Large-Scale Phosphoproteomic Study of Arabidopsis Membrane Proteins Reveals Early Signaling Events in Response to Cold

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
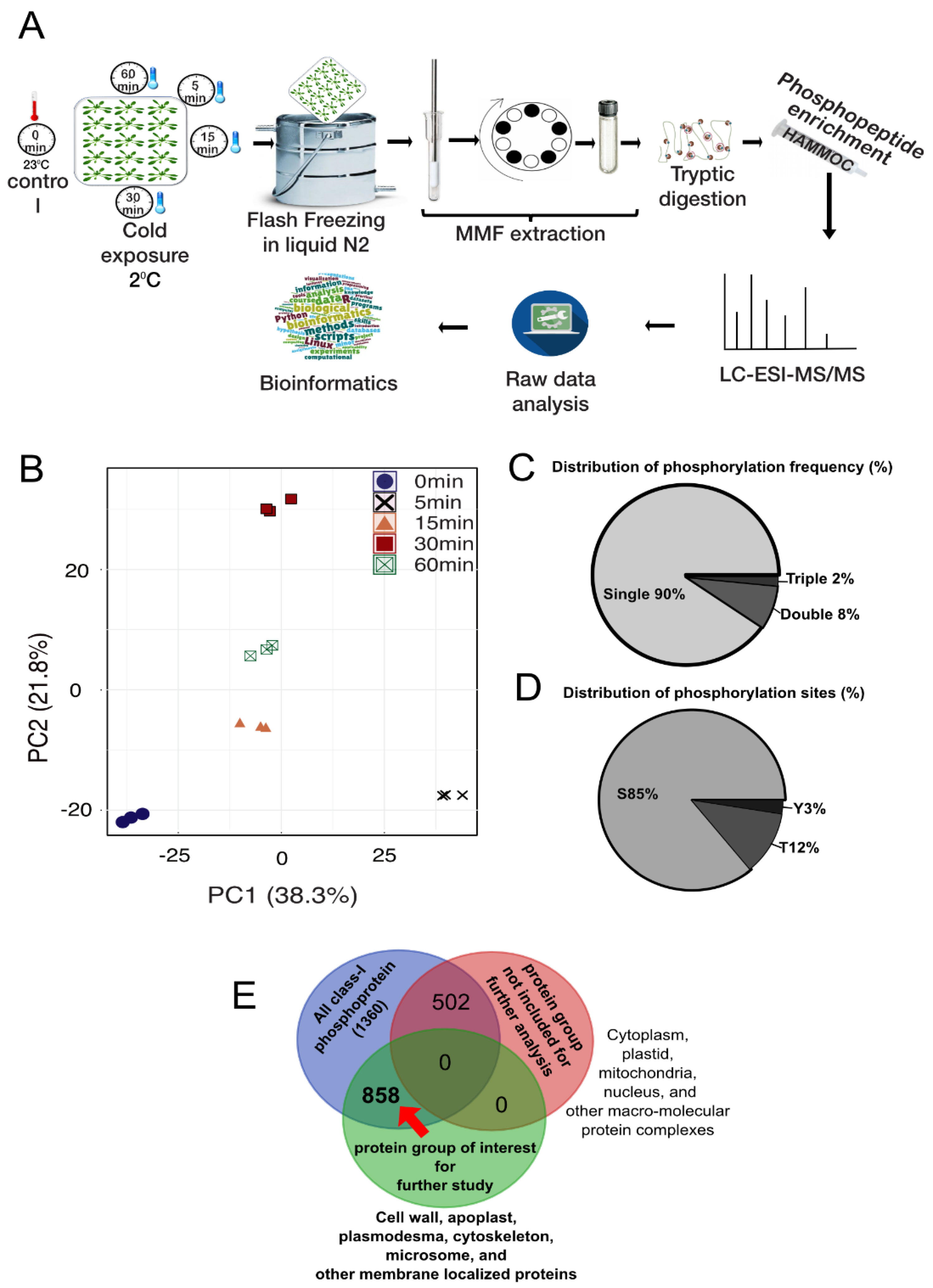
2.1. Description of the Phosphoproteomic Data of Early Cold Treatment in Arabidopsis
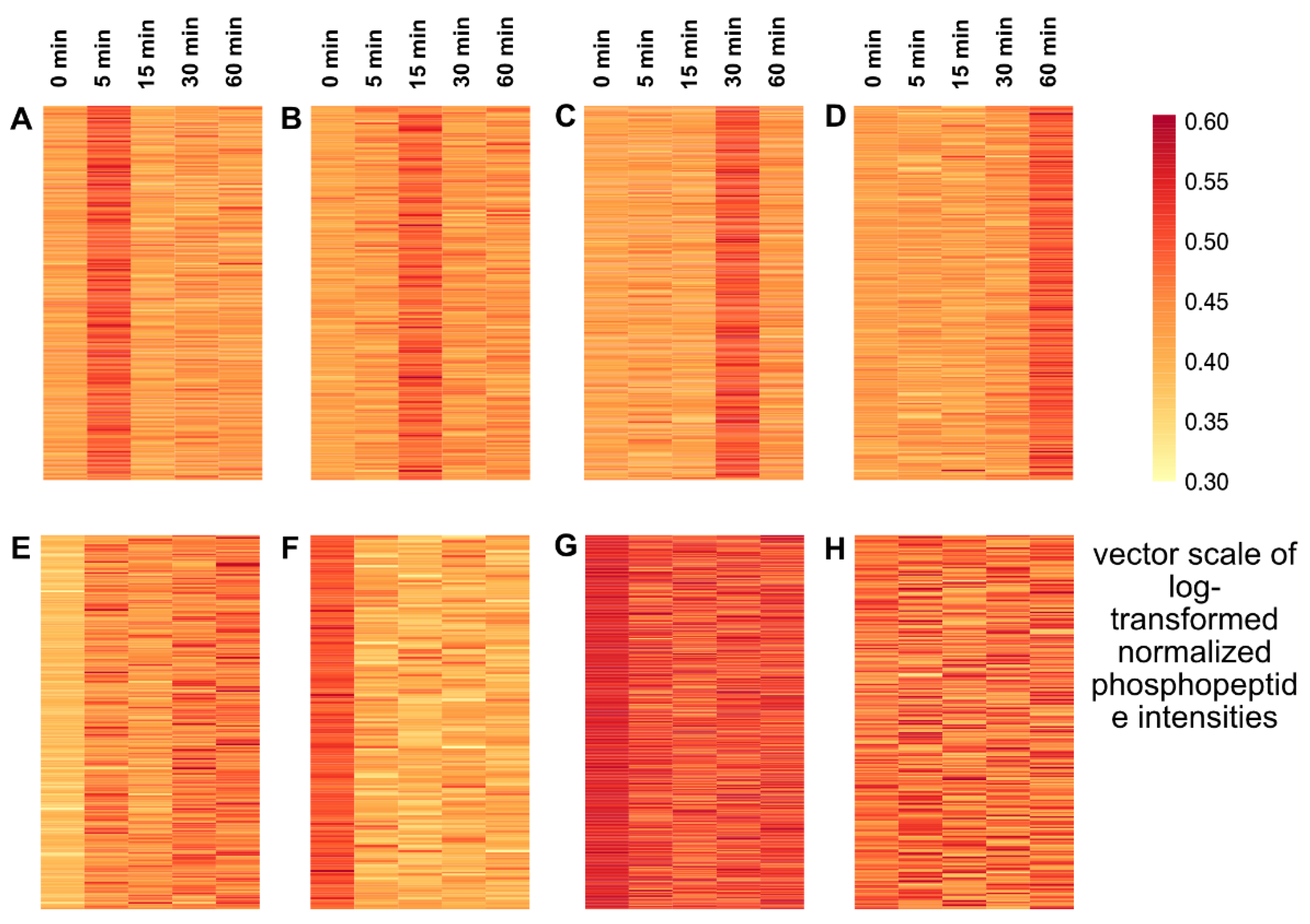
2.2. Temporal Profile of the Phosphopeptides in Response to Cold
2.3. Cold Signal Responsive Molecular Functions in Arabidopsis: GOMF
2.4. Cold Signal Responsive Biological Processes in Arabidopsis: GOBP
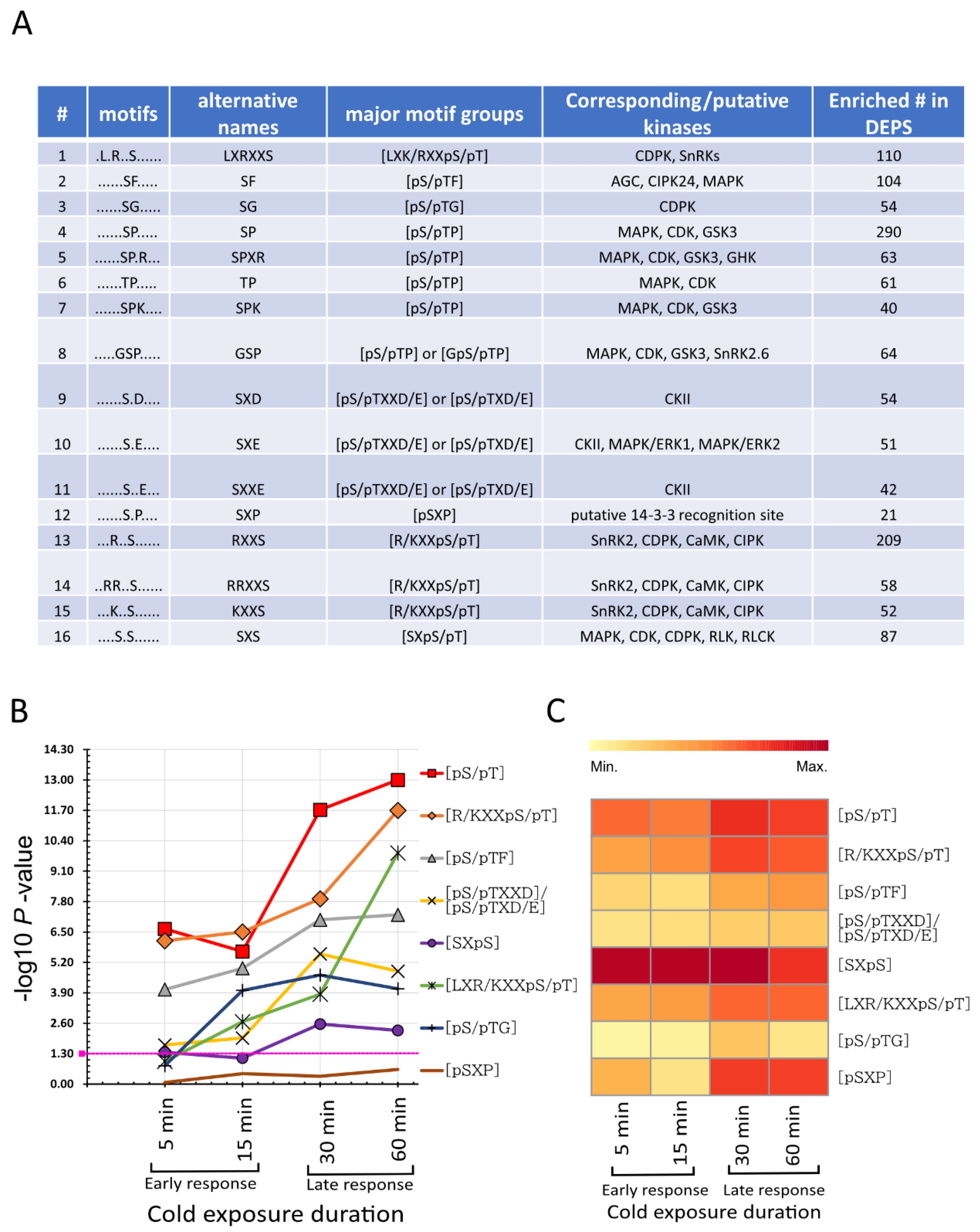
2.5. Cold-Responsive Phosphorylation Kinase Motifs
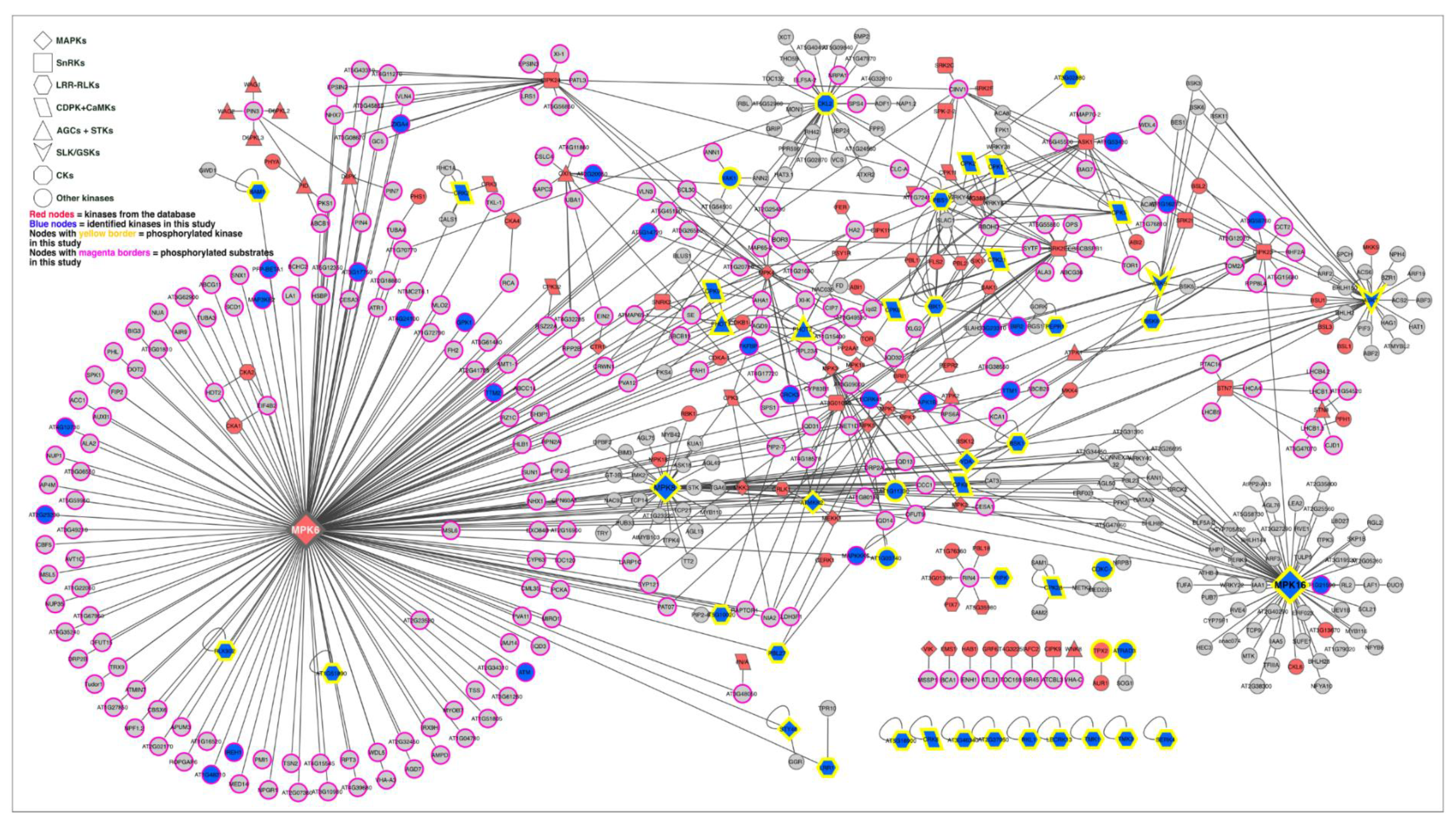
2.6. Kinase–Substrate Interaction Network
3. Discussion
3.1. Cold-Responsive Kinases and Signaling Proteins Might Play a Crucial Role in Early Cold Perception and Signal Transduction
3.2. Early Cold Response-Mediated Protein Phosphorylation Can Activate Phospholipid/Phosphoinositide Signaling Events
3.3. Early Cold Response Events Can Be Associated with Cytoskeletal and Vesicle Trafficking Systems
3.4. Transporters Respond to Early Cold Stress to Maintain Homeostasis
3.5. ROS Signaling under Cold Response
4. Materials and Methods
4.1. Plant Growth Conditions and Cold Exposure
4.2. Microsomal Membrane Fraction (MMF) Extraction
4.3. Phosphoproteomic Analysis
4.3.1. In-Solution Trypsin Digestion
4.3.2. Phosphopeptide Enrichment and Desalting
4.3.3. LC-ESI-MS/MS Analysis
4.3.4. Phosphopeptide Identification and Quantification
4.4. Bioinformatic Analysis
4.4.1. Gene Ontology Analysis
4.4.2. Phosphorylation Motif Dynamics Analysis
4.4.3. Kinase–Substrate Network Construction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MMF | Microsomal membrane fraction |
| LC-ESI-MS/MS | Liquid chromatography–electrospray ionization mass spectrometry/mass spectrometry |
References
- Levitt, J. Chilling, Freezing and High Temperature Stresses; Academic Press: Cambridge, MA, USA, 1980; Volume 1, pp. 27–64. [Google Scholar]
- Zhu, J.; Dong, C.-H.; Zhu, J.-K. Interplay between cold-responsive gene regulation, metabolism and RNA processing during plant cold acclimation. Curr. Opin. Plant Biol. 2007, 10, 290–295. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.; Zhu, J.-K. Cold stress regulation of gene expression in plants. Trends Plant Sci. 2007, 12, 444–451. [Google Scholar] [CrossRef]
- Miura, K.; Furumoto, T. Cold Signaling and Cold Response in Plants. Int. J. Mol. Sci. 2013, 14, 5312–5337. [Google Scholar] [CrossRef] [Green Version]
- Thomashow, M.F. Plant Cold Acclimation: Freezing Tolerance Genes and Regulatory Mechanisms. Annu. Rev. Plant Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [Green Version]
- Gilmour, S.J.; Hajela, R.K.; Thomashow, M.F. Cold Acclimation in Arabidopsis thaliana. Plant Physiol. 1988, 87, 745–750. [Google Scholar] [CrossRef] [Green Version]
- Miki, Y.; Takahashi, D.; Kawamura, Y.; Uemura, M. Temporal proteomics of Arabidopsis plasma membrane during cold- and de-acclimation. J. Proteom. 2019, 197, 71–81. [Google Scholar] [CrossRef]
- Takahashi, D.; Kawamura, Y.; Uemura, M. Changes of Detergent-Resistant Plasma Membrane Proteins in Oat and Rye during Cold Acclimation: Association with Differential Freezing Tolerance. J. Proteome Res. 2013, 12, 4998–5011. [Google Scholar] [CrossRef]
- Li, B.; Takahashi, D.; Kawamura, Y.; Uemura, M. Comparison of Plasma Membrane Proteomic Changes of Arabidopsis Suspension-Cultured Cells (T87 Line) after Cold and ABA Treatment in Association with Freezing Tolerance Development. Plant Cell Physiol. 2012, 53, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, D.; Imai, H.; Kawamura, Y.; Uemura, M. Lipid profiles of detergent resistant fractions of the plasma membrane in oat and rye in association with cold acclimation and freezing tolerance. Cryobiology 2016, 72, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, D.; Kawamura, Y.; Uemura, M. Cold acclimation is accompanied by complex responses of glycosylphosphatidylinositol (GPI)-anchored proteins in Arabidopsis. J. Exp. Bot. 2016, 67, 5203–5215. [Google Scholar] [CrossRef] [Green Version]
- Medina, J.; Catalá, R.; Salinas, J. The CBFs: Three arabidopsis transcription factors to cold acclimate. Plant Sci. 2011, 180, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Etakahashi, D.; Eli, B.; Enakayama, T.; Kawamura, Y.; Uemura, M. Plant plasma membrane proteomics for improving cold tolerance. Front. Plant Sci. 2013, 4, 90. [Google Scholar] [CrossRef] [Green Version]
- Osakabe, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S.P. Sensing the environment: Key roles of membrane-localized kinases in plant perception and response to abiotic stress. J. Exp. Bot. 2013, 64, 445–458. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.-W.; Guan, S.; Sun, Y.; Deng, Z.; Tang, W.; Shang, J.-X.; Sun, Y.; Burlingame, A.L.; Wang, Z.-Y. Brassinosteroid signal transduction from cell-surface receptor kinases to nuclear transcription factors. Nat. Cell Biol. 2009, 11, 1254–1260. [Google Scholar] [CrossRef]
- Marshall, A.; Aalen, R.B.; Audenaert, D.; Beeckman, T.; Broadley, M.R.; Butenko, M.A.; Caño-Delgado, A.I.; De Vries, S.; Dresselhaus, T.; Felix, G.; et al. Tackling Drought Stress: Receptor-like kinases present new approaches. Plant Cell 2012, 24, 2262–2278. [Google Scholar] [CrossRef]
- Mattei, B.; Spinelli, F.; Pontiggia, D.; De Lorenzo, G. Comprehensive Analysis of the Membrane Phosphoproteome Regulated by Oligogalacturonides in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, D.; Gorka, M.; Erban, A.; Graf, A.; Kopka, J.; Zuther, E.; Hincha, D.K. Both cold and sub-zero acclimation induce cell wall modification and changes in the extracellular proteome in Arabidopsis thaliana. Sci. Rep. 2019, 9, 2289. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Uemura, M. Protein and Lipid Compositions of Isolated Plasma Membranes from Orchard Grass (Dactylis glomerata L.) and Changes during Cold Acclimation. Plant Physiol. 1984, 75, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Willick, I.R.; Takahashi, D.; Fowler, D.B.; Uemura, M.; Tanino, K. Tissue-specific changes in apoplastic proteins and cell wall structure during cold acclimation of winter wheat crowns. J. Exp. Bot. 2018, 69, 1221–1234. [Google Scholar] [CrossRef] [Green Version]
- Novaković, L.; Guo, T.; Bacic, A.; Sampathkumar, A.; Johnson, K.L. Hitting the Wall—Sensing and Signaling Pathways Involved in Plant Cell Wall Remodeling in Response to Abiotic Stress. Plants 2018, 7, 89. [Google Scholar] [CrossRef] [Green Version]
