
Early Lipid Raft-Related Changes: Interplay between Unilateral Denervation and Hindlimb Suspension


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
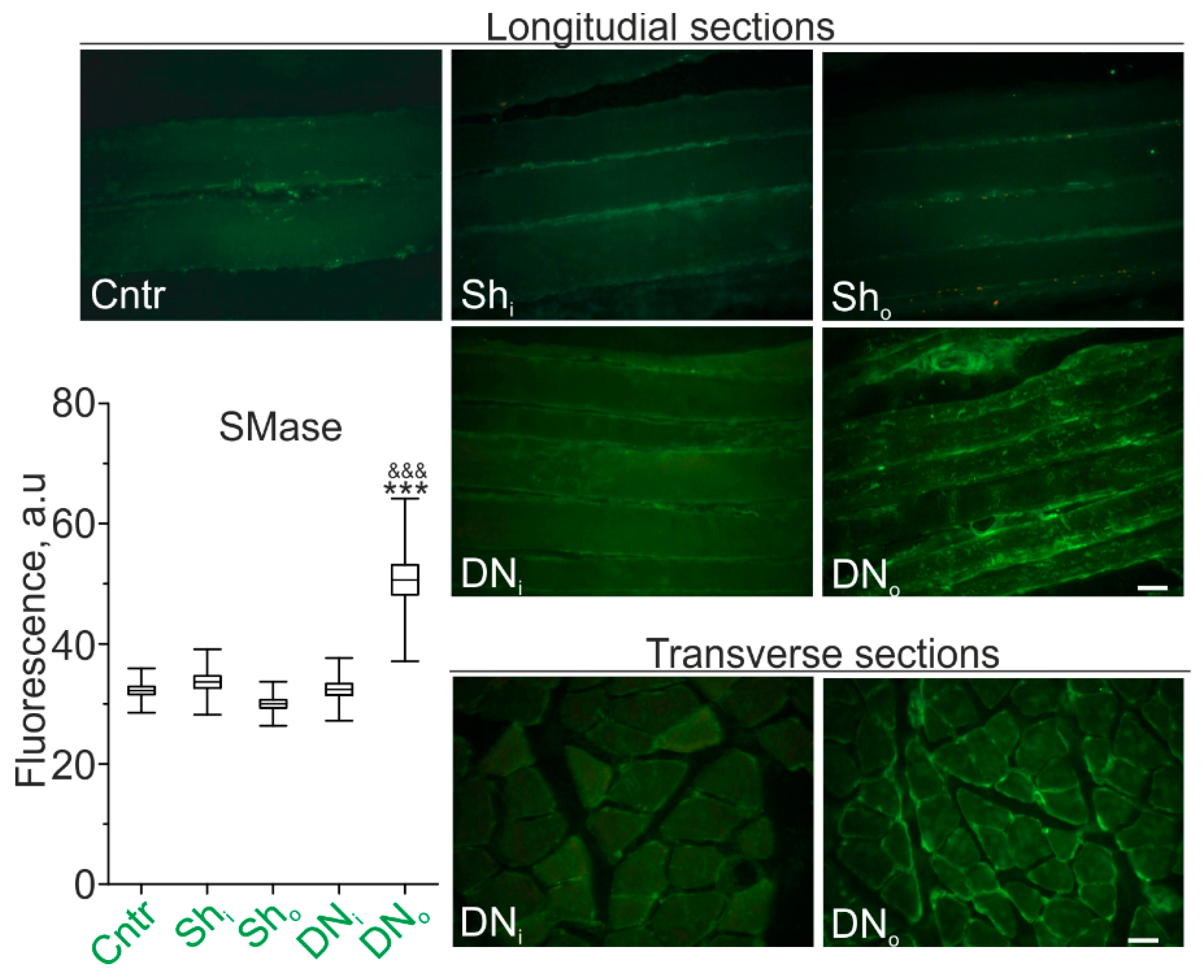
2.1. SMase Immunoexpression in the Soleus Muscle of Control (Sham-Operated Rats) and Denervated Animals
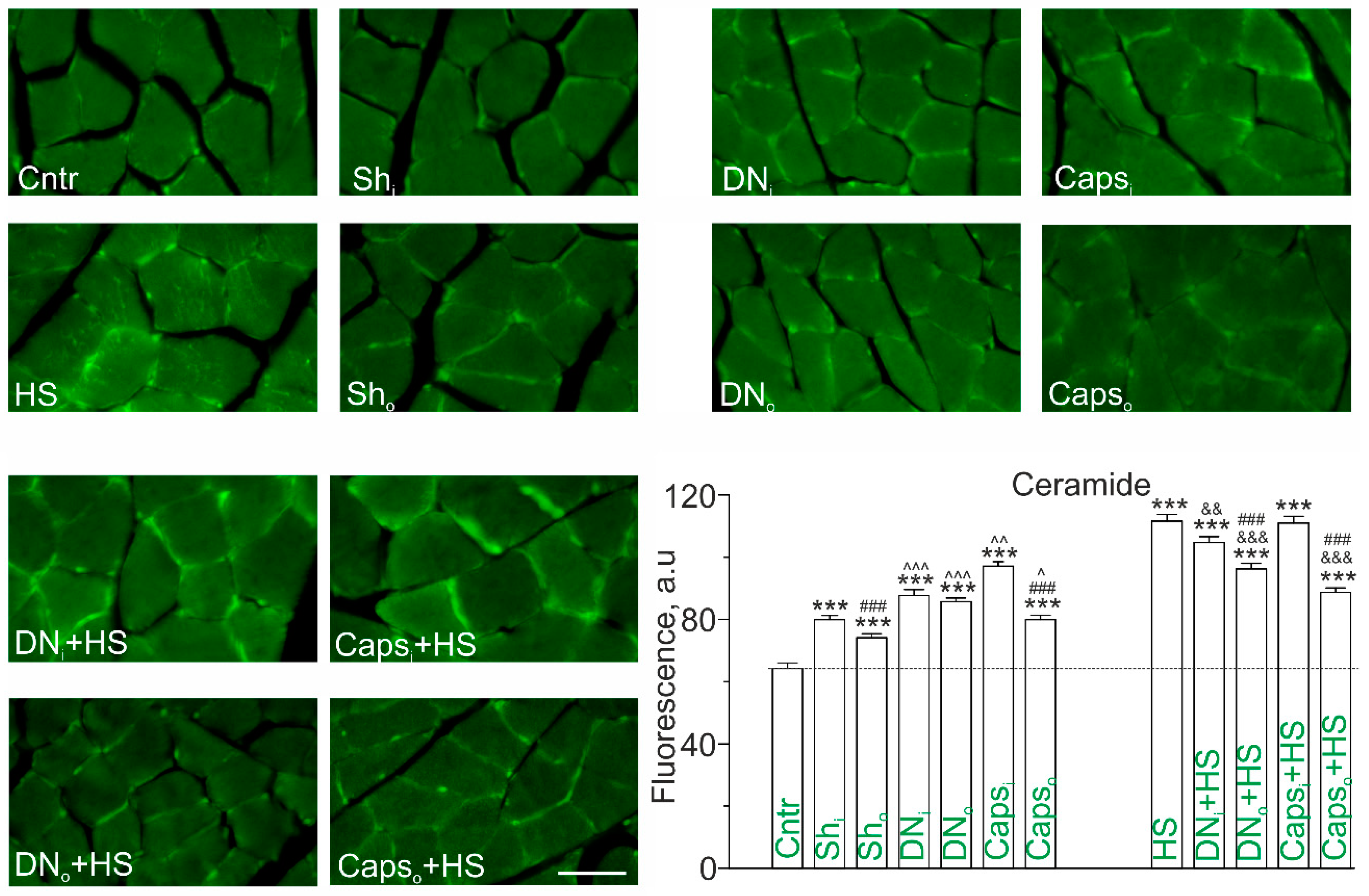
2.2. Immunodetection of Cer on Transverse Sections of Rat Soleus Muscle in HS Combined with Unilateral Denervation or Caps Treatment of Sciatic Nerve
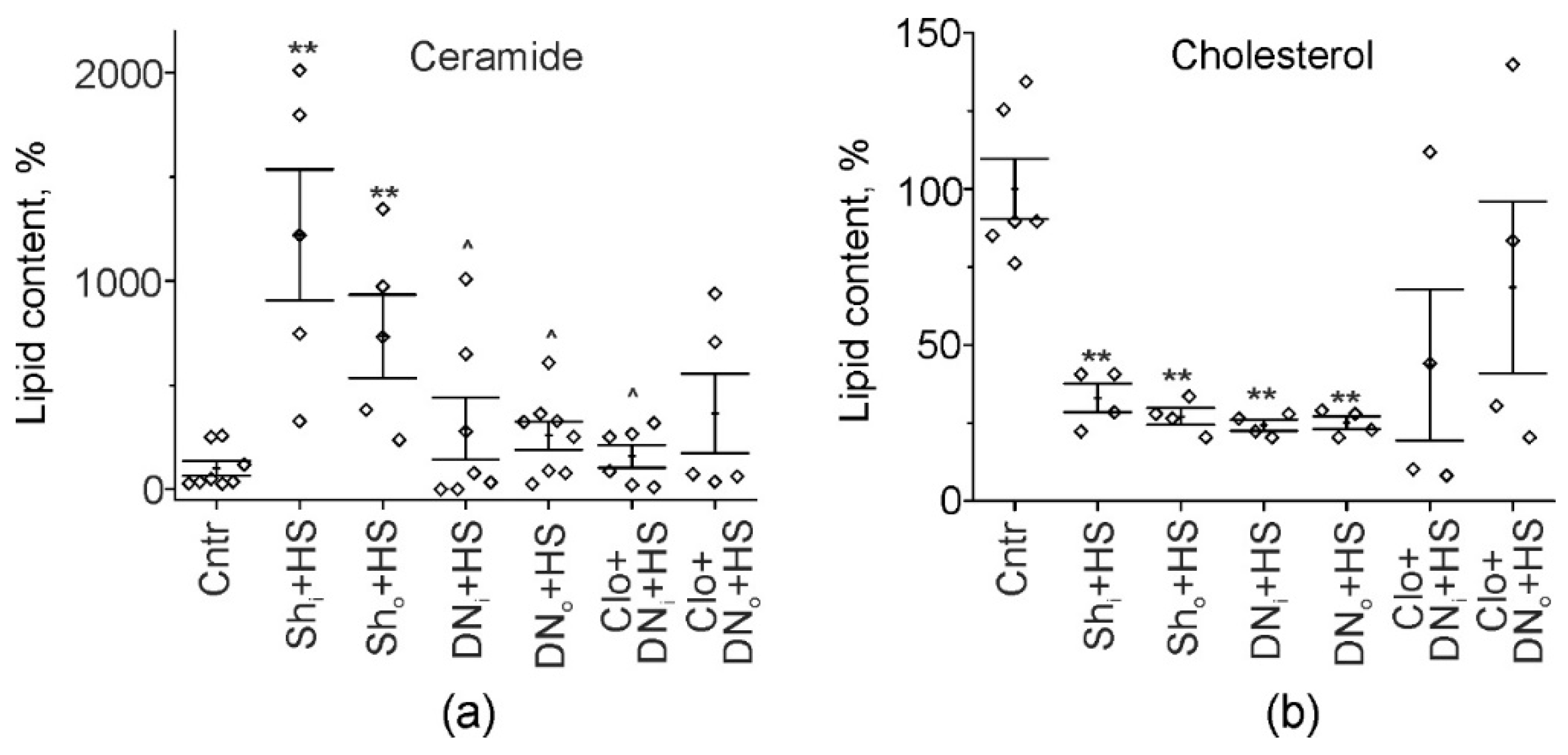
2.3. Cer and Chol Amount in Soleus Muscle Homogenates of Hindlimb Suspended and Unilaterally Denervated Rats
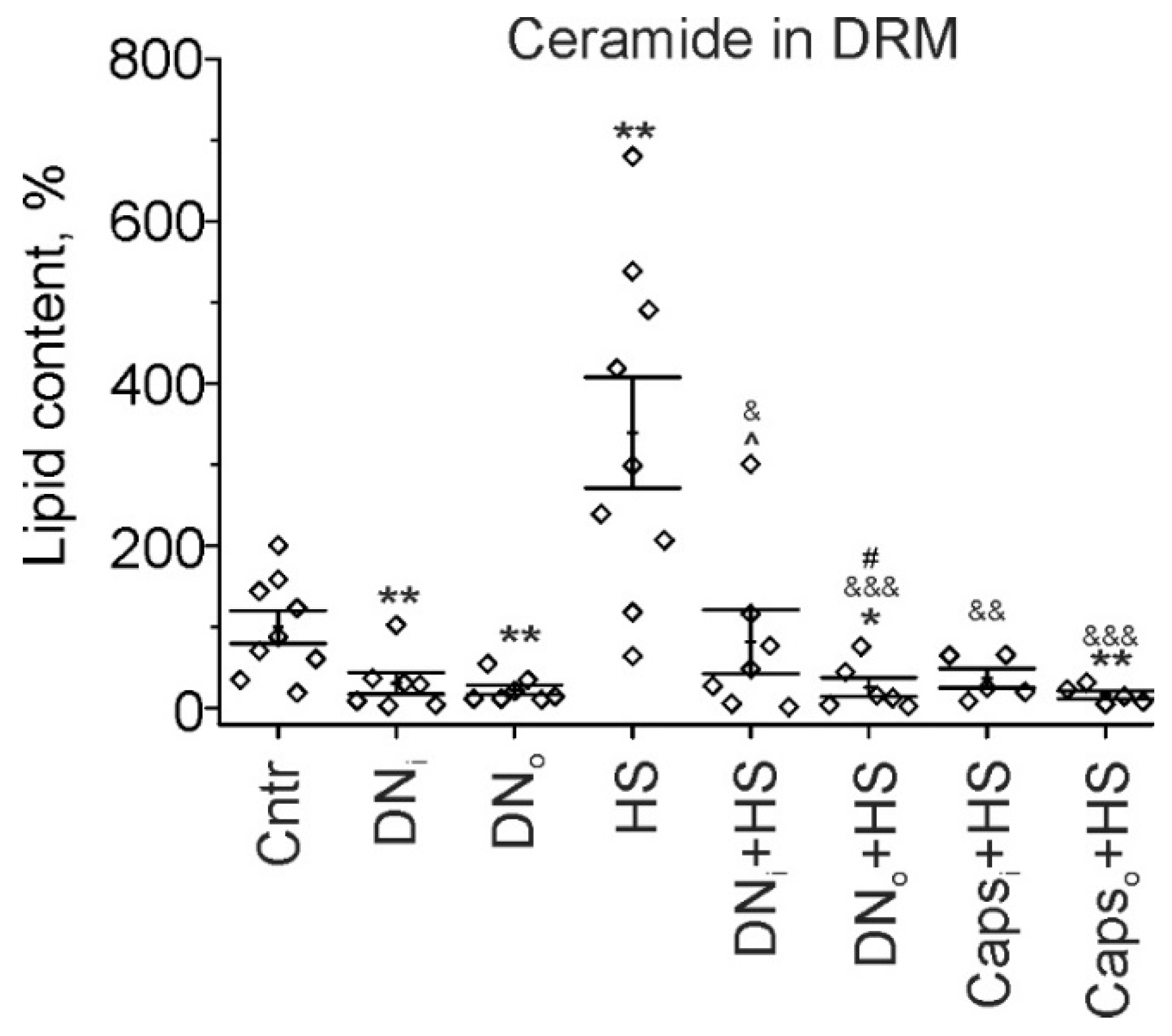
2.4. Cer in Detergent Resistant Membrane (DRM) Domains of Rat Soleus Muscle in HS Combined with Unilaterally Denervation or Caps Treatment of Sciatic Nerve
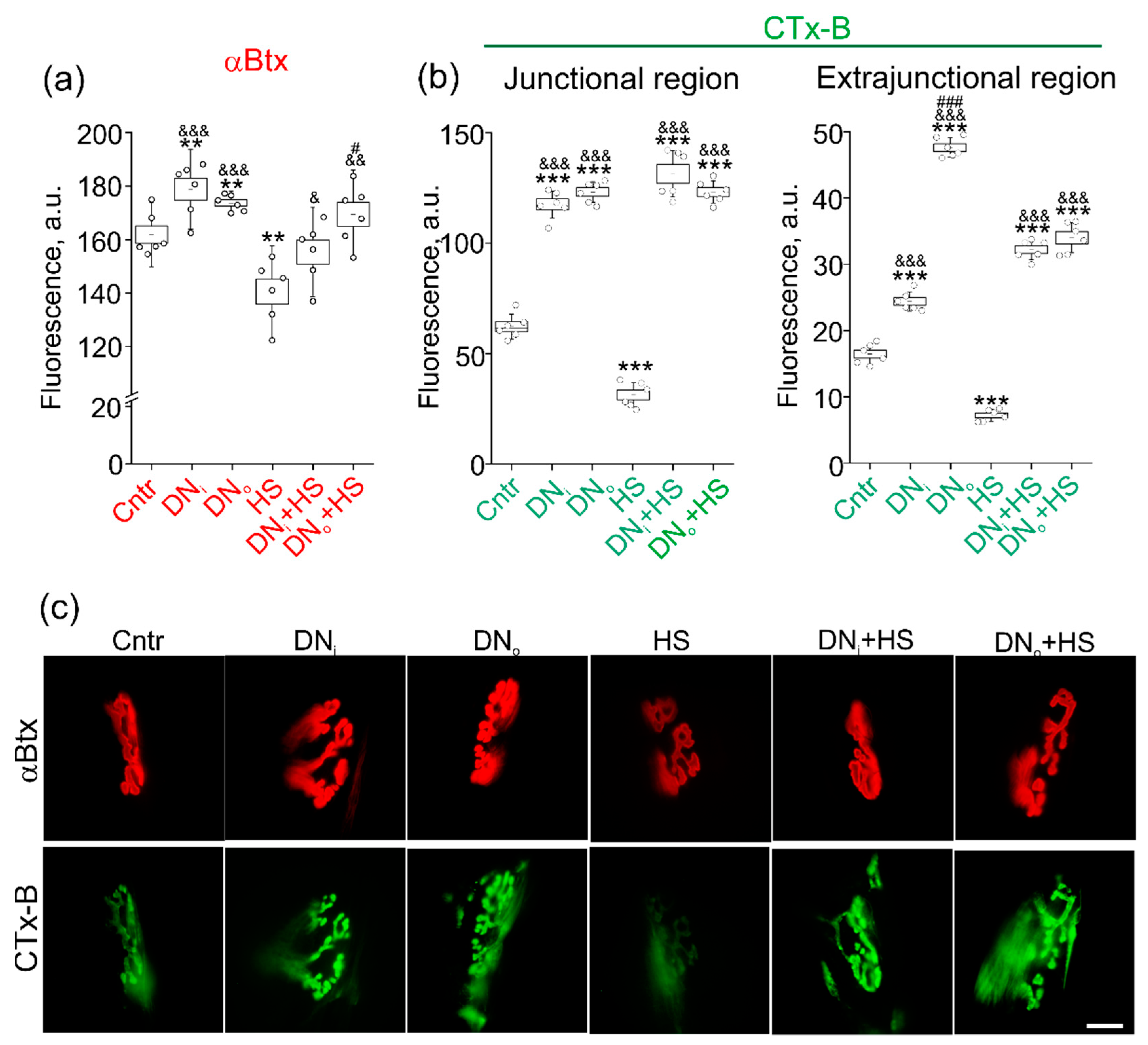
2.5. The Effect of Unilateral Denervation Itself and Its Combination with HS on Staining with Lipid Raft Marker
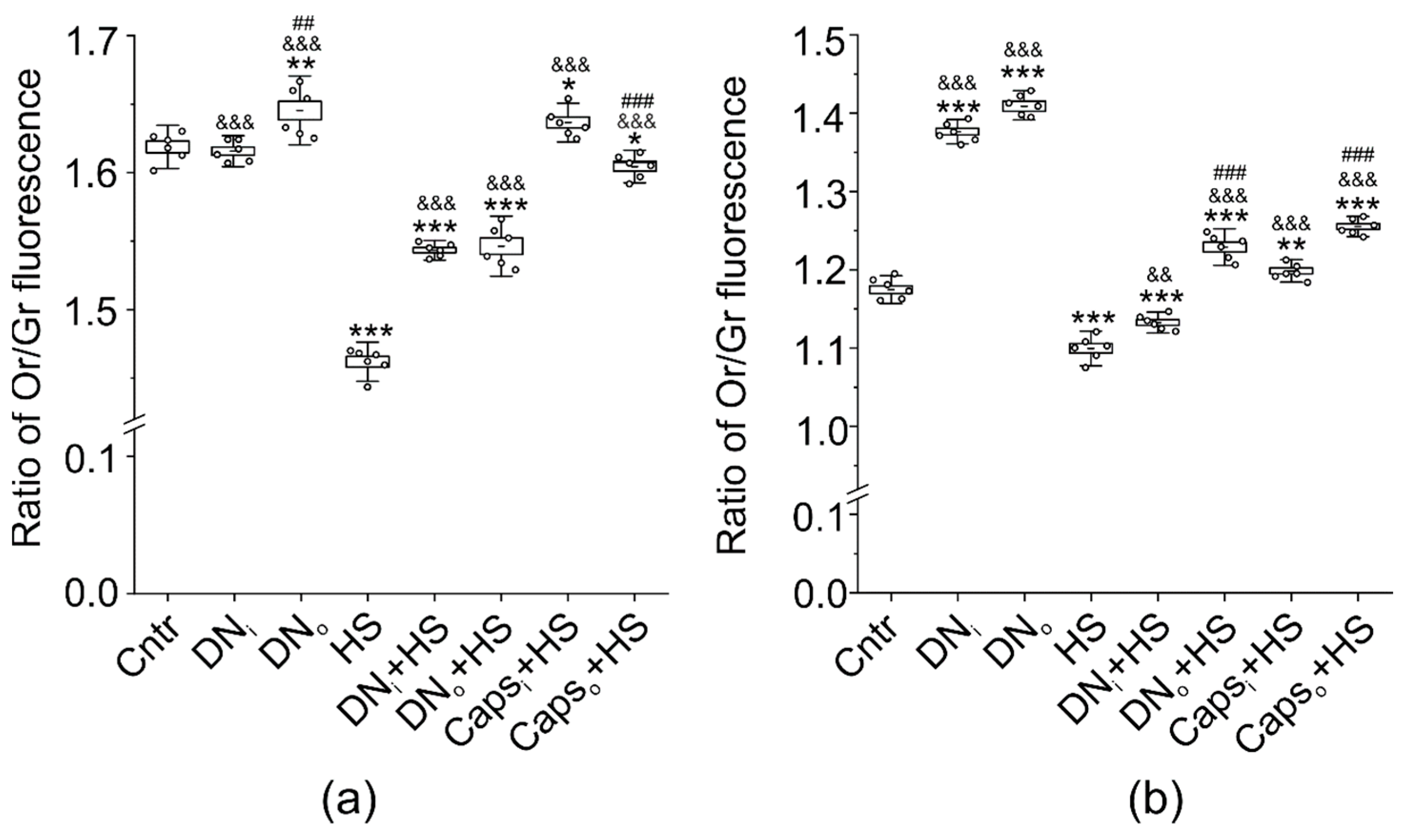
2.6. Influence of Unilateral Denervation and Caps-Induced Sensory Denervation on the Effect of HS on Lipid Distribution-Sensitive Dye F2N12S Fluorescence
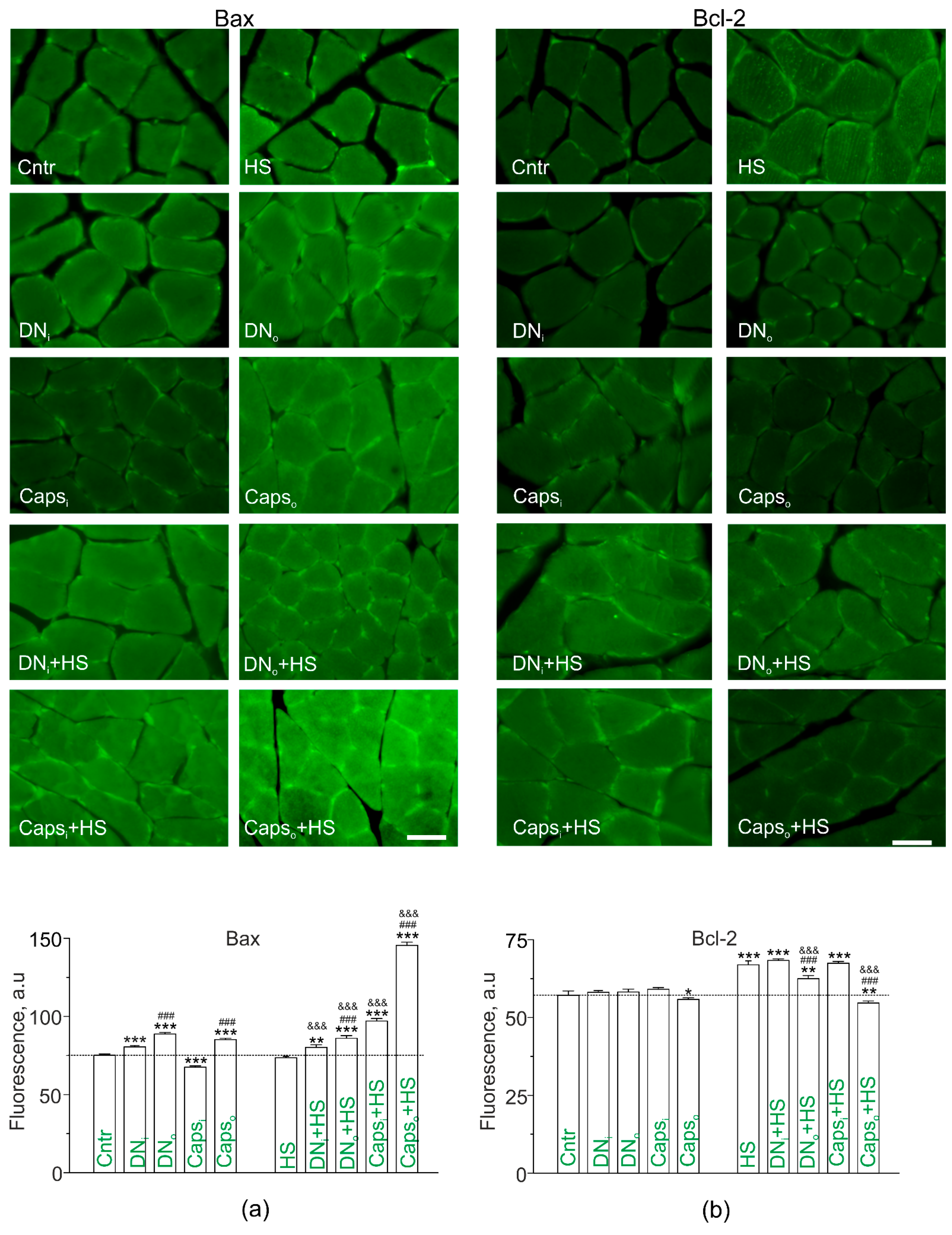
2.7. Study of Bax and Bcl-2 in Disused Soleus Muscles of Rats with Unilateral Surgical Denervation or Caps Treatment of the Sciatic Nerve
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Biochemical Study
4.3. Immunofluorescence Study
4.4. Confocal Fluorescence Microscopy
4.5. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HS | Hindlimb suspension |
| CSPA | Capsaicin-sensitive primary afferents |
| aSMase | Acid sphingomyelinase |
| Caps | Capsaicin |
| Cer | Ceramide |
| Chol | Cholesterol |
| DN | Denervation |
| DRM | Detergent resistant membrane fraction |
| CTx-B | Cholera toxin B subunit |
| α-Btx | α-bungarotoxin |
| TRPV1 | Transient receptor potential cation channel subfamily V member 1 |
References
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. (1985) 2002, 92, 1367–1377. [Google Scholar] [CrossRef]
- Baek, K.W.; Jung, Y.K.; Kim, J.S.; Park, J.S.; Hah, Y.S.; Kim, S.J.; Yoo, J.I. Rodent Model of Muscular Atrophy for Sarcopenia Study. J. Bone Metab. 2020, 27, 97–110. [Google Scholar] [CrossRef]
