases | ST3GAL1 | Ashkani 2016 [22]. | |
| ST3GAL3 | Tumor progression. Metastasis. | Ashkani 2016 [22]. Sethi 2014 [31]. | |
| ST6GALNAC2 | Metastasis. | Ashkani 2016 [22]. Murugaesu 2014 [52]. Ferrer 2014 [62]. | |
| ST6GALNAC3 | Tumor progression. | Ashkani 2016 [22]. Haldrup 2018 [63]. | |
| ST6GALNAC6 | Inflammation-driven carcinogenesis. | Ashkani 2016 [22]. Huang 2020 [64]. | |
| ST8SIA1 | Ashkani 2016 [22]. | ||
| ST8SIA3 | Ashkani 2016 [22]. | ||
| ST8SIA4 | Cell proliferation. Cell migration. Cell invasion. | Ashkani 2016 [22]. Ma 2017 [53]. | |
| ST8SIA5 | Ashkani 2016 [22]. | ||
3. Glycosylated Molecules in Colorectal Cancer
| Target Molecules | Glycosylation | Effects on Cancer Cells | References |
|---|---|---|---|
| Annexin A1 | GlcNAcylation | Mitosis. Apoptosis. Cell differentiation. | Li et al. [20]. Yang et al. [74]. |
| HSP90 | GlcNAcylation | Cell viability. Cancer progression. | Li et al. [20]. Zou et al. [78]. Overath et al. [80]. |
| Carcinoembryonic antigen (CEA) | ↑ Fucose ↑ Mannose ↑ Thomsen–Friedenreich antigen ↓ N-acetylgalactosamine ↓ N-acetylglucosamine ↓ Galactose | Immune tolerance. CRC tumorigenesis. CRC progression. | Zhao et al. [101]. van Gisbergen et al. [91]. |
| IGFBP3 | ↑Sialylation (α2,3) ↓Fucosylation ↓GlcNAcylation | Cell proliferation. Cell survival. Cell differentiation. Cell migration. | Zámorová et al. [93]. |
| Decorin | O-glycosylation. | CRC development and progression. Metastasis. Cell-cell adhesion. Cell migration. | Wei et al. [102]. |
| SORBS1 | O-glycosylation. | CRC development and progression. Metastasis. Cell-cell adhesion. | Wei et al. [102]. |
| EGFR | Sialylation (Loss of α2,6 sialylation) Modification of N- with LacdiNAc structures. | Cell proliferation. Tumor growth. Cancer cell survival. Attenuated cancer cell differentiation. Chemoresistance. | Li et al. [92]. Che et al. [23]. Park et al. [49]. |
| TGF-β receptors | Fucosylation | EMT. Metastasis. | Hirakawa et al. [42]. |
| MucinsMUC1 MUC2 | O-glycosylation | Alteration of the interactions of mammalian lectin receptors with tumor cells. Immune dysregulation. Loss of mucosal barrier integrity. Gut dysbiosis. | Pothuraju et al. [82]. Peixoto et al. [69]. Brockhausen et al. [103]. Venkitachalam et al. [17]. Kawashima et al. [19]. Arike et al. [86]. |
| Podoplanin | O-glycosylation | Cell migration. Cell invasion. | Liu et al. [104]. |
| β1 integrin | α2-6 sialylation | Cell adhesion. Cell motility. Cell migration. Tumor cell survival. | Seales et al. [88]. Zhuo et al. [105]. |
| Fas (CD95) | α2-6 sialylation | Anti-apoptotic effect. | Swindall et al. [51]. |
| PD-L1 | N-glycosylation | Immune response evasion. | Ruan et al. [106]. |
| CD147 | Modification with Beta1,6-branched polylactosamine structures. | Cell migration. Cell invasion. Metastasis. | Ni et al. [33]. |
| Glycosphingolipids | ↑ Fucosylation ↑ Sialylation ↓ Glycans acetylation ↓ Glycans sulfation | Cell proliferation. Cell migration. Chemoresistance. EMT. Metastasis. Reduced apoptosis. Poor cell differentiation. | Holst et al. [10]. Misonou et al. [100]. Cumin et al. [9]. Gb3: Distler et al. [107]. Gb4: Park et al. [108]. GCS: Haynes et al. [109]. NEU3: Yamaguchi et al. [110]. GD1a and GM1: Kwak et al. [111]. α-GalCer: Yoshioka et al. [112]. GM3: Chung et al. [113]. |
4. Implications of Glycosylation and Glycosyltransferases in Colorectal Cancer Therapy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Araghi, M.; Soerjomataram, I.; Jenkins, M.; Brierley, J.; Morris, E.; Bray, F.; Arnold, M. Global trends in colorectal cancer mortality: Projections to the year 2035. Int. J. Cancer 2019, 144, 2992–3000. [Google Scholar] [CrossRef] [Green Version]
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, J.C.M., Jr.; Morgado-Díaz, J.A. The role of N-glycans in colorectal cancer progression: Potential biomarkers and therapeutic applications. Oncotarget 2016, 7, 19395–19413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoja-Łukowicz, D.; Link-Lenczowski, P.; Carpentieri, A.; Amoresano, A.; Pocheć, E.; Artemenko, K.A.; Bergquist, J.; Lityńska, A. L1CAM from human melanoma carries a novel type of N-glycan with Galβ1-4Galβ1- motif. Involvement of N-linked glycans in migratory and invasive behaviour of melanoma cells. Glycoconj. J. 2013, 30, 205–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julien, S.; Ivetic, A.; Grigoriadis, A.; QiZe, D.; Burford, B.; Sproviero, D.; Picco, G.; Gillett, C.; Papp, S.L.; Schaffer, L.; et al. Selectin ligand sialyl-Lewis x antigen drives metastasis of hormone-dependent breast cancers. Cancer Res. 2011, 71, 7683–7693. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Ahn, Y.H.; Song, K.J.; Kang, J.G.; Lee, J.H.; Jeon, S.K.; Kim, H.-C.; Yoo, J.S.; Ko, J.-H. Overexpression and β-1,6-N-acetylglucosaminylation-initiated aberrant glycosylation of TIMP-1: A “double whammy” strategy in colon cancer progression. J. Biol. Chem. 2012, 287, 32467–32478. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Liu, Q.; He, F.; Zhu, W.; Hu, L.; Guo, L.; Zhang, J. The role of N-acetylglucosaminyltransferases V in the malignancy of human hepatocellular carcinoma. Exp. Mol. Pathol. 2012, 93, 8–17. [Google Scholar] [CrossRef]