- Schulze, W.X. Proteomics approaches to understand protein phosphorylation in pathway modulation. Curr. Opin. Plant Biol. 2010, 13, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Furuya, T.; Matsuoka, D.; Nanmori, T. Membrane rigidification functions upstream of the MEKK1-MKK2-MPK4 cascade during cold acclimation in Arabidopsis thaliana. FEBS Lett. 2014, 588, 2025–2030. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Furuya, T.; Matsuoka, D.; Nanmori, T. Phosphorylation of Arabidopsis thaliana MEKK1 via Ca2+ signaling as a part of the cold stress response. J. Plant Res. 2013, 126, 833–840. [Google Scholar] [CrossRef]
- Yang, T.; Chaudhuri, S.; Yang, L.; Du, L.; Poovaiah, B. A Calcium/Calmodulin-regulated Member of the Receptor-like Kinase Family Confers Cold Tolerance in Plants. J. Biol. Chem. 2010, 285, 7119–7126. [Google Scholar] [CrossRef] [Green Version]
- Teige, M.; Scheikl, E.; Eulgem, T.; Dóczi, R.; Ichimura, K.; Shinozaki, K.; Dangl, J.L.; Hirt, H. The MKK2 Pathway Mediates Cold and Salt Stress Signaling in Arabidopsis. Mol. Cell 2004, 15, 141–152. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, S.; He, W.-D.; Shao, X.-H.; Li, C.-Y.; Wei, Y.-R.; Deng, G.-M.; Kuang, R.-B.; Hu, C.-H.; Yi, G.-J.; et al. Comparative Phosphoproteomics Reveals an Important Role of MKK2 in Banana (Musa spp.) Cold Signal Network. Sci. Rep. 2017, 7, srep40852. [Google Scholar] [CrossRef]
- Yang, T.; Ali, G.S.; Yang, L.; Du, L.; Reddy, A.; Poovaiah, B. Calcium/calmodulin-regulated receptor-like kinase CRLK1 interacts with MEKK1 in plants. Plant Signal. Behav. 2010, 5, 991–994. [Google Scholar] [CrossRef]
- Benschop, J.J.; Mohammed, S.; O’Flaherty, M.; Heck, A.J.R.; Slijper, M.; Menke, F.L.H. Quantitative Phosphoproteomics of Early Elicitor Signaling inArabidopsis. Mol. Cell. Proteomics 2007, 6, 1198–1214. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Jia, Y.; Ding, Y.; Shi, Y.; Li, Z.; Guo, Y.; Gong, Z.; Yang, S. Plasma Membrane CRPK1-Mediated Phosphorylation of 14-3-3 Proteins Induces Their Nuclear Import to Fine-Tune CBF Signaling during Cold Response. Mol. Cell 2017, 66, 117–128.e5. [Google Scholar] [CrossRef] [Green Version]
- Osakabe, Y.; Mizuno, S.; Tanaka, H.; Maruyama, K.; Osakabe, K.; Todaka, D.; Fujita, Y.; Kobayashi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Overproduction of the Membrane-bound Receptor-like Protein Kinase 1, RPK1, Enhances Abiotic Stress Tolerance in Arabidopsis. J. Biol. Chem. 2010, 285, 9190–9201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tichy, A.; Salovska, B.; Rehulka, P.; Klimentova, J.; Vavrova, J.; Stulik, J.; Hernychova, L. Phosphoproteomics: Searching for a needle in a haystack. J. Proteomics 2011, 74, 2786–2797. [Google Scholar] [CrossRef] [PubMed]
- Kersten, B.; Agrawal, G.K.; Iwahashi, H.; Rakwal, R. Plant phosphoproteomics: A long road ahead. Proteomics 2006, 6, 5517–5528. [Google Scholar] [CrossRef]
- Chen, Y.; Weckwerth, W. Mass Spectrometry Untangles Plant Membrane Protein Signaling Networks. Trends Plant Sci. 2020, 25, 930–944. [Google Scholar] [CrossRef]
- Chen, Y.; Hoehenwarter, W. Changes in the Phosphoproteome and Metabolome Link Early Signaling Events to Rearrangement of Photosynthesis and Central Metabolism in Salinity and Oxidative Stress Response in Arabidopsis1. Plant Physiol. 2015, 169, 3021–3033. [Google Scholar] [CrossRef] [Green Version]
- Nühse, T.S.; Stensballe, A.; Jensen, O.N.; Peck, S.C. Large-scale Analysis of in Vivo Phosphorylated Membrane Proteins by Immobilized Metal Ion Affinity Chromatography and Mass Spectrometry. Mol. Cell. Proteomics 2003, 2, 1234–1243. [Google Scholar] [CrossRef] [Green Version]
- Niittylä, T.; Fuglsang, A.T.; Palmgren, M.G.; Frommer, W.B.; Schulze, W.X. Temporal Analysis of Sucrose-induced Phosphorylation Changes in Plasma Membrane Proteins of Arabidopsis. Mol. Cell. Proteom. 2007, 6, 1711–1726. [Google Scholar] [CrossRef] [Green Version]
- Engelsberger, W.R.; Schulze, W.X. Nitrate and ammonium lead to distinct global dynamic phosphorylation patterns when resupplied to nitrogen-starved Arabidopsis seedlings. Plant J. 2012, 69, 978–995. [Google Scholar] [CrossRef] [Green Version]
- Xue, L.; Wang, P.; Wang, L.; Renzi, E.; Radivojac, P.; Tang, H.; Arnold, R.; Zhu, J.-K.; Tao, W.A. Quantitative Measurement of Phosphoproteome Response to Osmotic Stress inArabidopsisBased on Library-Assisted eXtracted Ion Chromatogram (LAXIC). Mol. Cell. Proteomics 2013, 12, 2354–2369. [Google Scholar] [CrossRef] [Green Version]
- Stecker, K.E.; Minkoff, B.B.; Sussman, M.R. Phosphoproteomic Analyses Reveal Early Signaling Events in the Osmotic Stress Response. Plant Physiol. 2014, 165, 1171–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savas, J.N.; Stein, B.D.; Wu, C.C.; Yates, J.R. Mass spectrometry accelerates membrane protein analysis. Trends Biochem. Sci. 2011, 36, 388–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagami, H.; Sugiyama, N.; Mochida, K.; Daudi, A.; Yoshida, Y.; Toyoda, T.; Tomita, M.; Ishihama, Y.; Shirasu, K. Large-Scale Comparative Phosphoproteomics Identifies Conserved Phosphorylation Sites in Plants. Plant Physiol. 2010, 153, 1161–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Na Wu, X.; Xi, L.; Pertl-Obermeyer, H.; Liang-Cui, C.; Chu, L.-C.; Schulze, W.X. Highly Efficient Single-Step Enrichment of Low Abundance Phosphopeptides from Plant Membrane Preparations. Front. Plant Sci. 2017, 8, 1673. [Google Scholar] [CrossRef]
- Ishikawa, S.; Barrero, J.M.; Takahashi, F.; Peck, S.C.; Gubler, F.; Shinozaki, K.; Umezawa, T. Comparative Phosphoproteomic Analysis of Barley Embryos with Different Dormancy during Imbibition. Int. J. Mol. Sci. 2019, 20, 451. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-C.; Zhu, Y.; Arrington, J.V.; Paez, J.S.; Wang, P.; Zhu, P.; Chen, I.-H.; Zhu, J.-K.; Tao, W.A. Universal Plant Phosphoproteomics Workflow and Its Application to Tomato Signaling in Response to Cold Stress. Mol. Cell. Proteom. 2018, 17, 2068–2080. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F.H.; Na Wu, X.; Stintzi, A.; Schaller, A.; Schulze, W.X. The Systemin Signaling Cascade As Derived from Time Course Analyses of the Systemin-responsive Phosphoproteome. Mol. Cell. Proteom. 2019, 18, 1526–1542. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, S.; Takahashi, F.; Nakagami, H.; Peck, S.C.; Gubler, F.; Shinozaki, K.; Umezawa, T.; Barrero, J.M.; Barrero, J.M. Comparative Phosphoproteomic Analysis Reveals a Decay of ABA Signaling in Barley Embryos during After-Ripening. Plant Cell Physiol. 2019, 60, 2758–2768. [Google Scholar] [CrossRef]