- MacDonald, E.M.; Andres-Mateos, E.; Mejias, R.; Simmers, J.L.; Mi, R.; Park, J.S.; Ying, S.; Hoke, A.; Lee, S.J.; Cohn, R.D. Denervation atrophy is independent from Akt and mTOR activation and is not rescued by myostatin inhibition. Dis. Model. Mech. 2014, 7, 471–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrov, A.M.; Kravtsova, V.V.; Matchkov, V.V.; Vasiliev, A.N.; Zefirov, A.L.; Chibalin, A.V.; Heiny, J.A.; Krivoi, I.I. Membrane lipid rafts are disturbed in the response of rat skeletal muscle to short-term disuse. Am. J. Physiol. Cell Physiol. 2017, 312, C627–C637. [Google Scholar] [CrossRef]
- Bryndina, I.G.; Shalagina, M.N.; Sekunov, A.V.; Zefirov, A.L.; Petrov, A.M. Clomipramine counteracts lipid raft disturbance due to short-term muscle disuse. Neurosci. Lett. 2018, 664, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kasimov, M.R.; Giniatullin, A.R.; Zefirov, A.L.; Petrov, A.M. Effects of 5alpha-cholestan-3-one on the synaptic vesicle cycle at the mouse neuromuscular junction. Biochim. Biophys. Acta. 2015, 1851, 674–685. [Google Scholar] [CrossRef] [PubMed]
- Stetzkowski-Marden, F.; Recouvreur, M.; Camus, G.; Cartaud, A.; Marchand, S.; Cartaud, J. Rafts are required for acetylcholine receptor clustering. J. Mol. Neurosci. 2006, 30, 37–38. [Google Scholar] [CrossRef]
- Krivoi, I.I.; Petrov, A.M. Cholesterol and the Safety Factor for Neuromuscular Transmission. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega-Moreno, J.; Tirado-Cortes, A.; Alvarez, R.; Irles, C.; Mas-Oliva, J.; Ortega, A. Cholesterol depletion uncouples beta-dystroglycans from discrete sarcolemmal domains, reducing the mechanical activity of skeletal muscle. Cell Physiol. Biochem. 2012, 29, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Gazzerro, E.; Bonetto, A.; Minetti, C. Caveolinopathies: Translational implications of caveolin-3 in skeletal and cardiac muscle disorders. Handb. Clin. Neurol. 2011, 101, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Bryndina, I.G.; Shalagina, M.N.; Ovechkin, S.V.; Ovchinina, N.G. Sphingolipids in skeletal muscles of C57B1/6 mice after short-term simulated microgravity. Ross. Fiziol. Zh. Im. I. M. Sechenova 2014, 100, 1280–1286. [Google Scholar]
