
Allogenic Use of Human Placenta-Derived Stromal Cells as a Highly Active Subtype of Mesenchymal Stromal Cells for Cell-Based Therapies


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Development of the Concept of Cell Therapy for Pro-Regenerative Treatments
Stem Cells: Sources and Proposed Roles in Tissue Regeneration
2. Therapies Based on Mesenchymal Stem/Stromal Cell (MSC) Injections
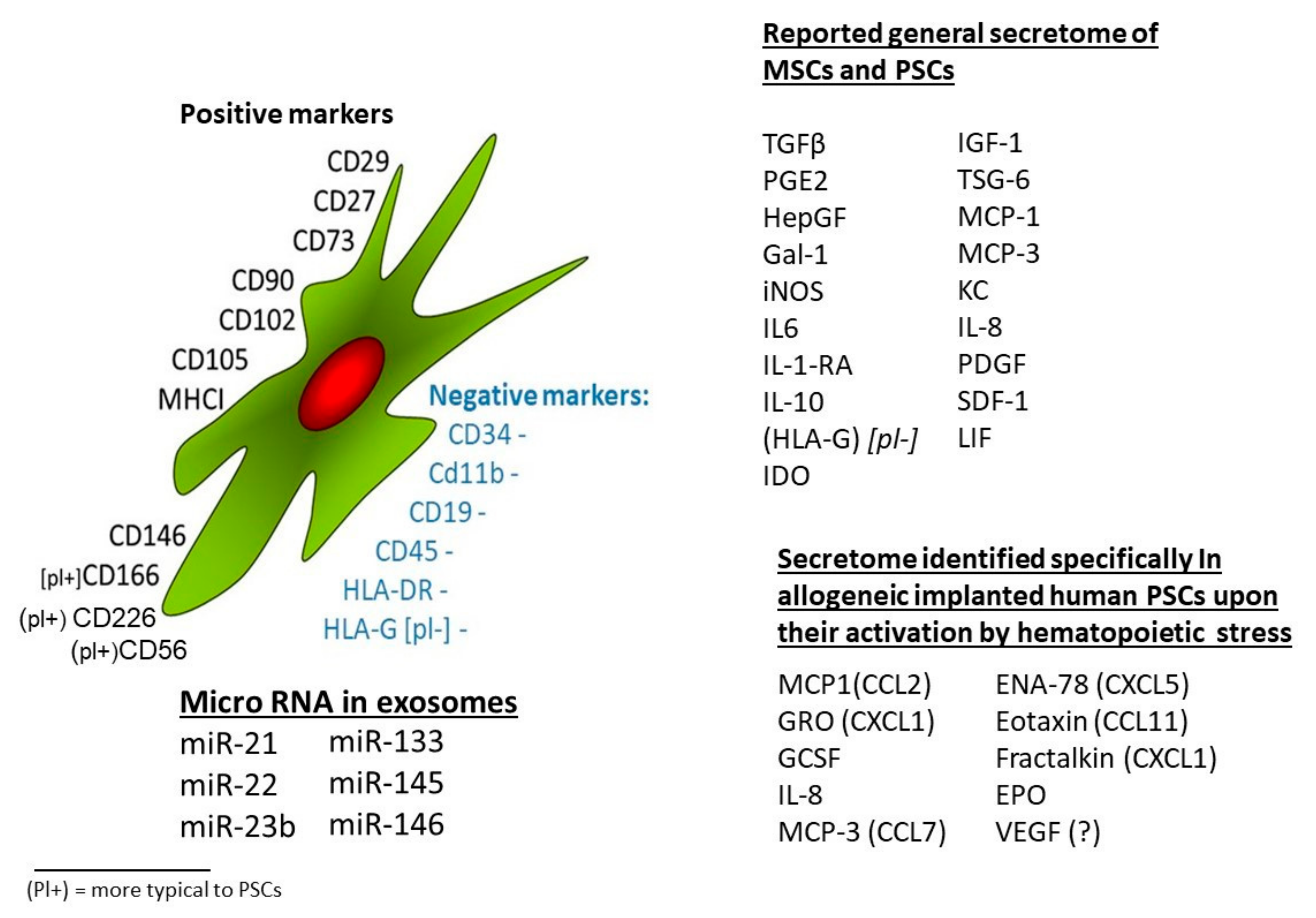
2.1. Properties and Phenotype of bmMSCs and MSCs Isolated from Other Tissues
2.2. Shared Features and Variations in the Phenotype of MSCs from Different Tissue Sources
2.3. Characterization of Different Tissues’ MSCs Based on Shared Surface Protein Markers
2.4. Characterization of MSCs from Different Sources by Their Secretion Profiles
2.5. MSCs for Modification of the Immune System Activity and Reduction of Inflammatory Responses
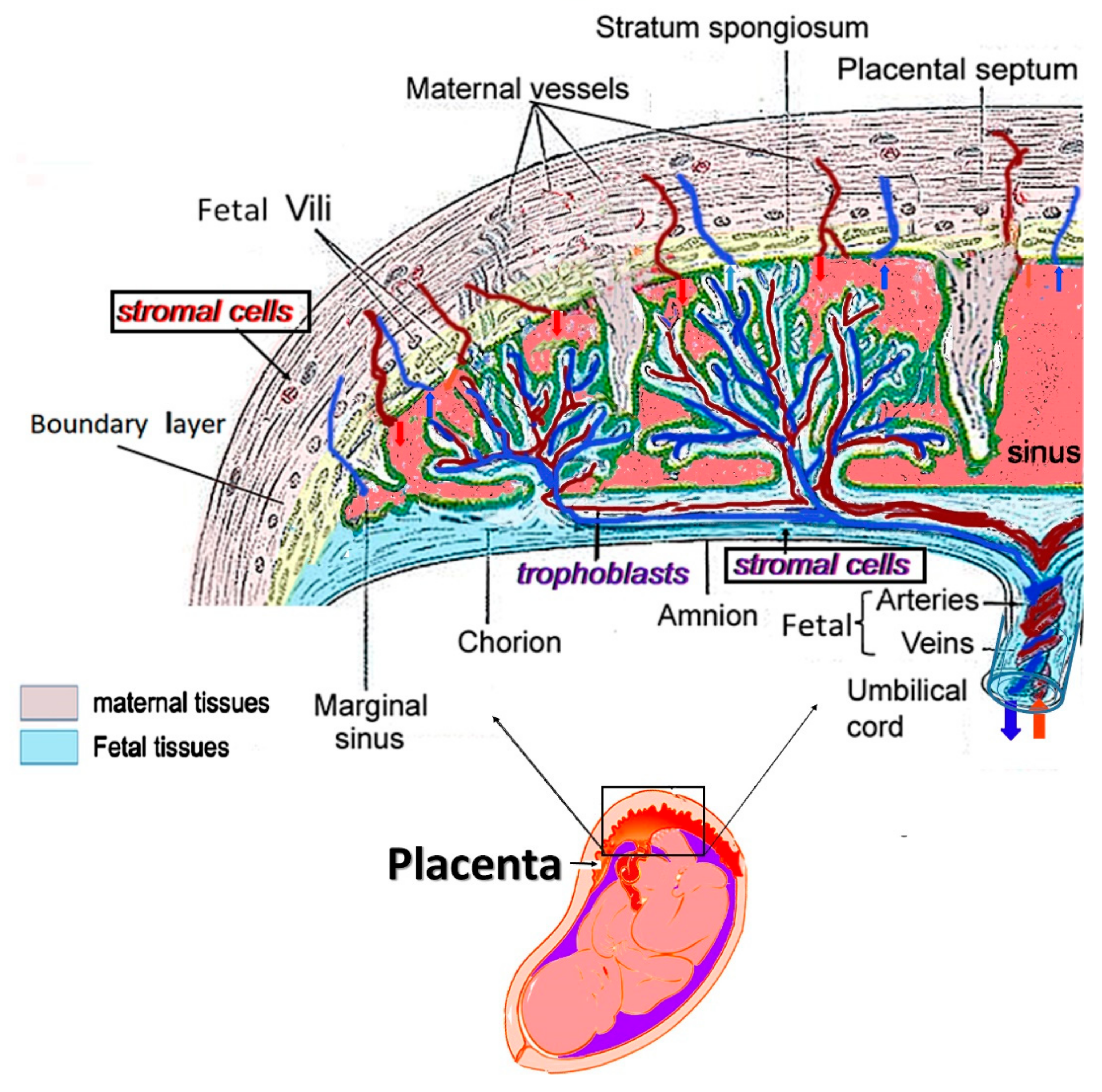
3. Application of hPSCs from Mature Placentae for Tissue Regeneration and Immune Modulation
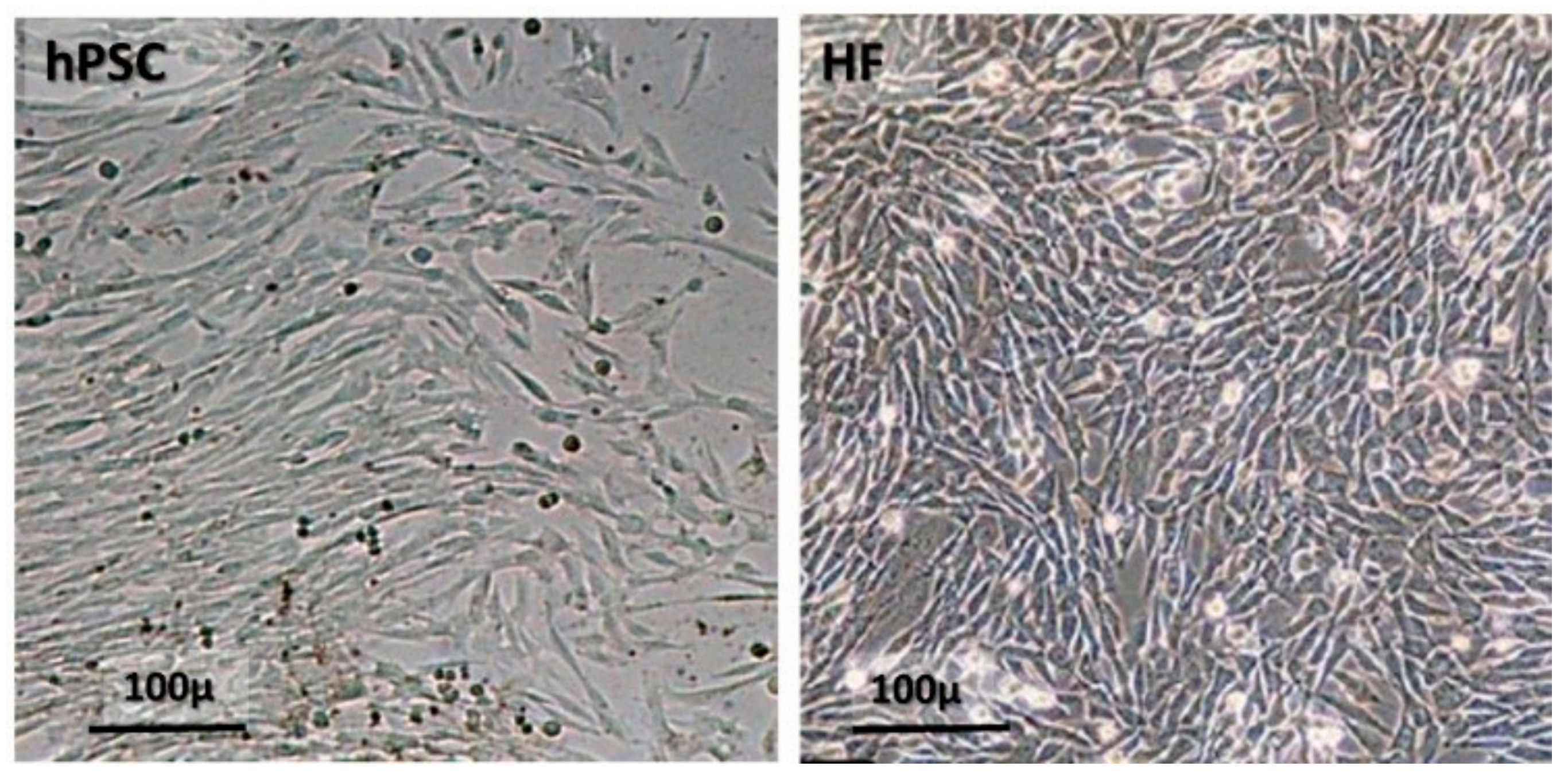
3.1. The Isolation of hPSCs from Full-Term Human Placenta and Their Expansion for Pre-Clinical and Clinical Studies
3.2. Suggested Modes of Action of hPSCs-Based Therapies
3.3. Pro-Regenerative Therapy with hPSCs from Commercial Sources
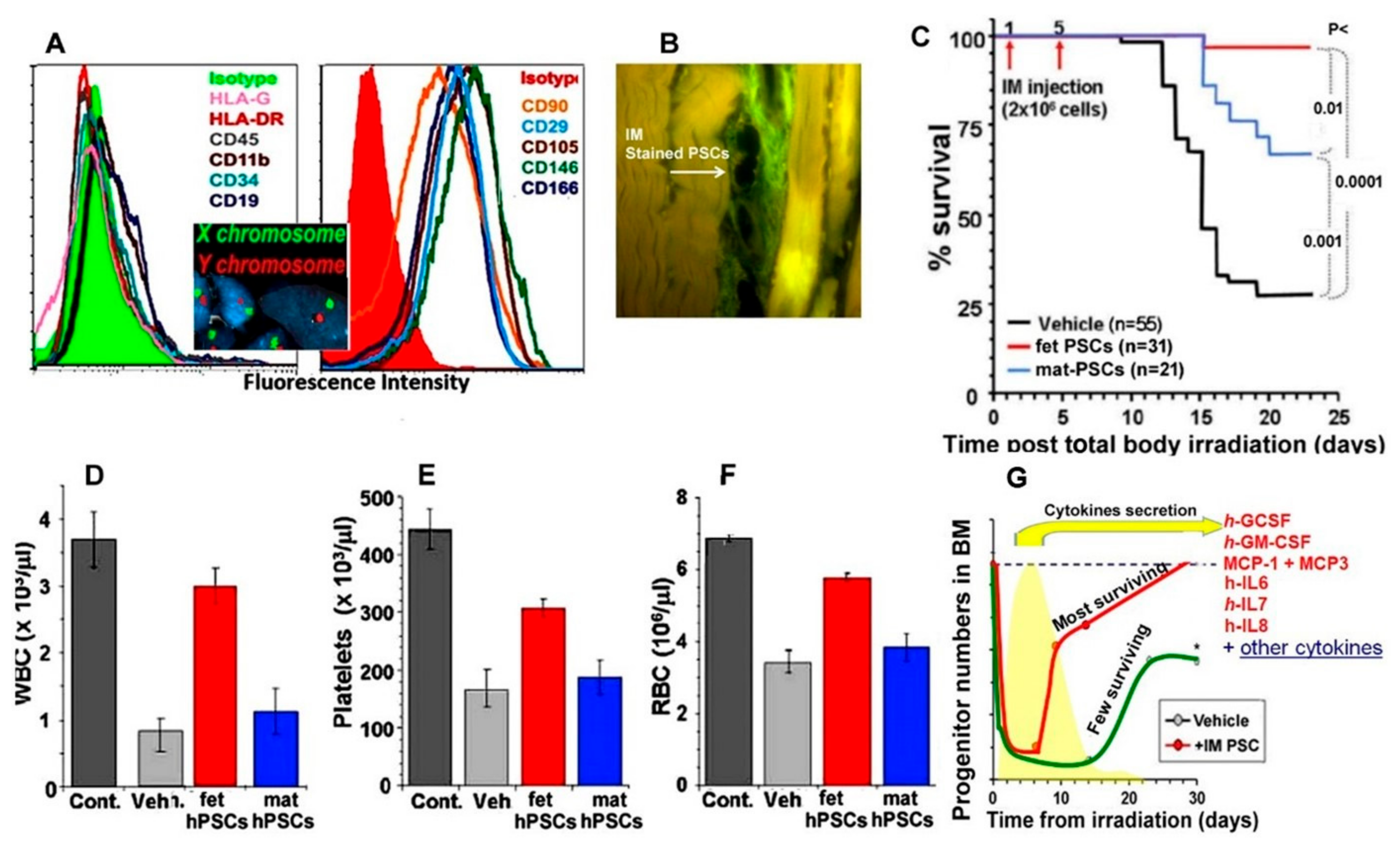
3.4. hPSCs Therapy as an Example of Their Use for Mitigation of Acute Radiation Syndrome (ARS)
3.5. The Anti-Inflammatory Effect of the hPSCs
4. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviation
| MSCs | mesenchymal stromal/stem cells |
| PSCs | placental stromal cells |
| f-hPSCs (or fet-hPSCs) | isolated and expanded human placental stromal cells, from fetal tissues |
| m-hPSCs or mat-hPSCs | isolated and expanded human placental stromal cells, from maternal tissues |
| HLA | human leukocyte antigen antibodies |
| BM | bone marrow |
| ES cells | multipotent embryonic stem cells |
| iPSCs | induced stem cells |
| IM, IP | intramuscular, intraperitoneal |
| atMSCs | adipose tissues MSCs |
| BBB | blood–brain barrier |
References
- Antunes, M.A.; Abreu, S.C.; Cruz, F.F.; Teixeira, A.C.; Lopes-Pacheco, M.; Bandeira, E.; Olsen, P.C.; Diaz, B.L.; Takyia, C.M.; Freitas, I.P.R.G.; et al. Effects of different mesenchymal stromal cell sources and delivery routes in experimental emphysema. Respir. Res. 2014, 15, 118. [Google Scholar] [CrossRef] [Green Version]
- Shufaro, Y.; Reubinoff, B.E. Therapeutic applications of embryonic stem cells. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 909–927. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, E.A.; Till, J.E. Effects of short-term culture on populations of hemopoietic progenitor cells from mouse marrow. Cell Tissue Kinet. 1971, 4, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Till, J.E. Proliferation and Differentiation of Stem Cells of the Blood-Forming System of the Mouse. NASA CR-673; NASA Contract Report, NASA CR; NASA: Washington, DC, USA, 1967; pp. 69–75.
- Guo, Y.; Lubbert, M.; Engelhardt, M. CD34-Hematopoietic Stem Cells: Current Concepts and Controversies. Stem Cells 2003, 21, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Verfaillie, C.M. Hematopoietic stem cells for transplantation. Nat. Immunol. 2002, 3, 314–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.H.; Pulin, A.A.; Seo, M.J.; Kota, D.J.; Ylostalo, J.; Larson, B.L.; Semprun-Prieto, L.; Delafontaine, P.; Prockop, D.J. Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6. Cell Stem Cell 2009, 5, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Azarpira, M.R.; Shahcheraghi, G.H.; Ayatollahi, M.; Geramizadeh, B. Tissue engineering strategy using mesenchymal stem cell-based chitosan scafolds in growth plate surgery: A preliminary study in rabbits. Orthop. Traumatol. Surg. Res. 2015, 101, 601–605. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ari, A.; Rivkin, R.; Frishman, M.; Gaberman, E.; Levdansky, L.; Gorodetsky, R. Isolation and implantation of bone marrow-derived mesenchymal stem cells with fibrin micro beads to repair a critical-size bone defect in mice. Tissue Eng. Part A 2009, 15, 2537–2546. [Google Scholar] [CrossRef]
- Bruder, S.P.; Kraus, K.H.; Goldberg, V.M.; Kadiyala, S. The effect of implants loaded with autologous mesenchymal stem cells on the healing of canine segmental bone defects. J. Bone Jt. Surg. Am. 1998, 80, 985–996. [Google Scholar] [CrossRef]
- Chen, L.; He, D.-M.; Zhang, Y. The differentiation of human placenta-derived mesenchymal stem cells into dopaminergic cells in vitro. Cell. Mol. Biol. Lett. 2009, 14, 528–536. [Google Scholar] [CrossRef]
- Fan, Z.X.; Lu, Y.; Deng, L.; Li, X.-Q.; Zhi, W.; Li-Ling, J.; Yang, Z.-M.; Xie, H.-Q. Placenta- versus bone-marrow-derived mesenchymal cells for the repair of segmental bone defects in a rabbit model. FEBS J. 2012, 279, 2455–2465. [Google Scholar] [CrossRef] [PubMed]
- Hirami, Y.; Osakada, F.; Takahashi, K.; Okita, K.; Yamanaka, S.; Ikeda, H.; Yoshimura, N.; Takahashi, M. Generation of retinal cells from mouse and human induced pluripotent stem cells. Neurosci. Lett. 2009, 458, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Holan, V.; Trosan, P.; Cejka, C.; Javorkova, E.; Zajicova, A.; Hermankova, B.; Chudickova, M.; Cejkova, J. A Comparative Study of the Therapeutic Potential of Mesenchymal Stem Cells and Limbal Epithelial Stem Cells for Ocular Surface Reconstruction. Stem Cells Transl. Med. 2015, 4, 1052–1063. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.Q.; Fox, R.; Chen, X.; Thibeault, S. Tissue regeneration of the vocal fold using bone marrow mesenchymal stem cells and synthetic extracellular matrix injections in rats. Laryngoscope 2010, 120, 537–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferro, F.; Falini, G.; Spelat, R.; D’Aurizio, F.; Puppato, E.; Pandolfi, M.; Beltrami, A.P.; Cesselli, D.; Beltrami, C.A.; Impiombato, F.S.A.; et al. Biochemical and biophysical analyses of tissue-engineered bone obtained from three-dimensional culture of a subset of bone marrow mesenchymal stem cells. Tissue Eng. Part A 2010, 16, 3657–3667. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.H.; Huang, T.-B.; Cheng, S.-J.; Weng, S.-Y.; Tsai, C.-L.; Tseng, C.-S.; Chen, D.C.; Liu, T.-Y.; Fu, K.-Y.; Yen, B.L. Chondrogenesis from human placenta-derived mesenchymal stem cells in three-dimensional scaffolds for cartilage tissue engineering. Tissue Eng. Part A 2011, 17, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Kittaka, M.; Kajiya, M.; Shiba, H.; Takewaki, M.; Takeshita, K.; Khung, R.; Fujita, T.; Iwata, T.; Nguyen, T.Q.; Ouhara, K.; et al. Clumps of a mesenchymal stromal cell/extracellular matrix complex can be a novel tissue engineering therapy for bone regeneration. Cytotherapy 2015, 17, 860–873. [Google Scholar] [CrossRef]
- Izal, I.; Aranda, P.; Sanz-Ramos, P.; Ripalda, P.; Mora, G.; Granero-Moltó, F.; Deplaine, H.; Gómez-Ribelles, J.L.; Ferrer, G.G.; Acosta, V.; et al. Culture of human bone marrow-derived mesenchymal stem cells on of poly(L-lactic acid) scaffolds: Potetial application for the tissue engineering of cartilage. Knee Surg. Sports Traumatol. Arthrosc. 2013, 21, 1737–1750. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, O.; Vexler, A.; Marx, G.; Prigozhina, T.; Levdansky, L.; Slavin, S.; Shimeliovich, I.; Gorodetsky, R. Fibrin microbeads for isolating and growing bone marrow-derived progenitor cells capable of forming bone tissue. Tissue Eng. 2002, 8, 661–672. [Google Scholar] [CrossRef]
- James, A.W.; Zhang, X.; Crisan, M.; Hardy, W.R.; Liang, P.; Meyers, C.A.; Lobo, S.; Lagishetty, V.; Childers, M.K.; Asatrian, G.; et al. Isolation and characterization of canine perivascular stem/stromal cells for bone tissue engineering. PLoS ONE 2017, 12, e0177308. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Xu, P.; Li, X.; Su, X.; Chen, Y.; Wan, L.; Fan, L.; Yin, K.; Liu, Y.; Zhao, R.C. Mesenchymal stem cells and cell therapy for bone repair. Curr. Mol. Pharmacol. 2016, 9, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shu, X.Z.; Prestwich, G.D. Osteochondral Defect Repair with Autologous Bone Marrow-Derived Mesenchymal Stem Cells in an Injectable, In Situ, Cross-Linked Synthetic Extracellular Matrix. Tissue Eng. 2006, 12, 3405–3416. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.N.; Law, J.B.K.; He, A.Y.; Low, H.Y.; Hui, J.H.; Lim, C.T.; Yang, Z.; Lee, E.H. Substrate topography determines the fate of chondrogenesis from human mesenchymal stem cells resulting in specific cartilage phenotype formation. Nanomedicine 2014, 10, 1507–1516. [Google Scholar] [CrossRef]
- Yeh, H.Y.; Lin, T.-Y.; Lin, C.-H.; Yen, B.L.; Tsai, C.-L.; Hsu, S.-H. Neocartilage formation from mesenchymal stem cells grown in type II collagen-hyaluronan composite scaffolds. Differentiation 2013, 86, 171–183. [Google Scholar] [CrossRef]
- Ahmed, T.A.; Hincke, M.T. Mesenchymal stem cell-based tissue engineering strategies for repair of articular cartilage. Histol. Histopathol. 2014, 29, 669–689. [Google Scholar]