- Umezawa, T.; Sugiyama, N.; Takahashi, F.; Anderson, J.C.; Ishihama, Y.; Peck, S.C.; Shinozaki, K. Genetics and Phosphoproteomics Reveal a Protein Phosphorylation Network in the Abscisic Acid Signaling Pathway in Arabidopsis thaliana. Sci. Signal. 2013, 6, rs8. [Google Scholar] [CrossRef]
- Wu, X.; Sklodowski, K.; Encke, B.; Schulze, W.X. A Kinase-Phosphatase Signaling Module with BSK8 and BSL2 Involved in Regulation of Sucrose-Phosphate Synthase. J. Proteome Res. 2014, 13, 3397–3409. [Google Scholar] [CrossRef]
- Balakrishnan, R.; Harris, M.A.; Huntley, R.P.; Van Auken, K.; Cherry, J.M. A guide to best practices for Gene Ontology (GO) manual annotation. Database 2013, 2013, bat054. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.D. The Gene Ontology and the Meaning of Biological Function. In The Gene Ontology Handbook; Springer: New York, NY, USA, 2017; Volume 1446, pp. 15–24. [Google Scholar]
- Van Wijk, K.J.; Friso, G.; Walther, D.; Schulze, W.X. Meta-Analysis of Arabidopsis thaliana Phospho-Proteomics Data Reveals Compartmentalization of Phosphorylation Motifs. Plant Cell 2014, 26, 2367–2389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulawski, M.; Braginets, R.; Schulze, W.X. PhosPhAt goes kinases—Searchable protein kinase target information in the plant phosphorylation site database PhosPhAt. Nucleic Acids Res. 2012, 41, D1176–D1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heazlewood, J.L.; Durek, P.; Hummel, J.; Selbig, J.; Weckwerth, W.; Walther, D.; Schulze, W.X. PhosPhAt: A database of phosphorylation sites in Arabidopsis thaliana and a plant-specific phosphorylation site predictor. Nucleic Acids Res. 2007, 36, D1015–D1021. [Google Scholar] [CrossRef] [PubMed]
- Durek, P.; Schmidt, R.; Heazlewood, J.L.; Jones, A.; MacLean, D.; Nagel, A.; Kersten, B.; Schulze, W.X. PhosPhAt: The Arabidopsis thaliana phosphorylation site database. An update. Nucleic Acids Res. 2010, 38, D828–D834. [Google Scholar] [CrossRef]
- Wang, P.; Hsu, C.-C.; Du, Y.; Zhu, P.; Zhao, C.; Fu, X.; Zhang, C.; Paez, J.S.; Macho, A.P.; Tao, W.A.; et al. Mapping proteome-wide targets of protein kinases in plant stress responses. Proc. Natl. Acad. Sci. USA 2020, 117, 3270–3280. [Google Scholar]
- Khan, M.; Rozhon, W.; Bigeard, J.; Pflieger, D.; Husar, S.; Pitzschke, A.; Teige, M.; Jonak, C.; Hirt, H.; Poppenberger, B. Brassinosteroid-regulated GSK3/Shaggy-like kinases phosphorylate mitogen-activated protein (MAP) kinase kinases, which control stomata development in Arabidopsis thaliana. J. Biol. Chem. 2013, 288, 7519–7527. [Google Scholar]
- Lv, D.; Li, X.; Zhang, M.; Gu, A.; Zhen, S.; Wang, C.; Li, X.; Yan, Y. Large-scale phosphoproteome analysis in seedling leaves of Brachypodium distachyon L. BMC Genom. 2014, 15, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, D.; Gygi, S.P. An iterative statistical approach to the identification of protein phosphorylation motifs from large-scale data sets. Nat. Biotechnol. 2005, 23, 1391–1398. [Google Scholar] [CrossRef]
- Zhang, S.H.; Kobayashi, R.; Graves, P.R.; Piwnica-Worms, H.; Tonks, N.K. Serine phosphorylation-dependent association of the band 4.1-related protein-tyrosine phosphatase PTPH1 with 14-3-3beta protein. J. Biol. Chem. 1997, 272, 27281–27287. [Google Scholar]
- Ku, N.-O.; Liao, J.; Omary, M.B. Phosphorylation of human keratin 18 serine 33 regulates binding to 14-3-3 proteins. EMBO J. 1998, 17, 1892–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pi, E.; Zhu, C.; Fan, W.; Huang, Y.; Qu, L.; Li, Y.; Zhao, Q.; Ding, F.; Qiu, L.; Wang, H.; et al. Quantitative Phosphoproteomic and Metabolomic Analyses Reveal GmMYB173 Optimizes Flavonoid Metabolism in Soybean under Salt Stress. Mol. Cell. Proteom. 2018, 17, 1209–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.-L.; Hsu, C.-L.; Hu, C.-W.; Ko, S.-Y.; Hsieh, H.-L.; Huang, H.-C.; Juan, H.-F. Integrating Phosphoproteomics and Bioinformatics to Study Brassinosteroid-Regulated Phosphorylation Dynamics in Arabidopsis. BMC Genom. 2015, 16, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, H.; Knight, M.R. Imaging spatial and cellular characteristics of low temperature calcium signature after cold acclimation in Arabidopsis. J. Exp. Bot. 2000, 51, 1679–1686. [Google Scholar] [CrossRef] [Green Version]
- Martí, M.C.; Stancombe, M.A.; Webb, A.A. Cell- and Stimulus Type-Specific Intracellular Free Ca2+ Signals in Arabidopsis. Plant Physiol. 2013, 163, 625–634. [Google Scholar] [CrossRef] [Green Version]
- Hiraki, H.; Uemura, M.; Kawamura, Y. Calcium Signaling-LinkedCBF/DREB1Gene Expression was Induced Depending on the Temperature Fluctuation in the Field: Views from the Natural Condition of Cold Acclimation. Plant Cell Physiol. 2018, 60, 303–317. [Google Scholar] [CrossRef]
- Yuan, P.; Yang, T.; Poovaiah, B. Calcium Signaling-Mediated Plant Response to Cold Stress. Int. J. Mol. Sci. 2018, 19, 3896. [Google Scholar] [CrossRef] [Green Version]
- Knight, H.; Trewavas, A.J.; Knight, M.R. Cold calcium signaling in Arabidopsis involves two cellular pools and a change in calcium signature after acclimation. Plant Cell 1996, 8, 489–503. [Google Scholar]
- Mao, J.; Manik, S.M.N.; Shi, S.; Chao, J.; Jin, Y.; Wang, Q.; Liu, H. Mechanisms and Physiological Roles of the CBL-CIPK Networking System in Arabidopsis thaliana. Genes 2016, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Luan, S. The CBL–CIPK network in plant calcium signaling. Trends Plant Sci. 2009, 14, 37–42. [Google Scholar] [CrossRef]
- Zeng, H.; Xu, L.; Singh, A.; Wang, H.; Du, L.; Poovaiah, B. Involvement of calmodulin and calmodulin-like proteins in plant responses to abiotic stresses. Front. Plant Sci. 2015, 6, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.; Takasaki, H.; Komatsu, S. Comprehensive Phosphoproteome Analysis in Rice and Identification of Phosphoproteins Responsive to Different Hormones/Stresses. J. Proteome Res. 2005, 4, 1592–1599. [Google Scholar] [CrossRef] [PubMed]
- Pi, Z.; Zhao, M.-L.; Peng, X.-J.; Shen, S. Phosphoproteomic Analysis of Paper Mulberry Reveals Phosphorylation Functions in Chilling Tolerance. J. Proteome Res. 2017, 16, 1944–1961. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, F.-F.; Peng, X.; Huang, J.; Shen, S. Global Phosphoproteomic Analysis Reveals the Defense and Response Mechanisms of Jatropha Curcas Seedling under Chilling Stress. Int. J. Mol. Sci. 2019, 20, 208. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Xun, Q.; Guo, Y.; Zhang, J.; Cheng, K.; Shi, T.; He, K.; Hou, S.; Gou, X.; Li, J. Genome-Wide Expression Pattern Analyses of the Arabidopsis Leucine-Rich Repeat Receptor-Like Kinases. Mol. Plant 2016, 9, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.-H.; Henderson, D.A.; Zhu, J.-K. The Arabidopsis Cold-Responsive Transcriptome and Its Regulation by ICE1. Plant Cell 2005, 17, 3155–3175. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, C.; Russinova, E.; Kemmerling, B.; Kwaaitaal, M.; De Vries, S.C. Arabidopsis somatic embryogenesis receptor kinase proteins serve brassinosteroid-dependent and -independent signaling pathways. Plant Physiol. 2008, 148, 611–619. [Google Scholar] [CrossRef] [Green Version]