- Salaun, E.; Lefeuvre-Orfila, L.; Cavey, T.; Martin, B.; Turlin, B.; Ropert, M.; Loreal, O.; Derbre, F. Myriocin prevents muscle ceramide accumulation but not muscle fiber atrophy during short-term mechanical unloading. J. Appl. Physiol. (1985) 2016, 120, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Kwon, O.S.; Tanner, R.E.; Barrows, K.M.; Runtsch, M.; Symons, J.D.; Jalili, T.; Bikman, B.T.; McClain, D.A.; O’Connell, R.M.; Drummond, M.J. MyD88 regulates physical inactivity-induced skeletal muscle inflammation, ceramide biosynthesis signaling, and glucose intolerance. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E11–E21. [Google Scholar] [CrossRef] [PubMed]
- Petrov, A.M.; Shalagina, M.N.; Protopopov, V.A.; Sergeev, V.G.; Ovechkin, S.V.; Ovchinina, N.G.; Sekunov, A.V.; Zefirov, A.L.; Zakirjanova, G.F.; Bryndina, I.G. Changes in Membrane Ceramide Pools in Rat Soleus Muscle in Response to Short-Term Disuse. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryndina, I.; Protopopov, V.; Sergeev, V.; Shalagina, M.; Ovechkin, S.; Yakovlev, A. Ceramide Enriched Membrane Domains in Rat Skeletal Muscle Exposed to Short-Term Hypogravitational Unloading. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Bandet, C.L.; Tan-Chen, S.; Bourron, O.; Le Stunff, H.; Hajduch, E. Sphingolipid Metabolism: New Insight into Ceramide-Induced Lipotoxicity in Muscle Cells. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- London, E. Ceramide selectively displaces cholesterol from ordered lipid domains (rafts): Implications for lipid raft structure and function. J. Biol. Chem. 2004, 279, 9997–10004. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Alterman, M.; Dobrowsky, R.T. Ceramide displaces cholesterol from lipid rafts and decreases the association of the cholesterol binding protein caveolin-1. J. Lipid. Res. 2005, 46, 1678–1691. [Google Scholar] [CrossRef] [Green Version]
- Turinsky, J.; Bayly, B.P.; O’Sullivan, D.M. 1,2-Diacylglycerol and ceramide levels in rat skeletal muscle and liver in vivo. Studies with insulin, exercise, muscle denervation, and vasopressin. J. Biol. Chem. 1990, 265, 7933–7938. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kim, W.J.; Lee, M.H.; Kim, S.Y.; Seo, D.H.; Kim, H.S.; Gelinsky, M.; Kim, T.J. Anti-skeletal muscle atrophy effect of Oenothera odorata root extract via reactive oxygen species-dependent signaling pathways in cellular and mouse model. Biosci. Biotechnol. Biochem. 2016, 80, 80–88. [Google Scholar] [CrossRef] [Green Version]
