Extracellular Vesicles from Follicular and Ampullary Fluid Isolated by Density Gradient Ultracentrifugation Improve Bovine Embryo Development and Quality

 ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
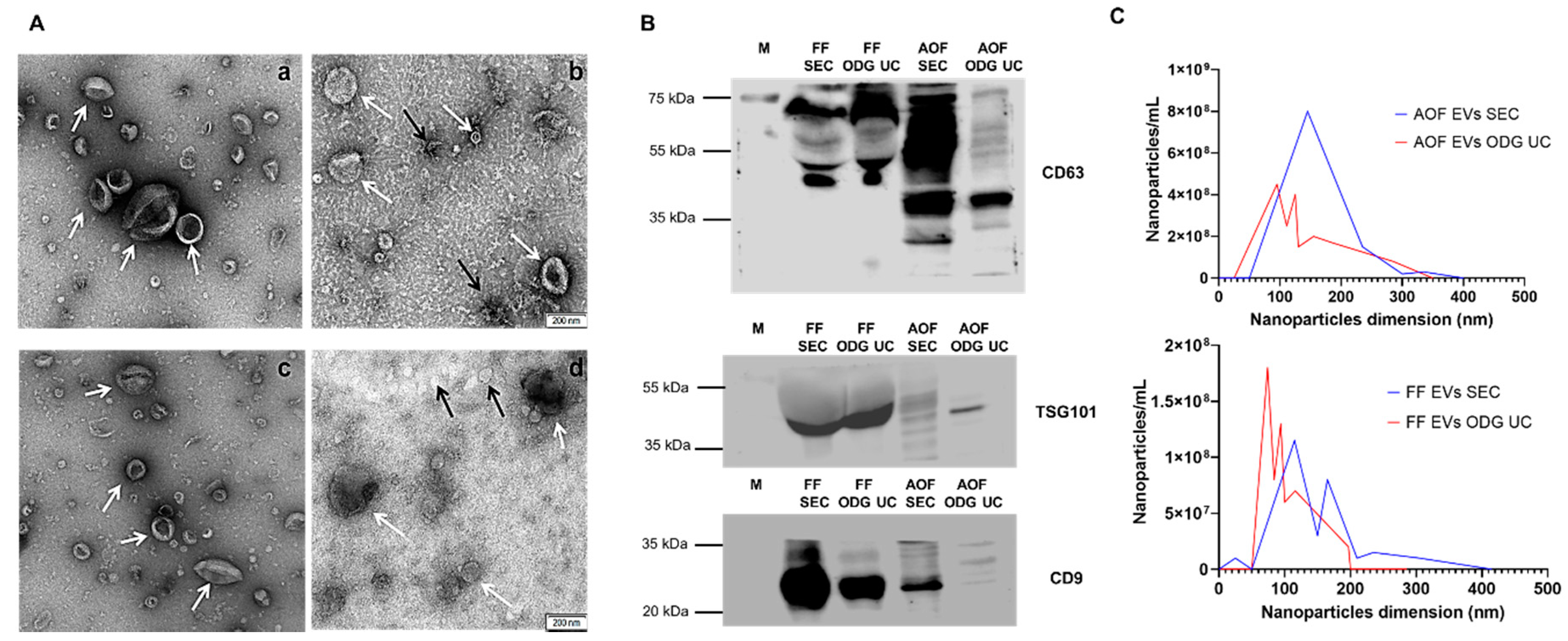
2.1. Equivalent Efficiency of OptiPrep™ Density Gradient Ultracentrifugation and Single-Step Size Exclusion Chromatography (qEV IZON™ Single Column) in Isolating Extracellular Vesicles
2.2. Higher Functionality of Extracellular Vesicles Isolated Using OptiPrep™ Density Gradient Ultracentrifugation Compared to Single-Step Size Exclusion Chromatography (qEV IZON™ Single Column)
2.2.1. Embryo Development
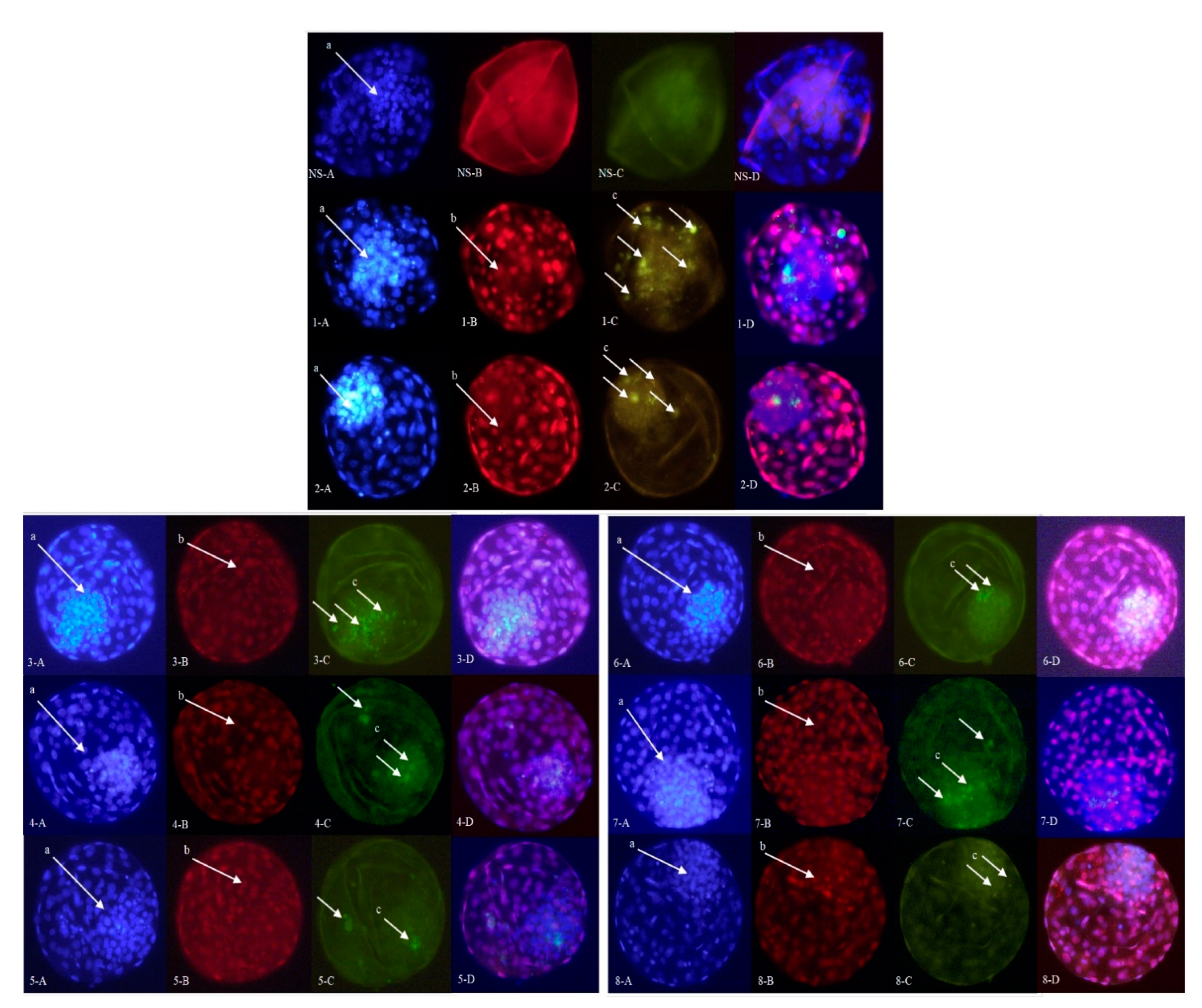
2.2.2. Embryo Quality
3. Discussion
4. Materials and Methods
4.1. Media and Reagents
4.2. Animal Handling, Bovine Follicular, and Ampullary Oviductal Fluid Collection
4.3. Extracellular Vesicles Isolation by OptiPrepTM Density Gradient Ultracentrifuge
4.4. Extracellular Vesicles Isolation by Single-Step Size Exclusion Chromatography
4.5. Characterization of Extracellular Vesicles
4.5.1. Transmission Electron Microscopy
4.5.2. Nanoparticle Tracking Analysis
4.5.3. Western Blotting
4.5.4. EV TRACK
4.6. In Vitro Embryo Production
4.7. Assessment of Embryo Development and Quality
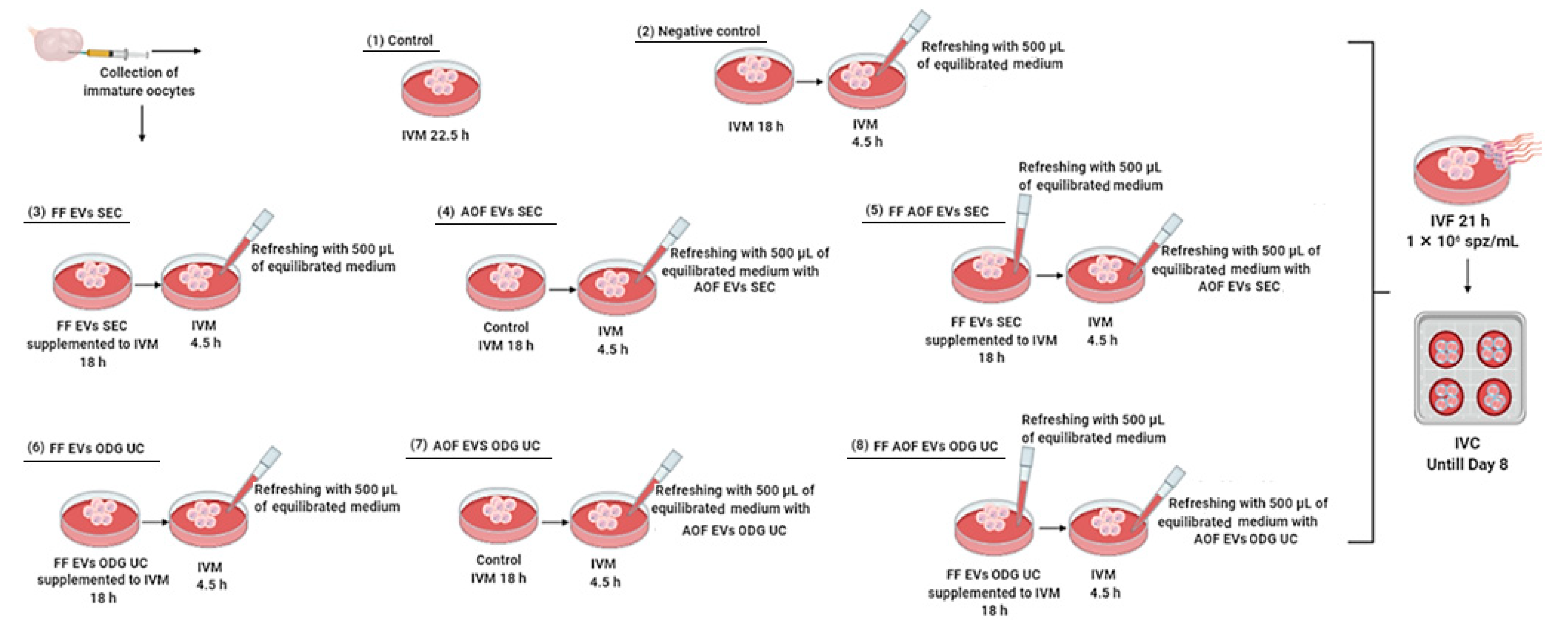
4.8. Experimental Design
4.8.1. Quality and Efficiency Assessment of Two Methods for Isolation of Extracellular Vesicles
4.8.2. Assessing Functionality of Extracellular Vesicles Derived from Follicular and Ampullary Oviductal Fluid during In Vitro Maturation
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EVs | Extracellular vesicles |
| FF | Follicular fluid |
| AOF | Ampullary oviduct fluid |
| ODG UC | OptiPrep™ density gradient ultracentrifugation |
| SEC | Size exclusion chromatography |
| ICM | Inner cell mass |
| TE | Trophectoderm |
| TCN | Total cell number |
| ACR | Apoptotic cell ratio |
| NTA | Nanoparticle tracking analysis |
| WB | Western blot |
| hpi | Hours post-insemination |
References
- Rizos, D.; Clemente, M.; Bermejo-Alvarez, P.; De La Fuente, J.; Lonergan, P.; Gutiérrez-Adán, A. Consequences of In Vitro Culture Conditions on Embryo Development and Quality. Reprod. Domest. Anim. 2008, 43, 44–50. [Google Scholar] [CrossRef]
