
The Role of microRNAs in Development of Colitis-Associated Colorectal Cancer

 ,
,  , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. MicroRNAs Overexpression Induces Colitis-Associated Colorectal Carcinogenesis
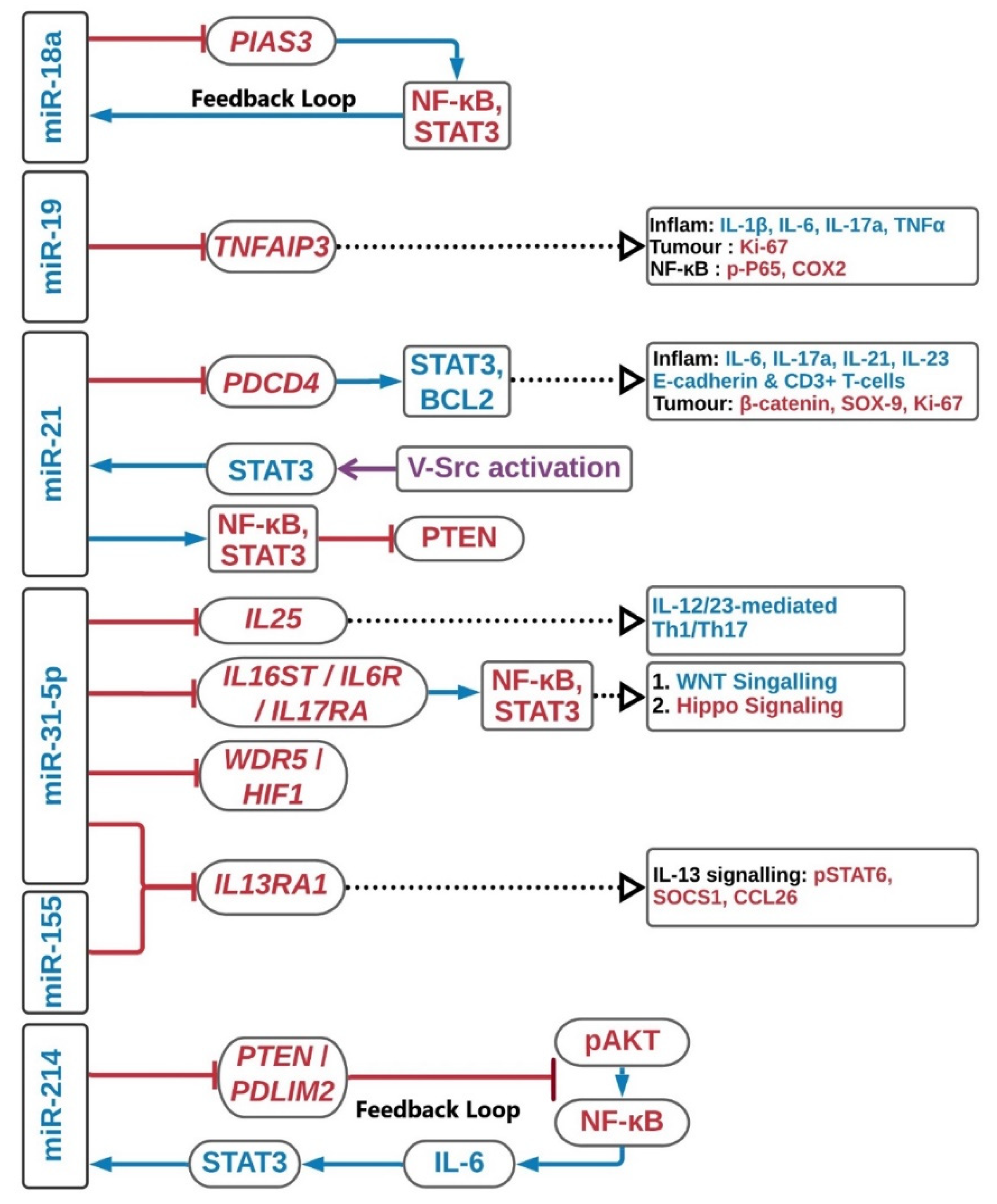
2.1. MiR-17-92 Cluster
2.2. MiR-21
2.3. MiR-31-5p
2.4. MiR-155
2.5. MiRNA-214
3. MicroRNAs as Suppressors of Colitis-Associated Colorectal Carcinogenesis
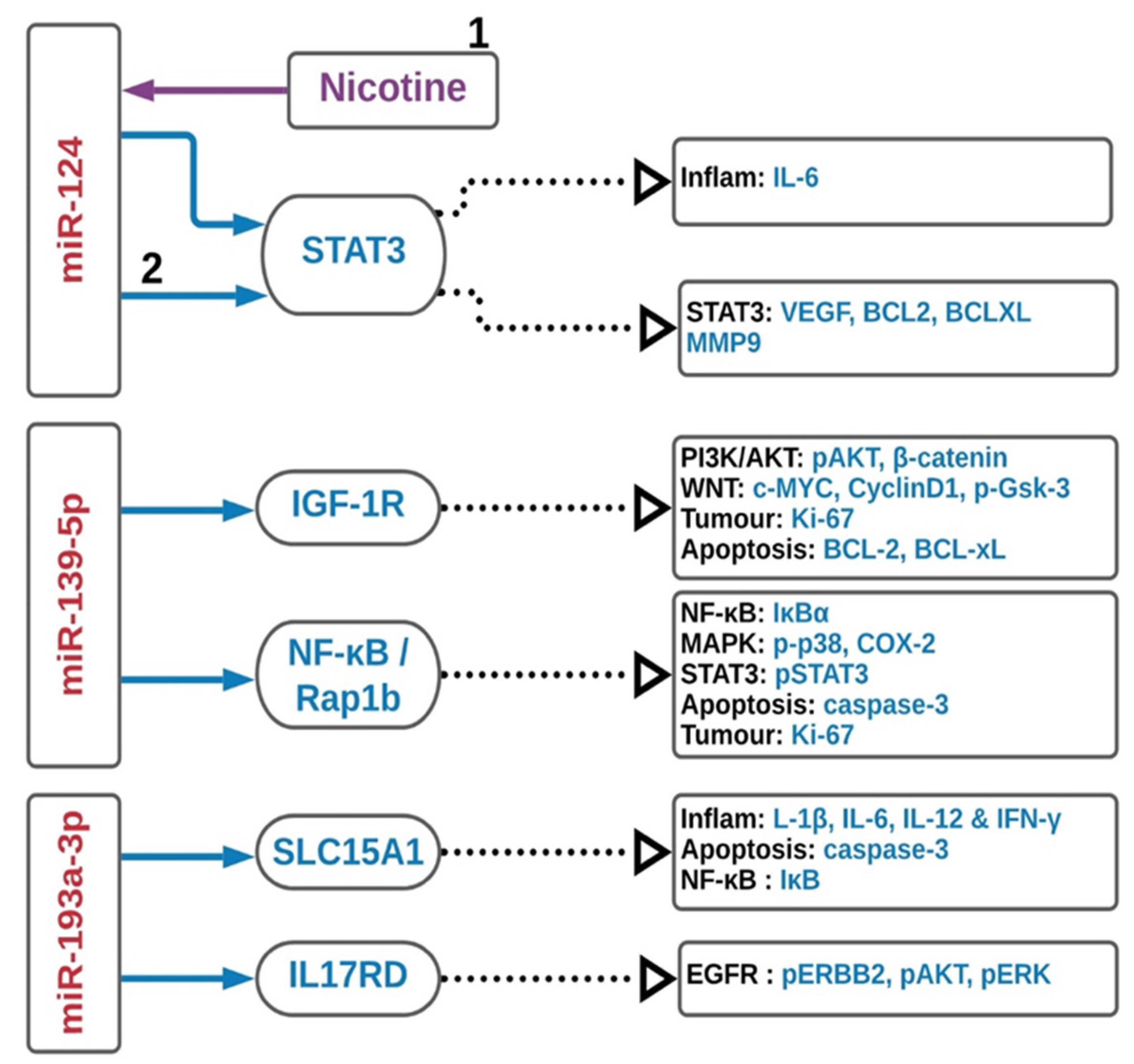
3.1. MiR-124
3.2. MiR-139-5p
3.3. MiR-193a-3p
4. MicroRNAs as a Treatment Tool for Inflammatory Bowel Disease
5. MicroRNAs as Predictive Inflammatory Bowel Disease Biomarker(s)
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BTG1 | BTG Anti-Proliferation Factor 1 |
| CCNDBP1 | Cyclin D1 Binding Protein 1 |
| CYLD | CYLD Lysine 63 Deubiquitinase |
| EGR1 | Early growth response protein 1 |
| EMT | Epithelial–mesenchymal transition |
| HIF1 | Hypoxia-inducible factor 1 |
| IGF-1R | Insulin-like growth factor 1 |
| IL13RA1 | Interleukin 13 Receptor Subunit Alpha 1 |
| IL6 | Interleukin-6 |
| IRAK1 | Interleukin-1 receptor-associated kinase 1 |
| MLCK | Myosin light chain kinase |
| PDCD4 | Programmed Cell Death 4 |
| PDLIM2 | PDZ And LIM Domain 2 |
| PIAS3 | Protein Inhibitor Of Activated STAT 3 |
| PTEN | Phosphatase and tensin homolog |
| RASA1 | RAS p21 protein activator 1 |
| ROCK1 | Rho Associated Coiled-Coil Containing Protein Kinase 1 |
| SLC15A1 | Solute Carrier Family 15 Member 1 |
| STAT3 | Signal transducer and activator of transcription 3 |
| TNFAIP3 | TNF alpha induced protein 3 |
| TRAF6 | TNF Receptor Associated Factor 6 |
| WDR5 | WD Repeat Domain 5 |
References
- Abraham, C.; Cho, J.H. Inflammatory bowel disease. N. Engl. J. Med. 2009, 361, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Molecular Basis of Epithelial Barrier Regulation. Am. J. Pathol. 2006, 169, 1901–1909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, C.N.; Blanchard, J.F.; Kliewer, E.; Wajda, A. Cancer risk in patients with inflammatory bowel disease: A population-based study. Cancer 2001, 91, 854–862. [Google Scholar] [CrossRef]
- Jess, T.; Rungoe, C.; Peyrin-Biroulet, L. Risk of colorectal cancer in patients with ulcerative colitis: A meta-analysis of population-based cohort studies. Clin. Gastroenterol. Hepatol. 2012, 10, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Rubin, D.T.; Huo, D.; Kinnucan, J.A.; Sedrak, M.S.; McCullom, N.E.; Bunnag, A.P.; Raun-Royer, E.P.; Cohen, R.D.; Hanauer, S.B.; Hart, J.; et al. Inflammation Is an Independent Risk Factor for Colonic Neoplasia in Patients with Ulcerative Colitis: A Case—Control Study. Clin. Gastroenterol. Hepatol. 2013, 11, 1601–1608. [Google Scholar] [CrossRef] [Green Version]
- Nieminen, U.; Jussila, A.; Nordling, S.; Mustonen, H.; Färkkilä, M.A. Inflammation and disease duration have a cumulative effect on the risk of dysplasia and carcinoma in IBD: A case-control observational study based on registry data. Int. J. Cancer 2014, 134, 189–196. [Google Scholar] [CrossRef]
- Eaden, J.A. The risk of colorectal cancer in ulcerative colitis: A meta-analysis. Gut 2001, 48, 526–535. [Google Scholar] [CrossRef] [Green Version]
- Canavan, C.; Abrams, K.R.; Mayberry, J. Meta-analysis: Colorectal and small bowel cancer risk in patients with Crohn’s disease. Aliment. Pharmacol. Ther. 2006, 23, 1097–1104. [Google Scholar] [CrossRef]
- Jensen, A.B.; Larsen, M.; Gislum, M.; Skriver, M.V.; Jepsen, P.; Norgaard, B.; Sorensen, H.T. Survival after Colorectal Cancer in Patients with Ulcerative Colitis: A Nationwide Population-Based Danish Study. Am. J. Gastroenterol. 2006, 101, 1283–1287. [Google Scholar] [CrossRef]
- Watanabe, T.; Konishi, T.; Kishimoto, J.; Kotake, K.; Muto, T.; Sugihara, K. Ulcerative colitis-associated colorectal cancer shows a poorer survival than sporadic colorectal cancer: A nationwide Japanese study. Inflamm. Bowel Dis. 2011, 17, 802–808. [Google Scholar] [CrossRef]
- Jess, T.; Loftus, E.V.; Velayos, F.S.; Harmsen, W.S.; Zinsmeister, A.R.; Smyrk, T.C.; Schleck, C.D.; Tremaine, W.J.; Melton, L.J.; Munkholm, P.; et al. Risk of Intestinal Cancer in Inflammatory Bowel Disease: A Population-Based Study from Olmsted County, Minnesota. Gastroenterology 2006, 130, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Itzkowitz, S.H. Cancer in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Rutter, M.; Saunders, B.; Wilkinson, K.; Rumbles, S.; Schofield, G.; Kamm, M.; Williams, C.; Price, A.; Talbot, I.; Forbes, A. Severity of inflammation is a risk factor for colorectal neoplasia in ulcerative colitis. Gastroenterology 2004, 126, 451–459. [Google Scholar] [CrossRef]
- Oh, S.Y.; Cho, K.-A.; Kang, J.L.; Kim, K.H.; Woo, S.-Y. Comparison of experimental mouse models of inflammatory bowel disease. Int. J. Mol. Med. 2014, 33, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Morgan, S.J.; Elangbam, C.S. Gastrointestinal Injury Models. In Drug Discovery Toxicology: From Target Assessment to Translational Biomarkers; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 273–278. [Google Scholar]