- Henriques, A.; Croixmarie, V.; Priestman, D.A.; Rosenbohm, A.; Dirrig-Grosch, S.; D’Ambra, E.; Huebecker, M.; Hussain, G.; Boursier-Neyret, C.; Echaniz-Laguna, A.; et al. Amyotrophic lateral sclerosis and denervation alter sphingolipids and up-regulate glucosylceramide synthase. Hum. Mol. Genet. 2015, 24, 7390–7405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tischler, M.E.; Satarug, S.; Eisenfeld, S.H.; Henriksen, E.J.; Rosenberg, S.B. Insulin effects in denervated and non-weight-bearing rat soleus muscle. Muscle Nerve 1990, 13, 593–600. [Google Scholar] [CrossRef]
- Kujawa, M.; Baran, W.; Jankowska-Steifer, E. Quantitative ultrastructural changes in satellite cells of rats immobilized after soleus muscle denervation. Exp. Mol. Pathol. 2005, 78, 78–85. [Google Scholar] [CrossRef]
- di Maso, N.A.; Haddad, F.; Zeng, M.; McCue, S.A.; Baldwin, K.M. Role of denervation in modulating IIb MHC gene expression in response to T(3) plus unloading state. J. Appl. Physiol. (1985) 2000, 88, 682–689. [Google Scholar] [CrossRef] [Green Version]
- Ohira, Y.; Yoshinaga, T.; Ohara, M.; Kawano, F.; Wang, X.D.; Higo, Y.; Terada, M.; Matsuoka, Y.; Roy, R.R.; Edgerton, V.R. The role of neural and mechanical influences in maintaining normal fast and slow muscle properties. Cells Tissues Organs 2006, 182, 129–142. [Google Scholar] [CrossRef] [PubMed]
- 2Picquet, F.; Falempin, M. Compared effects of hindlimb unloading versus terrestrial deafferentation on muscular properties of the rat soleus. Exp. Neurol. 2003, 182, 186–194. [Google Scholar] [CrossRef]
- Nemirovskaya, T.L.; Shenkman, B.S.; Mukhina, A.M.; Volodkovich, Y.Y.; Sayapina, M.M.; Brattseva, E.; Larina, O. Effects of deafferentation on the size and myosin phenotype of muscle fibers on stretching of the rat soleus muscle in conditions of gravitational unloading. Neurosci. Behav. Physiol. 2004, 34, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Szolcsanyi, J. Capsaicin and sensory neurones: A historical perspective. Prog. Drug Res. 2014, 68, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Saghy, E.; Szoke, E.; Payrits, M.; Helyes, Z.; Borzsei, R.; Erostyak, J.; Janosi, T.Z.; Setalo, G., Jr.; Szolcsanyi, J. Evidence for the role of lipid rafts and sphingomyelin in Ca2+-gating of Transient Receptor Potential channels in trigeminal sensory neurons and peripheral nerve terminals. Pharmacol. Res. 2015, 100, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Petrov, A.M.; Yakovleva, A.A.; Zefirov, A.L. Role of membrane cholesterol in spontaneous exocytosis at frog neuromuscular synapses: Reactive oxygen species-calcium interplay. J. Physiol. 