- Lonergan, P.; Fair, T. In vitro-produced bovine embryos—Dealing with the warts. Theriogenology 2008, 69, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Sirard, M.-A.; Richard, F.; Blondin, P.; Robert, C. Contribution of the oocyte to embryo quality. Theriogenology 2006, 65, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Matzuk, M.M.; Burns, K.H.; Viveiros, M.M.; Eppig, J.J. Intercellular Communication in the Mammalian Ovary: Oocytes Carry the Conversation. Science 2002, 296, 2178. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, R.J.; Irving-Rodgers, H.F. Formation of the Ovarian Follicular Antrum and Follicular Fluid1. Biol. Reprod. 2010, 82, 1021–1029. [Google Scholar] [CrossRef] [Green Version]
- Albertini, D.F.; Combelles, C.; Benecchi, E.; Carabatsos, M.J. Cellular basis for paracrine regulation of ovarian follicle development. Reproduction 2001, 121, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, J.C.; Veeramachaneni, D.N.R.; Winger, Q.A.; Carnevale, E.M.; Bouma, G.J. Cell-Secreted Vesicles in Equine Ovarian Follicular Fluid Contain miRNAs and Proteins: A Possible New Form of Cell Communication Within the Ovarian Follicle1. Biol. Reprod. 2012, 86, 71. [Google Scholar] [CrossRef] [PubMed]
- Lopera-Vasquez, R.; Hamdi, M.; Maillo, V.; Gutierrez-Adan, A.; Bermejo-Alvarez, P.; Ramírez, M.Á.; Yáñez-Mó, M.; Rizos, D. Effect of bovine oviductal extracellular vesicles on embryo development and quality in vitro. Reproduction 2017, 153, 461–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373. [Google Scholar] [CrossRef] [Green Version]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [Green Version]
- Alenquer, M.; Amorim, M.J. Exosome Biogenesis, Regulation, and Function in Viral Infection. Viruses 2015, 7, 5066–5083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavani, K.C.; Hendrix, A.; Van Den Broeck, W.; Couck, L.; Szymanska, K.; Lin, X.; De Koster, J.; Van Soom, A.; Leemans, B. Isolation and Characterization of Functionally Active Extracellular Vesicles from Culture Medium