- Antoniou, E.; Margonis, G.A.; Angelou, A.; Pikouli, A.; Argiri, P.; Karavokyros, I.; Papalois, A.; Pikoulis, E. The TNBS-induced colitis animal model: An overview. Ann. Med. Surg. 2016, 11, 9–15. [Google Scholar] [CrossRef]
- Pizarro, T.T.; Arseneau, K.O.; Cominelli, F. Lessons from genetically engineered animal models XI. Novel mouse models to study pathogenic mechanisms of Crohn’s disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 278, G665–G669. [Google Scholar] [CrossRef]
- Byrne, F.R.; Morony, S.; Warmington, K.; Geng, Z.; Brown, H.L.; Flores, S.A.; Fiorino, M.; Yin, S.L.; Hill, D.; Porkess, V.; et al. CD4+CD45RBHi T cell transfer induced colitis in mice is accompanied by osteopenia which is treatable with recombinant human osteoprotegerin. Gut 2005, 54, 78–86. [Google Scholar] [CrossRef]
- Seamons, A.; Treuting, P.M.; Brabb, T.; Maggio-Price, L. Characterization of Dextran Sodium Sulfate-Induced Inflammation and Colonic Tumorigenesis in Smad3−/− Mice with Dysregulated TGFβ. PLoS ONE 2013, 8, e79182. [Google Scholar] [CrossRef]
- Lee, H.-N.; Yum, H.-W.; Surh, Y.-J. The Azoxymethane Plus Dextran Sulfate Sodium-Induced Mouse Colon Cancer Model for the Study of Dietary Chemoprevention of Inflammation-Associated Carcinogenesis. In Cancer Prevention; Humana Press: New York, NY, USA, 2014; pp. 155–172. [Google Scholar]
- Berg, D.J.; Davidson, N.; Kühn, R.; Müller, W.; Menon, S.; Holland, G.; Thompson-Snipes, L.; Leach, M.W.; Rennick, D. Enterocolitis and colon cancer in interleukin-10-deficient mice are associated with aberrant cytokine production and CD4(+) TH1-like responses. J. Clin. Investig. 1996, 98, 1010–1020. [Google Scholar] [CrossRef]
- Feagins, L.A.; Souza, R.F.; Spechler, S.J. Carcinogenesis in IBD: Potential targets for the prevention of colorectal cancer. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 297–305. [Google Scholar] [CrossRef]
- Robles, A.I.; Traverso, G.; Zhang, M.; Roberts, N.J.; Khan, M.A.; Joseph, C.; Lauwers, G.Y.; Selaru, F.M.; Popoli, M.; Pittman, M.E.; et al. Whole-Exome Sequencing Analyses of Inflammatory Bowel Disease−Associated Colorectal Cancers. Gastroenterology 2016, 150, 931–943. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, A.W.; Nemunaitis, J. Modulation of miRNA activity in human cancer: A new paradigm for cancer gene therapy? Cancer Gene Ther. 2008, 15, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Yang, Y.; Fu, Y.; Guo, F.; Zhang, X.; Xiao, S.; Zhu, W.; Huang, Z.; Zhang, J.; Chen, J. PIAS3-mediated feedback loops promote chronic colitis-associated malignant transformation. Theranostics 2018, 8, 3022–3037. [Google Scholar] [CrossRef]
- Wang, T.; Xu, X.; Xu, Q.; Ren, J.; Shen, S.; Fan, C.; Hou, Y. miR-19a promotes colitis-associated colorectal cancer by regulating tumor necrosis factor alpha-induced protein 3-NF-κB feedback loops. Oncogene 2017, 36, 3240–3251. [Google Scholar] [CrossRef]
- Schetter, A.J.; Leung, S.Y.; Sohn, J.J.; Zanetti, K.A.; Bowman, E.D.; Yanaihara, N.; Yuen, S.T.; Chan, T.L.; Kwong, D.L.W.; Au, G.K.H.; et al. MicroRNA expression profiles associated with prognosis and therapeutic outcome in colon adenocarcinoma. JAMA 2008, 299, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Yang, Y.; Xia, Y.; Okugawa, Y.; Yang, J.; Liang, Y.; Chen, H.; Zhang, P.; Wang, F.; Han, H.; et al. Novel evidence for an oncogenic role of microRNA-21 in colitis-associated colorectal cancer. Gut 2016, 65, 1470–1481. [Google Scholar] [CrossRef]
- Iliopoulos, D.; Jaeger, S.A.; Hirsch, H.A.; Bulyk, M.L.; Struhl, K. STAT3 Activation of miR-21 and miR-181b-1 via PTEN and CYLD Are Part of the Epigenetic Switch Linking Inflammation to Cancer. Mol. Cell 2010, 39, 493–506. [Google Scholar] [CrossRef] [Green Version]
- Benderska, N.; Dittrich, A.-L.; Knaup, S.; Rau, T.T.; Neufert, C.; Wach, S.; Fahlbusch, F.B.; Rauh, M.; Wirtz, R.M.; Agaimy, A.; et al. miRNA-26b Overexpression in Ulcerative Colitis-associated Carcinogenesis. Inflamm. Bowel Dis. 2015, 21, 2039–2051. [Google Scholar] [CrossRef] [Green Version]
- Olaru, A.V.; Selaru, F.M.; Mori, Y.; Vazquez, C.; David, S.; Paun, B.; Cheng, Y.; Jin, Z.; Yang, J.; Agarwal, R.; et al. Dynamic changes in the expression of MicroRNA-31 during inflammatory bowel disease-associated neoplastic transformation. Inflamm. Bowel Dis. 2011, 17, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Bai, J.; Zhang, L.; Lou, F.; Ke, F.; Cai, W.; Wang, H. Conditional knockout of microRNA-31 promotes the development of colitis associated cancer. Biochem. Biophys. Res. Commun. 2017, 490, 62–68. [Google Scholar] [CrossRef]
- El-Daly, S.M.; Omara, E.A.; Hussein, J.; Youness, E.R.; El-Khayat, Z. Differential expression of miRNAs regulating NF-κB and STAT3 crosstalk during colitis-associated tumorigenesis. Mol. Cell. Probes 2019, 47, 101442. [Google Scholar] [CrossRef]
- Shi, T.; Xie, Y.; Fu, Y.; Zhou, Q.; Ma, Z.; Ma, J.; Huang, Z.; Zhang, J.; Chen, J. The signaling axis of microRNA-31/interleukin-25 regulates Th1/Th17-mediated inflammation response in colitis. Mucosal Immunol. 2017, 10, 983–995. [Google Scholar] [CrossRef]