2014, 592, 4995–5009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabados, T.; Gomori, K.; Palvolgyi, L.; Gorbe, A.; Baczko, I.; Helyes, Z.; Jancso, G.; Ferdinandy, P.; Bencsik, P. Capsaicin-Sensitive Sensory Nerves and the TRPV1 Ion Channel in Cardiac Physiology and Pathologies. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Jancso, G.; Kiraly, E.; Jancso-Gabor, A. Pharmacologically induced selective degeneration of chemosensitive primary sensory neurones. Nature 1977, 270, 741–743. [Google Scholar] [CrossRef]
- Szolcsanyi, J. Forty years in capsaicin research for sensory pharmacology and physiology. Neuropeptides 2004, 38, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Pettorossi, V.E.; Bortolami, R.; Della Torre, G.; Brunetti, O. Effects of capsaicin in the motor nerve. Exp. Neurol. 1994, 128, 284–289. [Google Scholar] [CrossRef]
- Kaufman, M.P.; Longhurst, J.C.; Rybicki, K.J.; Wallach, J.H.; Mitchell, J.H. Effects of static muscular contraction on impulse activity of groups III and IV afferents in cats. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1983, 55, 105–112. [Google Scholar] [CrossRef]
- Brunetti, O.; Barazzoni, A.M.; Della Torre, G.; Clavenzani, P.; Pettorossi, V.E.; Bortolami, R. Partial transformation from fast to slow muscle fibers induced by deafferentation of capsaicin-sensitive muscle afferents. Muscle Nerve 1997, 20, 1404–1413. [Google Scholar] [CrossRef]
- Helyes, Z.; Nemeth, J.; Than, M.; Bolcskei, K.; Pinter, E.; Szolcsanyi, J. Inhibitory effect of anandamide on resiniferatoxin-induced sensory neuropeptide release in vivo and neuropathic hyperalgesia in the rat. Life Sci. 2003, 73, 2345–2353. [Google Scholar] [CrossRef]
- Russell, F.A.; King, R.; Smillie, S.J.; Kodji, X.; Brain, S.D. Calcitonin gene-related peptide: Physiology and pathophysiology. Physiol. Rev. 2014, 94, 1099–1142. [Google Scholar] [CrossRef] [Green Version]