Conditioned by Bovine Embryos In Vitro. Int. J. Mol. Sci. 2018, 20, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohel, M.M.H.; Hoelker, M.; Noferesti, S.S.; Salilew-Wondim, D.; Tholen, E.; Looft, C.; Rings, F.; Uddin, M.J.; Spencer, T.E.; Schellander, K.; et al. Exosomal and Non-Exosomal Transport of Extra-Cellular microRNAs in Follicular Fluid: Implications for Bovine Oocyte Developmental Competence. PLoS ONE 2013, 8, e78505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, W.-T.; Hong, X.; Christenson, L.K.; McGinnis, L.K. Extracellular Vesicles from Bovine Follicular Fluid Support Cumulus Expansion. Biol. Reprod. 2015, 93, 117. [Google Scholar] [CrossRef] [PubMed]
- Hung, W.-T.; Navakanitworakul, R.; Khan, T.; Zhang, P.; Davis, J.S.; McGinnis, L.K.; Christenson, L.K. Stage-specific follicular extracellular vesicle uptake and regulation of bovine granulosa cell proliferation. Biol. Reprod. 2017, 97, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, J.C.; Andrade, G.M.; del Collado, M.; Sampaio, R.V.; Sangalli, J.R.; Silva, L.A.; Pinaffi, F.V.L.; Jardim, I.B.; Cesar, M.C.; Nogueira, M.F.G.; et al. Supplementation with small-extracellular vesicles from ovarian follicular fluid during in vitro production modulates bovine embryo development. PLoS ONE 2017, 12, e0179451. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, Y.; Onuma, A.; Fujioka, Y.A.; Yasuhara, K.; Fujii, W.; Naito, K.; Sugiura, K. Effects of exosome-like vesicles on cumulus expansion in pigs in vitro. J. Reprod. Dev. 2017, 63, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ávila, A.C.F.C.M.; Bridi, A.; Andrade, G.M.; del Collado, M.; Sangalli, J.R.; Nociti, R.P.; da Silva Junior, W.A.; Bastien, A.; Robert, C.; Meirelles, F.V.; et al. Estrous cycle impacts microRNA content in extracellular vesicles that modulate bovine cumulus cell transcripts during in vitro maturation. Biol. Reprod. 2020, 102, 362–375. [Google Scholar] [CrossRef]
- Hailay, T.; Hoelker, M.; Poirier, M.; Gebremedhn, S.; Rings, F.; Saeed-Zidane, M.; Salilew-Wondim, D.; Dauben, C.; Tholen, E.; Neuhoff, C.; et al. Extracellular vesicle-coupled miRNA profiles in follicular fluid of cows with divergent post-calving metabolic status. Sci. Rep. 2019, 9, 12851. [Google Scholar] [CrossRef] [Green Version]