- Kuznetsov, S.A.; Hailu-Lazmi, A.; Cherman, N.; De Castro, L.F.; Robey, P.G.; Gorodetsky, R. In Vivo Formation of Stable Hyaline Cartilage by Naive Human Bone Marrow Stromal Cells with Modified Fibrin Microbeads. Stem Cells Transl. Med. 2019, 8, 586–592. [Google Scholar] [CrossRef] [Green Version]
- Mardones, R.; Jofre, C.M.; Minguell, J.J. Cell Therapy and Tissue Engineering Approaches for Cartilage Repair and/or Regeneration. Int. J. Stem Cells 2015, 8, 48–53. [Google Scholar] [CrossRef]
- Bonvillain, R.W.; Danchuk, S.; Sullivan, D.E.; Betancourt, A.M.; Semon, J.A.; Eagle, M.E.; Mayeux, J.P.; Gregory, A.N.; Wang, G.; Townley, I.K.; et al. A nonhuman primate model of lung regeneration: Detergent-mediated decellularization and initial in vitro recellularization with mesenchymal stem cells. Tissue Eng. Part A 2012, 18, 2437–2452. [Google Scholar] [CrossRef] [Green Version]
- Gorodetsky, R. The use of fibrin based matrices and fibrin microbeads (FMB) for cell based tissue regeneration. Expert Opin. Biol. Ther. 2008, 8, 1831–1846. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.; Yang, X. Mesenchymal stem cells and bone regeneration: Current Status. Injury 2011, 42, 562–568. [Google Scholar] [CrossRef]
- Tsaryk, R.; Gloria, A.; Russo, T.; Anspach, L.; De Santis, R.; Ghanaati, S.; Unger, R.E.; Ambrosio, L.; Kirkpatrick, C.J. Collagen-low molecular weight hyaluronic acid semi-interpenetrating network loaded with gelatin microspheres for cell and growth factor delivery for nucleus pulposus regeneration. Acta Biomater. 2015, 20, 10–21. [Google Scholar] [CrossRef]
- Xia, H.; Li, X.; Gao, W.; Fu, X.; Fang, R.H.; Zhang, L.; Zhang, K. Tissue repair and regeneration with endogenous stem cells. Nat. Rev. Mater. 2018, 3, 174–193. [Google Scholar] [CrossRef]
- Ohnuki, M.; Takahashi, K.; Yamanaka, S. Generation and characterization of human induced pluripotent stem cells. Curr. Protoc. Stem Cell Biol. 2009, 9, 4A.2.1–4A.2.25. [Google Scholar] [CrossRef]
- Yoshida, Y.; Yamanaka, S. Recent stem cell advances: Induced pluripotent stem cells for disease modeling and stem cell-based regeneration. Circulation 2010, 122, 80–87. [Google Scholar] [CrossRef]
- Hentze, H.; Graichen, R.; Colman, A. Cell therapy and the safety of embryonic stem cell-derived grafts. Trends Biotechnol. 2007, 25, 24–32. [Google Scholar] [CrossRef]
- Shibata, H.; Ageyama, N.; Tanaka, Y.; Kishi, Y.; Sasaki, K.; Nakamura, S.; Muramatsu, S.-I.; Hayashi, S.; Kitano, Y.; Terao, K.; et al. Improved safety of hematopoietic transplantation with monkey embryonic stem cells in the allogeneic setting. Stem Cells 2006, 24, 1450–1457. [Google Scholar] [CrossRef]
- Chien, C.C.; Yen, B.L.; Lee, F.-K.; Lai, T.-H.; Chen, Y.-C.; Chan, S.-H.; Huang, H.-I. In vitro differentiation of human placenta-derived multipotent cells into hepatocyte-like cells. Stem Cells 2006, 24, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Nagao, Y.; Kitano, Y.; Hasegawa, H.; Shibata, H.; Takatoku, M.; Hayashi, S.; Ozawa, K.; Hanazono, Y. Hematopoietic microchimerism in sheep after in utero transplantation of cultured cynomolgus embryonic stem cells. Transplantation 2005, 79, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Idelson, M.; Alper, R.; Obolensky, A.; Ben-Shushan, E.; Hemo, I.; Yachimovich-Cohen, N.; Khaner, H.; Smith, Y.; Wiser, O.; Gropp, M.; et al. Directed differentiation of human embryonic stem cells into functional retinal pigment epithelium cells. Cell Stem Cell 2009, 5, 396–408. [Google Scholar] [CrossRef] [Green Version]
- Idelson, M.; Alper, R.; Obolensky, A.; Yachimovich-Cohen, N.; Rachmilewitz, J.; Ejzenberg, A.; Beider, E.; Banin, E.; Reubinoff, B. Immunological Properties of Human Embryonic Stem Cell-Derived Retinal Pigment Epithelial Cells. Stem Cell Rep. 2018, 11, 681–695. [Google Scholar] [CrossRef] [Green Version]
- Vailly, J.; Gagnoux-Palacios, L.; Dell’Ambra, E.; Roméro, C.; Pinola, M.; Zambruno, G.; De Luca, M.; Ortonne, J.P.; Meneguzzi, G. Corrective gene transfer of keratinocytes from patients with junctional epidermolysis bullosa restores assembly of hemidesmosomes in reconstructed epithelia. Gene Ther. 1998, 5, 1322–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latella, M.C.; Cocchiarella, F.; De Rosa, L.; Turchiano, G.; Goncalves, M.; Larcher, F.; De Luca, M.; Recchia, A. Correction of Recessive Dystrophic Epidermolysis Bullosa by Transposon-Mediated Integration of COL7A1 in Transplantable Patient-Derived Primary Keratinocytes. J. Investig. Dermatol. 2017, 137, 836–844. [Google Scholar] [CrossRef] [Green Version]
- Colter, D.C.; Sekiya, I.; Prockop, D.J. Identification of a subpopulation of rapidly self-renewing and multipotential adult stem cells in colonies of human marrow stromal cells. Proc. Natl. Acad. Sci. USA 2001, 98, 7841–7845. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Gong, S.P.; Kim, B.; Kwon, H.S.; Yang, W.S.; Jeong, J.-W.; Ahn, J.; Lim, J.M. The co-injection of somatic cells with embryonic stem cells affects teratoma formation and the properties of teratoma-derived stem cell-like cells. PLoS ONE 2014, 9, e105975. [Google Scholar] [CrossRef]
- Gropp, M.; Shilo, V.; Vainer, G.; Gov, M.; Gil, Y.; Khaner, H.; Matzrafi, L.; Idelson, M.; Kopolovic, J.; Zak, N.B.; et al. Standardization of the teratoma assay for analysis of pluripotency of human ES cells and biosafety of their differentiated progeny. PLoS ONE 2012, 7, e45532. [Google Scholar] [CrossRef]
- Bandekar, M.; Maurya, D.K.; Sharma, D.; Checker, R.; Gota, V.; Mishra, N.; Sandur, S.K. Xenogeneic transplantation of human WJ-MSCs rescues mice from acute radiation syndrome via Nrf-2-dependent regeneration of damaged tissues. Am. J. Transplant. 2020, 20, 2044–2057. [Google Scholar] [CrossRef]
- Berardis, S.; Sattwika, P.D.; Najimi, M.; Sokal, E.M. Use of mesenchymal stem cells to treat liver fibrosis: Current situation and future prospects. World J. Gastroenterol. 2015, 21, 742–758. [Google Scholar] [CrossRef]
- Chen, H.; Min, X.H.; Wang, Q.Y.; Leung, F.W.; Shi, L.; Zhou, Y.; Yu, T.; Wang, C.M.; An, G.; Sha, W.H.; et al. Pre-activation of mesenchymal stem cells with TNF-alpha, IL-1beta and nitric oxide enhances its paracrine effects on radiation-induced intestinal injury. Sci. Rep. 2015, 5, 8718. [Google Scholar] [CrossRef] [Green Version]
- Gore, A.V.; Bible, L.E.; Livingston, D.H.; Mohr, A.M.; Sifri, Z.C. Mesenchymal stem cells enhance lung recovery after injury, shock, and chronic stress. Surgery 2016, 159, 1430–1435. [Google Scholar] [CrossRef] [Green Version]
- Hess, D.C.; Borlongan, C.V. Stem cells and neurological diseases. Cell Prolif. 2008, 41 (Suppl. 1), 94–114. [Google Scholar] [CrossRef] [PubMed]
- Newman, R.E.; Yoo, D.; Leroux, M.A.; Danilkovitch-Miagkova, A. Treatment of inflammatory diseases with mesenchymal stem cells. Inflamm. Allergy Drug Targets 2009, 8, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Stabler, C.T.; Lecht, S.; Lazarovici, P.; Lelkes, P.I. Mesenchymal stem cells for therapeutic applications in pulmonary medicine. Br. Med. Bull. 2015, 115, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Zaher, W.; Harkness, L.; Jafari, A.; Kassem, M. An update of human mesenchymal stem cell biology and their clinical uses. Arch. Toxicol. 2014, 88, 1069–1082. [Google Scholar] [CrossRef]
- Griffin, T.P.; Martin, W.P.; Islam, N.; O’Brien, T.; Griffin, M. The Promise of Mesenchymal Stem Cell Therapy for Diabetic Kidney Disease. Curr. Diabetes Rep. 2016, 16, 42. [Google Scholar] [CrossRef]
- Ogata, K.; Katagiri, W.; Osugi, M.; Kawai, T.; Sugimura, Y.; Hibi, H.; Nakamura, S.; Ueda, M. Evaluation of the therapeutic effects of conditioned media from mesenchymal stem cells in a rat bisphosphonate-related osteonecrosis of the jaw-like model. Bone 2015, 74, 95–105. [Google Scholar] [CrossRef]
- Amiri, F.; Jahanian-Najafabadi, A.; Roudkenar, M.H. In vitro augmentation of mesenchymal stem cells viability in stressful microenvironments: In vitro augmentation of mesenchymal stem cells viability. Cell Stress Chaperones 2015, 20, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Galipeau, J.; Krampera, M.; Barrett, J.; Dazzi, F.; Deans, R.J.; DeBruijn, J.; Dominici, M.; Fibbe, W.E.; Gee, A.P.; Gimble, J.M.; et al. International Society for Cellular Therapy perspective on immune functional assays for mesenchymal stromal cells as potency release criterion for advanced phase clinical trials. Cytotherapy 2016, 18, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Attar-Schneider, O.; Zismanov, V.; Drucker, L.; Gottfried, M. Secretome of human bone marrow mesenchymal stem cells: An emerging player in lung cancer progression and mechanisms of translation initiation. Tumor Biol. 2015, 37, 4755–4765. [Google Scholar] [CrossRef]
- Kawai, T.; Katagiri, W.; Osugi, M.; Sugimura, Y.; Hibi, H.; Ueda, M. Secretomes from bone marrow-derived mesenchymal stromal cells enhance periodontal tissue regeneration. Cytotherapy 2015, 17, 369–381. [Google Scholar] [CrossRef]