- Helyes, Z.; Pinter, E.; Sandor, K.; Elekes, K.; Banvolgyi, A.; Keszthelyi, D.; Szoke, E.; Toth, D.M.; Sandor, Z.; Kereskai, L.; et al. Impaired defense mechanism against inflammation, hyperalgesia, and airway hyperreactivity in somatostatin 4 receptor gene-deleted mice. Proc. Natl. Acad. Sci. USA 2009, 106, 13088–13093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- New, H.V.; Mudge, A.W. Calcitonin gene-related peptide regulates muscle acetylcholine receptor synthesis. Nature 1986, 323, 809–811. [Google Scholar] [CrossRef]
- Giniatullin, A.; Petrov, A.; Giniatullin, R. The involvement of P2Y12 receptors, NADPH oxidase, and lipid rafts in the action of extracellular ATP on synaptic transmission at the frog neuromuscular junction. Neuroscience 2015, 285, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Petrov, A.M.; Kasimov, M.R.; Giniatullin, A.R.; Tarakanova, O.I.; Zefirov, A.L. The role of cholesterol in the exo- and endocytosis of synaptic vesicles in frog motor nerve endings. Neurosci. Behav. Physiol. 2010, 40, 894–901. [Google Scholar] [CrossRef]
- Kravtsova, V.V.; Petrov, A.M.; Vasil’ev, A.N.; Zefirov, A.L.; Krivoi, I.I. Role of cholesterol in the maintenance of endplate electrogenesis in rat diaphragm. Bull. Exp. Biol. Med. 2015, 158, 298–300. [Google Scholar] [CrossRef]
- Petrov, A.M.; Naumenko, N.V.; Uzinskaya, K.V.; Giniatullin, A.R.; Urazaev, A.K.; Zefirov, A.L. Increased non-quantal release of acetylcholine after inhibition of endocytosis by methyl-beta-cyclodextrin: The role of vesicular acetylcholine transporter. Neuroscience 2011, 186, 1–12. [Google Scholar] [CrossRef]
- Mukhutdinova, K.A.; Kasimov, M.R.; Giniatullin, A.R.; Zakyrjanova, G.F.; Petrov, A.M. 24S-hydroxycholesterol suppresses neuromuscular transmission in SOD1(G93A) mice: A possible role of NO and lipid rafts. Mol. Cell. Neurosci. 2018, 88, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Kasimov, M.R.; Zakyrjanova, G.F.; Giniatullin, A.R.; Zefirov, A.L.; Petrov, A.M. Similar oxysterols may lead to opposite effects on synaptic transmission: Olesoxime versus 5alpha-cholestan-3-one at the frog neuromuscular junction. Biochim. Biophys. Acta 2016, 1861, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Shynkar, V.V.; Klymchenko, A.S.; Kunzelmann, C.; Duportail, G.; Muller, C.D.; Demchenko, A.P.; Freyssinet, J.M.; Mely, Y. Fluorescent biomembrane probe for ratiometric detection of apoptosis. J. Am. Chem. Soc. 2007, 129, 2187–2193. [Google Scholar] [CrossRef] [PubMed]