- Asaadi, A.; Kafi, M.; Atashi, H.; Azari, M.; Hostens, M. Frozen—Thawed ampullary cell monolayer improves bovine embryo in vitro development and quality. Zygote 2019, 27, 337–346. [Google Scholar] [CrossRef]
- Davachi, N.D.; Kohram, H.; Shahneh, A.Z.; Zhandi, M.; Goudarzi, A.; Fallahi, R.; Masoudi, R.; Yousefi, A.R.; Bartlewski, P.M. The effect of conspecific ampulla oviductal epithelial cells during in vitro maturation on oocyte developmental competence and maturation-promoting factor (MPF) activity in sheep. Theriogenology 2017, 88, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Coy, P.; García-Vázquez, F.A.; Visconti, P.E.; Avilés, M. Roles of the oviduct in mammalian fertilization. Reproduction 2012, 144, 649–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavani, K.C.; Lin, X.; Hamacher, J.; Broeck, W.V.D.; Couck, L.; Peelman, L.; Hendrix, A.; Van Soom, A. The Separation and Characterization of Extracellular Vesicles from Medium Conditioned by Bovine Embryos. Int. J. Mol. Sci. 2020, 21, 2942. [Google Scholar] [CrossRef] [PubMed]
- Lamparski, H.G.; Metha-Damani, A.; Yao, J.-Y.; Patel, S.; Hsu, D.-H.; Ruegg, C.; Le Pecq, J.-B. Production and characterization of clinical grade exosomes derived from dendritic cells. J. Immunol. Methods 2002, 270, 211–226. [Google Scholar] [CrossRef]
- Van Deun, J.; Mestdagh, P.; Sormunen, R.; Cocquyt, V.; Vermaelen, K.; Vandesompele, J.; Bracke, M.; De Wever, O.; Hendrix, A. The impact of disparate isolation methods for extracellular vesicles on downstream RNA profiling. J. Extracell. Vesicles 2014, 3, 24858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mol, E.A.; Goumans, M.-J.; Doevendans, P.A.; Sluijter, J.P.G.; Vader, P. Higher functionality of extracellular vesicles isolated using size-exclusion chromatography compared to ultracentrifugation. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 2061–2065. [Google Scholar] [CrossRef] [PubMed]