- He, K.; Gou, X.; Yuan, T.; Lin, H.; Asami, T.; Yoshida, S.; Russell, S.D.; Li, J. BAK1 and BKK1 Regulate Brassinosteroid-Dependent Growth and Brassinosteroid-Independent Cell-Death Pathways. Curr. Biol. 2007, 17, 1109–1115. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.-H.; Wang, X.; Kota, U.; Goshe, M.B.; Clouse, S.D.; Huber, S.C. Tyrosine phosphorylation of the BRI1 receptor kinase emerges as a component of brassinosteroid signaling in Arabidopsis. Tyrosine phosphorylation of the BRI1 receptor kinase emerges as a component of brassinosteroid signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 658–663. [Google Scholar]
- Kong, Q.; Sun, T.; Qu, N.; Ma, J.; Li, M.; Cheng, Y.-T.; Zhang, Q.; Wu, D.; Zhang, Z.; Zhang, Y. Two redundant receptor-like cytoplasmic kinases function downstream of pattern recognition receptors to regulate activation of SA biosynthesis in Arabidopsis. Plant Physiol. 2016, 171, 1344–1354. [Google Scholar] [CrossRef] [Green Version]
- Ou, Y.; Lu, X.; Zi, Q.; Xun, Q.; Zhang, J.; Wu, Y.; Shi, H.; Wei, Z.; Zhao, B.; Zhang, X.; et al. RGF1 INSENSITIVE 1 to 5, a group of LRR receptor-like kinases, are essential for the perception of root meristem growth factor 1 in Arabidopsis thaliana. Cell Res. 2016, 26, 686–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.; Liu, L.; Wang, J.; Wu, Z.; Zhang, H.; Tang, J.; Lin, G.; Wang, Y.; Wenyang, L.; Li, W.; et al. Signature motif-guided identification of receptors for peptide hormones essential for root meristem growth. Cell Res. 2016, 26, 674–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Dai, N.; Chen, J.; Nagawa, S.; Cao, M.; Li, H.; Zhou, Z.; Chen, X.; De Rycke, R.; Rakusová, H.; et al. Cell Surface ABP1-TMK Auxin-Sensing Complex Activates ROP GTPase Signaling. Science 2014, 343, 1025–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.; Chen, R.; Li, P.; Yu, Y.; Zheng, R.; Ge, D.; Zheng, W.; Wang, X.; Gu, Y.; Gelová, Z.; et al. TMK1-mediated auxin signalling regulates differential growth of the apical hook. Nat. Cell Biol. 2019, 568, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, M.; Yu, L.; Zhou, Z.; Liang, X.; Liu, Z.; Cai, G.; Gao, L.; Zhang, X.; Wang, Y.; et al. The FLS2-Associated Kinase BIK1 Directly Phosphorylates the NADPH Oxidase RbohD to Control Plant Immunity. Cell Host Microbe 2014, 15, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Wu, Y.; Yang, F.; Zhang, Y.; Chen, S.; Xie, Q.; Tian, X.; Zhou, J.-M. BIK1 interacts with PEPRs to mediate ethylene-induced immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 6205–6210. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Lu, D.; Gao, X.; Jiang, S.; Ma, X.; Wang, Z.; Mengiste, T.; He, P.; Shan, L. Inverse modulation of plant immune and brassinosteroid signaling pathways by the receptor-like cytoplasmic kinase BIK1. Proc. Natl. Acad. Sci. USA 2013, 110, 12114–12119. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Yamaguchi, K.; Shirakawa, T.; Nakagami, H.; Mine, A.; Ishikawa, K.; Fujiwara, M.; Narusaka, M.; Narusaka, Y.; Ichimura, K.; et al. The Arabidopsis CERK 1-associated kinase PBL 27 connects chitin perception to MAPK activation. EMBO J. 2016, 35, 2468–2483. [Google Scholar] [CrossRef]
- Champion, A.; Picaud, A.; Henry, Y. Reassessing the MAP3K and MAP4K relationships. Trends Plant Sci. 2004, 9, 123–129. [Google Scholar] [CrossRef]
- Sun, H.; King, A.J.; Diaz, H.; Marshall, M.S. Regulation of the protein kinase Raf-1 by oncogenic Ras through phosphatidylinositol 3-kinase, Cdc42/Rac and Pak. Curr. Biol. 2000, 10, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Miki, Y. Shotgun Proteomics and Phosphoproteomics Analysis of Cold Acclimation and De-Acclimation in Arabidopsis Thaliana. Master’s Thesis, Iwate University, Iwate, Japan, 2015. [Google Scholar]
- Chaiwongsar, S.; Strohm, A.K.; Su, S.-H.; Krysan, P.J. Genetic analysis of the Arabidopsis protein kinases MAP3Kε1 and MAP3Kε2 indicates roles in cell expansion and embryo development. Front. Plant Sci. 2012, 3, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiwongsar, S.; Otegui, M.S.; Jester, P.J.; Monson, S.S.; Krysan, P.J. The protein kinase genes MAP3K epsilon 1 and MAP3K epsilon 2 are required for pollen viability in Arabidopsis thaliana. Plant. J. 2006, 48, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, D.C.; Lukowitz, W.; Somerville, C.R. Stomatal Development and Pattern Controlled by a MAPKK Kinase. Science 2004, 304, 1494–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Wang, H.; He, Y.; Liu, Y.; Walker, J.C.; Torii, K.U.; Zhang, S. A MAPK Cascade Downstream of ERECTA Receptor-Like Protein Kinase Regulates Arabidopsis Inflorescence Architecture by Promoting Localized Cell Proliferation. Plant Cell 2012, 24, 4948–4960. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Nam, K.H.; Vafeados, D.; Chory, J. BIN2, a New Brassinosteroid-Insensitive Locus in Arabidopsis. Plant Physiol. 2001, 127, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Sreeramulu, S.; Mostizky, Y.; Sunitha, S.; Shani, E.; Nahum, H.; Salomon, D.; Ben Hayun, L.; Gruetter, C.; Rauh, D.; Ori, N.; et al. BSKs are partially redundant positive regulators of brassinosteroid signaling in Arabidopsis. Plant J. 2013, 74, 905–919. [Google Scholar] [CrossRef]
- Mosblech, A.; König, S.; Stenzel, I.; Grzeganek, P.; Feussner, I.; Heilmann, I. Phosphoinositide and Inositolpolyphosphate Signalling in Defense Responses of Arabidopsis thaliana Challenged by Mechanical Wounding. Mol. Plant 2008, 1, 249–261. [Google Scholar] [CrossRef] [Green Version]
- Mishkind, M.; Vermeer, J.E.M.; Darwish, E.; Munnik, T. Heat stress activates phospholipase D and triggers PIP2accumulation at the plasma membrane and nucleus. Plant J. 2009, 60, 10–21. [Google Scholar] [CrossRef]
- Dewald, D.B.; Torabinejad, J.; Jones, C.A.; Shope, J.C.; Cangelosi, A.R.; Thompson, J.E.; Prestwich, G.D.; Hama, H. Rapid Accumulation of Phosphatidylinositol 4,5-Bisphosphate and Inositol 1,4,5-Trisphosphate Correlates with Calcium Mobilization in Salt-Stressed Arabidopsis. Plant Physiol. 2001, 126, 759–769. [Google Scholar] [CrossRef] [Green Version]