- Tian, Y.; Xu, J.; Li, Y.; Zhao, R.; Du, S.; Lv, C.; Wu, W.; Liu, R.; Sheng, X.; Song, Y.; et al. MicroRNA-31 Reduces Inflammatory Signaling and Promotes Regeneration in Colon Epithelium, and Delivery of Mimics in Microspheres Reduces Colitis in Mice. Gastroenterology 2019, 156, 2281–2296.e6. [Google Scholar] [CrossRef]
- Gwiggner, M.; Martinez-Nunez, R.T.; Whiteoak, S.R.; Bondanese, V.P.; Claridge, A.; Collins, J.E.; Cummings, J.R.F.; Sanchez-Elsner, T. MicroRNA-31 and MicroRNA-155 Are Overexpressed in Ulcerative Colitis and Regulate IL-13 Signaling by Targeting Interleukin 13 Receptor α-1. Genes 2018, 9, 85. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, K.T.; Enos, R.T.; McClellan, J.L.; Cranford, T.L.; Chatzistamou, I.; Singh, U.P.; Nagarkatti, M.; Nagarkatti, P.S.; Fan, D.; Murphy, E.A. MicroRNA-155 deletion promotes tumorigenesis in the azoxymethane-dextran sulfate sodium model of colon cancer. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G347–G358. [Google Scholar] [CrossRef] [Green Version]
- Polytarchou, C.; Hommes, D.W.; Palumbo, T.; Hatziapostolou, M.; Koutsioumpa, M.; Koukos, G.; van der Meulen-de Jong, A.E.; Oikonomopoulos, A.; van Deen, W.K.; Vorvis, C.; et al. MicroRNA214 Is Associated With Progression of Ulcerative Colitis, and Inhibition Reduces Development of Colitis and Colitis-Associated Cancer in Mice. Gastroenterology 2015, 149, 981–992.e11. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Wang, C.; Long, S.; Ma, Y.; Guo, Y.; Huang, Z.; Chen, X.; Zhang, C.; Chen, J.; Zhang, J. C/EBP-β-activated microRNA-223 promotes tumour growth through targeting RASA1 in human colorectal cancer. Br. J. Cancer 2015, 112, 1491–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Yu, T.; Shi, Y.; Ma, C.; Yang, W.; Fang, L.; Sun, M.; Wu, W.; Xiao, F.; Guo, F.; et al. MicroRNA 301A Promotes Intestinal Inflammation and Colitis-Associated Cancer Development by Inhibiting BTG1. Gastroenterology 2017, 152, 1434–1448. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Long, J.; Yin, Y.; Guo, H.; Jiang, E.; Li, Y.; He, Q.; Zeng, C.; Sun, Y. MicroRNA-34a suppresses the invasion and migration of colorectal cancer cells by enhancing EGR1 and inhibiting vimentin. Exp. Ther. Med. 2019. [Google Scholar] [CrossRef]
- Koukos, G.; Polytarchou, C.; Kaplan, J.L.; Morley–Fletcher, A.; Gras–Miralles, B.; Kokkotou, E.; Baril–Dore, M.; Pothoulakis, C.; Winter, H.S.; Iliopoulos, D. MicroRNA-124 Regulates STAT3 Expression and Is Down-regulated in Colon Tissues of Pediatric Patients With Ulcerative Colitis. Gastroenterology 2013, 145, 842–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Z.; Wan, J.-J.; Sun, Y.; Wu, T.; Wang, P.-Y.; Du, P.; Su, D.-F.; Yang, Y.; Liu, X. Nicotine protects against DSS colitis through regulating microRNA-124 and STAT3. J. Mol. Med. 2017, 95, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Zou, F.; Mao, R.; Yang, L.; Lin, S.; Lei, K.; Zheng, Y.; Ding, Y.; Zhang, P.; Cai, G.; Liang, X.; et al. Targeted deletion of miR-139-5p activates MAPK, NF-κB and STAT3 signaling and promotes intestinal inflammation and colorectal cancer. FEBS J. 2016, 283, 1438–1452. [Google Scholar] [CrossRef]
- Maoa, R.; Zou, F.; Yang, L.; Lin, S.; Li, Y.; Ma, M.; Yin, P.; Liang, X.; Liu, J. The loss of MiR-139-5p promotes colitis-associated tumorigenesis by mediating PI3K/AKT/Wnt signaling. Int. J. Biochem. Cell Biol. 2015, 69, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Cao, Y.; Wang, Z.; He, J.; Chen, H.; Xiong, H.; Ren, L.; Shen, C.; Zhang, X.; Yan, Y.; et al. CCAT1 lncRNA Promotes Inflammatory Bowel Disease Malignancy by Destroying Intestinal Barrier via Downregulating miR-185-3p. Inflamm. Bowel Dis. 2019, 25, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Chen, X.; Chen, Q.; Shi, L.; Liang, H.; Zhou, Z.; Liu, Q.; Pang, W.; Hou, D.; Wang, C.; et al. MicroRNA-193a-3p Reduces Intestinal Inflammation in Response to Microbiota via Down-regulation of Colonic PepT1. J. Biol. Chem. 2015, 290, 16099–16115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pekow, J.; Meckel, K.; Dougherty, U.; Huang, Y.; Chen, X.; Almoghrabi, A.; Mustafi, R.; Ayaloglu-Butun, F.; Deng, Z.; Haider, H.I.; et al. miR-193a-3p is a Key Tumor Suppressor in Ulcerative Colitis-Associated Colon Cancer and Promotes Carcinogenesis through Upregulation of IL17RD. Clin. Cancer Res. 2017, 23, 5281–5291. [Google Scholar] [CrossRef] [Green Version]
- Cellura, D.; Pickard, K.; Quaratino, S.; Parker, H.; Strefford, J.C.; Thomas, G.J.; Mitter, R.; Mirnezami, A.H.; Peake, N.J. miR-19-Mediated Inhibition of Transglutaminase-2 Leads to Enhanced Invasion and Metastasis in Colorectal Cancer. Mol. Cancer Res. 2015, 13, 1095–1105. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, R.; Yang, F.; Cheng, R.; Chen, X.; Cui, S.; Gu, Y.; Sun, W.; You, C.; Liu, Z.; et al. miR-19a promotes colorectal cancer proliferation and migration by targeting TIA1. Mol. Cancer 2017, 16, 53. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Wang, X.; Wen, C.; Yang, X.; Song, M.; Chen, J.; Wang, C.; Zhang, B.; Wang, L.; Iwamoto, A.; et al. Hsa-miR-19a is associated with lymph metastasis and mediates the TNF-α induced epithelial-to-mesenchymal transition in colorectal cancer. Sci. Rep. 2015, 5, 13350. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; She, S.; Li, D.; Liu, Z.; Yang, X.; Zeng, Z.; Liu, F. Role of miR-19a targeting TNF-α in mediating ulcerative colitis. Scand. J. Gastroenterol. 2013, 48, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, J.; Wang, Q.; Meng, G.; Lv, X.; Zhou, H.; Li, W.; Zhang, J. The relationship between microRNAs and the STAT3-related signaling pathway in cancer. Tumor Biol. 2017, 39, 101042831771986. [Google Scholar] [CrossRef] [Green Version]