- Cumin, C.; Huang, Y.L.; Everest-Dass, A.; Jacob, F. Deciphering the Importance of Glycosphingolipids on Cellular and Molecular Mechanisms Associated with Epithelial-to-Mesenchymal Transition in Cancer. Biomolecules 2021, 11, 62. [Google Scholar] [CrossRef]
- Holst, S.; Stavenhagen, K.; Balog, C.I.; Koeleman, C.A.; McDonnell, L.M.; Mayboroda, O.A.; Verhoeven, A.; Mesker, W.E.; Tollenaar, R.A.; Deelder, A.M.; et al. Investigations on aberrant glycosylation of glycosphingolipids in colorectal cancer tissues using liquid chromatography and matrix-assisted laser desorption time-of-flight mass spectrometry (MALDI-TOF-MS). Mol. Cell. Proteom. 2013, 12, 3081–3093. [Google Scholar] [CrossRef] [Green Version]
- Holst, S.; Wuhrer, M.; Rombouts, Y. Glycosylation characteristics of colorectal cancer. Adv. Cancer Res. 2015, 126, 203–256. [Google Scholar]
- Varki, A. Biological roles of glycans. Glycobiology 2017, 27, 3–49. [Google Scholar] [CrossRef] [Green Version]
- Marth, J.D.; Grewal, P.K. Mammalian glycosylation in immunity. Nat. Rev. Immunol. 2008, 8, 874–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef]
- Stanley, P. Golgi glycosylation. Cold Spring Harb. Perspect. Biol. 2011, 3, a005199. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.P.; Sánchez-Martínez, R.; Vargas, T.; Herranz, J.; Martín-Hernández, R.; Mendiola, M.; Hardisson, D.; Reglero, G.; Feliu, J.; Redondo, A.; et al. The role of glycosyltransferase enzyme GCNT3 in colon and ovarian cancer prognosis and chemoresistance. Sci. Rep. 2018, 8, 8485. [Google Scholar] [CrossRef]
- Venkitachalam, S.; Guda, K. Altered glycosyltransferases in colorectal cancer. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 5–7. [Google Scholar] [CrossRef]
- Venkitachalam, S.; Revoredo, L.; Varadan, V.; Fecteau, R.E.; Ravi, L.; Lutterbaugh, J.; Markowitz, S.D.; Willis, J.E.; Gerken, T.A.; Guda, K. Biochemical and functional characterization of glycosylation-associated mutational landscapes in colon cancer. Sci. Rep. 2016, 6, 23642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawashima, H. Roles of the gel-forming MUC2 mucin and its O-glycosylation in the protection against colitis and colorectal cancer. Biol. Pharm. Bull. 2012, 35, 1637–1641. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wen, T.; Zhu, M.; Li, L.; Wei, J.; Wu, X.; Guo, M.; Liu, S.; Zhao, H.; Xia, S.; et al. Glycoproteomic analysis of tissues from patients with colon cancer using lectin microarrays and nanoLC-MS/MS. Mol. Biosyst. 2013, 9, 1877–1887. [Google Scholar] [CrossRef]
- Dall’olio, F. Protein glycosylation in cancer biology: An overview. Clin. Mol. Pathol. 1996, 49, M126–M135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashkani, J.; Naidoo, K.J. Glycosyltransferase Gene Expression Profiles Classify Cancer Types and Propose Prognostic Subtypes. Sci. Rep. 2016, 6, 26451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, M.I.; Huang, J.; Hung, J.S.; Lin, Y.C.; Huang, M.J.; Lai, H.S.; Hsu, W.M.; Liang, J.T.; Huang, M.C. β1, 4-N-acetylgalactosaminyltransferase III modulates cancer stemness through EGFR signaling pathway in colon cancer cells. Oncotarget 2014, 5, 3673–3684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Vallinas, M.; Vargas, T.; Moreno-Rubio, J.; Molina, S.; Herranz, J.; Cejas, P.; Burgos, E.; Aguayo, C.; Custodio, A.; Reglero, G.; et al. Clinical relevance of the differential expression of the glycosyltransferase gene GCNT3 in colon cancer. Eur. J. Cancer 2015, 51, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.C.; Chen, H.Y.; Huang, H.C.; Huang, J.; Liang, J.T.; Shen, T.L.; Lin, N.Y.; Ho, C.C.; Cho, I.M.; Hsu, S.M. C2GnT-M is downregulated in colorectal cancer and its re-expression causes growth inhibition of colon cancer cells. Oncogene 2006, 25, 3267–3276. [Google Scholar] [CrossRef]
- Hung, J.S.; Huang, J.; Lin, Y.C.; Huang, M.J.; Lee, P.H.; Lai, H.S.; Liang, J.T.; Huang, M.C. C1GALT1 overexpression promotes the invasive behavior of colon cancer cells through modifying O-glycosylation of FGFR2. Oncotarget 2014, 5, 2096–2106. [Google Scholar] [CrossRef] [Green Version]
- Ishida, H.; Togayachi, A.; Sakai, T.; Iwai, T.; Hiruma, T.; Sato, T.; Okubo, R.; Inaba, N.; Kudo, T.; Gotoh, M.; et al. A novel beta1,3-N-acetylglucosaminyltransferase (beta3Gn-T8), which synthesizes poly-N-acetyllactosamine, is dramatically upregulated in colon cancer. FEBS Lett. 2005, 579, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Iwai, T.; Kudo, T.; Kawamoto, R.; Kubota, T.; Togayachi, A.; Hiruma, T.; Okada, T.; Kawamoto, T.; Morozumi, K.; Narimatsu, H. Core 3 synthase is down-regulated in colon carcinoma and profoundly suppresses the metastatic potential of carcinoma cells. Proc. Natl. Acad. Sci. USA 2005, 102, 4572–4577. [Google Scholar] [CrossRef] [Green Version]