- Sacchetti, B.; Funari, A.; Michienzi, S.; Di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.G.; Riminucci, M.; et al. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianco, P.; Robey, P.G.; Saggio, I.; Riminucci, M. “Mesenchymal” stem cells in human bone marrow (skeletal stem cells): A critical discussion of their nature, identity, and significance in incurable skeletal disease. Hum. Gene Ther. 2010, 21, 1057–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Rivkin, R.; Ben-Ari, A.; Kassis, I.; Zangi, L.; Gaberman, E.; Levdansky, L.; Marx, G.; Gorodetsky, R. High-yield isolation, expansion, and differentiation of murine bone marrow-derived mesenchymal stem cells using fibrin microbeads (FMB). Cloning Stem Cells 2007, 9, 157–175. [Google Scholar] [CrossRef]
- Zangi, L.; Rivkin, R.; Kassis, I.; Levdansky, L.; Marx, G.; Gorodetsky, R. High-yield isolation, expansion, and differentiation of rat bone marrow-derived mesenchymal stem cells with fibrin microbeads. Tissue Eng. 2006, 12, 2343–2354. [Google Scholar] [CrossRef]
- Martire, A.; Bedada, F.B.; Uchida, S.; Pöling, J.; Krüger, M.; Warnecke, H.; Richter, M.; Kubin, T.; Herold, S.; Braun, T. Mesenchymal stem cells attenuate inflammatory processes in the heart and lung via inhibition of TNF signaling. Basic Res. Cardiol. 2016, 111, 54. [Google Scholar] [CrossRef] [Green Version]
- Caplan, A.I.; Dennis, J.E. Mesenchymal stem cells as trophic mediators. J. Cell Biochem. 2006, 98, 1076–1084. [Google Scholar] [CrossRef]
- Caplan, A.I.; Correa, D. The MSC: An injury drugstore. Cell Stem Cell 2011, 9, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Reinders, M.E.; De Fijter, J.W.; Roelofs, H.; Bajema, I.M.; De Vries, D.K.; Schaapherder, A.F.; Claas, F.H.; Van Miert, P.P.; Roelen, D.L.; Van Kooten, C.; et al. Autologous bone marrow-derived mesenchymal stromal cells for the treatment of allograft rejection after renal transplantation: Results of a phase I study. Stem Cells Transl. Med. 2013, 2, 107–111. [Google Scholar] [CrossRef]
- Fu, L.; Liu, Y.; Zhang, D.; Xie, J.; Guan, H.; Shang, T. Beneficial effect of human umbilical cord-derived mesenchymal stem cells on an endotoxin-induced rat model of preeclampsia. Exp. Ther. Med. 2015, 10, 1851–1856. [Google Scholar] [CrossRef]
- Rahimzadeh, A.; Mirakabad, F.S.T.; Movassaghpour, A.; Shamsasenjan, K.; Kariminekoo, S.; Talebi, M.; Shekari, A.; Zeighamian, V.; Ghalhar, M.G.; Akbarzadeh, A. Biotechnological and biomedical applications of mesenchymal stem cells as a therapeutic system. Artif. Cells Nanomed. Biotechnol. 2014, 44, 1–12. [Google Scholar]
- Li, D.; Pan, X.; Zhao, J.; Chi, C.; Wu, G.; Wang, Y.; Liao, S.; Wang, C.; Ma, J.; Pan, J. Bone Marrow Mesenchymal Stem Cells Suppress Acute Lung Injury Induced by Lipopolysaccharide Through Inhibiting the TLR2, 4/NF-kappaB Pathway in Rats with Multiple Trauma. Shock 2016, 45, 641–646. [Google Scholar] [CrossRef]
- Squillaro, T.; Peluso, G.; Galderisi, U. Clinical Trials with Mesenchymal Stem Cells: An Update. Cell Transplant. 2016, 25, 829–848. [Google Scholar] [CrossRef] [Green Version]
- Ottoboni, L.; De Feo, D.; Merlini, A.; Martino, G. Commonalities in immune modulation between mesenchymal stem cells (MSCs) and neural stem/precursor cells (NPCs). Immunol. Lett. 2015, 168, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Laranjeira, P.; Pedrosa, M.; Pedreiro, S.; Gomes, J.; Martinho, A.; Antunes, B.; Ribeiro, T.; Santos, F.; Trindade, H.; Paiva, A. Effect of human bone marrow mesenchymal stromal cells on cytokine production by peripheral blood naive, memory, and effector T cells. Stem Cell Res. Ther. 2015, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Hinden, L.; Shainer, R.; Almogi-Hazan, O.; Or, R. Ex Vivo Induced Regulatory Human/Murine Mesenchymal Stem Cells as Immune Modulators. Stem Cells 2015, 33, 2256–2267. [Google Scholar] [CrossRef]
- Castro-Manrreza, M.E.; Mayani, H.; Monroy-García, A.; Flores-Figueroa, E.; Chávez-Rueda, K.; Legorreta-Haquet, V.; Santiago-Osorio, E.; Montesinos, J.J. Human mesenchymal stromal cells from adult and neonatal sources: A comparative in vitro analysis of their immunosuppressive properties against T cells. Stem Cells Dev. 2014, 23, 1217–1232. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, D.; Liu, X.; Tang, S.; Wei, F. Human umbilical cord mesenchymal stem cells reduce systemic inflammation and attenuate LPS-induced acute lung injury in rats. J. Inflamm. 2012, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- Al Jumah, M.A.; Abumaree, M.H. The Immunomodulatory and Neuroprotective Effects of Mesenchymal Stem Cells (MSCs) in Experimental Autoimmune Encephalomyelitis (EAE): A Model of Multiple Sclerosis (MS). Int. J. Mol. Sci. 2012, 13, 9298–92331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvinen, L.; Badri, L.; Wettlaufer, S.; Ohtsuka, T.; Standiford, T.J.; Toews, G.B.; Pinsky, D.J.; Peters-Golden, M.; Lama, V.N. Lung resident mesenchymal stem cells isolated from human lung allografts inhibit T cell proliferation via a soluble mediator. J. Immunol. 2008, 181, 4389–4396. [Google Scholar] [CrossRef] [Green Version]
- Hafez, P.; Jose, S.; Chowdhury, S.R.; Ng, M.H.; Ruszymah, B.H.I.; Mohd, R.A.R. Cardiomyogenic differentiation of human sternal bone marrow mesenchymal stem cells using a combination of basic fibroblast growth factor and hydrocortisone. Cell Biol. Int. 2016, 40, 55–64. [Google Scholar] [CrossRef]
- Sabapathy, V.; Sundaram, B.; Sreelakshmi, V.M.; Mankuzhy, P.; Kumar, S. Human Wharton’s Jelly Mesenchymal Stem Cells plasticity augments scar-free skin wound healing with hair growth. PLoS ONE 2014, 9, e93726. [Google Scholar] [CrossRef] [PubMed]
- Cartarozzi, L.P.; Spejo, A.B.; Ferreira, R.S.; Barraviera, B.; Duek, E.; Carvalho, J.L.; Góes, A.M.; Oliveira, A.L. Mesenchymal stem cells engrafted in a fibrin scaffold stimulate Schwann cell reactivity and axonal regeneration following sciatic nerve tubulization. Brain Res. Bull. 2015, 112, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Pelled, G.; Turgeman, G.; Aslan, H.; Gazit, Z.; Gazit, D. Mesenchymal stem cells for bone gene therapy and tissue engineering. Curr. Pharm. Des. 2002, 8, 1917–1928. [Google Scholar] [CrossRef]
- Manzini, B.M.; Duarte, A.D.S.S.; Sankaramanivel, S.; Ramos, A.L.; Latuf-Filho, P.; Escanhoela, C.; Kharmandayan, P.; Saad, S.T.O.; Boin, I.; Luzo, Â.C.M. Useful properties of undifferentiated mesenchymal stromal cells and adipose tissue as the source in liver-regenerative therapy studied in an animal model of severe acute fulminant hepatitis. Cytotherapy 2015, 17, 1052–1065. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Eleuteri, S.; Fierabracci, A. Insights into the Secretome of Mesenchymal Stem Cells and Its Potential Applications. Int. J. Mol. Sci 2019, 20, 4597. [Google Scholar] [CrossRef] [Green Version]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast precursors in normal and irradiated mouse hematopoietic organs. Exp. Hematol. 1976, 4, 267–274. [Google Scholar]
- Caplan, A.I.; Bruder, S.P. Mesenchymal stem cells: Building blocks for molecular medicine in the 21st century. Trends Mol. Med. 2001, 7, 259–264. [Google Scholar] [CrossRef]
- Popov, B.V.; Serikov, V.B.; Petrov, N.S.; Izusova, T.V.; Gupta, N.; Matthay, M.A. Lung epithelial cells induce endodermal differentiation in mouse mesenchymal bone marrow stem cells by paracrine mechanism. Tissue Eng. 2007, 13, 2441–2450. [Google Scholar] [CrossRef]
- Tremolada, C.; Ricordi, C.; Caplan, A.I.; Ventura, C. Mesenchymal Stem Cells in Lipogems, a Reverse Story: From Clinical Practice to Basic Science. Methods Mol. Biol. 2016, 1416, 109–122. [Google Scholar] [PubMed]
- Parekkadan, B.; Tilles, A.W.; Yarmush, M.L. Bone marrow-derived mesenchymal stem cells ameliorate autoimmune enteropathy independently of regulatory T cells. Stem Cells 2008, 26, 1913–1919. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorodetsky, R.; McBride, W.H.; Withers, H.R.; Miller, G.G. Effect of fibroblast implants on wound healing of irradiated skin: Assay of wound strength and quantitative immunohistology of collagen. Radiat. Res. 1991, 125, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.W.; Gorodetsky, R.; Micevicz, E.D.; Mackenzie, N.C.; Gaberman, E.; Levdansky, L.; McBride, W.H. Marrow-derived stromal cell delivery on fibrin microbeads can correct radiation-induced wound-healing deficits. J. Invest. Dermatol. 2013, 133, 553–561. [Google Scholar] [CrossRef] [Green Version]
- Krueger, W.W.; Goepfert, H.; Romsdahl, M.; Herson, J.; Withers, R.H.; Jesse, R.H. Fibroblast implantation enhances wound healing as indicated by breaking strength determinations. Otolaryngology 1978, 86, ORL-804–811. [Google Scholar] [CrossRef]
- Hocking, A.M. The Role of Chemokines in Mesenchymal Stem Cell Homing to Wounds. Adv. Wound Care 2015, 4, 623–630. [Google Scholar] [CrossRef]
- Bank, J.R.; Rabelink, T.J.; De Fijter, J.W.; Reinders, M.E.J. Safety and Efficacy Endpoints for Mesenchymal Stromal Cell Therapy in Renal Transplant Recipients. J. Immunol. Res. 2015, 2015, 391797. [Google Scholar] [CrossRef]