- Heilmann, I. Phosphoinositide signaling in plant development. Development 2016, 143, 2044–2055. [Google Scholar] [CrossRef] [Green Version]
- Dove, S.K.; Cooke, F.T.; Douglas, M.R.; Sayers, L.G.; Parker, P.J.; Michell, R.H. Osmotic stress activates phosphatidylinositol-3,5-bisphosphate synthesis. Nat. Cell Biol. 1997, 390, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Bonangelino, C.J.; Nau, J.J.; Duex, J.E.; Brinkman, M.; Wurmser, A.E.; Gary, J.D.; Emr, S.D.; Weisman, L.S. Osmotic stress–induced increase of phosphatidylinositol 3,5-bisphosphate requires Vac14p, an activator of the lipid kinase Fab1p. J. Cell Biol. 2002, 156, 1015–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, C.Y.; Alghamdi, T.A.; Botelho, R.J. Phosphatidylinositol-3,5-Bisphosphate: No Longer the Poor PIP2. Traffic 2011, 13, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Munnik, T.; Sato, M.H. Phosphatidylinositol 3-phosphate 5-kinase, fab1/pikfyve kinase mediates endosome maturation to establish endosome-cortical microtubule interaction in Arabidopsis. Plant Physiol. 2015, 169, 1961–1974. [Google Scholar] [CrossRef] [Green Version]
- Van Gisbergen, P.A.; Li, M.; Wu, S.-Z.; Bezanilla, M. Class II formin targeting to the cell cortex by binding PI(3,5)P2 is essential for polarized growth. J. Cell Biol. 2012, 198, 235–250. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Matsuzawa, T.; Takegawa, K.; Sato, M.H. Loss-of-Function and Gain-of-Function Mutations in FAB1A/B Impair Endomembrane Homeostasis, Conferring Pleiotropic Developmental Abnormalities in Arabidopsis. Plant Physiol. 2010, 155, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Gary, J.D.; Wurmser, A.E.; Bonangelino, C.J.; Weisman, L.S.; Emr, S.D. Fab1p Is Essential for PtdIns(3)P 5-Kinase Activity and the Maintenance of Vacuolar Size and Membrane Homeostasis. J. Cell Biol. 1998, 143, 65–79. [Google Scholar] [CrossRef]
- Cooke, F.T.; Dove, S.K.; McEwen, R.K.; Painter, G.; Holmes, A.B.; Hall, M.N.; Michell, R.H.; Parker, P.J. The stress-activated phosphatidylinositol 3-phosphate 5-kinase Fab1p is essential for vacuole function in S. cerevisiae. Curr. Biol. 1998, 8, 1219–S2. [Google Scholar] [CrossRef] [Green Version]
- Suh, B.-C.; Hille, B. Regulation of ion channels by phosphatidylinositol 4,5-bisphosphate. Curr. Opin. Neurobiol. 2005, 15, 370–378. [Google Scholar] [CrossRef]
- Yao, H.-Y.; Xue, H.-W. Signals and mechanisms affecting vesicular trafficking during root growth. Curr. Opin. Plant Biol. 2011, 14, 571–579. [Google Scholar] [CrossRef]
- Hunt, L.; Otterhag, L.; Lee, J.C.; Lasheen, T.; Hunt, J.; Seki, M.; Shinozaki, K.; Sommarin, M.; Gilmour, D.J.; Pical, C.; et al. Gene-specific expression and calcium activation of Arabidopsis thaliana phospholipase C isoforms. New Phytol. 2004, 162, 643–654. [Google Scholar] [CrossRef]
- Kim, D.H.; Eu, Y.J.; Yoo, C.M.; Kim, Y.W.; Pih, K.T.; Jin, J.B.; Kim, S.J.; Stenmark, H.; Hwang, I. Trafficking of phosphatidylinositol 3-phosphate from the trans-Golgi network to the lumen of the central vacuole in plant cells. Plant Cell 2001, 13, 287–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munnik, T. Phosphatidic acid: An emerging plant lipid second messenger. Trends Plant Sci. 2001, 6, 227–233. [Google Scholar] [CrossRef]
- Di Paolo, G.; De Camilli, P. Phosphoinositides in cell regulation and membrane dynamics. Nature 2006, 443, 651–657. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, L.-J.; Huang, R.-D. Cytoskeleton and plant salt stress tolerance. Plant Signal. Behav. 2011, 6, 29–31. [Google Scholar] [CrossRef] [Green Version]
- Xuan, J.; Song, Y.; Zhang, H.; Liu, J.; Guo, Z.-R.; Hua, Y. Comparative Proteomic Analysis of the Stolon Cold Stress Response between the C4 Perennial Grass Species Zoysia japonica and Zoysia metrella. PLoS ONE 2013, 8, e75705. [Google Scholar] [CrossRef]
- Yan, S.-P.; Zhang, Q.-Y.; Tang, Z.-C.; Su, W.-A.; Sun, W.-N. Comparative Proteomic Analysis Provides New Insights into Chilling Stress Responses in Rice. Mol. Cell. Proteomics 2006, 5, 484–496. [Google Scholar] [CrossRef] [Green Version]
- Peremyslov, V.V.; Morgun, E.A.; Kurth, E.G.; Makarova, K.S.; Koonin, E.V.; Dolja, V.V. Identification of Myosin XI Receptors in Arabidopsis Defines a Distinct Class of Transport Vesicles. Plant Cell 2013, 25, 3022–3038. [Google Scholar] [CrossRef] [Green Version]
- Nick, P. Microtubules, signalling and abiotic stress. Plant J. 2013, 75, 309–323. [Google Scholar] [CrossRef]
- Smertenko, A.; Chang, H.Y.; Sonobe, S.; Fenyk, S.I.; Weingartner, M.; Bögre, L.; Hussey, P.J. Control of the AtMAP65-1 interaction with microtubules through the cell cycle. J. Cell Sci. 2006, 119, 3227–3237. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Takahashi, D.; Kawamura, Y.; Uemura, M. Plasma membrane proteome analyses of Arabidopsis thaliana suspension-cultured cells during cold or ABA treatment: Relationship with freezing tolerance and growth phase. J. Proteom. 2019, 211, 103528. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Rahman, A. Cold stress response in Arabidopsis thaliana is mediated by GNOM ARF-GEF. Plant J. 2018, 97, 500–516. [Google Scholar] [CrossRef] [Green Version]
- Minami, A.; Fujiwara, M.; Furuto, A.; Fukao, Y.; Yamashita, T.; Kamo, M.; Kawamura, Y.; Uemura, M. Alterations in Detergent-Resistant Plasma Membrane Microdomains in Arabidopsis thaliana During Cold Acclimation. Plant Cell Physiol. 2008, 50, 341–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conde, A.; Chaves, M.M.; Gerós, H. Membrane Transport, Sensing and Signaling in Plant Adaptation to Environmental Stress. Plant Cell Physiol. 2011, 52, 1583–1602. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; et al. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nat. Cell Biol. 2014, 514, 367–371. [Google Scholar] [CrossRef]
- Hou, C.; Tian, W.; Kleist, T.; He, K.; Garcia, V.; Bai, F.; Hao, Y.; Luan, S.; Li, L. DUF221 proteins are a family of osmosensitive calcium-permeable cation channels conserved across eukaryotes. Cell Res. 2014, 24, 632–635. [Google Scholar] [CrossRef] [Green Version]
- Bose, J.; Pottosin, I.I.; Shabala, S.S.; Palmgren, M.G.; Shabala, S. Calcium Efflux Systems in Stress Signaling and Adaptation in Plants. Front. Plant Sci. 2011, 2, 85. [Google Scholar] [CrossRef] [Green Version]
- Carpaneto, A.; Ivashikina, N.; Levchenko, V.; Krol, E.; Jeworutzki, E.; Zhu, J.-K.; Hedrich, R. Cold Transiently Activates Calcium-Permeable Channels in Arabidopsis Mesophyll Cells. Plant Physiol. 2006, 143, 487–494. [Google Scholar] [CrossRef] [Green Version]