- Petrov, A.M.; Mast, N.; Li, Y.; Denker, J.; Pikuleva, I.A. Brain sterol flux mediated by cytochrome P450 46A1 affects membrane properties and membrane-dependent processes. Brain Commun. 2020, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannion, R.J.; Doubell, T.P.; Coggeshall, R.E.; Woolf, C.J. Collateral sprouting of uninjured primary afferent A-fibers into the superficial dorsal horn of the adult rat spinal cord after topical capsaicin treatment to the sciatic nerve. J. Neurosci. 1996, 16, 5189–5195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitts, R.H.; Riley, D.R.; Widrick, J.J. Physiology of a microgravity environment invited review: Microgravity and skeletal muscle. J. Appl. Physiol. (1985) 2000, 89, 823–839. [Google Scholar] [CrossRef] [Green Version]
- Adams, G.R.; Caiozzo, V.J.; Baldwin, K.M. Skeletal muscle unweighting: Spaceflight and ground-based models. J. Appl. Physiol. (1985) 2003, 95, 2185–2201. [Google Scholar] [CrossRef] [Green Version]
- Tan-Chen, S.; Guitton, J.; Bourron, O.; Le Stunff, H.; Hajduch, E. Sphingolipid Metabolism and Signaling in Skeletal Muscle: From Physiology to Physiopathology. Front. Endocrinol. 2020, 11, 491. [Google Scholar] [CrossRef] [PubMed]
- Shenkman, B.S.; Kozlovskaya, I.B. Cellular Responses of Human Postural Muscle to Dry Immersion. Front. Physiol. 2019, 10, 187. [Google Scholar] [CrossRef]
- Garcia-Ruiz, C.; Morales, A.; Fernandez-Checa, J.C. Glycosphingolipids and cell death: One aim, many ways. Apoptosis 2015, 20, 607–620. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, Y.; Kohyama-Koganeya, A.; Hirabayashi, Y. New insights on glucosylated lipids: Metabolism and functions. Biochim. Biophys. Acta 2013, 1831, 1475–1485. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, A.; Hall, P.; Wall, P.D.; Allt, G.; MacKenzie, M.L.; Gibson, S.; Polak, J.M. Effects of capsaicin applied locally to adult peripheral nerve. II. Anatomy and enzyme and peptide chemistry of peripheral nerve and spinal cord. Pain 1981, 11, 379–388. [Google Scholar] [CrossRef]
- Ilin, E.A.; Novikov, V.E. Stand for modelling the physiological effects of weightlessness in laboratory experiments with rats. Kosm. Biol. Aviakosm. Med. 1980, 14, 79–80. [Google Scholar] [PubMed]