- Nagaishi, K.; Arimura, Y.; Fujimiya, M. Stem cell therapy for inflammatory bowel disease. J. Gastroenterol. 2015, 50, 280–286. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, P.; Aghayan, H.R.; Larijani, B.; Soleimani, M.; Dehpour, A.-R.; Sahebjam, M.; Ghaderi, F.; Arjmand, B. Stem cell-based approach for the treatment of Parkinson’s disease. Med. J. Islam. Repub. Iran 2015, 29, 168. [Google Scholar]
- Hashmi, S.; Ahmed, M.; Murad, M.H.; Litzow, M.R.; Adams, R.H.; Ball, L.M.; Prasad, V.K.; Kebriaei, P.; Ringden, O. Survival after mesenchymal stromal cell therapy in steroid-refractory acute graft-versus-host disease: Systematic review and meta-analysis. Lancet Haematol. 2016, 3, e45–e52. [Google Scholar] [CrossRef]
- Kim, G.; Eom, Y.W.; Baik, S.K.; Shin, Y.; Lim, Y.L.; Kim, M.Y.; Kwon, S.O.; Chang, S.J. Therapeutic Effects of Mesenchymal Stem Cells for Patients with Chronic Liver Diseases: Systematic Review and Meta-analysis. J. Korean Med. Sci. 2015, 30, 1405–1415. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-Y.; Li, X.-Z.; Wang, L.-B. Therapeutic implications of mesenchymal stem cells in acute lung injury/acute respiratory distress syndrome. Stem Cell Res. Ther. 2013, 4, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.R.; Tang, Y.-L.; Xuan, M.; Chang, Z.; Wang, X.-Y.; Liang, X.-H. Transplantation of autologous mesenchymal stem cells for end-stage liver cirrhosis: A meta-analysis based on seven controlled trials. Gastroenterol. Res. Pract. 2015, 2015, 908275. [Google Scholar] [CrossRef] [Green Version]
- Zuk, A.P.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Zannettino, A.C.W.; Paton, S.; Arthur, A.; Khor, F.; Itescu, S.; Gimble, J.; Gronthos, S. Multipotential human adipose-derived stromal stem cells exhibit a perivascular phenotype in vitro and in vivo. J. Cell. Physiol. 2007, 214, 413–421. [Google Scholar] [CrossRef]
- Lee, O.K.; Kuo, T.K.; Chen, W.-M.; Lee, K.-D.; Hsieh, S.-L.; Chen, T.-H. Isolation of multipotent mesenchymal stem cells from umbilical cord blood. Blood 2004, 103, 1669–1675. [Google Scholar] [CrossRef] [Green Version]
- Romanov, Y.A.; Svintsitskaya, V.A.; Smirnov, V.N. Searching for Alternative Sources of Postnatal Human Mesenchymal Stem Cells: Candidate MSC-Like Cells from Umbilical Cord. Stem Cells 2003, 21, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Melo, F.R.; Bressan, R.B.; Forner, S.; Martini, A.C.; Rode, M.; Delben, P.B.; Rae, G.A.; Figueiredo, C.P.; Trentin, A.G. Transplantation of Human Skin-Derived Mesenchymal Stromal Cells Improves Locomotor Recovery After Spinal Cord Injury in Rats. Cell. Mol. Neurobiol. 2017, 37, 941–947. [Google Scholar] [CrossRef]
- Da Silva Meirelles, L.; Chagastelles, C.; Nardi, N.B. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J. Cell Sci. 2006, 119 Pt 11, 2204–2213. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Sha, X.-J.; Li, G.-H.; Yang, F.-S.; Ji, K.; Wen, L.-Y.; Liu, S.-Y.; Chen, L.; Ding, Y.; Xuan, K. Comparative characterization of stem cells from human exfoliated deciduous teeth and dental pulp stem cells. Arch. Oral Biol. 2012, 57, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Wang, Q.; Li, Y.; Wei, H.; Shi, J.; Li, A. A novel method for banking stem cells from human exfoliated deciduous teeth: Lentiviral TERT immortalization and phenotypical analysis. Stem Cell Res. Ther. 2016, 7, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinzur, L.; Akyuez, L.; Levdansky, L.; Blumenfeld, M.; Volinsky, E.; Aberman, Z.; Reinke, P.; Ofir, R.; Volk, H.-D.; Gorodetsky, R. Rescue from lethal acute radiation syndrome (ARS) with severe weight loss by secretome of intramuscularly injected human placental stromal cells. J. Cachexia Sarcopenia Muscle 2018, 9, 1079–1092. [Google Scholar] [CrossRef] [PubMed]
- Abumaree, M.H.; Al Jumah, M.A.; Kalionis, B.; Jawdat, D.; Al Khaldi, A.; Altalabani, A.A.; Knawy, B.A. Phenotypic and functional characterization of mesenchymal stem cells from chorionic villi of human term placenta. Stem Cell Rev. Rep. 2013, 9, 16–31. [Google Scholar] [CrossRef]
- Adani, B.; Basheer, M.; Hailu, A.L.; Fogel, T.; Israeli, E.; Volinsky, E.; Gorodetsky, R. Isolation and expansion of high yield of pure mesenchymal stromal cells from fresh and cryopreserved placental tissues. Cryobiology 2019, 89, 100–103. [Google Scholar] [CrossRef]
- Ilic, N.; Atkinson, K. Manufacturing and use of human placenta-derived mesenchymal stromal cells for phase I clinical trials: Establishment and evaluation of a protocol. Vojnosanit. Pregl. 2014, 71, 651–659. [Google Scholar] [CrossRef] [Green Version]
- In ‘t Anker, P.S.; Scherjon, S.A.; Der Keur, C.K.-V.; De Groot-Swings, G.M.; Claas, F.H.; Fibbe, W.E.; Kanhai, H.H. Isolation of mesenchymal stem cells of fetal or maternal origin from human placenta. Stem Cells 2004, 22, 1338–1345. [Google Scholar] [CrossRef]
- Kanematsu, D.; Shofuda, T.; Yamamoto, A.; Ban, C.; Ueda, T.; Yamasaki, M.; Kanemura, Y. Isolation and cellular properties of mesenchymal cells derived from the decidua of human term placenta. Differentiation 2011, 82, 77–88. [Google Scholar] [CrossRef]
- Miao, Z.; Jin, J.; Chen, L.; Zhu, J.; Huang, W.; Zhao, J.; Qian, H.; Zhang, X. Isolation of mesenchymal stem cells from human placenta: Comparison with human bone marrow mesenchymal stem cells. Cell Biol. Int. 2006, 30, 681–687. [Google Scholar] [CrossRef]
- Patel, J.; Shafiee, A.; Wang, W.; Fisk, N.M.; Khosrotehrani, K. Novel isolation strategy to deliver pure fetal-origin and maternal-origin mesenchymal stem cell (MSC) populations from human term placenta. Placenta 2014, 35, 969–971. [Google Scholar] [CrossRef]
- Rus Ciuca, D.; SoriŢău, O.; Suşman, S.; I Pop, V.; Mihu, C.M. Isolation and characterization of chorionic mesenchyal stem cells from the placenta. Rom. J. Morphol. Embryol. 2011, 52, 803–808. [Google Scholar]
- Parolini, O.; Alviano, F.; Bergwerf, I.; Boraschi, D.; De Bari, C.; De Waele, P.; Dominici, M.; Evangelista, M.; Falk, W.; Hennerbichler, S.; et al. Toward Cell Therapy Using Placenta-Derived Cells: Disease Mechanisms, Cell Biology, Preclinical Studies, and Regulatory Aspects at the Round Table. Stem Cells Dev. 2010, 19, 143–154. [Google Scholar] [CrossRef]
- Fukuchi, Y.; Nakajima, H.; Sugiyama, D.; Hirose, I.; Kitamura, T.; Tsuji, K. Human Placenta-Derived Cells Have Mesenchymal Stem/Progenitor Cell Potential. Stem Cells 2004, 22, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Abumaree, M.H.; Abomaray, F.M.; AlShehri, N.A.; Almutairi, A.; Alaskar, A.S.; Kalionis, B.; Al Jumah, M.A. Phenotypic and Functional Characterization of Mesenchymal Stem/Multipotent Stromal Cells From Decidua Parietalis of Human Term Placenta. Reprod. Sci. 2016, 23, 1193–1207. [Google Scholar] [CrossRef]
- Talwadekar, M.D.; Kale, V.P.; Limaye, L.S. Placenta-derived mesenchymal stem cells possess better immunoregulatory properties compared to their cord-derived counterparts-a paired sample study. Sci. Rep. 2015, 5, 15784. [Google Scholar] [CrossRef] [Green Version]
- Vogel, W.; Grünebach, F.; A Messam, C.; Kanz, L.; Brugger, W.; Bühring, H.-J. Heterogeneity among human bone marrow-derived mesenchymal stem cells and neural progenitor cells. Haematologica 2003, 88, 126–133. [Google Scholar]
- Buhring, H.J.; Battula, V.L.; Treml, S.; Schewe, B.; Kanz, L.; Vogel, W. Novel markers for the prospective isolation of human MSC. Ann. N. Y. Acad. Sci. 2007, 1106, 262–271. [Google Scholar] [CrossRef]
- Alvarez-Viejo, M.; Menendez-Menendez, Y.; Otero-Hernandez, J. CD271 as a marker to identify mesenchymal stem cells from diverse sources before culture. World J. Stem Cells 2015, 7, 470–476. [Google Scholar] [CrossRef]
- Turco, M.Y.; Gardner, L.; Kay, R.G.; Hamilton, R.S.; Prater, M.; Hollinshead, M.S.; McWhinnie, A.; Esposito, L.; Fernando, R.; Skelton, H.; et al. Trophoblast organoids as a model for maternal-fetal interactions during human placentation. Nature 2018, 564, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Sivasubramaniyan, K.; Harichandan, A.; Schumann, S.; Sobiesiak, M.; Lengerke, C.; Maurer, A.; Kalbacher, H.; Bühring, H.J. Prospective isolation of mesenchymal stem cells from human bone marrow using novel antibodies directed against Sushi domain containing 2. Stem Cells Dev. 2013, 22, 1944–1954. [Google Scholar] [CrossRef]
- Abruzzese, T.; Aicher, W.K. Factors contained in the expansion medium regulate the expression stem cell markers CD146 and SUSD2 on human placenta-derived mesenchymal stromal cells and modulate their differentiation capacity in vitro. FASEB J. 2019, 33, 495.1. [Google Scholar]
- Maleki, M.; Ghanbarvand, F.; Behvarz, M.R.; Ejtemaei, M.; Ghadirkhomi, E. Comparison of mesenchymal stem cell markers in multiple human adult stem cells. Int. J. Stem Cells 2014, 7, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohart, F.; Mason, E.; Matigian, N.; Mosbergen, R.; Korn, O.; Chen, T.; Butcher, S.; Patel, J.; Atkinson, K.; Khosrotehrani, K.; et al. A molecular classification of human mesenchymal stromal cells. PeerJ 2016, 4, e1845. [Google Scholar] [CrossRef] [PubMed]