- Takov, K.; Yellon, D.M.; Davidson, S.M. Comparison of small extracellular vesicles isolated from plasma by ultracentrifugation or size-exclusion chromatography: Yield, purity and functional potential. J. Extracell. Vesicles 2018, 8, 1560809. [Google Scholar] [CrossRef] [PubMed]
- Lopera-Vásquez, R.; Hamdi, M.; Fernandez-Fuertes, B.; Maillo, V.; Beltrán-Breña, P.; Calle, A.; Redruello, A.; López-Martín, S.; Gutierrez-Adán, A.; Yañez-Mó, M.; et al. Extracellular Vesicles from BOEC in In Vitro Embryo Development and Quality. PLoS ONE 2016, 11, e0148083. [Google Scholar] [CrossRef]
- Navakanitworakul, R.; Hung, W.-T.; Gunewardena, S.; Davis, J.S.; Chotigeat, W.; Christenson, L.K. Characterization and Small RNA Content of Extracellular Vesicles in Follicular Fluid of Developing Bovine Antral Follicles. Sci. Rep. 2016, 6, 25486. [Google Scholar] [CrossRef]
- Davachi, N.D.; Shahneh, A.Z.; Kohram, H.; Zhandi, M.; Shamsi, H.; Hajiyavand, A.M.; Saadat, M. Differential influence of ampullary and isthmic derived epithelial cells on zona pellucida hardening and in vitro fertilization in ovine. Reprod. Biol. 2016, 16, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Oh, H.J.; Kim, M.J.; Kim, G.A.; Choi, Y.B.; Jo, Y.K.; Setyawan, E.N.; Lee, B.C. Effect of co-culture canine cumulus and oviduct cells with porcine oocytes during maturation and subsequent embryo development of parthenotes in vitro. Theriogenology 2018, 106, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Anna, L.-C.; Claudia, P.; Giulia, A.; Silvia, M.; Valentina, L.; Eleonora, O.; Paola, E.; Fausto, C. Oviductal microvesicles and their effect on in vitro maturation of canine oocytes. Reproduction 2017, 154, 167–180. [Google Scholar]
- Lamy, J.; Liere, P.; Pianos, A.; Aprahamian, F.; Mermillod, P.; Saint-Dizier, M. Steroid hormones in bovine oviductal fluid during the estrous cycle. Theriogenology 2016, 86, 1409–1420. [Google Scholar] [CrossRef] [PubMed]
- Consortium, E.-T.; Van Deun, J.; Mestdagh, P.; Agostinis, P.; Akay, Ö.; Anand, S.; Anckaert, J.; Martinez, Z.A.; Baetens, T.; Beghein, E.; et al. EV-TRACK: Transparent reporting and centralizing knowledge in extracellular vesicle research. Nat. Methods 2017, 14, 228. [Google Scholar]