- Radeva, G.; Sharom, F.J. Isolation and characterization of lipid rafts with different properties from RBL-2H3 (rat basophilic leukaemia) cells. Biochem. J. 2004, 380, 219–230. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Higgins, J.A.; Fieldsend, J.K. Phosphatidylcholine synthesis for incorporation into membranes or for secretion as plasma lipoproteins by Golgi membranes of rat liver. J. Lipid. Res. 1987, 28, 268–278. [Google Scholar] [CrossRef]
- Boath, A.; Graf, C.; Lidome, E.; Ullrich, T.; Nussbaumer, P.; Bornancin, F. Regulation and traffic of ceramide 1-phosphate produced by ceramide kinase: Comparative analysis to glucosylceramide and sphingomyelin. J. Biol. Chem. 2008, 283, 8517–8526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudnick, J.; Puttmann, B.; Tesch, P.A.; Alkner, B.; Schoser, B.G.; Salanova, M.; Kirsch, K.; Gunga, H.C.; Schiffl, G.; Luck, G.; et al. Differential expression of nitric oxide synthases (NOS 1-3) in human skeletal muscle following exercise countermeasure during 12 weeks of bed rest. FASEB J. 2004, 18, 1228–1230. [Google Scholar] [CrossRef] [PubMed]
- Margheri, G.; D’Agostino, R.; Trigari, S.; Sottini, S.; Del Rosso, M. The beta-subunit of cholera toxin has a high affinity for ganglioside GM1 embedded into solid supported lipid membranes with a lipid raft-like composition. Lipids 2014, 49, 203–206. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryndina, I.G.; Shalagina, M.N.; Protopopov, V.A.; Sekunov, A.V.; Zefirov, A.L.; Zakirjanova, G.F.; Petrov, A.M. Early Lipid Raft-Related Changes: Interplay between Unilateral Denervation and Hindlimb Suspension. Int. J. Mol. Sci. 2021, 22, 2239. https://doi.org/10.3390/ijms22052239
Bryndina IG, Shalagina MN, Protopopov VA, Sekunov AV, Zefirov AL, Zakirjanova GF, Petrov AM. Early Lipid Raft-Related Changes: Interplay between Unilateral Denervation and Hindlimb Suspension. International Journal of Molecular Sciences. 2021; 22(5):2239. https://doi.org/10.3390/ijms22052239
Chicago/Turabian StyleBryndina, Irina G., Maria N. Shalagina, Vladimir A. Protopopov, Alexey V. Sekunov, Andrey L. Zefirov, Guzalia F. Zakirjanova, and Alexey M. Petrov. 2021. "Early Lipid Raft-Related Changes: Interplay between Unilateral Denervation and Hindlimb Suspension" International Journal of Molecular Sciences 22, no. 5: 2239. https://doi.org/10.3390/ijms22052239