- Yau, T.O.; Wu, C.W.; Dong, Y.; Tang, C.-M.; Ng, S.S.M.; Chan, F.K.L.; Sung, J.J.Y.; Yu, J. microRNA-221 and microRNA-18a identification in stool as potential biomarkers for the non-invasive diagnosis of colorectal carcinoma. Br. J. Cancer 2014, 111, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Yau, T.O.; Tang, C.-M.; Harriss, E.K.; Dickins, B.; Polytarchou, C. Faecal microRNAs as a non-invasive tool in the diagnosis of colonic adenomas and colorectal cancer: A meta-analysis. Sci. Rep. 2019, 9, 9491. [Google Scholar] [CrossRef] [PubMed]
- Svrcek, M.; El-Murr, N.; Wanherdrick, K.; Dumont, S.; Beaugerie, L.; Cosnes, J.; Colombel, J.-F.; Tiret, E.; Fléjou, J.-F.; Lesuffleur, T.; et al. Overexpression of microRNAs-155 and 21 targeting mismatch repair proteins in inflammatory bowel diseases. Carcinogenesis 2013, 34, 828–834. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Liang, Y.; Yang, J.; Xia, Y.; Chen, H.; Han, H.; Yang, Y.; Wu, W.; Gao, R.; Qin, H. MicroRNA-21 Knockout Improve the Survival Rate in DSS Induced Fatal Colitis through Protecting against Inflammation and Tissue Injury. PLoS ONE 2013, 8, e66814. [Google Scholar] [CrossRef] [Green Version]
- Ando, Y.; Mazzurana, L.; Forkel, M.; Okazaki, K.; Aoi, M.; Schmidt, P.T.; Mjösberg, J.; Bresso, F. Downregulation of MicroRNA-21 in Colonic CD3+ T Cells in UC Remission. Inflamm. Bowel Dis. 2016, 22, 2788–2793. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Dong, F.; Arendovich, N.; Zhang, J.; Huang, Y.; Kwon, J.H. Divergent influence of microRNA-21 deletion on murine colitis phenotypes. Inflamm. Bowel Dis. 2014, 20, 1972–1985. [Google Scholar] [CrossRef]
- Nosho, K.; Igarashi, H.; Nojima, M.; Ito, M.; Maruyama, R.; Yoshii, S.; Naito, T.; Sukawa, Y.; Mikami, M.; Sumioka, W.; et al. Association of microRNA-31 with BRAF mutation, colorectal cancer survival and serrated pathway. Carcinogenesis 2014, 35, 776–783. [Google Scholar] [CrossRef] [Green Version]
- Kiss, I.; Mlcochova, J.; Bortlicek, Z.; Poprach, A.; Drabek, J.; Vychytilova-Faltejskova, P.; Svoboda, M.; Buchler, T.; Batko, S.; Ryska, A.; et al. Efficacy and Toxicity of Panitumumab After Progression on Cetuximab and Predictive Value of MiR-31-5p in Metastatic Wild-type KRAS Colorectal Cancer Patients. Anticancer Res. 2016, 36, 4955–4960. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Mitsuhashi, K.; Igarashi, H.; Nosho, K.; Naito, T.; Yoshii, S.; Takahashi, H.; Fujita, M.; Sukawa, Y.; Yamamoto, E.; et al. MicroRNA-31 expression in relation to BRAF mutation, CpG island methylation and colorectal continuum in serrated lesions. Int. J. Cancer 2014, 135, 2507–2515. [Google Scholar] [CrossRef]
- Sun, D.; Yu, F.; Ma, Y.; Zhao, R.; Chen, X.; Zhu, J.; Zhang, C.-Y.; Chen, J.; Zhang, J. MicroRNA-31 Activates the RAS Pathway and Functions as an Oncogenic MicroRNA in Human Colorectal Cancer by Repressing RAS p21 GTPase Activating Protein 1 (RASA1). J. Biol. Chem. 2013, 288, 9508–9518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, I.; Nosho, K.; Kanno, S.; Igarashi, H.; Kurihara, H.; Ishigami, K.; Ishiguro, K.; Mitsuhashi, K.; Maruyama, R.; Koide, H.; et al. EZH2 expression is a prognostic biomarker in patients with colorectal cancer treated with anti-EGFR therapeutics. Oncotarget 2017, 8, 17810. [Google Scholar] [CrossRef]
- Aoki, H.; Nosho, K.; Igarashi, H.; Ito, M.; Mitsuhashi, K.; Naito, T.; Yamamoto, E.; Tanuma, T.; Nomura, M.; Maguchi, H.; et al. MicroRNA-31 expression in colorectal serrated pathway progression. World J. Gastroenterol. 2014, 20, 12346–12349. [Google Scholar] [CrossRef]
- Kurihara, H.; Maruyama, R.; Ishiguro, K.; Kanno, S.; Yamamoto, I.; Ishigami, K.; Mitsuhashi, K.; Igarashi, H.; Ito, M.; Tanuma, T.; et al. The relationship between EZH2 expression and microRNA-31 in colorectal cancer and the role in evolution of the serrated pathway. Oncotarget 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Yau, T.O. Precision treatment in colorectal cancer: Now and the future. JGH Open J. Gastroenterol. Hepatol. 2019, 3, 361–369. [Google Scholar] [CrossRef]
- Igarashi, H.; Kurihara, H.; Mitsuhashi, K.; Ito, M.; Okuda, H.; Kanno, S.; Naito, T.; Yoshii, S.; Takahashi, H.; Kusumi, T.; et al. Association of MicroRNA-31-5p with Clinical Efficacy of Anti-EGFR Therapy in Patients with Metastatic Colorectal Cancer. Ann. Surg. Oncol. 2015, 22, 2640–2648. [Google Scholar] [CrossRef]