- Murata, K.; Miyoshi, E.; Kameyama, M.; Ishikawa, O.; Kabuto, T.; Sasaki, Y.; Hiratsuka, M.; Ohigashi, H.; Ishiguro, S.; Ito, S.; et al. Expression of N-acetylglucosaminyltransferase V in colorectal cancer correlates with metastasis and poor prognosis. Clin. Cancer Res. 2000, 6, 1772–1777. [Google Scholar] [PubMed]
- Noda, M.; Okayama, H.; Tachibana, K.; Sakamoto, W.; Saito, K.; Thar Min, A.K.; Ashizawa, M.; Nakajima, T.; Aoto, K.; Momma, T.; et al. Glycosyltransferase Gene Expression Identifies a Poor Prognostic Colorectal Cancer Subtype Associated with Mismatch Repair Deficiency and Incomplete Glycan Synthesis. Clin. Cancer Res. 2018, 24, 4468–4481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethi, M.K.; Thaysen-Andersen, M.; Smith, J.T.; Baker, M.S.; Packer, N.H.; Hancock, W.S.; Fanayan, S. Comparative N-glycan profiling of colorectal cancer cell lines reveals unique bisecting GlcNAc and α-2,3-linked sialic acid determinants are associated with membrane proteins of the more metastatic/aggressive cell lines. J. Proteome Res. 2014, 13, 277–288. [Google Scholar] [CrossRef]
- Xu, D.; Wang, W.; Bian, T.; Yang, W.; Shao, M.; Yang, H. Increased expression of O-GlcNAc transferase (OGT) is a biomarker for poor prognosis and allows tumorigenesis and invasion in colon cancer. Int. J. Clin. Exp. Pathol. 2019, 12, 1305–1314. [Google Scholar]
- Ni, J.; Jiang, Z.; Shen, L.; Gao, L.; Yu, M.; Xu, X.; Zou, S.; Hua, D.; Wu, S. beta3GnT8 regulates the metastatic potential of colorectal carcinoma cells by altering the glycosylation of CD147. Oncol. Rep. 2014, 31, 1795–1801. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Hemler, M.E. Regulation of MMP-1 and MMP-2 production through CD147/extracellular matrix metalloproteinase inducer interactions. Cancer Res. 2001, 61, 2276–2281. [Google Scholar]
- Very, N.; Lefebvre, T.; El Yazidi-Belkoura, I. Drug resistance related to aberrant glycosylation in colorectal cancer. Oncotarget 2017, 9, 1380–1402. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Zhou, M.; Peng, L.; Kong, S.; Miao, R.; Shi, Y.; Sheng, H.; Li, L. Upregulation of CD147 promotes cell invasion, epithelial-to-mesenchymal transition and activates MAPK/ERK signaling pathway in colorectal cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 7432–7441. [Google Scholar] [PubMed]
- Zhang, C.; Deng, X.; Qiu, L.; Peng, F.; Geng, S.; Shen, L.; Luo, Z. Knockdown of C1GalT1 inhibits radioresistance of human esophageal cancer cells through modifying β1-integrin glycosylation. J. Cancer 2018, 9, 2666–2677. [Google Scholar] [CrossRef]
- Kim, Y.S.; Hwang, S.Y.; Kang, H.Y.; Sohn, H.; Oh, S.; Kim, J.Y.; Yoo, J.S.; Kim, Y.H.; Kim, C.H.; Jeon, J.H.; et al. Functional proteomics study reveals that N-Acetylglucosaminyltransferase V reinforces the invasive/metastatic potential of colon cancer through aberrant glycosylation on tissue inhibitor of metalloproteinase-1. Mol. Cell. Proteom. 2008, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croci, D.O.; Cerliani, J.P.; Dalotto-Moreno, T.; Mendez-Huergo, S.P.; Mascanfroni, I.D.; Dergan-Dylon, S.; Toscano, M.A.; Caramelo, J.J.; Garcia-Vallejo, J.J.; Ouyang, J.; et al. Glycosylation-dependent lectin-receptor interactions preserve angiogenesis in anti-VEGF refractory tumors. Cell 2014, 156, 744–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastian, K.; Scott, E.; Elliott, D.J.; Munkley, J. FUT8 Alpha-(1,6)-Fucosyltransferase in Cancer. Int. J. Mol. Sci. 2021, 22, 455. [Google Scholar] [CrossRef]
- Cheng, L.; Luo, S.; Jin, C.; Ma, H.; Zhou, H.; Jia, L. FUT family mediates the multidrug resistance of human hepatocellular carcinoma via the PI3K/Akt signaling pathway. Cell Death Dis. 2013, 4, e923. [Google Scholar] [CrossRef]
- Hirakawa, M.; Takimoto, R.; Tamura, F.; Yoshida, M.; Ono, M.; Murase, K.; Sato, Y.; Osuga, T.; Sato, T.; Iyama, S.; et al. Fucosylated TGF-β receptors transduces a signal for epithelial-mesenchymal transition in colorectal cancer cells. Br. J. Cancer 2014, 110, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Lai, T.-Y.; Chen, I.J.; Lin, R.-J.; Liao, G.-S.; Yeo, H.-L.; Ho, C.-L.; Wu, J.-C.; Chang, N.-C.; Lee, A.C.-L.; Yu, A.L. Fucosyltransferase 1 and 2 play pivotal roles in breast cancer cells. Cell Death Discov. 2019, 5, 74. [Google Scholar] [CrossRef] [PubMed]
- Padró, M.; Cobler, L.; Garrido, M.; de Bolós, C. Down-regulation of FUT3 and FUT5 by shRNA alters Lewis antigens expression and reduces the adhesion capacities of gastric cancer cells. Biochim. Biophys. Acta 2011, 1810, 1141–1149. [Google Scholar] [CrossRef]
- Tu, C.-F.; Wu, M.-Y.; Lin, Y.-C.; Kannagi, R.; Yang, R.-B. FUT8 promotes breast cancer cell invasiveness by remodeling TGF-β receptor core fucosylation. Breast Cancer Res. 2017, 19, 111. [Google Scholar] [CrossRef] [Green Version]
- Auslander, N.; Cunningham, C.E.; Toosi, B.M.; McEwen, E.J.; Yizhak, K.; Vizeacoumar, F.S.; Parameswaran, S.; Gonen, N.; Freywald, T.; Bhanumathy, K.K.; et al. An integrated computational and experimental study uncovers FUT9 as a metabolic driver of colorectal cancer. Mol. Syst. Biol. 2017, 13, 956. [Google Scholar] [CrossRef] [PubMed]