- Guy, C.L.; Huber, J.L.A.; Huber, S.C. Sucrose Phosphate Synthase and Sucrose Accumulation at Low Temperature. Plant Physiol. 1992, 100, 502–508. [Google Scholar] [CrossRef] [Green Version]
- Le Hir, R.; Spinner, L.; Klemens, P.A.; Chakraborti, D.; De Marco, F.; Vilaine, F.; Wolff, N.; Lemoine, R.; Porcheron, B.; Géry, C.; et al. Disruption of the Sugar Transporters AtSWEET11 and AtSWEET12 Affects Vascular Development and Freezing Tolerance in Arabidopsis. Mol. Plant 2015, 8, 1687–1690. [Google Scholar] [CrossRef] [Green Version]
- Durand, M.; Porcheron, B.; Hennion, N.; Maurousset, L.; Lemoine, R.; Pourtau, N. Water Deficit Enhances C Export to the Roots in Arabidopsis thaliana Plants with Contribution of Sucrose Transporters in Both Shoot and Roots. Plant Physiol. 2016, 170, 1460–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, W.X.; Schneider, T.; Starck, S.; Martinoia, E.; Trentmann, O. Cold acclimation induces changes in Arabidopsis tonoplast protein abundance and activity and alters phosphorylation of tonoplast monosaccharide transporters. Plant J. 2012, 69, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Wingenter, K.; Trentmann, O.; Winschuh, I.; Hörmiller, I.I.; Heyer, A.G.; Reinders, J.; Schulz, A.; Geiger, D.; Hedrich, R.; Neuhaus, H.E. A member of the mitogen-activated protein 3-kinase family is involved in the regulation of plant vacuolar glucose uptake. Plant J. 2011, 68, 890–900. [Google Scholar] [CrossRef] [PubMed]
- Olsson, A.; Svennelid, F.; Ek, B.; Sommarin, M.; Larsson, C. A Phosphothreonine Residue at the C-Terminal End of the Plasma Membrane H+-ATPase Is Protected by Fusicoccin-Induced 14–3–3 Binding. Plant Physiol. 1998, 118, 551–555. [Google Scholar] [CrossRef] [Green Version]
- Axelsen, K.B.; Venema, K.; Jahn, T.; Baunsgaard, L.; Palmgren, M.G. Molecular Dissection of the C-Terminal Regulatory Domain of the Plant Plasma Membrane H+-ATPase AHA2: Mapping of Residues that When Altered Give Rise to an Activated Enzyme. Biochemistry 1999, 38, 7227–7234. [Google Scholar] [CrossRef]
- Haruta, M.; Sabat, G.; Stecker, K.; Minkoff, B.B.; Sussman, M.R. A Peptide Hormone and Its Receptor Protein Kinase Regulate Plant Cell Expansion. Science 2014, 343, 408–411. [Google Scholar] [CrossRef] [Green Version]
- Viotti, C.; Luoni, L.; Morandini, P.; De Michelis, M.I. Characterization of the interaction between the plasma membrane H+-ATPase of Arabidopsis thaliana and a novel interactor (PPI1). FEBS J. 2005, 272, 5864–5871. [Google Scholar] [CrossRef]
- Hwang, J.U.; Song, W.Y.; Hong, D.; Ko, D.; Yamaoka, Y.; Jang, S.; Yim, S.; Lee, E.; Khare, D.; Kim, K.; et al. Plant ABC transporters enable many unique aspects of a terrestrial plant’s lifestyle. Mol. Plant 2016, 9, 338–355. [Google Scholar]
- Kang, J.; Park, J.; Choi, H.; Burla, B.; Kretzschmar, T.; Lee, Y.; Martinoia, E. Plant ABC Transporters. Arab. Book 2011, 9, e0153. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.Y.; Jin, J.Y.; Alejandro, S.; Martinoia, E.; Lee, Y.S. Overexpression of AtABCG36 improves drought and salt stress resistance in Arabidopsis. Physiol. Plant. 2010, 139, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.-P.; Li, Z.-S.; Rea, P.A. AtMRP1 gene of Arabidopsis encodes a glutathione S-conjugate pump: Isolation and functional definition of a plant ATP-binding cassette transporter gene. Proc. Natl. Acad. Sci. USA 1997, 94, 8243–8248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecchetti, V.; Brunetti, P.; Napoli, N.; Fattorini, L.; Altamura, M.M.; Costantino, P.; Cardarelli, M. ABCB1 and ABCB19 auxin transporters have synergistic effects on early and late Arabidopsisanther development. J. Integr. Plant Biol. 2015, 57, 1089–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, D.R.; Wu, G.; Ljung, K.; Spalding, E.P. Auxin transport into cotyledons and cotyledon growth depend similarly on the ABCB19 Multidrug Resistance-like transporter. Plant J. 2009, 60, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Růžička, K.; Strader, L.C.; Bailly, A.; Yang, H.; Blakeslee, J.; Łangowski, Ł.; Nejedlá, E.; Fujita, H.; Itoh, H.; Syōno, K.; et al. Arabidopsis PIS1 encodes the ABCG37 transporter of auxinic compounds including the auxin precursor indole-3-butyric acid. Proc. Natl. Acad. Sci. USA 2010, 107, 10749–10753. [Google Scholar]
- Christie, J.M.; Yang, H.; Richter, G.L.; Sullivan, S.; Thomson, C.E.; Lin, J.; Titapiwatanakun, B.; Ennis, M.; Kaiserli, E.; Lee, O.R.; et al. phot1 Inhibition of ABCB19 Primes Lateral Auxin Fluxes in the Shoot Apex Required For Phototropism. PLoS Biol. 2011, 9, e1001076. [Google Scholar] [CrossRef]
- Shibasaki, K.; Uemura, M.; Tsurumi, S.; Rahman, A. Auxin Response in Arabidopsis under Cold Stress: Underlying Molecular Mechanisms. Plant Cell 2009, 21, 3823–3838. [Google Scholar] [CrossRef] [Green Version]
- Bielach, A.; Hrtyan, M.; Tognetti, V.B. Plants under Stress: Involvement of Auxin and Cytokinin. Int. J. Mol. Sci. 2017, 18, 1427. [Google Scholar] [CrossRef] [Green Version]
- Grunewald, W.; Friml, J. The march of the PINs: Developmental plasticity by dynamic polar targeting in plant cells. EMBO J. 2010, 29, 2700–2714. [Google Scholar] [CrossRef]
- Huang, F.; Zago, M.K.; Abas, L.; Van Marion, A.; Galván-Ampudia, C.S.; Offringa, R. Phosphorylation of Conserved PIN Motifs Directs Arabidopsis PIN1 Polarity and Auxin Transport. Plant Cell 2010, 22, 1129–1142. [Google Scholar] [CrossRef] [Green Version]
- Mulekar, J.J.; Bu, Q.; Chen, F.; Huq, E. Casein kinase II α subunits affect multiple developmental and stress-responsive pathways in Arabidopsis. Plant J. 2012, 69, 343–354. [Google Scholar] [CrossRef]
- Moreno-Romero, J.; Espunya, M.C.; Platara, M.; Ariño, J.; Martínez, M.C. A role for protein kinase CK2 in plant development: Evidence obtained using a dominant-negative mutant. Plant J. 2008, 55, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Kaldenhoff, R. On the pH Regulation of Plant Aquaporins. J. Biol. Chem. 2008, 283, 33889–33892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tournaire-Roux, C.; Sutka, M.; Javot, H.; Gout, E.; Gerbeau, P.; Luu, D.-T.; Bligny, R.; Maurel, C. Cytosolic pH regulates root water transport during anoxic stress through gating of aquaporins. Nat. Cell Biol. 2003, 425, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Shelden, M.C.; Howitt, S.M.; Kaiser, B.N.; Tyerman, S.D. Identification and functional characterisation of aquaporins in the grapevine, Vitis vinifera. Funct. Plant Biol. 2009, 36, 1065–1078. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Chung, G.C.; Jang, J.Y.; Ahn, S.J.; Zwiazek, J.J. Overexpression of PIP2;5 Aquaporin Alleviates Effects of Low Root Temperature on Cell Hydraulic Conductivity and Growth in Arabidopsis. Plant Physiol. 2012, 159, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Prak, S.; Hem, S.; Boudet, J.; Viennois, G.; Sommerer, N.; Rossignol, M.; Maurel, C.; Santoni, V. Multiple phosphorylations in the C-terminal tail of plant plasma membrane aquaporins: Role in subcellular trafficking of AtPIP2;1 in response to salt stress. Mol. Cell Proteom. 2008, 7, 1019–1030. [Google Scholar] [CrossRef] [Green Version]