- Chen, T.; Yao, L.-Q.; Shi, Q.; Ren, Z.; Ye, L.-C.; Xu, J.-M.; Zhou, P.-H.; Zhong, Y.-S. MicroRNA-31 contributes to colorectal cancer development by targeting factor inhibiting HIF-1α (FIH-1). Cancer Biol. Ther. 2014, 15, 516–523. [Google Scholar] [CrossRef] [Green Version]
- Necela, B.M.; Carr, J.M.; Asmann, Y.W.; Thompson, E.A. Differential expression of microRNAs in tumors from chronically inflamed or genetic (APC(Min/+)) models of colon cancer. PLoS ONE 2011, 6, e18501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Y.-L.; Wang, H.-F.; Sun, Z.-Q.; Tang, Y.; Han, X.-N.; Yu, X.-B.; Liu, K. Up-regulated miR-155-5p promotes cell proliferation, invasion and metastasis in colorectal carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 6988–6994. [Google Scholar] [PubMed]
- Khoshinani, H.M.; Afshar, S.; Pashaki, A.S.; Mahdavinezhad, A.; Nikzad, S.; Najafi, R.; Amini, R.; Gholami, M.H.; Khoshghadam, A.; Saidijam, M. Involvement of miR-155/FOXO3a and miR-222/PTEN in acquired radioresistance of colorectal cancer cell line. Jpn. J. Radiol. 2017, 35, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Al-Haidari, A.A.; Syk, I.; Thorlacius, H. MiR-155-5p positively regulates CCL17-induced colon cancer cell migration by targeting RhoA. Oncotarget 2017, 8, 14887–14896. [Google Scholar] [CrossRef] [Green Version]
- Bakirtzi, K.; Hatziapostolou, M.; Karagiannides, I.; Polytarchou, C.; Jaeger, S.; Iliopoulos, D.; Pothoulakis, C. Neurotensin signaling activates microRNAs-21 and -155 and Akt, promotes tumor growth in mice, and is increased in human colon tumors. Gastroenterology 2011, 141, 1749–1761.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceppi, M.; Pereira, P.M.; Dunand-Sauthier, I.; Barras, E.; Reith, W.; Santos, M.A.; Pierre, P. MicroRNA-155 modulates the interleukin-1 signaling pathway in activated human monocyte-derived dendritic cells. Proc. Natl. Acad. Sci. USA 2009, 106, 2735–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, R.M.; Taganov, K.D.; Boldin, M.P.; Cheng, G.; Baltimore, D. MicroRNA-155 is induced during the macrophage inflammatory response. Proc. Natl. Acad. Sci. USA 2007, 104, 1604–1609. [Google Scholar] [CrossRef] [Green Version]
- Min, M.; Peng, L.; Yang, Y.; Guo, M.; Wang, W.; Sun, G. MicroRNA-155 Is Involved in the Pathogenesis of Ulcerative Colitis by Targeting FOXO3a. Inflamm. Bowel Dis. 2014, 20, 652–659. [Google Scholar] [CrossRef]
- Cremer, T.J.; Ravneberg, D.H.; Clay, C.D.; Piper-Hunter, M.G.; Marsh, C.B.; Elton, T.S.; Gunn, J.S.; Amer, A.; Kanneganti, T.-D.; Schlesinger, L.S.; et al. MiR-155 Induction by F. novicida but Not the Virulent F. tularensis Results in SHIP Down-Regulation and Enhanced Pro-Inflammatory Cytokine Response. PLoS ONE 2009, 4, e8508. [Google Scholar] [CrossRef]
- Pathak, S.; Grillo, A.R.; Scarpa, M.; Brun, P.; D’Incà, R.; Nai, L.; Banerjee, A.; Cavallo, D.; Barzon, L.; Palù, G.; et al. MiR-155 modulates the inflammatory phenotype of intestinal myofibroblasts by targeting SOCS1 in ulcerative colitis. Exp. Mol. Med. 2015, 47, e164. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhu, F.; Li, H.; Fan, H.; Wu, H.; Dong, Y.; Chu, S.; Tan, C.; Wang, Q.; He, H.; et al. MiR-155 contributes to intestinal barrier dysfunction in DSS-induced mice colitis via targeting HIF-1α/TFF-3 axis. Aging 2020, 12, 14966–14977. [Google Scholar] [CrossRef]
- Singh, U.P.; Murphy, A.E.; Enos, R.T.; Shamran, H.A.; Singh, N.P.; Guan, H.; Hegde, V.L.; Fan, D.; Price, R.L.; Taub, D.D.; et al. miR-155 deficiency protects mice from experimental colitis by reducing T helper type 1/type 17 responses. Immunology 2014, 143, 478–489. [Google Scholar] [CrossRef]
- Chandrasekaran, K.S.; Sathyanarayanan, A.; Karunagaran, D. MicroRNA-214 suppresses growth, migration and invasion through a novel target, high mobility group AT-hook 1, in human cervical and colorectal cancer cells. Br. J. Cancer 2016, 115, 741–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.L.; He, G.Y.; Lan, X.L.; Zeng, Z.C.; Guan, J.; Ding, Y.; Qian, X.L.; Liao, W.T.; Ding, Y.Q.; Liang, L. Inhibition of ATG12-mediated autophagy by miR-214 enhances radiosensitivity in colorectal cancer. Oncogenesis 2018, 7, 16. [Google Scholar] [CrossRef]
- He, G.Y.; Hu, J.L.; Zhou, L.; Zhu, X.H.; Xin, S.N.; Zhang, D.; Lu, G.F.; Liao, W.T.; Ding, Y.Q.; Liang, L. The FOXD3/miR-214/MED19 axis suppresses tumour growth and metastasis in human colorectal cancer. Br. J. Cancer 2016, 115, 1367–1378. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.-L.; Wang, Z.; Zeng, Z.-L.; Wu, W.-J.; Zhang, D.-S.; Luo, H.-Y.; Wang, F.-H.; Qiu, M.-Z.; Wang, D.; Ren, C.; et al. Identification of microRNA-214 as a negative regulator of colorectal cancer liver metastasis by way of regulation of fibroblast growth factor receptor 1 expression. Hepatology 2014, 60, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Cristóbal, I.; Caramés, C.; Madoz-Gúrpide, J.; Rojo, F.; Aguilera, O.; García-Foncillas, J. Downregulation of miR-214 is specific of liver metastasis in colorectal cancer and could play a role determining the metastatic niche. Int. J. Colorectal Dis. 2014, 29, 885. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, J.; Wang, X.; Zheng, C.; Ma, W. Downregulation of microRNA-214 and overexpression of FGFR-1 contribute to hepatocellular carcinoma metastasis. Biochem. Biophys. Res. Commun. 2013, 439, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Kanaan, Z.; Rai, S.N.; Eichenberger, M.R.; Barnes, C.; Dworkin, A.M.; Weller, C.; Cohen, E.; Roberts, H.; Keskey, B.; Petras, R.E.; et al. Differential MicroRNA expression tracks neoplastic progression in inflammatory bowel disease-associated colorectal cancer. Hum. Mutat. 2012, 33, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Jinushi, T.; Shibayama, Y.; Kinoshita, I.; Oizumi, S.; Jinushi, M.; Aota, T.; Takahashi, T.; Horita, S.; Dosaka-Akita, H.; Iseki, K. Low expression levels of microRNA-124-5p correlated with poor prognosis in colorectal cancer via targeting of SMC4. Cancer Med. 2014, 3, 1544–1552. [Google Scholar] [CrossRef]
- Taniguchi, K.; Sugito, N.; Kumazaki, M.; Shinohara, H.; Yamada, N.; Nakagawa, Y.; Ito, Y.; Otsuki, Y.; Uno, B.; Uchiyama, K.; et al. MicroRNA-124 inhibits cancer cell growth through PTB1/PKM1/PKM2 feedback cascade in colorectal cancer. Cancer Lett. 2015, 363, 17–27. [Google Scholar] [CrossRef]
- Sun, Y.; Zhao, X.; Zhou, Y.; Hu, Y. miR-124, miR-137 and miR-340 regulate colorectal cancer growth via inhibition of the Warburg effect. Oncol. Rep. 2012, 28, 1346–1352. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, K.; Sugito, N.; Kumazaki, M.; Shinohara, H.; Yamada, N.; Matsuhashi, N.; Futamura, M.; Ito, Y.; Otsuki, Y.; Yoshida, K.; et al. Positive feedback of DDX6/c-Myc/PTB1 regulated by miR-124 contributes to maintenance of the Warburg effect in colon cancer cells. Biochim. Biophys. Acta 2015, 1852, 1971–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warburg, O. On the Origin of Cancer Cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, J.; Manley, J.L. Turning on a Fuel Switch of Cancer: hnRNP Proteins Regulate Alternative Splicing of Pyruvate Kinase mRNA. Cancer Res. 2010, 70, 8977–8980. [Google Scholar] [CrossRef] [Green Version]
- Christofk, H.R.; Vander Heiden, M.G.; Harris, M.H.; Ramanathan, A.; Gerszten, R.E.; Wei, R.; Fleming, M.D.; Schreiber, S.L.; Cantley, L.C. The M2 splice isoform of pyruvate kinase is important for cancer metabolism and tumour growth. Nature 2008, 452, 230–233. [Google Scholar] [CrossRef]
- Demaria, M.; Poli, V. PKM2, STAT3 and HIF-1α. JAK-STAT 2012, 1, 194–196. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhao, X.; Luo, M.; Zhou, Y.; Ren, W.; Wu, K.; Li, X.; Shen, J.; Hu, Y. The Pro-Apoptotic Role of the Regulatory Feedback Loop between miR-124 and PKM1/HNF4α in Colorectal Cancer Cells. Int. J. Mol. Sci. 2014, 15, 4318–4332. [Google Scholar] [CrossRef] [Green Version]
- Demaria, M.; Giorgi, C.; Lebiedzinska, M.; Esposito, G.; D’Angeli, L.; Bartoli, A.; Gough, D.J.; Turkson, J.; Levy, D.E.; Watson, C.J.; et al. A STAT3-mediated metabolic switch is involved in tumour transformation and STAT3 addiction. Aging 2010, 2, 823–842. [Google Scholar] [CrossRef] [Green Version]
- Marzec, M.; Liu, X.; Wong, W.; Yang, Y.; Pasha, T.; Kantekure, K.; Zhang, P.; Woetmann, A.; Cheng, M.; Odum, N.; et al. Oncogenic kinase NPM/ALK induces expression of HIF1α mRNA. Oncogene 2011, 30, 1372–1378. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-F.; Lai, R. STAT3 in Cancer—Friend or Foe? Cancers 2014, 6, 1408–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, K. Role of STAT3 in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 5110–5114. [Google Scholar] [CrossRef] [Green Version]
- Musso, A.; Dentelli, P.; Carlino, A.; Chiusa, L.; Repici, A.; Sturm, A.; Fiocchi, C.; Rizzetto, M.; Pegoraro, L.; Sategna-Guidetti, C.; et al. Signal transducers and activators of transcription 3 signaling pathway: An essential mediator of inflammatory bowel disease and other forms of intestinal inflammation. Inflamm. Bowel Dis. 2005, 11, 91–98. [Google Scholar] [CrossRef]
- Atreya, R.; Neurath, M.F. Involvement of IL-6 in the pathogenesis of inflammatory bowel disease and colon cancer. Clin. Rev. Allergy Immunol. 2005, 28, 187–196. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, Y.; Yue, X.; Li, H.; Luo, X.; Wang, Y.; Wang, K.; Wan, J. MiR-124 suppresses growth of human colorectal cancer by inhibiting STAT3. PLoS ONE 2013, 8, e70300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Ji, P.; Zhang, Y.; LaComb, J.F.; Tian, X.; Li, E.; Williams, J.L. Aberrant DNA Methylation: Implications in Racial Health Disparity. PLoS ONE 2016, 11, e0153125. [Google Scholar] [CrossRef]
- Harada, T.; Yamamoto, E.; Yamano, H.; Nojima, M.; Maruyama, R.; Kumegawa, K.; Ashida, M.; Yoshikawa, K.; Kimura, T.; Harada, E.; et al. Analysis of DNA methylation in bowel lavage fluid for detection of colorectal cancer. Cancer Prev. Res. 2014, 7, 1002–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Dong, Y.; Zhu, N.; Tsoi, H.; Zhao, Z.; Wu, C.W.; Wang, K.; Zheng, S.; Ng, S.S.; Chan, F.K.; et al. microRNA-139-5p exerts tumor suppressor function by targeting NOTCH1 in colorectal cancer. Mol. Cancer 2014, 13, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Yin, Y.; Zhang, J.; Zhang, B.; Bian, Z.; Quan, C.; Zhou, L.; Hu, Y.; Wang, Q.; Ni, S.; et al. MiR-139-5p inhibits migration and invasion of colorectal cancer by downregulating AMFR and NOTCH1. Protein Cell 2014, 5, 851–861. [Google Scholar] [CrossRef] [Green Version]