- Roson-Burgo, B.; Sanchez-Guijo, F.; Del Cañizo, C.; Rivas, J.D.L. Insights into the human mesenchymal stromal/stem cell identity through integrative transcriptomic profiling. BMC Genom. 2016, 17, 944. [Google Scholar] [CrossRef] [Green Version]
- Verpoorten, A.; Abruzzese, T.A.; Pils, E.; Abele, H.; Hart, M.L.; Aicher, W.K. Expression of CD146 on Human Placenta-Derived Mesenchymal Stromal Cells and their Osteogenic Differentiation Capacity are Modulated by Factors Contained in Platelet Lysate. J. Regen. Med. 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Matigian, N.; Brooke, G.; Zaibak, F.; Rossetti, T.; Kollar, K.; Pelekanos, R.; Heazlewood, C.; Mackay-Sim, A.; Wells, C.A.; Atkinson, K. Multipotent human stromal cells isolated from cord blood, term placenta and adult bone marrow show distinct differences in gene expression pattern. Genom. Data 2015, 3, 70–74. [Google Scholar] [CrossRef]
- Pasquinelli, G.; Tazzari, P.; Ricci, F.; Vaselli, C.; Buzzi, M.; Conte, R.; Orrico, C.; Foroni, L.; Stella, A.; Alviano, F.; et al. Ultrastructural characteristics of human mesenchymal stromal (stem) cells derived from bone marrow and term placenta. Ultrastruct. Pathol. 2007, 31, 23–31. [Google Scholar] [CrossRef]
- Roson-Burgo, B.; Sanchez-Guijo, F.; Del Cañizo, C.; Rivas, J.D.L. Transcriptomic portrait of human Mesenchymal Stromal/Stem cells isolated from bone marrow and placenta. BMC Genom. 2014, 15, 910. [Google Scholar] [CrossRef] [Green Version]
- Clabaut, A.; Grare, C.; Léger, T.; Hardouin, P.; Broux, O. Variations of secretome profiles according to conditioned medium preparation: The example of human mesenchymal stem cell-derived adipocytes. Electrophoresis 2015, 36, 2587–2593. [Google Scholar] [CrossRef]
- Dong, L.; Wang, Y.; Zheng, T.; Pu, Y.; Ma, Y.; Qi, X.; Zhang, W.; Xue, F.; Shan, Z.; Liu, J.; et al. Hypoxic hUCMSC-derived extracellular vesicles attenuate allergic airway inflammation and airway remodeling in chronic asthma mice. Stem Cell Res. Ther. 2021, 12, 4. [Google Scholar] [CrossRef]
- Mao, C.; Li, D.; Zhou, E.; Gao, E.; Zhang, T.; Sun, S.; Gao, L.; Fan, Y.; Wang, C. Extracellular vesicles from anoxia preconditioned mesenchymal stem cells alleviate myocardial ischemia/reperfusion injury. Aging 2021, 13, 6156–6170. [Google Scholar] [CrossRef]
- Harrell, C.R.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Mesenchymal Stem Cell-Derived Exosomes and Other Extracellular Vesicles as New Remedies in the Therapy of Inflammatory Diseases. Cells 2019, 8, 1605. [Google Scholar] [CrossRef] [Green Version]
- Damania, A.; Jaiman, D.; Teotia, A.K.; Kumar, A. Mesenchymal stromal cell-derived exosome-rich fractionated secretome confers a hepatoprotective effect in liver injury. Stem Cell Res. Ther. 2018, 9, 31. [Google Scholar] [CrossRef] [Green Version]
- Billing, A.M.; Ben Hamidane, H.; Dib, S.S.; Cotton, R.J.; Bhagwat, A.M.; Kumar, P.; Hayat, S.; Yousri, N.A.; Goswami, N.; Suhre, K.; et al. Comprehensive transcriptomic and proteomic characterization of human mesenchymal stem cells reveals source specific cellular markers. Sci. Rep. 2016, 6, 21507. [Google Scholar] [CrossRef] [Green Version]
- Spitzhorn, L.-S.; Megges, M.; Wruck, W.; Rahman, M.S.; Otte, J.; Degistirici, Ö.; Meisel, R.; Sorg, R.V.; Oreffo, R.O.C.; Adjaye, J. Human iPSC-derived MSCs (iMSCs) from aged individuals acquire a rejuvenation signature. Stem Cell Res. Ther. 2019, 10, 100. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.; Lee, J.; Kwon, Y.; Park, K.-S.; Jeong, J.-H.; Choi, S.-J.; Bang, S.I.; Chang, J.W.; Lee, C. Comparative Proteomic Analysis of the Mesenchymal Stem Cells Secretome from Adipose, Bone Marrow, Placenta and Wharton’s Jelly. Int. J. Mol. Sci. 2021, 22, 845. [Google Scholar] [CrossRef]
- Ulrich, C.; Rolauffs, B.; Abele, H.; Bonin, M.; Nieselt, K.; Hart, M.L.; Aicher, W.K. Low osteogenic differentiation potential of placenta-derived mesenchymal stromal cells correlates with low expression of the transcription factors Runx2 and Twist2. Stem Cells Dev. 2013, 22, 2859–2872. [Google Scholar] [CrossRef] [Green Version]
- Maerz, J.K.; Roncoroni, L.P.; Goldeck, D.; Abruzzese, T.; Kalbacher, H.; Rolauffs, B.; Dezwart, P.; Nieselt, K.; Hart, M.L.; Klein, G.; et al. Bone marrow-derived mesenchymal stromal cells differ in their attachment to fibronectin-derived peptides from term placenta-derived mesenchymal stromal cells. Stem Cell Res. Ther. 2016, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Muraglia, A.; Cancedda, R.; Quarto, R. Clonal mesenchymal progenitors from human bone marrow differentiate in vitro according to a hierarchical model. J. Cell Sci. 2000, 113 Pt 7, 1161–1166. [Google Scholar] [CrossRef]
- Robinson, A.M.; Miller, S.; Payne, N.; Boyd, R.; Sakkal, S.; Nurgali, K. Neuroprotective Potential of Mesenchymal Stem Cell-Based Therapy in Acute Stages of TNBS-Induced Colitis in Guinea-Pigs. PLoS ONE 2015, 10, e0139023. [Google Scholar] [CrossRef]
- Piscioneri, A.; Morelli, S.; Mele, M.; Canonaco, M.; Bilotta, E.; Pantano, P.; Drioli, E.; De Bartolo, L. Neuroprotective effect of human mesenchymal stem cells in a compartmentalized neuronal membrane system. Acta Biomater. 2015, 24, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.S.H.; Mei, S.H.J.; Stewart, D.J. The Immunomodulatory and Therapeutic Effects of Mesenchymal Stromal Cells for Acute Lung Injury and Sepsis. J. Cell. Physiol. 2015, 230, 2606–2617. [Google Scholar] [CrossRef] [PubMed]
- Hendijani, F.; Javanmard, S.H.; Rafiee, L.; Sadeghi-Aliabadi, H. Effect of human Wharton’s jelly mesenchymal stem cell secretome on proliferation, apoptosis and drug resistance of lung cancer cells. Res. Pharm. Sci. 2015, 10, 134–142. [Google Scholar]
- Succar, P.; Breen, E.J.; Kuah, D.; Herbert, B.R. Alterations in the Secretome of Clinically Relevant Preparations of Adipose-Derived Mesenchymal Stem Cells Cocultured with Hyaluronan. Stem Cells Int. 2015, 2015, 421253. [Google Scholar] [CrossRef] [Green Version]
- Phinney, D.G.; Pittenger, M.F. Concise Review: MSC-Derived Exosomes for Cell-Free Therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rani, S.; Ryan, A.E.; Griffin, M.D.; Ritter, T. Mesenchymal Stem Cell-derived Extracellular Vesicles: Toward Cell-free Therapeutic Applications. Mol. Ther. 2015, 23, 812–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, S.; Deregibus, M.C.; Camussi, G. The secretome of mesenchymal stromal cells: Role of extracellular vesicles in immunomodulation. Immunol. Lett. 2015, 168, 154–158. [Google Scholar] [CrossRef]
- Saraswati, S.; Guo, Y.; Atkinson, J.; Young, P.P. Prolonged hypoxia induces monocarboxylate transporter-4 expression in mesenchymal stem cells resulting in a secretome that is deleterious to cardiovascular repair. Stem Cells 2015, 33, 1333–1344. [Google Scholar] [CrossRef] [Green Version]
- Curley, G.F.; Hayes, M.; Ansari, B.; Shaw, G.; Ryan, A.; Barry, F.; O’Brien, T.; O’Toole, D.; Laffey, J.G. Mesenchymal stem cells enhance recovery and repair following ventilator-induced lung injury in the rat. Thorax 2012, 67, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Stahl, P.D.; Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles, Integrators of Homeostasis. Physiology 2019, 34, 169–177. [Google Scholar] [CrossRef]
- Maacha, S.; Sidahmed, H.; Jacob, S.; Gentilcore, G.; Calzone, R.; Grivel, J.-C.; Cugno, C. Paracrine Mechanisms of Mesenchymal Stromal Cells in Angiogenesis. Stem Cells Int. 2020, 2020, 4356359. [Google Scholar] [CrossRef]
- Ringden, O.; Uzunel, M.; Rasmusson, I.; Remberger, M.; Sundberg, B.; Lönnies, H.; Marschall, H.-U.; Dlugosz, A.; Szakos, A.; Hassan, Z.; et al. Mesenchymal stem cells for treatment of therapy-resistant graft-versus-host disease. Transplantation 2006, 81, 1390–1397. [Google Scholar] [CrossRef]
- Sun, B.; Yu, K.-R.; Bhandari, D.R.; Jung, J.-W.; Kang, S.-K.; Kang, K.-S. Human umbilical cord blood mesenchymal stem cell-derived extracellular matrix prohibits metastatic cancer cell MDA-MB-231 proliferation. Cancer Lett. 2010, 296, 178–185. [Google Scholar] [CrossRef]
- Sun, L.; Wang, D.; Liang, J.; Zhang, H.; Feng, X.; Wang, H.; Hua, B.; Liu, B.; Ye, S.; Hu, X.; et al. Umbilical cord mesenchymal stem cell transplantation in severe and refractory systemic lupus erythematosus. Arthritis Rheum. 2010, 62, 2467–2475. [Google Scholar] [CrossRef]
- Tyndall, A.; Houssiau, F.A. Mesenchymal stem cells in the treatment of autoimmune diseases. Ann. Rheum. Dis. 2010, 69, 1413–1414. [Google Scholar] [CrossRef] [Green Version]
- Machado, C.D.V.; Telles, P.D.; Nascimento, I.L. Immunological characteristics of mesenchymal stem cells. Rev. Bras. Hematol. Hemoter. 2013, 35, 62–67. [Google Scholar] [CrossRef]
- Opitz, C.A.; Litzenburger, U.M.; Lutz, C.; Lanz, T.V.; Tritschler, I.; Köppel, A.; Tolosa, E.; Hoberg, M.; Anderl, J.; Aicher, W.K.; et al. Toll-Like Receptor Engagement Enhances the Immunosuppressive Properties of Human Bone Marrow-Derived Mesenchymal Stem Cells by Inducing Indoleamine-2,3-dioxygenase-1 via Interferon-β and Protein Kinase R. Stem Cells 2009, 27, 909–919. [Google Scholar] [CrossRef]