- Blanas, A.; Zaal, A.; van der Haar Àvila, I.; Kempers, M.; Kruijssen, L.; de Kok, M.; Popovic, M.A.; van der Horst, J.C.; van Vliet, S.J. FUT9-Driven Programming of Colon Cancer Cells towards a Stem Cell-Like State. Cancers 2020, 12, 2580. [Google Scholar] [CrossRef]
- Ogawa, T.; Hirohashi, Y.; Murai, A.; Nishidate, T.; Okita, K.; Wang, L.; Ikehara, Y.; Satoyoshi, T.; Usui, A.; Kubo, T.; et al. ST6GALNAC1 plays important roles in enhancing cancer stem phenotypes of colorectal cancer via the Akt pathway. Oncotarget 2017, 8, 112550–112564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.J.; Yi, J.Y.; Jin, Y.B.; Lee, Y.J.; Lee, J.S.; Lee, Y.S.; Ko, Y.G.; Lee, M. Sialylation of epidermal growth factor receptor regulates receptor activity and chemosensitivity to gefitinib in colon cancer cells. Biochem. Pharmacol. 2012, 83, 849–857. [Google Scholar] [CrossRef]
- Schultz, M.J.; Holdbrooks, A.T.; Chakraborty, A.; Grizzle, W.E.; Landen, C.N.; Buchsbaum, D.J.; Conner, M.G.; Arend, R.C.; Yoon, K.J.; Klug, C.A.; et al. The Tumor-Associated Glycosyltransferase ST6Gal-I Regulates Stem Cell Transcription Factors and Confers a Cancer Stem Cell Phenotype. Cancer Res. 2016, 76, 3978–3988. [Google Scholar] [CrossRef] [Green Version]
- Swindall, A.F.; Bellis, S.L. Sialylation of the Fas death receptor by ST6Gal-I provides protection against Fas-mediated apoptosis in colon carcinoma cells. J. Biol. Chem. 2011, 286, 22982–22990. [Google Scholar] [CrossRef] [Green Version]
- Murugaesu, N.; Iravani, M.; van Weverwijk, A.; Ivetic, A.; Johnson, D.A.; Antonopoulos, A.; Fearns, A.; Jamal-Hanjani, M.; Sims, D.; Fenwick, K.; et al. An in vivo functional screen identifies ST6GalNAc2 sialyltransferase as a breast cancer metastasis suppressor. Cancer Discov. 2014, 4, 304–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Zhao, X.; Liang, L.; Wang, G.; Li, Y.; Miao, X.; Zhao, Y. miR-146a and miR-146b promote proliferation, migration and invasion of follicular thyroid carcinoma via inhibition of ST8SIA4. Oncotarget 2017, 8, 28028–28041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall’Olio, F.; Trinchera, M. Epigenetic Bases of Aberrant Glycosylation in Cancer. Int. J. Mol. Sci. 2017, 18, 998. [Google Scholar] [CrossRef] [Green Version]
- Barkeer, S.; Chugh, S.; Karmakar, S.; Kaushik, G.; Rauth, S.; Rachagani, S.; Batra, S.K.; Ponnusamy, M.P. Novel role of O-glycosyltransferases GALNT3 and B3GNT3 in the self-renewal of pancreatic cancer stem cells. BMC Cancer 2018, 18, 1157. [Google Scholar] [CrossRef]
- Shen, L.; Yu, M.; Xu, X.; Gao, L.; Ni, J.; Luo, Z.; Wu, S. Knockdown of β3GnT8 reverses 5-fluorouracil resistance in human colorectal cancer cells via inhibition the biosynthesis of polylactosamine-type N-glycans. Int. J. Oncol. 2014, 45, 2560–2568. [Google Scholar] [CrossRef] [Green Version]
- Petretti, T.; Kemmner, W.; Schulze, B.; Schlag, P.M. Altered mRNA expression of glycosyltransferases in human colorectal carcinomas and liver metastases. Gut 2000, 46, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, L.; Xu, L.; Yang, Y.; Zhou, L.; Chang, Y.; Shi, T.; Tan, C.; An, H.; Zhu, Y.; Xu, J. High expression of FUT3 is linked to poor prognosis in clear cell renal cell carcinoma. Oncotarget 2017, 8, 61036–61047. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Nagy, T.; Pierce, M. Post-translational glycoprotein modifications regulate colon cancer stem cells and colon adenoma progression in Apc(min/+) mice through altered Wnt receptor signaling. J. Biol. Chem. 2014, 289, 31534–31549. [Google Scholar] [CrossRef] [Green Version]
- Marcos, N.T.; Pinho, S.; Grandela, C.; Cruz, A.; Samyn-Petit, B.; Harduin-Lepers, A.; Almeida, R.; Silva, F.; Morais, V.; Costa, J.; et al. Role of the human ST6GalNAc-I and ST6GalNAc-II in the synthesis of the cancer-associated sialyl-Tn antigen. Cancer Res. 2004, 64, 7050–7057. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Leon, F.; Rauth, S.; Batra, S.K.; Ponnusamy, M.P. A Systematic Review on the Implications of O-linked Glycan Branching and Truncating Enzymes on Cancer Progression and Metastasis. Cells 2020, 9, 446. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, C.M.; Reginato, M.J. Sticking to sugars at the metastatic site: Sialyltransferase ST6GalNAc2 acts as a breast cancer metastasis suppressor. Cancer Discov. 2014, 4, 275–277. [Google Scholar] [CrossRef] [Green Version]
- Haldrup, C.; Pedersen, A.L.; Øgaard, N.; Strand, S.H.; Høyer, S.; Borre, M.; Ørntoft, T.F.; Sørensen, K.D. Biomarker potential of ST6GALNAC3 and ZNF660 promoter hypermethylation in prostate cancer tissue and liquid biopsies. Mol. Oncol. 2018, 12, 545–560. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.C.; Cai, B.H.; Suen, C.S.; Lee, H.Y.; Hwang, M.J.; Liu, F.T.; Kannagi, R. BGN/TLR4/NF-B Mediates Epigenetic Silencing of Immunosuppressive Siglec Ligands in Colon Cancer Cells. Cells 2020, 9, 397. [Google Scholar] [CrossRef] [Green Version]
- Dube, D.H.; Bertozzi, C.R. Glycans in cancer and inflammation—Potential for therapeutics and diagnostics. Nat. Rev. Drug Discov. 2005, 4, 477–488. [Google Scholar] [CrossRef]