- Aroca, R.F.; Amodeo, G.; Fernández-Illescas, S.; Herman, E.M.; Chaumont, F.; Chrispeels, M.J. The Role of Aquaporins and Membrane Damage in Chilling and Hydrogen Peroxide Induced Changes in the Hydraulic Conductance of Maize Roots12. Plant Physiol. 2005, 137, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Prado, K.; Boursiac, Y.; Tournaire-Roux, C.; Monneuse, J.-M.; Postaire, O.; Da Ines, O.; Schäffner, A.R.; Hem, S.; Santoni, V.; Maurel, C. Regulation of Arabidopsis Leaf Hydraulics Involves Light-Dependent Phosphorylation of Aquaporins in Veins[C][W]. Plant Cell 2013, 25, 1029–1039. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Kawamura, Y.; Maeshima, M.; Rahman, A.; Uemura, M. Plasma Membrane Aquaporin Members PIPs Act in Concert to Regulate Cold Acclimation and Freezing Tolerance Responses in Arabidopsis thaliana. Plant Cell Physiol. 2020, 61, 787–802. [Google Scholar] [CrossRef]
- Kadota, Y.; Sklenar, J.; Derbyshire, P.; Stransfeld, L.; Asai, S.; Ntoukakis, V.; Jones, J.D.; Shirasu, K.; Menke, F.; Jones, A.; et al. Direct Regulation of the NADPH Oxidase RBOHD by the PRR-Associated Kinase BIK1 during Plant Immunity. Mol. Cell 2014, 54, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Kadota, Y.; Shirasu, K.; Zipfel, C. Regulation of the NADPH Oxidase RBOHD During Plant Immunity. Plant Cell Physiol. 2015, 56, 1472–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudsocq, M.; Willmann, M.R.; McCormack, M.P.; Lee, H.; Shan, L.; He, P.; Bush, J.; Cheng, S.-H.; Sheen, J. Differential innate immune signalling via Ca2+ sensor protein kinases. Nat. Cell Biol. 2010, 464, 418–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uemura, M.; Joseph, R.A.; Steponkus, P.L. Cold Acclimation of Arabidopsis thaliana (Effect on Plasma Membrane Lipid Composition and Freeze-Induced Lesions). Plant Physiol. 1995, 109, 15–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sugiyama, N.; Masuda, T.; Shinoda, K.; Nakamura, A.; Tomita, M.; Ishihama, Y. Phosphopeptide Enrichment by Aliphatic Hydroxy Acid-modified Metal Oxide Chromatography for Nano-LC-MS/MS in Proteomics Applications. Mol. Cell. Proteom. 2007, 6, 1103–1109. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, M.K.; Nomura, Y.; Wang, L.; Nakagami, H.; Somers, D.E. Quantitative Circadian Phosphoproteomic Analysis of Arabidopsis Reveals Extensive Clock Control of Key Components in Physiological, Metabolic, and Signaling Pathways. Mol. Cell. Proteom. 2015, 14, 2243–2260. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.-Y.; Krishnakumar, V.; Chan, A.P.; Schobel, S.; Town, C. Araport11: A complete reannotation of the Arabidopsis thaliana reference genome. Plant J. 2017, 89, 047308. [Google Scholar] [CrossRef] [Green Version]
- Huala, E. The Arabidopsis Information Resource (TAIR): A comprehensive database and web-based information retrieval, analysis, and visualization system for a model plant. Nucleic Acids Res. 2001, 29, 102–105. [Google Scholar] [CrossRef] [Green Version]
- Olsen, J.V.; Ong, S.-E.; Mann, M. Trypsin Cleaves Exclusively C-terminal to Arginine and Lysine Residues. Mol. Cell. Proteom. 2004, 3, 608–614. [Google Scholar] [CrossRef] [Green Version]
- Savitski, M.M.; Lemeer, S.; Boesche, M.; Lang, M.; Mathieson, T.; Bantscheff, M.; Kuster, B. Confident Phosphorylation Site Localization Using the Mascot Delta Score. Mol. Cell. Proteom. 2011, 10, 003830. [Google Scholar] [CrossRef] [Green Version]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpievitch, Y.V.; Dabney, A.R.; Smith, R.D. Normalization and missing value imputation for label-free LC-MS analysis. BMC Bioinform. 2012, 13, S5. [Google Scholar] [CrossRef] [Green Version]
- Karpievitch, Y.V.; Taverner, T.; Adkins, J.N.; Callister, S.J.; Anderson, G.A.; Smith, R.D.; Dabney, A.R. Normalization of peak intensities in bottom-up MS-based proteomics using singular value decomposition. Bioinformatics 2009, 25, 2573–2580. [Google Scholar] [CrossRef] [PubMed]
- Stetson, L.C.; Ostrom, Q.T.; Schlatzer, D.; Liao, P.; Devine, K.; Waite, K.; Couce, M.E.; Harris, P.L.R.; Kerstetter-Fogle, A.; Berens, M.E.; et al. Proteins inform survival-based differences in patients with glioblastoma. Neurooncol. Adv. 2020, 2. [Google Scholar] [CrossRef] [PubMed]
- Karpievitch, Y.V.; Stuart, T.; Mohamed, S. ProteoMM: Multi-dataset model-based differential expression proteomics analysis platform, R package version 1.8.0. Bioconductor 2020. [Google Scholar] [CrossRef]
- Howe, E.; Holton, K.; Nair, S.; Schlauch, D.; Sinha, R.; Quackenbush, J. MeV: MultiExperiment Viewer. In Biomedical Informatics for Cancer Research; Ochs, M.F., Casagrande, J.T., Davuluri, R.V., Eds.; Springer: Boston, MA, USA, 2010; pp. 267–277. [Google Scholar]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. agriGO: A GO analysis toolkit for the agricultural community. Nucleic Acids Res. 2010, 38, W64–W70. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Huang Da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang Da, W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS: A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g: Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, C.M.; Castleden, I.R.; Tanz, S.K.; Aryamanesh, N.; Harvey Millar, A. SUBA4: The interactive data analysis centre for Arabidopsis subcellular protein locations. Nucl. Acids Res. 2017, 45, 1064–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [Green Version]
- Chou, M.F.; Schwartz, D. Biological Sequence Motif Discovery Usingmotif-x. Curr. Protoc. Bioinform. 2011, 35, 13.15.1–13.15.24. [Google Scholar] [CrossRef] [Green Version]
- Wagih, O.; Sugiyama, N.; Ishihama, Y.; Beltrao, P. Uncovering Phosphorylation-Based Specificities through Functional Interaction Networks. Mol. Cell. Proteomics 2016, 15, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TAIR Accession | TAIR Symbol | Protein Description | Localization | Bin Names of Functional Categories | Phosphopeptides | Phospho-residue | Phospho-sites | KMC Clusters |
|---|---|---|---|---|---|---|---|---|
| AT1G59610 | DRP2B | dynamin-like 3 | PM | Vesicle trafficking | RYS[+80]DPAQNGEDSSGSGGSSR | S | 883 | E,C * |