- Shen, K.; Liang, Q.; Xu, K.; Cui, D.; Jiang, L.; Yin, P.; Lu, Y.; Li, Q.; Liu, J. MiR-139 inhibits invasion and metastasis of colorectal cancer by targeting the type I insulin-like growth factor receptor. Biochem. Pharmacol. 2012, 84, 320–330. [Google Scholar] [CrossRef]
- Liu, H.; Yin, Y.; Hu, Y.; Feng, Y.; Bian, Z.; Yao, S.; Li, M.; You, Q.; Huang, Z. miR-139-5p sensitizes colorectal cancer cells to 5-fluorouracil by targeting NOTCH-1. Pathol.-Res. Pract. 2016, 212, 643–649. [Google Scholar] [CrossRef]
- Li, Q.; Liang, X.; Wang, Y.; Meng, X.; Xu, Y.; Cai, S.; Wang, Z.; Liu, J.; Cai, G. miR-139-5p Inhibits the Epithelial-Mesenchymal Transition and Enhances the Chemotherapeutic Sensitivity of Colorectal Cancer Cells by Downregulating BCL2. Sci. Rep. 2016, 6, 27157. [Google Scholar] [CrossRef]
- Bian, Z.; Zhang, J.; Li, M.; Feng, Y.; Yao, S.; Song, M.; Qi, X.; Fei, B.; Yin, Y.; Hua, D.; et al. Long non-coding RNA LINC00152 promotes cell proliferation, metastasis, and confers 5-FU resistance in colorectal cancer by inhibiting miR-139-5p. Oncogenesis 2017, 6, 395. [Google Scholar] [CrossRef]
- Mamoori, A.; Wahab, R.; Islam, F.; Lee, K.; Vider, J.; Lu, C.-T.; Gopalan, V.; Lam, A.K. Clinical and biological significance of miR-193a-3p targeted KRAS in colorectal cancer pathogenesis. Hum. Pathol. 2018, 71, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Duan, B.; Hu, J.; Yu, H.; Sheng, H.; Gao, H.; Huang, J. Decreased expression of miR-193a-3p is associated with poor prognosis in colorectal cancer. Oncol. Lett. 2017, 14, 1061–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, H.; Takahashi, M.; Ohnuma, S.; Unno, M.; Yoshino, Y.; Ouchi, K.; Takahashi, S.; Yamada, Y.; Shimodaira, H.; Ishioka, C. microRNA-193a-3p is specifically down-regulated and acts as a tumor suppressor in BRAF-mutated colorectal cancer. BMC Cancer 2017, 17, 723. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Fabbri, M.; Calin, G.A. MicroRNAs and other non-coding RNAs as targets for anticancer drug development. Nat. Rev. Drug Discov. 2013, 12, 847–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, V.; Winkler, J. miRNA-based therapies: Strategies and delivery platforms for oligonucleotide and non-oligonucleotide agents. Future Med. Chem. 2014, 6, 1967–1984. [Google Scholar] [CrossRef] [Green Version]
- Chapman, C.G.; Pekow, J. The emerging role of miRNAs in inflammatory bowel disease: A review. Therap. Adv. Gastroenterol. 2015, 8, 4–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.-J.; Wu, J.-J.; Jiang, W.-L.; Xiao, J.-H.; Tao, K.-Z.; Ma, L.; Zheng, P.; Wan, R.; Wang, X.-P. MicroRNA-155 promotes the pathogenesis of experimental colitis by repressing SHIP-1 expression. World J. Gastroenterol. 2017, 23, 976–985. [Google Scholar] [CrossRef]
- Selbach, M.; Schwanhäusser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread changes in protein synthesis induced by microRNAs. Nature 2008, 455, 58–63. [Google Scholar] [CrossRef]
- López-Urrutia, E.; Bustamante Montes, L.P.; Ladrón de Guevara Cervantes, D.; Pérez-Plasencia, C.; Campos-Parra, A.D. Crosstalk Between Long Non-coding RNAs, Micro-RNAs and mRNAs: Deciphering Molecular Mechanisms of Master Regulators in Cancer. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Wen, Z.-H.; Agoramoorthy, G.; Lin, C.-S. Therapeutic microRNA Delivery Strategies with Special Emphasis on Cancer Therapy and Tumorigenesis: Current Trends and Future Challenges. Curr. Drug Metab. 2016, 17, 469–477. [Google Scholar] [CrossRef]
- Tenore, G.C.; Ritieni, A.; Campiglia, P.; Stiuso, P.; Di Maro, S.; Sommella, E.; Pepe, G.; D’Urso, E.; Novellino, E. Antioxidant peptides from “Mozzarella di Bufala Campana DOP” after simulated gastrointestinal digestion: In vitro intestinal protection, bioavailability, and anti-haemolytic capacity. J. Funct. Foods 2015, 15, 365–375. [Google Scholar] [CrossRef]
- Nielsen, O.H.; Ainsworth, M.A. Tumor necrosis factor inhibitors for inflammatory bowel disease. N. Engl. J. Med. 2013, 369, 754–762. [Google Scholar] [CrossRef] [Green Version]
- Ben-Horin, S.; Chowers, Y. Review article: Loss of response to anti-TNF treatments in Crohn’s disease. Aliment. Pharmacol. Ther. 2011, 33, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Morilla, I.; Uzzan, M.; Laharie, D.; Cazals-Hatem, D.; Denost, Q.; Daniel, F.; Belleannee, G.; Bouhnik, Y.; Wainrib, G.; Panis, Y.; et al. Colonic MicroRNA Profiles, Identified by a Deep Learning Algorithm, That Predict Responses to Therapy of Patients With Acute Severe Ulcerative Colitis. Clin. Gastroenterol. Hepatol. 2019, 17, 905–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.W.; Ng, S.C.; Dong, Y.; Tian, L.; Ng, S.S.M.; Leung, W.W.; Law, W.T.; Yau, T.O.; Chan, F.K.L.; Sung, J.J.Y.; et al. Identification of microRNA-135b in Stool as a Potential Noninvasive Biomarker for Colorectal Cancer and Adenoma. Clin. Cancer Res. 2014, 20, 2994–3002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yau, T.O.; Wu, C.W.; Tang, C.-M.; Chen, Y.; Fang, J.; Dong, Y.; Liang, Q.; Ng, S.S.M.; Chan, F.K.L.; Sung, J.J.Y.; et al. microRNA-20a in human faeces as a non-invasive biomarker for colorectal cancer. Oncotarget 2016, 7, 1559–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tepus, M.; Yau, T.O. Non-Invasive Colorectal Cancer Screening: An Overview. Gastrointest. Tumors 2020, 3–4, 62–73. [Google Scholar] [CrossRef]