- Orntoft, T.F.; Vestergaard, E.M. Clinical aspects of altered glycosylation of glycoproteins in cancer. Electrophoresis 1999, 20, 362–371. [Google Scholar] [CrossRef]
- Christiansen, M.N.; Chik, J.; Lee, L.; Anugraham, M.; Abrahams, J.L.; Packer, N.H. Cell surface protein glycosylation in cancer. Proteomics 2014, 14, 525–546. [Google Scholar] [CrossRef]
- Jóźwiak, P.; Forma, E.; Bryś, M.; Krześlak, A. O-GlcNAcylation and Metabolic Reprograming in Cancer. Front. Endocrinol. 2014, 5, 145. [Google Scholar]
- Peixoto, A.; Relvas-Santos, M.; Azevedo, R.; Santos, L.L.; Ferreira, J.A. Protein Glycosylation and Tumor Microenvironment Alterations Driving Cancer Hallmarks. Front. Oncol. 2019, 9, 380. [Google Scholar] [CrossRef] [Green Version]
- Shental-Bechor, D.; Levy, Y. Effect of glycosylation on protein folding: A close look at thermodynamic stabilization. Proc. Natl. Acad. Sci. USA 2008, 105, 8256–8261. [Google Scholar] [CrossRef] [Green Version]
- Vagin, O.; Kraut, J.A.; Sachs, G. Role of N-glycosylation in trafficking of apical membrane proteins in epithelia. Am. J. Physiol. Ren. Physiol. 2009, 296, F459–F469. [Google Scholar] [CrossRef] [Green Version]
- Moraes, L.A.; Ampomah, P.B.; Lim, L.H.K. Annexin A1 in inflammation and breast cancer: A new axis in the tumor microenvironment. Cell Adhes. Migr. 2018, 12, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Lizarbe, M.A.; Barrasa, J.I.; Olmo, N.; Gavilanes, F.; Turnay, J. Annexin-Phospholipid Interactions. Functional Implications. Int. J. Mol. Sci. 2013, 14, 2652–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Qian, K. Protein O-GlcNAcylation: Emerging mechanisms and functions. Nat. Rev. Mol. Cell Biol. 2017, 18, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.P.; Zhang, K.; Wu, J.; Yang, X. O-GlcNAc signaling in cancer metabolism and epigenetics. Cancer Lett. 2015, 356, 244–250. [Google Scholar] [CrossRef] [Green Version]
- Fisi, V.; Miseta, A.; Nagy, T.A.-O. The Role of Stress-Induced O-GlcNAc Protein Modification in the Regulation of Membrane Transport. Oxidative Med. Cell. Longev. 2017, 2017, 1308692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuehlke, A.D.; Beebe, K.; Neckers, L.; Prince, T. Regulation and function of the human HSP90AA1 gene. Gene 2015, 570, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, M.; Bhatia, A.; Dong, H.; Jayaprakash, P.; Guo, J.; Sahu, D.; Hou, Y.; Tsen, F.; Tong, C.; O’Brien, K.; et al. Evolutionarily conserved dual lysine motif determines the non-chaperone function of secreted Hsp90alpha in tumour progression. Oncogene 2017, 36, 2160–2171. [Google Scholar] [CrossRef]
- Trepel, J.; Mollapour, M.; Giaccone, G.; Neckers, L. Targeting the dynamic HSP90 complex in cancer. Nat. Rev. Cancer 2010, 10, 537–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overath, T.; Kuckelkorn, U.; Henklein, P.; Strehl, B.; Bonar, D.; Kloss, A.; Siele, D.; Kloetzel, P.M.; Janek, K. Mapping of O-GlcNAc sites of 20 S proteasome subunits and Hsp90 by a novel biotin-cystamine tag. Mol. Cell Proteom. 2012, 11, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeland, E.; Belo, A.I.; Mongera, S.; van Die, I.; Meijer, G.A.; van Kooyk, Y. Differential glycosylation of MUC1 and CEACAM5 between normal mucosa and tumour tissue of colon cancer patients. Int. J. Cancer 2012, 131, 117–128. [Google Scholar] [CrossRef]
- Pothuraju, R.; Krishn, S.R.; Gautam, S.K.; Pai, P.; Ganguly, K.; Chaudhary, S.; Rachagani, S.; Kaur, S.; Batra, S.K. Mechanistic and Functional Shades of Mucins and Associated Glycans in Colon Cancer. Cancers (Basel) 2020, 12, 649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Ponce, C.; Navarro Quiroz, R.; Díaz Perez, A.; Aroca Martinez, G.; Cadena Bonfanti, A.; Acosta Hoyos, A.; Gómez Escorcia, L.; Hernández Agudelo, S.; Orozco Sánchez, C.; Villarreal Camacho, J.; et al. MicroRNAs overexpressed in Crohn’s disease and their interactions with mechanisms of epigenetic regulation explain novel aspects of Crohn’s disease pathogenesis. Clin. Epigenet. 2021, 13, 39. [Google Scholar] [CrossRef]
- Nishida, A.; Lau, C.W.; Zhang, M.; Andoh, A.; Shi, H.N.; Mizoguchi, E.; Mizoguchi, A. The membrane-bound mucin Muc1 regulates T helper 17-cell responses and colitis in mice. Gastroenterology 2012, 142, 865–874.e2. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, B.; Gupta, N.; Konowalchuk, J.D. MUC1 Mucin: A Putative Regulatory (Checkpoint) Molecule of T Cells. Front. Immunol. 2018, 9, 2391. [Google Scholar] [CrossRef] [Green Version]
- Arike, L.; Hansson, G.C. The Densely O-Glycosylated MUC2 Mucin Protects the Intestine and Provides Food for the Commensal Bacteria. J. Mol. Biol. 2016, 428, 3221–3229. [Google Scholar] [CrossRef] [Green Version]
- Läubli, H.; Borsig, L. Altered Cell Adhesion and Glycosylation Promote Cancer Immune Suppression and Metastasis. Front. Immunol. 2019, 10, 2120. [Google Scholar] [CrossRef] [Green Version]
- Seales, E.C.; Jurado, G.A.; Brunson, B.A.; Wakefield, J.K.; Frost, A.R.; Bellis, S.L. Hypersialylation of beta1 integrins, observed in colon adenocarcinoma, may contribute to cancer progression by up-regulating cell motility. Cancer Res. 2005, 65, 4645–4652. [Google Scholar] [CrossRef] [Green Version]