- Wydooghe, E.; Heras, S.; Dewulf, J.; Piepers, S.; Van den Abbeel, E.; De Sutter, P.; Vandaele, L.; Van Soom, A. Replacing serum in culture medium with albumin and insulin, transferrin and selenium is the key to successful bovine embryo development in individual culture. Reprod. Fertil. Dev. 2014, 26, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Wydooghe, E.; Vandaele, L.; Beek, J.; Favoreel, H.; Heindryckx, B.; De Sutter, P.; Van Soom, A. Differential apoptotic staining of mammalian blastocysts based on double immunofluorescent CDX2 and active caspase-3 staining. Anal. Biochem. 2011, 416, 228–230. [Google Scholar] [CrossRef] [PubMed]
- Böing, A.N.; van der Pol, E.; Grootemaat, A.E.; Coumans, F.A.W.; Sturk, A.; Nieuwland, R. Single-step isolation of extracellular vesicles by size-exclusion chromatography. J. Extracell. Vesicles 2014, 3, 23430. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of EVs | Follicular Fluid | Ampullary Oviductal Fluid | ||
|---|---|---|---|---|
| Isolation Technique | SEC | ODG UC | SEC | ODG UC |
| Mean | 136.7 ± 3.9 nm | 107.9 ± 5.4 nm | 165.5 ± 5.8 nm | 166.9 ± 9.7 nm |
| Mode | 109.1 ± 2.5 nm | 79.9 ± 6.5 nm | 128.1 ± 14.3 nm | 145.5 ± 5.3 nm |
| SD | 52.1 ± 4.6 nm | 47.8 ± 5.9 nm | 84.0 ± 7.6 nm | 58.7 ± 8.2 nm |
| D10 | 91.1 ± 5.5 nm | 67.2 ± 2.5 nm | 91.7 ± 2.5 nm | 107.6 ± 2.2 nm |
| D50 | 122.5 ± 1.8 nm | 95.5 ± 4.8 nm | 139.1 ± 6.8 nm | 151.6 ± 5.7 nm |
| D90 | 179.9 ± 3.6 nm | 162.4 ± 15.9 nm | 311.8 ± 34.8 nm | 240.9 ± 23.7 nm |
| Nanoparticles/mL | 7.2 ± 0.79 × 1012 | 2.4 ± 0.2 × 1012 | 6.4 ± 0.5 × 1012 | 1.8 ± 0.2 × 1013 |
| Treatment 1 | Fertilized Oocytes | Cleavage Rate (%) 2 | Blastocyst Rate (%) 3 |
|---|---|---|---|
| (1) Control | 224 | 84.04 ± 1.15 a | 34.76 ± 1.69 c |
| (2) Negative control | 222 | 86.83 ± 1.15 a | 36.45 ± 1.69 bc |
| (3) FF EVs SEC | 216 | 87.52 ± 1.15 a | 35.85 ± 1.69 bc |
| (4) AOF EVs SEC | 221 | 87.85 ± 1.15 a | 39.79 ± 1.69 b |
| (5) FF EVs + AOF EVs SEC | 212 | 84.93 ± 1.15 a | 37.27 ± 1.69 bc |
| (6) FF EVs ODG UC | 218 | 86.72 ± 1.15 a | 45.41 ± 1.69 a |
| (7) AOF EVs ODG UC | 220 | 88.34 ± 1.15 a | 48.74 ± 1.69 a |
| (8) FF EVs+ AOF EVs ODG UC | 207 | 87.02 ± 1.15 a | 46.56 ± 1.69 a |