- Krueger, T.E.; Thorek, D.L.J.; Meeker, A.K.; Isaacs, J.T.; Brennen, W.N. Tumor-infiltrating mesenchymal stem cells: Drivers of the immunosuppressive tumor microenvironment in prostate cancer? Prostate 2019, 79, 320–330. [Google Scholar] [CrossRef]
- Jiang, D.; Chen, F.-X.; Zhou, H.; Lu, Y.-Y.; Tan, H.; Yu, S.-J.; Yuan, J.; Liu, H.; Meng, W.; Jin, Z.-B. Bioenergetic Crosstalk between Mesenchymal Stem Cells and various Ocular Cells through the intercellular trafficking of Mitochondria. Theranostics 2020, 10, 7260–7272. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.M.; Nakhle, J.; Griessinger, E.; Vignais, M.-L. Intercellular mitochondria trafficking highlighting the dual role of mesenchymal stem cells as both sensors and rescuers of tissue injury. Cell Cycle 2018, 17, 712–721. [Google Scholar] [CrossRef]
- Jorgensen, C.; Khoury, M. Musculoskeletal Progenitor/Stromal Cell-Derived Mitochondria Modulate Cell Differentiation and Therapeutical Function. Front. Immunol. 2021, 12, 606781. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, C.-D.; Jiang, X.-X.; Li, H.-L.; Tang, P.-H.; Mao, N. Comparison of mesenchymal stem cells from human placenta and bone marrow. Chin. Med. J. 2004, 117, 882–887. [Google Scholar] [PubMed]
- Aengenheister, L.; Favaro, R.R.; Morales-Prieto, D.M.; Furer, L.A.; Gruber, M.; Wadsack, C.; Markert, U.R.; Buerki-Thurnherr, T. Research on nanoparticles in human perfused placenta: State of the art and perspectives. Placenta 2021, 104, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Haghighi Poodeh, S.; Salonurmi, T.; Nagy, I.; Koivunen, P.; Vuoristo, J.; Räsänen, J.; Sormunen, R.; Vainio, S.; Savolainen, M. Alcohol-induced premature permeability in mouse placenta-yolk sac barriers in vivo. Placenta 2012, 33, 866–873. [Google Scholar] [CrossRef]
- Merz, G.; Schwenk, V.; Shah, R.; Salafia, C.; Necaise, P.; Joyce, M.; Villani, T.; Johnson, M.; Crider, N. Three-dimensional Rendering and Analysis of Immunolabeled, Clarified Human Placental Villous Vascular Networks. J. Vis. Exp. 2018, 133, e57099. [Google Scholar] [CrossRef]
- Rath, G.; Dhuria, R.; Salhan, S.; Jain, A.K. Morphology and morphometric analysis of stromal capillaries in full term human placental villi of smoking mothers: An electron microscopic study. Clin. Ter. 2011, 162, 301–305. [Google Scholar]
- Wagner, G.P.; Kin, K.; Muglia, L.; Pavlicev, M. Evolution of mammalian pregnancy and the origin of the decidual stromal cell. Int. J. Dev. Biol. 2014, 58, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Zigic, Z.; Marković, S.; Grbesa, D.; Ramić, S.; Halilović, A. Quantitative research of capillaries in terminal villi of mature placentae. Bosn. J. Basic Med. Sci. 2010, 10, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Saulnier, N.; Loriau, J.; Febre, M.; Robert, C.; Rakic, R.; Bonte, T.; Buff, S.; Maddens, S. Canine placenta: A promising potential source of highly proliferative and immunomodulatory mesenchymal stromal cells? Vet. Immunol. Immunopathol. 2016, 171, 47–55. [Google Scholar] [CrossRef]
- Li, G.; Zhang, X.-A.; Wang, H.; Wang, X.; Meng, C.-L.; Chan, C.-Y.; Yew, D.T.W.; Tsang, K.S.; Li, K.; Tsai, S.-N.; et al. Comparative proteomic analysis of mesenchymal stem cells derived from human bone marrow, umbilical cord and placenta: Implication in the migration. Adv. Exp. Med. Biol. 2011, 720, 51–68. [Google Scholar]
- Jaramillo-Ferrada, P.A.; Wolvetang, E.J.; Cooper-White, J.J. Differential mesengenic potential and expression of stem cell-fate modulators in mesenchymal stromal cells from human-term placenta and bone marrow. J. Cell. Physiol. 2012, 227, 3234–3242. [Google Scholar] [CrossRef] [PubMed]
- Mohr, S.; Portmann-Lanz, C.B.; Schoeberlein, A.; Sager, R.; Surbek, D.V. Generation of an osteogenic graft from human placenta and placenta-derived mesenchymal stem cells. Reprod. Sci. 2010, 17, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Parolini, O.; Soncini, M. Human placenta: A source for progenitor/stem cells? J. Reproduktionsmed. Endocrinol 2006, 3, 117–126. [Google Scholar]
- Koo, B.K.; Park, I.Y.; Kim, J.; Kim, J.-H.; Kwon, A.; Kim, M.; Kim, Y.; Shin, J.C.; Kim, J.-H. Isolation and characterization of chorionic mesenchymal stromal cells from human full term placenta. J. Korean Med. Sci. 2012, 27, 857–863. [Google Scholar] [CrossRef] [Green Version]
- Brooke, G.; Rossetti, T.; Pelekanos, R.; Ilic, N.; Murray, P.; Hancock, S.; Antonenas, V.; Huang, G.; Gottlieb, D.; Bradstock, K.; et al. Manufacturing of human placenta-derived mesenchymal stem cells for clinical trials. Br. J. Haematol. 2009, 144, 571–579. [Google Scholar] [CrossRef]
- Silini, A.R.; Di Pietro, R.; Lang-Olip, I.; Alviano, F.; Banerjee, A.; Basile, M.; Borutinskaite, V.; Eissner, G.; Gellhaus, A.; Giebel, B.; et al. Perinatal Derivatives: Where Do We Stand? A Roadmap of the Human Placenta and Consensus for Tissue and Cell Nomenclature. Front. Bioeng. Biotechnol. 2020, 8, 610544. [Google Scholar] [CrossRef]
- Ulrich, C.; Abruzzese, T.; Maerz, J.K.; Ruh, M.; Amend, B.; Benz, K.; Rolauffs, B.; Abele, H.; Hart, M.L.; Aicher, W.K. Human Placenta-Derived CD146-Positive Mesenchymal Stromal Cells Display a Distinct Osteogenic Differentiation Potential. Stem Cells Dev. 2015, 24, 1558–1569. [Google Scholar] [CrossRef]
- Pipino, C.; Shangaris, P.; Resca, E.; Zia, S.; Deprest, J.; Sebire, N.J.; David, A.L.; Guillot, P.V.; De Coppi, P. Placenta as a reservoir of stem cells: An underutilized resource? Br. Med. Bull. 2012, 105, 43–68. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.Z.; Ji, H. In vitro differentiation of osteocytes and adipocytes from human placenta-derived cells. J. Int. Med. Res. 2012, 40, 761–767. [Google Scholar] [CrossRef] [Green Version]
- Kadam, S.; Govindasamy, V.; Bhonde, R. Generation of functional islets from human umbilical cord and placenta derived mesenchymal stem cells. Methods Mol. Biol. 2012, 879, 291–313. [Google Scholar]
- Semenov, O.V.; Koestenbauer, S.; Riegel, M.; Zech, N.; Zimmermann, R.; Zisch, A.H.; Malek, A. Multipotent mesenchymal stem cells from human placenta: Critical parameters for isolation and maintenance of stemness after isolation. Am. J. Obstet. Gynecol. 2010, 202, 193.e1–193.e13. [Google Scholar] [CrossRef] [Green Version]
- Portmann-Lanz, C.B.; Schoeberlein, A.; Portmann, R.; Mohr, S.; Rollini, P.; Sager, R.; Surbek, D.V. Turning placenta into brain: Placental mesenchymal stem cells differentiate into neurons and oligodendrocytes. Am. J. Obstet. Gynecol. 2010, 202, 294.e1–294.e11. [Google Scholar] [CrossRef]
- Brooke, G.; Tong, H.; Lévesque, J.-P.; Atkinson, K. Molecular trafficking mechanisms of multipotent mesenchymal stem cells derived from human bone marrow and placenta. Stem Cells Dev. 2008, 17, 929–940. [Google Scholar] [CrossRef]
- Barlow, S.; Brooke, G.; Chatterjee, K.; Price, G.; Pelekanos, R.; Rossetti, T.; Doody, M.; Venter, D.; Pain, S.; Gilshenan, K.; et al. Comparison of human placenta- and bone marrow-derived multipotent mesenchymal stem cells. Stem Cells Dev. 2008, 17, 1095–1107. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Mitsuru, A.; Igura, K.; Takahashi, K.; Ichinose, S.; Yamaguchi, S.; Takahashi, T.A. Mesenchymal progenitor cells derived from chorionic villi of human placenta for cartilage tissue engineering. Biochem. Biophys. Res. Commun. 2006, 340, 944–952. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, Z.; Huang, Y.; Wang, P.; Luo, Y.; Gao, Y.; Du, Z. The subtype CD200-positive, chorionic mesenchymal stem cells from the placenta promote regeneration of human hepatocytes. Biotechnol. Lett. 2014, 36, 1335–1341. [Google Scholar] [CrossRef]
- Passipieri, J.A.; Kasai-Brunswick, T.H.; Suhett, G.; Martins, A.B.; Brasil, G.V.; Campos, D.B.; Rocha, N.N.; Ramos, I.P.; Mello, D.B.; Rodrigues, D.C.; et al. Improvement of cardiac function by placenta-derived mesenchymal stem cells does not require permanent engraftment and is independent of the insulin signaling pathway. Stem Cell Res. Ther. 2014, 5, 102. [Google Scholar] [CrossRef] [Green Version]
- Manochantr, S.; U-Pratya, Y.; Kheolamai, P.; Rojphisan, S.; Chayosumrit, M.; Tantrawatpan, C.; Supokawej, A.; Issaragrisil, S. Immunosuppressive properties of mesenchymal stromal cells derived from amnion, placenta, Wharton’s jelly and umbilical cord. Intern. Med. J. 2013, 43, 430–439. [Google Scholar] [CrossRef]
- Kong, P.; Xie, X.; Li, F.; Liu, Y.; Lu, Y. Placenta mesenchymal stem cell accelerates wound healing by enhancing angiogenesis in diabetic Goto-Kakizaki (GK) rats. Biochem. Biophys. Res. Commun. 2013, 438, 410–419. [Google Scholar] [CrossRef]
- Lee, M.J.; Jung, J.; Na, K.-H.; Moon, J.S.; Lee, H.-J.; Kim, J.-H.; Kim, G.I.; Kwon, S.-W.; Hwang, S.-G. Anti-fibrotic effect of chorionic plate-derived mesenchymal stem cells isolated from human placenta in a rat model of CCl(4)-injured liver: Potential application to the treatment of hepatic diseases. J. Cell. Biochem. 2010, 111, 1453–1463. [Google Scholar] [CrossRef]