- Deschepper, F.M.; Zoppi, R.; Pirro, M.; Hensbergen, P.J.; Dall’Olio, F.; Kotsias, M.; Gardner, R.A.; Spencer, D.I.R.; Videira, P.A. L1CAM as an E-selectin Ligand in Colon Cancer. Int. J. Mol. Sci. 2020, 21, 8286. [Google Scholar] [CrossRef]
- De Bousser, E.; Meuris, L.; Callewaert, N.; Festjens, N. Human T cell glycosylation and implications on immune therapy for cancer. Hum. Vaccines Immunother. 2020, 16, 2374–2388. [Google Scholar] [CrossRef] [Green Version]
- Van Gisbergen, K.P.; Aarnoudse, C.A.; Meijer, G.A.; Geijtenbeek, T.B.; van Kooyk, Y. Dendritic cells recognize tumor-specific glycosylation of carcinoembryonic antigen on colorectal cancer cells through dendritic cell-specific intercellular adhesion molecule-3-grabbing nonintegrin. Cancer Res. 2005, 65, 5935–5944. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Al-Japairai, K.; Tao, Y.; Xiang, Z. RPN2 promotes colorectal cancer cell proliferation through modulating the glycosylation status of EGFR. Oncotarget 2017, 8, 72633–72651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zámorová, M.; Holazová, A.; Miljuš, G.; Robajac, D.; Šunderić, M.; Malenković, V.; Đukanović, B.; Gemeiner, P.; Katrlík, J.; Nedić, O. Analysis of changes in the glycan composition of serum, cytosol and membrane glycoprotein biomarkers of colorectal cancer using a lectin-based protein microarray. Anal. Methods 2017, 9, 2660–2666. [Google Scholar] [CrossRef]
- Baxter, R.C. Insulin-like growth factor binding protein-3 (IGFBP-3): Novel ligands mediate unexpected functions. J. Cell Commun. Signal. 2013, 7, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Dozmorov, M.; Oh, Y. IGFBP-3/IGFBP-3 Receptor System as an Anti-Tumor and Anti-Metastatic Signaling in Cancer. Cells 2020, 9, 1261. [Google Scholar] [CrossRef]
- Grkovic, S.; O’Reilly, V.C.; Han, S.; Hong, M.; Baxter, R.C.; Firth, S.M. IGFBP-3 binds GRP78, stimulates autophagy and promotes the survival of breast cancer cells exposed to adverse microenvironments. Oncogene 2013, 32, 2412–2420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, G.E.; Wilson, E.M.; Powell, D.; Oh, Y. Butyrate, a histone deacetylase inhibitor, activates the human IGF binding protein-3 promoter in breast cancer cells: Molecular mechanism involves an Sp1/Sp3 multiprotein complex. Endocrinology 2001, 142, 3817–3827. [Google Scholar] [CrossRef]
- Williams, A.C.; Smartt, H.; AM, H.Z.; Macfarlane, M.; Paraskeva, C.; Collard, T.J. Insulin-like growth factor binding protein 3 (IGFBP-3) potentiates TRAIL-induced apoptosis of human colorectal carcinoma cells through inhibition of NF-kappaB. Cell Death Differ. 2007, 14, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baricević, I.; Masnikosa, R.; Lagundzin, D.; Golubović, V.; Nedić, O. Alterations of insulin-like growth factor binding protein 3 (IGFBP-3) glycosylation in patients with breast tumours. Clin. Biochem. 2010, 43, 725–731. [Google Scholar] [CrossRef]
- Misonou, Y.; Shida, K.; Korekane, H.; Seki, Y.; Noura, S.; Ohue, M.; Miyamoto, Y. Comprehensive Clinico-Glycomic Study of 16 Colorectal Cancer Specimens: Elucidation of Aberrant Glycosylation and Its Mechanistic Causes in Colorectal Cancer Cells. J. Proteome Res. 2009, 8, 2990–3005. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhan, T.; Deng, Z.; Li, Q.; Liu, Y.; Yang, S.; Ji, D.; Li, Y. Glycan analysis of colorectal cancer samples reveals stage-dependent changes in CEA glycosylation patterns. Clin. Proteom. 2018, 15, 9. [Google Scholar] [CrossRef]
- Wei, H.; Cheng, Z.; Ouyang, C.; Zhang, Y.; Hu, Y.; Chen, S.; Wang, C.; Lu, F.; Zhang, J.; Wang, Y.; et al. Glycoprotein screening in colorectal cancer based on differentially expressed Tn antigen. Oncol. Rep. 2016, 36, 1313–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brockhausen, I. Mucin-type O-glycans in human colon and breast cancer: Glycodynamics and functions. EMBO Rep. 2006, 7, 599–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Cui, Y.; Yang, F.; Xu, Z.; Da, L.T.; Zhang, Y. Inhibition of polypeptide N-acetyl-α-galactosaminyltransferases is an underlying mechanism of dietary polyphenols preventing colorectal tumorigenesis. Bioorg. Med. Chem. 2019, 27, 3372–3382. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Y.; Chammas, R.; Bellis, S.L. Sialylation of beta1 integrins blocks cell adhesion to galectin-3 and protects cells against galectin-3-induced apoptosis. J. Biol. Chem. 2008, 283, 22177–22185. [Google Scholar] [CrossRef]
- Ruan, Z.; Liang, M.; Lai, M.; Shang, L.; Deng, X.; Su, X. KYA1797K down-regulates PD-L1 in colon cancer stem cells to block immune evasion by suppressing the β-catenin/STT3 signaling pathway. Int. Immunopharmacol. 2020, 78, 106003. [Google Scholar] [CrossRef]
- Distler, U.; Souady, J.; Hülsewig, M.; Drmić-Hofman, I.; Haier, J.; Friedrich, A.W.; Karch, H.; Senninger, N.; Dreisewerd, K.; Berkenkamp, S.; et al. Shiga toxin receptor Gb3Cer/CD77: Tumor-association and promising therapeutic target in pancreas and colon cancer. PLoS ONE 2009, 4, e6813. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Kwak, C.Y.; Shayman, J.A.; Kim, J.H. Globoside promotes activation of ERK by interaction with the epidermal growth factor receptor. Biochim. Biophys. Acta 2012, 1820, 1141–1148. [Google Scholar] [CrossRef] [Green Version]