| Treatment 1 | No. of Blast. 2 | Cell Number | ACR (%) 6 | ||
|---|---|---|---|---|---|
| TCN ± SE 3 | ICM ± SE 4 | TE ± SE 5 | |||
| (1) Control | 42 | 135.97 ±1.37 d | 36.97 ± 0.66 d | 99.00 ± 0.95 d | 12.49 ± 0.19 a |
| (2) Negative control | 46 | 138.23 ± 1.31 d | 38.71 ± 0.63 d | 99.52 ± 0.91 d | 11.59 ± 0.18 b |
| (3) FF EVs SEC | 48 | 149.02 ± 1.28 c | 41.75 ± 0.61 c | 107.27 ± 0.89 c | 8.06 ± 0.18 c |
| (4) AOF EVs SEC | 46 | 153.00 ± 1.31 c | 42.97 ± 0.63 bc | 110.02 ± 0.91 c | 7.85 ± 0.18 c |
| (5) FF EVs + AOF EVs SEC | 47 | 152.48 ± 1.30 c | 43.12 ± 0.62 bc | 109.36 ± 0.90 c | 6.55 ± 0.18 d |
| (6) FF EVs ODG UC | 51 | 161.94 ± 1.24 b | 44.39 ± 0.60 b | 117.54 ± 0.87 b | 6.18 ± 0.17 d |
| (7) AOF EVs ODG UC | 58 | 166.93 ± 1.17 ab | 46.91 ± 0.56 a | 120.01 ± 0.81 ab | 6.60 ± 0.16 d |
| (8) FF EVs+ AOF EVs ODG UC | 56 | 168.69 ± 1.19 a | 47.01 ± 0.57 a | 121.67 ± 0.83 a | 5.94 ± 0.16 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asaadi, A.; Dolatabad, N.A.; Atashi, H.; Raes, A.; Van Damme, P.; Hoelker, M.; Hendrix, A.; Pascottini, O.B.; Van Soom, A.; Kafi, M.; et al. Extracellular Vesicles from Follicular and Ampullary Fluid Isolated by Density Gradient Ultracentrifugation Improve Bovine Embryo Development and Quality. Int. J. Mol. Sci. 2021, 22, 578. https://doi.org/10.3390/ijms22020578
Asaadi A, Dolatabad NA, Atashi H, Raes A, Van Damme P, Hoelker M, Hendrix A, Pascottini OB, Van Soom A, Kafi M, et al. Extracellular Vesicles from Follicular and Ampullary Fluid Isolated by Density Gradient Ultracentrifugation Improve Bovine Embryo Development and Quality. International Journal of Molecular Sciences. 2021; 22(2):578. https://doi.org/10.3390/ijms22020578
Chicago/Turabian StyleAsaadi, Anise, Nima Azari Dolatabad, Hadi Atashi, Annelies Raes, Petra Van Damme, Michael Hoelker, An Hendrix, Osvaldo Bogado Pascottini, Ann Van Soom, Mojtaba Kafi, and et al. 2021. "Extracellular Vesicles from Follicular and Ampullary Fluid Isolated by Density Gradient Ultracentrifugation Improve Bovine Embryo Development and Quality" International Journal of Molecular Sciences 22, no. 2: 578. https://doi.org/10.3390/ijms22020578