- Prather, W.R.; Toren, A.; Meiron, M.; Ofir, R.; Tschope, C.; Horwitz, E.M. The role of placental-derived adherent stromal cell (PLX-PAD) in the treatment of critical limb ischemia. Cytotherapy 2009, 11, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Shim, S.S.; Seok, O.S.; Lee, H.Y.; Woo, S.K.; Kim, B.H.; Song, H.R.; Lee, J.K.; Park, Y.K. Comparison of cytokine expression in mesenchymal stem cells from human placenta, cord blood, and bone marrow. J. Korean Med. Sci. 2009, 24, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Alviano, F.; Fossati, V.; Marchionni, C.; Arpinati, M.; Bonsi, L.; Franchina, M.; Lanzoni, G.; Cantoni, S.; Cavallini, C.; Bianchi, F.; et al. Term Amniotic membrane is a high throughput source for multipotent Mesenchymal Stem Cells with the ability to differentiate into endothelial cells in vitro. BMC Dev. Biol. 2007, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, S.; Meinhardt, G.; Saleh, L.; Kunihs, V.; Gamperl, M.; Kaindl, U.; Ellinger, A.; Burkard, T.R.; Fiala, C.; Pollheimer, J.; et al. Self-Renewing Trophoblast Organoids Recapitulate the Developmental Program of the Early Human Placenta. Stem Cell Rep. 2018, 11, 537–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaberman, E.; Pinzur, L.; Levdansky, L.; Tsirlin, M.; Netzer, N.; Aberman, Z.; Gorodetsky, R. Mitigation of Lethal Radiation Syndrome in Mice by Intramuscular Injection of 3D Cultured Adherent Human Placental Stromal Cells. PLoS ONE 2013, 8, e66549. [Google Scholar] [CrossRef]
- Chatterjee, P.; Chiasson, V.L.; Pinzur, L.; Raveh, S.; Abraham, E.; Jones, K.A.; Bounds, K.R.; Ofir, R.; Flaishon, L.; Chajut, A.; et al. Human placenta-derived stromal cells decrease inflammation, placental injury and blood pressure in hypertensive pregnant mice. Clin. Sci. 2016, 130, 513–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Yang, Y.; Zhang, Y.; Hao, G.; Liu, T.; Wang, L.; Yang, T.; Wang, Q.; Zhang, G.; Wei, J.; et al. Placental mesenchymal stem cells of fetal and maternal origins demonstrate different therapeutic potentials. Stem Cell Res. Ther. 2014, 5, 48. [Google Scholar] [CrossRef] [Green Version]
- Adams, K.M.; Yan, Z.; Stevens, A.M.; Nelson, J.L. The changing maternal “self” hypothesis: A mechanism for maternal tolerance of the fetus. Placenta 2007, 28, 378–382. [Google Scholar] [CrossRef]
- Eide, I.P.; Isaksen, C.V.; Salvesen, K.Å.; Langaas, M.; Günther, C.-C.; Iversen, A.-C.; Austgulen, R. Fetal growth restriction is associated with reduced FasL expression by decidual cells. J. Reprod. Immunol. 2007, 74, 7–14. [Google Scholar] [CrossRef]
- Gregori, S.; Amodio, G.; Quattrone, F.; Panina-Bordignon, P. HLA-G Orchestrates the Early Interaction of Human Trophoblasts with the Maternal Niche. Front. Immunol. 2015, 6, 128. [Google Scholar] [CrossRef] [Green Version]
- Jauniaux, E.; Jurkovic, D.; Gulbis, B.; Liesnard, C.; Lees, C.; Campbell, S. Materno-fetal immunoglobulin transfer and passive immunity during the first trimester of human pregnancy. Hum. Reprod. 1995, 10, 3297–3300. [Google Scholar] [CrossRef]
- Luria, O.; Bar, J.; Kovo, M.; Golan, A.; Barnea, O. Feto-maternal interaction: A mathematical model simulating placental response in hypertensive disorders of pregnancy. Reprod. Sci. 2010, 17, 963–969. [Google Scholar] [CrossRef]
- Wicherek, L.; Klimek, M.; Dutsch-Wicherek, M. The level of maternal immune tolerance and fetal maturity. Neuro Endocrinol. Lett. 2005, 26, 561–566. [Google Scholar]
- Mathews, S.; Rao, K.L.; Prasad, K.S.; Kanakavalli, M.K.; Reddy, A.G.; Raj, T.A.; Thangaraj, K.; Pande, G. Propagation of pure fetal and maternal mesenchymal stromal cells from terminal chorionic villi of human term placenta. Sci. Rep. 2015, 5, 10054. [Google Scholar] [CrossRef] [Green Version]
- Igura, K.; Zhang, X.; Takahashi, K.; Mitsuru, A.; Yamaguchi, S.; Takahashi, T. Isolation and characterization of mesenchymal progenitor cells from chorionic villi of human placenta. Cytotherapy 2004, 6, 543–553. [Google Scholar] [CrossRef]
- Kim, M.J.; Shin, K.S.; Jeon, J.H.; Lee, D.R.; Shim, S.H.; Kim, J.K.; Cha, D.-H.; Yoon, T.K.; Kim, G.J. Human chorionic-plate-derived mesenchymal stem cells and Wharton’s jelly-derived mesenchymal stem cells: A comparative analysis of their potential as placenta-derived stem cells. Cell Tissue Res. 2011, 346, 53–64. [Google Scholar] [CrossRef]
- Vulcano, F.; Milazzo, L.; Ciccarelli, C.; Eramo, A.; Sette, G.; Mauro, A.; Macioce, G.; Martinelli, A.; La Torre, R.; Casalbore, P.; et al. Wharton’s jelly mesenchymal stromal cells have contrasting effects on proliferation and phenotype of cancer stem cells from different subtypes of lung cancer. Exp. Cell Res. 2016, 345, 190–198. [Google Scholar] [CrossRef]
- Rylova, Y.V.; Milovanova, N.V.; Gordeeva, M.N.; Savilova, A.M. Characteristics of Multipotent Mesenchymal Stromal Cells from Human Terminal Placenta. Bull. Exp. Biol. Med. 2015, 159, 253–257. [Google Scholar] [CrossRef]
- Zhao, G.; Liu, F.; Lan, S.; Li, P.; Wang, L.; Kou, J.; Qi, X.; Fan, R.; Hao, D.; Wu, C.; et al. Large-scale expansion of Wharton’s jelly-derived mesenchymal stem cells on gelatin microbeads, with retention of self-renewal and multipotency characteristics and the capacity for enhancing skin wound healing. Stem Cell Res. Ther. 2015, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Veryasov, V.N.; Savilova, A.M.; Buyanovskaya, O.A.; Chulkina, M.M.; Pavlovich, S.V.; Sukhikh, G.T. Isolation of mesenchymal stromal cells from extraembryonic tissues and their characteristics. Bull. Exp. Biol. Med. 2014, 157, 119–124. [Google Scholar] [CrossRef]
- Liu, X.; Zheng, P.; Wang, X.; Dai, G.; Cheng, H.; Zhang, Z.; Hua, R.; Niu, X.; Shi, J.; An, Y. A preliminary evaluation of efficacy and safety of Wharton’s jelly mesenchymal stem cell transplantation in patients with type 2 diabetes mellitus. Stem Cell Res. Ther. 2014, 5, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tantrawatpan, C.; Manochantr, S.; Kheolamai, P.; U-Pratya, Y.; Supokawej, A.; Issaragrisil, S. Pluripotent gene expression in mesenchymal stem cells from human umbilical cord Wharton’s jelly and their differentiation potential to neural-like cells. J. Med. Assoc. Thail. 2013, 96, 1208–1217. [Google Scholar]
- Mathew, S.A.; Naik, C.; Cahill, P.A.; Bhonde, R.R. Placental mesenchymal stromal cells as an alternative tool for therapeutic angiogenesis. Cell. Mol. Life Sci. 2020, 77, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.F.; Zhong, Z.-N.; Fu, X.-F.; Peng, D.-X.; Lu, G.-H.; Li, W.-H.; Xu, H.-Y.; Hu, H.-B.; He, J.-M.; Su, W.-Y.; et al. Comparison of cell proliferation, apoptosis, cellular morphology and ultrastructure between human umbilical cord and placenta-derived mesenchymal stem cells. Neurosci. Lett. 2013, 541, 77–82. [Google Scholar] [CrossRef]
- Shaer, A.; Aghdaie, M.H.; Esfandiari, E. Isolation and characterization of Human Mesenchymal Stromal Cells Derived from Placental Decidua Basalis; Umbilical cord Wharton’s Jelly and Amniotic Membrane. Pak. J. Med. Sci. 2014, 30, 1022–1026. [Google Scholar]
- Soma, H.; Ehrmann, R.L.; Hertig, A.T. Human trophoblast in tissue culture. Obstet. Gynecol. 1961, 18, 704–718. [Google Scholar]
- Taylor, P.V.; Hancock, K.W. Viability of human trophoblast in vitro. J. Obstet. Gynaecol. Br. Commonw. 1973, 80, 834–838. [Google Scholar] [CrossRef]
- Yu, J.; Su, X.; Zhu, C.; Pan, Q.; Yang, J.; Ma, J.; Shen, L.; Cao, H.; Qiaoling, P. GFP Labeling and Hepatic Differentiation Potential of Human Placenta-Derived Mesenchymal Stem Cells. Cell. Physiol. Biochem. 2015, 35, 2299–2308. [Google Scholar] [CrossRef]
- Lankford, L.; Selby, T.; Becker, J.; Ryzhuk, V.; Long, C.; Farmer, D.; Wang, A. Early gestation chorionic villi-derived stromal cells for fetal tissue engineering. World J. Stem Cells 2015, 7, 195–207. [Google Scholar] [CrossRef]
- Martini, M.M.; Jeremias, T.D.S.; Kohler, M.C.; Marostica, L.L.; Trentin, A.G.; Alvarez-Silva, M. Human placenta-derived mesenchymal stem cells acquire neural phenotype under the appropriate niche conditions. DNA Cell Biol. 2013, 32, 58–65. [Google Scholar] [CrossRef]
- Makhoul, G.; Chiu, R.C.; Cecere, R. Placental mesenchymal stem cells: A unique source for cellular cardiomyoplasty. Ann. Thorac. Surg. 2013, 95, 1827–1833. [Google Scholar] [CrossRef]
- Zahavi-Goldstein, E.; Blumenfeld, M.; Fuchs-Telem, D.; Pinzur, L.; Rubin, S.; Aberman, Z.; Sher, N.; Ofir, R. Placenta-derived PLX-PAD mesenchymal-like stromal cells are efficacious in rescuing blood flow in hind limb ischemia mouse model by a dose- and site-dependent mechanism of action. Cytotherapy 2017, 19, 1438–1446. [Google Scholar] [CrossRef]
- Shapira, I.; Fainstein, N.; Tsirlin, M.; Stav, I.; Volinsky, E.; Moresi, C.; Ben-Hur, T.; Gorodetsky, R. Placental Stromal Cell Therapy for Experimental Autoimmune Encephalomyelitis: The Role of Route of Cell Delivery. Stem Cells Transl. Med. 2017, 6, 1286–1294. [Google Scholar] [CrossRef]