- Haynes, T.A.; Filippov, V.; Filippova, M.; Yang, J.; Zhang, K.; Duerksen-Hughes, P.J. DNA damage induces down-regulation of UDP-glucose ceramide glucosyltransferase, increases ceramide levels and triggers apoptosis in p53-deficient cancer cells. Biochim. Biophys. Acta 2012, 1821, 943–953. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, K.; Shiozaki, K.; Moriya, S.; Koseki, K.; Wada, T.; Tateno, H.; Sato, I.; Asano, M.; Iwakura, Y.; Miyagi, T. Reduced susceptibility to colitis-associated colon carcinogenesis in mice lacking plasma membrane-associated sialidase. PLoS ONE 2012, 7, e41132. [Google Scholar] [CrossRef] [Green Version]
- Kwak, D.H.; Ryu, J.-S.; Kim, C.-H.; Ko, K.; Ma, J.Y.; Hwang, K.-A.; Choo, Y.-K. Relationship between ganglioside expression and anti-cancer effects of the monoclonal antibody against epithelial cell adhesion molecule in colon cancer. Exp. Mol. Med. 2011, 43, 693–701. [Google Scholar] [CrossRef]
- Yoshioka, K.; Ueno, Y.; Tanaka, S.; Nagai, K.; Onitake, T.; Hanaoka, R.; Watanabe, H.; Chayama, K. Role of natural killer T cells in the mouse colitis-associated colon cancer model. Scand. J. Immunol. 2012, 75, 16–26. [Google Scholar] [CrossRef]
- Chung, T.-W.; Choi, H.-J.; Kim, S.-J.; Kwak, C.-H.; Song, K.-H.; Jin, U.-H.; Chang, Y.-C.; Chang, H.W.; Lee, Y.-C.; Ha, K.-T.; et al. The ganglioside GM3 is associated with cisplatin-induced apoptosis in human colon cancer cells. PLoS ONE 2014, 9, e92786. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.G.M.; Solomon, M.J.; Mahon, K.; O’Shannassy, S. Management of colorectal cancer. BMJ 2019, 366, l4561. [Google Scholar] [CrossRef]
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; van de Velde, C.J.; Watanabe, T. Colorectal cancer. Nat. Rev. Dis. Primers 2015, 1, 15065. [Google Scholar] [CrossRef] [Green Version]
- Messersmith, W.A. NCCN Guidelines Updates: Management of Metastatic Colorectal Cancer. J. Natl. Compr. Cancer Netw. 2019, 17, 599–601. [Google Scholar]
- Van Cutsem, E.; Cervantes, A.; Nordlinger, B.; Arnold, D.; Group, E.G.W. Metastatic colorectal cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2014, 25 (Suppl. S3), iii1–iii9. [Google Scholar] [CrossRef]
- Sanchez-Gundin, J.; Fernandez-Carballido, A.M.; Martinez-Valdivieso, L.; Barreda-Hernandez, D.; Torres-Suarez, A.I. New Trends in the Therapeutic Approach to Metastatic Colorectal Cancer. Int. J. Med. Sci. 2018, 15, 659–665. [Google Scholar] [CrossRef] [Green Version]
- Van der Stok, E.P.; Spaander, M.C.W.; Grunhagen, D.J.; Verhoef, C.; Kuipers, E.J. Surveillance after curative treatment for colorectal cancer. Nat. Rev. Clin. Oncol. 2017, 14, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.H.; Chen, Y.X.; Fang, J.Y. Comprehensive review of targeted therapy for colorectal cancer. Signal. Transduct. Target. Ther. 2020, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Yaffee, P.; Osipov, A.; Tan, C.; Tuli, R.; Hendifar, A. Review of systemic therapies for locally advanced and metastatic rectal cancer. J. Gastrointest. Oncol. 2015, 6, 185–200. [Google Scholar] [PubMed]
- Steentoft, C.; Migliorini, D.; King, T.R.; Mandel, U.; June, C.H.; Posey, A.D., Jr. Glycan-directed CAR-T cells. Glycobiology 2018, 28, 656–669. [Google Scholar] [CrossRef]
- Dominguez-Villar, M.; Fernandez-Ponce, C.; Munoz-Suano, A.; Gomez, E.; Rodríguez-Iglesias, M.; Garcia-Cozar, F. Up-regulation of FOXP3 and induction of suppressive function in CD4+ Jurkat T-cells expressing hepatitis C virus core protein. Clin. Sci. 2012, 123, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Ponce, C.; Dominguez-Villar, M.; Aguado, E.; Garcia-Cozar, F. CD4+ primary T cells expressing HCV-core protein upregulate Foxp3 and IL-10, suppressing CD4 and CD8 T cells. PLoS ONE 2014, 9, e85191. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Ponce, C.; Dominguez-Villar, M.; Muñoz-Miranda, J.P.; Arbulo-Echevarria, M.M.; Litrán, R.; Aguado, E.; García-Cozar, F. Immune modulation by the hepatitis C virus core protein. J. Viral Hepat. 2017, 24, 350–356. [Google Scholar] [CrossRef]
- Fernández-Ponce, C.; Durán-Ruiz, M.C.; Narbona-Sánchez, I.; Muñoz-Miranda, J.P.; Arbulo-Echevarria, M.M.; Serna-Sanz, A.; Baumann, C.; Litrán, R.; Aguado, E.; Bloch, W.; et al. Ultrastructural Localization and Molecular Associations of HCV Capsid Protein in Jurkat T Cells. Front. Microbiol. 2018, 8, 2595. [Google Scholar] [CrossRef] [Green Version]
- Sasawatari, S.; Okamoto, Y.; Kumanogoh, A.; Toyofuku, T. Blockade of N-Glycosylation Promotes Antitumor Immune Response of T Cells. J. Immunol. 2020, 204, 1373–1385. [Google Scholar] [CrossRef]