- Verdier, J.; Breunig, I.R.; Ohse, M.C.; Roubrocks, S.; Kleinfeld, S.; Roy, S.; Streetz, K.; Trautwein, C.; Roderburg, C.; Sellge, G. Faecal Micro-RNAs in Inflammatory Bowel Diseases. J. Crohns. Colitis 2020, 14, 110–117. [Google Scholar] [CrossRef]
- Ji, Y.; Li, X.; Zhu, Y.; Li, N.; Zhang, N.; Niu, M. Faecal microRNA as a biomarker of the activity and prognosis of inflammatory bowel diseases. Biochem. Biophys. Res. Commun. 2018, 503, 2443–2450. [Google Scholar] [CrossRef]
- Monaghan, T.M.; Seekatz, A.M.; Markham, N.O.; Yau, T.O.; Hatziapostolou, M.; Jilani, T.; Christodoulou, N.; Roach, B.; Birli, E.; Pomenya, O.; et al. Fecal microbiota transplantation for recurrent Clostridioides difficile infection associates with functional alterations in circulating microRNAs. Gastroenterology 2021. [Google Scholar] [CrossRef]
- Schönauen, K.; Le, N.; von Arnim, U.; Schulz, C.; Malfertheiner, P.; Link, A. Circulating and Fecal microRNAs as Biomarkers for Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2018, 24, 1547–1557. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Type | Method | Prevalent of Response | Limitations |
|---|---|---|---|
| Colitis Models | |||
| Chemically Induced | DSS | Epithelial damage | Does not require T and/or B cell responses [14], high severity variability [15] |
| TNBS/DNBS | Epithelial damage, Immune-driven | Aetiopathogenesis not clear [16] | |
| Oxazolone | Epithelial damage, Immune-driven | International administration required [15] | |
| Spontaneous Mutation | SAMP1/Yit | Immune-driven | Affect small intestine only [17], low breeding rate [15] |
| C3H/HeJBir | Immune-driven | Greatly influenced by caging conditions [15] | |
| Adoptive T Cell Transfer | CD4+CD45RBhi | Immune-driven | Lack of a full overview of colitis development [18], expensive |
| Genetically Engineered | IL-10−/− | Immune-driven | Lack of focal granulomatous inflammation and Transmural inflammation [17] |
| Colitis-Associated Colorectal Tumorigenesis Models | |||
| Chemically Induced | DSS | Epithelial damage | Low cancer incidents [19]. |
| AOM/DSS | Epithelial damage | The most common CAC model [20] | |
| Genetically Engineered | IL-10−/− | Immune-driven | ~60% of cancer Incidence [21] |
| miRNA | Target Gene(s) | Function | Reference(s) |
|---|---|---|---|
| Upregulation | |||
| miR-18a | PIAS3 | Proliferation, cell apoptosis | [26] |
| miR-19a | TNFAIP3 | Activate NF-κB signaling | [27] |
| miR-21 | PDCD4, PTEN | Invasion, intravasation, metastasis, apoptosis | [28,29,30] |
| miR-26b | CCNDBP1 | Tumorigenesis and development of digestive diseases | [31] |
| miR-31 | HIF1, WDR5, IL13RA1 | Activate RAS signaling, stimulating tumorigenesis and correlates with serrated CRC | [32,33,34,35,36] |
| miR-146b | TRAF6, IRAK1 | [34] | |
| miR-155 | IL13RA1 | Negative feedback loop controlling IL-1β | [34,37,38] |
| miR-181b-1 | CYLD | Cellular transformation | [30,34] |
| miR-214 | PDLIM2, PTEN | Malignant transformation | [39] |
| miR-221 | PDLIM2 | [34] | |
| miR-223 | RASA1 | Cell proliferation | [40] |
| miR-301a | BTG1 | Promote intestinal inflammation | [41] |
| Downregulation | |||
| miR-34a | IL6/EMT/EGR1 | Suppresses migration and invasion | [34,42] |
| miR-124 | STAT3/ROCK1 | Inhibits neoplastic transformation | [43,44] |
| miR-139-5p | IGF-1R | Maintain intestinal homeostasis | [45,46] |
| miR-185-3p | MLCK | Regulate via lncRNA CCAT1 | [47] |
| miR-193a-3p | SLC15A1 | Suppress NF-κB signaling | [48,49] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bocchetti, M.; Ferraro, M.G.; Ricciardiello, F.; Ottaiano, A.; Luce, A.; Cossu, A.M.; Scrima, M.; Leung, W.-Y.; Abate, M.; Stiuso, P.; et al. The Role of microRNAs in Development of Colitis-Associated Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 3967. https://doi.org/10.3390/ijms22083967
Bocchetti M, Ferraro MG, Ricciardiello F, Ottaiano A, Luce A, Cossu AM, Scrima M, Leung W-Y, Abate M, Stiuso P, et al. The Role of microRNAs in Development of Colitis-Associated Colorectal Cancer. International Journal of Molecular Sciences. 2021; 22(8):3967. https://doi.org/10.3390/ijms22083967
Chicago/Turabian StyleBocchetti, Marco, Maria Grazia Ferraro, Filippo Ricciardiello, Alessandro Ottaiano, Amalia Luce, Alessia Maria Cossu, Marianna Scrima, Wing-Yan Leung, Marianna Abate, Paola Stiuso, and et al. 2021. "The Role of microRNAs in Development of Colitis-Associated Colorectal Cancer" International Journal of Molecular Sciences 22, no. 8: 3967. https://doi.org/10.3390/ijms22083967