- Thierry, D.; Bertho, J.M.; Chapel, A.; Gourmelon, P. Cell therapy for the treatment of accidental radiation overexposure. Br. J. Radiol. 2005, 78, 175–179. [Google Scholar] [CrossRef]
- Lange, C.; Brunswig-Spickenheier, B.; Cappallo-Obermann, H.; Eggert, K.; Gehling, U.M.; Rudolph, C.; Schlegelberger, B.; Cornils, K.; Zustin, J.; Spiess, A.-N.; et al. Radiation rescue: Mesenchymal stromal cells protect from lethal irradiation. PLoS ONE 2012, 6, e14486. [Google Scholar] [CrossRef]
- Kawakatsu, M.; Urata, Y.; Goto, S.; Ono, Y.; Li, T.-S. Placental extract protects bone marrow-derived stem/progenitor cells against radiation injury through anti-inflammatory activity. J. Radiat. Res. 2013, 54, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Ofir, R.; Pinzur, L.; Levent, A.; Aberman, Z.; Gorodetsky, R.; Volk, H.D. Mechanism of action of PLX-R18, a placental-derived cellular therapy for the treatment of radiation-induced bone marrow failure. Blood 2015, 126, 2417. [Google Scholar] [CrossRef]
- Sher, N.; Ofir, R. Placenta-Derived Adherent Stromal Cell Therapy for Hematopoietic Disorders: A Case Study of PLX-R18. Cell Transplant. 2018, 27, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Metheny, L.; Eid, S.; Lingas, K.; Ofir, R.; Pinzur, L.; Meyerson, H.; Lazarus, H.M.; Huang, A.Y. Posttransplant Intramuscular Injection of PLX-R18 Mesenchymal-Like Adherent Stromal Cells Improves Human Hematopoietic Engraftment in A Murine Transplant Model. Front. Med. 2018, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Shim, S.; Lee, S.B.; Lee, J.G.; Jang, W.S.; Lee, S.J.; Park, S.; Lee, S.S. Mitigating effects of hUCB-MSCs on the hematopoietic syndrome resulting from total body irradiation. Exp. Hematol. 2013, 41, 346–353.e2. [Google Scholar] [CrossRef]
- Bustos, M.L.; Huleihel, L.; Meyer, E.M.; Donnenberg, A.D.; Donnenberg, V.S.; Sciurba, J.D.; Mroz, L.; McVerry, B.J.; Ellis, B.M.; Kaminski, N.; et al. Activation of human mesenchymal stem cells impacts their therapeutic abilities in lung injury by increasing interleukin (IL)-10 and IL-1RN levels. Stem Cells Transl. Med. 2013, 2, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Quimby, J.M.; Webb, T.L.; Randall, E.; Marolf, A.; Valdes-Martinez, A.; Dow, S.W. Assessment of intravenous adipose-derived allogeneic mesenchymal stem cells for the treatment of feline chronic kidney disease: A randomized, placebo-controlled clinical trial in eight cats. J. Feline Med. Surg. 2015, 18, 165–1711. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Jiang, L.; Long, X.; Fu, C.; Wang, X.; Wu, X.; Liu, Z.; Zhu, F.; Shi, J.; Li, S. Bone-marrow-derived mesenchymal stem cells inhibit gastric aspiration lung injury and inflammation in rats. J. Cell. Mol. Med. 2016, 20, 1706–1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; He, H.; Liu, A.; Xu, J.; Han, J.; Chen, Q.; Hu, S.; Xu, X.; Huang, Y.; Guo, F.; et al. Therapeutic Effects of Bone Marrow-Derived Mesenchymal Stem Cells in Models of Pulmonary and Extrapulmonary Acute Lung Injury. Cell Transplant. 2015, 24, 2629–2642. [Google Scholar] [CrossRef] [Green Version]
- Phinney, D.G.; Hill, K.; Michelson, C.; DuTreil, M.; Hughes, C.; Humphries, S.; Wilkinson, R.; Baddoo, M.; Bayly, E. Biological activities encoded by the murine mesenchymal stem cell transcriptome provide a basis for their developmental potential and broad therapeutic efficacy. Stem Cells 2006, 24, 186–198. [Google Scholar] [CrossRef]
- Chinnadurai, R.; Ng, S.; Velu, V.; Galipeau, J. Challenges in animal modelling of mesenchymal stromal cell therapy for inflammatory bowel disease. World J. Gastroenterol. 2015, 21, 4779–4787. [Google Scholar] [CrossRef]
- Tang, Y.; Chen, Y.; Wang, X.; Song, G.; Li, Y.; Shi, L. Combinatorial Intervention with Mesenchymal Stem Cells and Granulocyte Colony-Stimulating Factor in a Rat Model of Ulcerative Colitis. Dig. Dis. Sci. 2015, 60, 1948–1957. [Google Scholar] [CrossRef]
- Tanaka, Y. Human mesenchymal stem cells as a tool for joint repair in rheumatoid arthritis. Clin. Exp. Rheumatol. 2015, 33, S58–S62. [Google Scholar]
- Da Costa Goncalves, F.; Schneider, N.; Pinto, F.O.; Meyer, F.S.; Visioli, F.; Pfaffenseller, B.; Lopez, P.L.; Passos, E.P.; Cirne-Lima, E.O.; Meurer, L.; et al. Intravenous vs intraperitoneal mesenchymal stem cells administration: What is the best route for treating experimental colitis? World J. Gastroenterol. 2014, 20, 18228–18239. [Google Scholar] [CrossRef]
- Fransson, M.; Piras, E.; Wang, H.; Burman, J.; Duprez, I.; Harris, R.A.; Leblanc, K.; Magnusson, P.U.; Brittebo, E.; Loskog, A.S.I. Intranasal delivery of central nervous system-retargeted human mesenchymal stromal cells prolongs treatment efficacy of experimental autoimmune encephalomyelitis. Immunology 2014, 142, 431–441. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Cao, W.; Shi, Y. Plasticity of mesenchymal stem cells in immunomodulation: Pathological and therapeutic implications. Nat. Immunol. 2014, 15, 1009–1016. [Google Scholar] [CrossRef]
- Kadam, S.; Muthyala, S.; Nair, P.; Bhonde, R. Human placenta-derived mesenchymal stem cells and islet-like cell clusters generated from these cells as a novel source for stem cell therapy in diabetes. Rev. Diabet. Stud. 2010, 7, 168–182. [Google Scholar] [CrossRef] [Green Version]
- Thakkar, U.G.; Trivedi, H.L.; Vanikar, A.V.; Dave, S.D. Insulin-secreting adipose-derived mesenchymal stromal cells with bone marrow-derived hematopoietic stem cells from autologous and allogenic sources for type 1 diabetes mellitus. Cytotherapy 2015, 17, 940–947. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Y.; Gong, H.; Yu, C.; Guo, C.; Wang, F.; Yan, S.; Xu, H. Long term effect and safety of Wharton’s jelly-derived mesenchymal stem cells on type 2 diabetes. Exp. Ther. Med. 2016, 12, 1857–1866. [Google Scholar] [CrossRef]
- Fogel, T.; Gorodetsky, R.; Weksler-Zangen, S. Placenta stromal cell therapy modulates regeneration of pancreas tissue in a dia-betes rat model. Abstract# 397; 55. In Proceedings of the EASD meeting, Barcelona, Spain, 16–20 September 2019. (#3b0e23a4-030c-43f5-b699-9625c30689e7). [Google Scholar]
- Covas, D.T.; Panepucci, R.A.; Fontes, A.M.; Silva, W.A.; Orellana, M.D.; Freitas, M.C.; Neder, L.; Santos, A.R.; Peres, L.C.; Jamur, M.C.; et al. Multipotent mesenchymal stromal cells obtained from diverse human tissues share functional properties and gene-expression profile with CD146+ perivascular cells and fibroblasts. Exp. Hematol. 2008, 36, 642–654. [Google Scholar] [CrossRef]
- Najar, M.; Raicevic, G.; Fayyad-Kazan, H.; Bron, D.; Toungouz, M.; Lagneaux, L. Mesenchymal stromal cells and immunomodulation: A gathering of regulatory immune cells. Cytotherapy 2016, 18, 160–171. [Google Scholar] [CrossRef]
- Shigemoto-Kuroda, T.; Oh, J.Y.; Kim, D.-K.; Jeong, H.J.; Park, S.Y.; Lee, H.J.; Park, J.W.; Kim, T.W.; An, S.Y.; Prockop, D.J.; et al. MSC-derived Extracellular Vesicles Attenuate Immune Responses in Two Autoimmune Murine Models: Type 1 Diabetes and Uveoretinitis. Stem Cell Rep. 2017, 8, 1214–1225. [Google Scholar] [CrossRef] [Green Version]
- Ragni, E.; Papait, A.; Orfei, C.P.; Silini, A.R.; Colombini, A.; Viganò, M.; Libonati, F.; Parolini, O.; de Girolamo, L. Amniotic membrane-mesenchymal stromal cells secreted factors and extracellular vesicle-miRNAs: Anti-inflammatory and regenerative features for musculoskeletal tissues. Stem Cells Transl. Med. 2021. [Google Scholar] [CrossRef]
- Mikkola, H.K.; Gekas, C.; Orkin, S.H.; Dieterlen-Lievre, F. Placenta as a site for hematopoietic stem cell development. Exp. Hematol. 2005, 33, 1048–1054. [Google Scholar] [CrossRef]
- Rhodes, K.E.; Gekas, C.; Wang, Y.; Lux, C.T.; Francis, C.S.; Chan, D.N.; Conway, S.; Orkin, S.H.; Yoder, M.C.; Mikkola, H.K. The emergence of hematopoietic stem cells is initiated in the placental vasculature in the absence of circulation. Cell Stem Cell 2008, 2, 252–263. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.G.; Keller, B.A.; Lankford, L.; Pivetti, C.D.; Hirose, S.; Farmer, D.L.; Wang, A. Age Does Matter: A Pilot Comparison of Placenta-Derived Stromal Cells for in utero Repair of Myelomeningocele Using a Lamb Model. Fetal Diagn. Ther. 2016, 39, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorodetsky, R.; Aicher, W.K. Allogenic Use of Human Placenta-Derived Stromal Cells as a Highly Active Subtype of Mesenchymal Stromal Cells for Cell-Based Therapies. Int. J. Mol. Sci. 2021, 22, 5302. https://doi.org/10.3390/ijms22105302
Gorodetsky R, Aicher WK. Allogenic Use of Human Placenta-Derived Stromal Cells as a Highly Active Subtype of Mesenchymal Stromal Cells for Cell-Based Therapies. International Journal of Molecular Sciences. 2021; 22(10):5302. https://doi.org/10.3390/ijms22105302
Chicago/Turabian StyleGorodetsky, Raphael, and Wilhelm K. Aicher. 2021. "Allogenic Use of Human Placenta-Derived Stromal Cells as a Highly Active Subtype of Mesenchymal Stromal Cells for Cell-Based Therapies" International Journal of Molecular Sciences 22, no. 10: 5302. https://doi.org/10.3390/ijms22105302