- Steenackers, A.; Olivier-Van Stichelen, S.; Baldini, S.F.; Dehennaut, V.; Toillon, R.A.; Le Bourhis, X.; El Yazidi-Belkoura, I.; Lefebvre, T. Silencing the Nucleocytoplasmic O-GlcNAc Transferase Reduces Proliferation, Adhesion, and Migration of Cancer and Fetal Human Colon Cell Lines. Front. Endocrinol. 2016, 7, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Warburg, O. On respiratory impairment in cancer cells. Science 1956, 124, 269–270. [Google Scholar]
- Siegel, R.; Desantis, C.; Jemal, A. Colorectal cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 104–117. [Google Scholar] [CrossRef]
- Giovannucci, E.; Michaud, D. The role of obesity and related metabolic disturbances in cancers of the colon, prostate, and pancreas. Gastroenterology 2007, 132, 2208–2225. [Google Scholar] [CrossRef]
- Babae, N.; Bourajjaj, M.; Liu, Y.; Van Beijnum, J.R.; Cerisoli, F.; Scaria, P.V.; Verheul, M.; Van Berkel, M.P.; Pieters, E.H.; Van Haastert, R.J.; et al. Systemic miRNA-7 delivery inhibits tumor angiogenesis and growth in murine xenograft glioblastoma. Oncotarget 2014, 5, 6687–6700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, P.; He, T.; Jiang, R.; Li, G. MicroRNA-423-5p targets O-GlcNAc transferase to induce apoptosis in cardiomyocytes. Mol. Med. Rep. 2015, 12, 1163–1168. [Google Scholar] [CrossRef]
- Vaiana, C.A.; Kurcon, T.; Mahal, L.K. MicroRNA-424 Predicts a Role for β-1,4 Branched Glycosylation in Cell Cycle Progression. J. Biol. Chem. 2016, 291, 1529–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, M.; Xu, B.; Li, X.; Shang, Y.; Chu, Y.; Wang, W.; Chen, D.; Wu, N.; Hu, S.; Zhang, S.; et al. O-GlcNAcylation promotes colorectal cancer metastasis via the miR-101-O-GlcNAc/EZH2 regulatory feedback circuit. Oncogene 2019, 38, 301–316. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Zhang, B.; Nairn, A.V.; Nagy, T.; Moremen, K.W.; Buckhaults, P.; Pierce, M. O-Linked N-Acetylglucosamine (O-GlcNAc) Expression Levels Epigenetically Regulate Colon Cancer Tumorigenesis by Affecting the Cancer Stem Cell Compartment via Modulating Expression of Transcriptional Factor MYBL1. J. Biol. Chem. 2017, 292, 4123–4137. [Google Scholar] [CrossRef] [Green Version]
- Holm, M.; Nummela, P.; Heiskanen, A.; Satomaa, T.; Kaprio, T.; Mustonen, H.; Ristimäki, A.; Haglund, C. N-glycomic profiling of colorectal cancer according to tumor stage and location. PLoS ONE 2020, 15, e0234989. [Google Scholar] [CrossRef]
- Bennett, E.P.; Hassan, H.; Mandel, U.; Mirgorodskaya, E.; Roepstorff, P.; Burchell, J.; Taylor-Papadimitriou, J.; Hollingsworth, M.A.; Merkx, G.; van Kessel, A.G.; et al. Cloning of a Human UDP-N-Acetyl-α-d-Galactosamine:PolypeptideN-Acetylgalactosaminyltransferase That Complements Other GalNAc-Transferases in Complete O-Glycosylation of the MUC1 Tandem Repeat. J. Biol. Chem. 1998, 273, 30472–30481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Short, H.J.; Qian, K.X.; Elhammer, A.P.; Geng, J.G. Characterization of glycoprotein ligands for P-selectin on a human small cell lung cancer cell line NCI-H345. Biochem. Biophys. Res. Commun. 2001, 288, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Ramachandran, V.; Kang, J.; Kishimoto, T.K.; Cummings, R.D.; McEver, R.P. Identification of N-terminal residues on P-selectin glycoprotein ligand-1 required for binding to P-selectin. J. Biol. Chem. 1998, 273, 7078–7087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, J.J.; Qu, X.Y.; Zhou, D.Z. miR-4262 inhibits colon cancer cell proliferation via targeting of GALNT4. Mol. Med. Rep. 2017, 16, 3731–3736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamaguchi, J.; Nakagawa, H.; Takahashi, M.; Kudo, T.; Kamiyama, N.; Sun, B.; Oshima, T.; Sato, Y.; Deguchi, K.; Todo, S.; et al. Swainsonine reduces 5-fluorouracil tolerance in the multistage resistance of colorectal cancer cell lines. Mol. Cancer 2007, 6, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, F.M.; Latorre, A.O.; Hueza, I.M.; Sanches, D.S.; Lippi, L.L.; Gardner, D.R.; Spinosa, H.S. Increased antitumor efficacy by the combined administration of swainsonine and cisplatin in vivo. Phytomedicine 2011, 18, 1096–1101. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Ponce, C.; Geribaldi-Doldán, N.; Sánchez-Gomar, I.; Navarro Quiroz, R.; Atencio Ibarra, L.; Gomez Escorcia, L.; Fernández-Cisnal, R.; Aroca Martinez, G.; García-Cózar, F.; Navarro Quiroz, E. The Role of Glycosyltransferases in Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 5822. https://doi.org/10.3390/ijms22115822
Fernández-Ponce C, Geribaldi-Doldán N, Sánchez-Gomar I, Navarro Quiroz R, Atencio Ibarra L, Gomez Escorcia L, Fernández-Cisnal R, Aroca Martinez G, García-Cózar F, Navarro Quiroz E. The Role of Glycosyltransferases in Colorectal Cancer. International Journal of Molecular Sciences. 2021; 22(11):5822. https://doi.org/10.3390/ijms22115822
Chicago/Turabian StyleFernández-Ponce, Cecilia, Noelia Geribaldi-Doldán, Ismael Sánchez-Gomar, Roberto Navarro Quiroz, Linda Atencio Ibarra, Lorena Gomez Escorcia, Ricardo Fernández-Cisnal, Gustavo Aroca Martinez, Francisco García-Cózar, and Elkin Navarro Quiroz. 2021. "The Role of Glycosyltransferases in Colorectal Cancer" International Journal of Molecular Sciences 22, no. 11: 5822. https://doi.org/10.3390/ijms22115822