| AT1G10290 | DRP2A | dynamin-like protein 6 | G,PM | Vesicle trafficking | RYS[+80]DPAQNGDAASPGSGSNRR | S | 877 | A * |
| AT1G13980 | GN | sec7 domain-containing protein | G,PM | Vesicle trafficking | FSQLLS[+80]LDTEEPR | S | 961 | C * |
| AT5G06140 | SNX1 | sorting nexin 1 | C,PM | Vesicle trafficking | NISGSMQS[+80]PR | S | 16 | C * |
| AT3G11820 | SYP121 | syntaxin of plants 121 | PM | Vesicle trafficking | TLDRLIS[+80]TGESER | S | 190 | C * |
| AT5G08080 | SYP132 | syntaxin of plants 132 | PM | Vesicle trafficking | GQS[+80]SREGDVELGEQQGGDQGLEDFFKK | S | 16 | D * |
| AT5G61210 | SNAP33 | soluble N-ethylmaleimide-sensitive factor adaptor protein 33 | PM | Vesicle trafficking | TTS[+80]EPSLADMTNPFGGER | S | 47 | E,C * |
| AT5G58440 | SNX2A | sorting nexin 2A | G,EM | Vesicle trafficking | SPS[+80]SSSSDYIK | S | 146 | A * |
| AT1G08190 | VPS41 | vacuolar protein sorting 41 | G,EM | Vesicle trafficking | EDNNRSS[+80]FSQR | S | 860 | B * |
| AT5G06560 | MYOB7 | myosin-binding protein (Protein of unknown function 2C DUF593) | EM | unknown | FKNDTADGYAMS[+80]PR | S | 385 | E,A * |
| AT4G27500 | PPI1 | proton pump interactor 1 | ER,PM | unknown | KKTGGNTETETEEVPEAS[+80]EEEIEAPVQEEKPQK | S | 540 | E,C * |
| AT1G72160 | PATL3 | Sec14p-like phosphatidylinositol transfer family protein | PM | Solute transport | SMIPQNLGS[+80]FKEESSKLSDLSNSEK | S | 108 | E,C * |
| AT2G41560 | ACA4 | autoinhibited Ca2+-ATPase 2C isoform 4 | V,PM | Solute transport | SSVS[+80]IVKNR | S | 28 | A * |
| AT1G27770 | ACA1 | autoinhibited Ca2+-ATPase 1 | PM | Solute transport | FTANLS[+80]KR | S | 46 | A * |
| AT4G04340 | AtOSCA1.1 | ERD (early-responsive to dehydration stress) family protein | PM | Solute transport | RNT[+80]PAPSR | T | 750 | B * |
| AT4G22120 | AtOSCA1.2 | ERD (early-responsive to dehydration stress) family protein | PM | Solute transport | NTPAPSIIS[+80]GDDSPSLPFSGK | S | 757 | C * |
| AT5G23660 | SWEET12 | bidirectional sugar transporter SWEET12-like protein | PM | Solute transport | LGTLTS[+80]PEPVAITVVR | S | 248 | A * |
| AT1G08930 | ERD6 | Major facilitator superfamily protein | ER,PM | Solute transport | SLS[+80]IRER | S | 17 | A * |
| AT3G18830 | PLT5 | polyol/monosaccharide transporter 5 | PM | Solute transport | TVPNPEVEIGS[+80]NKQWKEGDTQSS | S | 527 | D * |
| AT1G20840 | TMT1 | tonoplast monosaccharide transporter1 | V,PM | Solute transport | LYGTHENQSYLARPVPEQNS[+80]SLGLR | S | 277 | E,A * |
| AT1G20840 | TMT1 | tonoplast monosaccharide transporter1 | V,PM | Solute transport | YYLKEDGAES[+80]R | S | 446 | B* |
| AT4G35300 | TMT2 | tonoplast monosaccharide transporter2 | V,PM | Solute transport | IYLHQEGFPGS[+80]RR | S | 448 | E,A * |
| AT4G35300 | TMT2 | tonoplast monosaccharide transporter2 | V,PM | Solute transport | HGS[+80]TMSR | S | 287 | D * |
| AT1G75220 | ERDL6 | Major facilitator superfamily protein | V,PM | Solute transport | RPFIHTGS[+80]WYR | S | 23 | D * |
| AT2G18960 | AHA1 | H[+]-ATPase 1 | PM | Solute transport | T[+80]LHGLQPKEDVNIFPEKGSYR | T | 881 | D * |
| AT2G18960 | AHA1 | H[+]-ATPase 1 | PM | Solute transport | GLDIDTAGHHYT[+80]V | T | 948 | D * |
| AT2G18960 | AHA1 | H[+]-ATPase 1 | PM | Solute transport | EDVNIFPEKGS[+80]YRELSEIAEQAK | S | 899 | E,D * |
| AT3G47950 | AHA4 | H[+]-ATPase 4 | PM | Solute transport | GLDIETIQQAYT[+80]V | T | 959 | E,A * |
| AT3G54820 | PIP2-5 | plasma membrane intrinsic protein 2%3B5 | PM | Solute transport | ALGS[+80]FRSQPHV | S | 279 | E,D * |
| AT2G39010 | PIP2-6 | plasma membrane intrinsic protein 2E | PM | Solute transport | AYGS[+80]VRS[+80]QLHELHA | S;S | 279;282 | D * |
| AT4G35100 | PIP2-7 | plasma membrane intrinsic protein 3 | PM | Solute transport | ALGSFRS[+80]NATN | S | 276 | C * |
| AT2G36910 | ABCB1 | ATP binding cassette subfamily B1 | PM | Solute transport | NSVSS[+80]PIMTR | S | 634 | A * |
| AT1G30400 | ABCC1 | multidrug resistance-associated protein 1 | V,PD | Solute transport | SIT[+80]LENKR | T | 1485 | A * |
| AT1G59870 | ABCG36 | ABC-2 and Plant PDR ABC-type transporter family protein | PM | Solute transport | SLS[+80]TADGNRRGEVAMGR | S | 825 | B * |
| AT3G53480 | ABCG37 | pleiotropic drug resistance 9 | PM | Solute transport | MNLS[+80]YWR | S | 1187 | C * |
| AT5G01240 | LAX1 | like AUXIN RESISTANT 1 | PM | Solute transport | QAEESIVVS[+80]GEDEVAGR | S | 14 | E,A * |
| AT5G01240 | LAX1 | like AUXIN RESISTANT 1 | PM | Solute transport | KVEDS[+80]AAEEDIDGNGGNGFSMK | S | 27 | B * |
| AT5G47910 | RBOHD | respiratory burst oxidase homologue D | PM | Redox homeostasis | VFS[+80]RRPSPAVR | S | 148 | B * |
| AT5G47910 | RBOHD | respiratory burst oxidase homologue D | PM | Redox homeostasis | TSS[+80]AAIHALKGLK | S | 163 | C * |
| AT5G47910 | RBOHD | respiratory burst oxidase homologue D | PM | Redox homeostasis | ILSQMLS[+80]QK | S | 347 | B * |
| AT3G15220 | AT3G15220 | Protein kinase superfamily protein | MT, CT | Protein modification | RQEVS[+80]PNRISQR | S | 364 | D * |
| AT5G14720 | AT5G14720 | Protein kinase superfamily protein | PM | Protein modification | YLEQTSAKQPGS[+80]PETNVDDLLQTPPATSR | S | 613 | E,C * |
| AT4G24100 | AT4G24100 | Protein kinase superfamily protein | PM | Protein modification | SDS[+80]NGNVEPVASERER | S | 654 | E,D * |
| AT4G10730 | AT4G10730 | Protein kinase superfamily protein | PM | Protein modification | KSAS[+80]VGNWILDSK | S | 579 | C * |
| AT1G63700 | YDA | Protein kinase superfamily protein | PM | Protein modification | S[+80]LPCLDSEDATNYQQK | S | 692 | D * |
| AT5G66850 | MAPKKK5 | mitogen-activated protein kinase kinase kinase 5 | PM | Protein modification | SPS[+80]AFTAVPR | S | 90 | C * |
| AT3G07980 | MAP3KE2 | mitogen-activated protein kinase kinase kinase 6 | V | Protein modification | KIS[+80]GQLDYVK | S | 925 | E,D * |
| AT3G13530 | M3KE1 | mitogen-activated protein kinase kinase kinase 7 | PM | Protein modification | S[+80]GQLDPNNPIFGQNETSSLSMIDQPDVLK | S | 788 | C,A * |
| AT5G19010 | MPK16 | mitogen-activated protein kinase 16 | PM | Protein modification | VAFNDTPTAIFWTDY[+80]VATR | Y | 189 | D,A * |
| AT1G18150 | MPK8 | Protein kinase superfamily protein | PM | Protein modification | AAAAVASTLESEEADNGGGYS[+80]AR | S | 539 | D * |
| AT2G31010 | Raf13 | Protein kinase superfamily protein | PM | Protein modification | KLSNTSHS[+80]EPNVATVFWR | S | 335 | D * |
| AT1G16270 | Raf18 | kinase superfamily with octicosapeptide | PM | Protein modification | NT[+80]LVSGGVRGTLPWMAPELLNGSSSKVSEK | T | 1024 | E *,D * |
| AT2G24360 | Raf22 | Protein kinase superfamily protein | PM | Protein modification | HYS[+80]LSVGQSVFRPGR | S | 81 | A * |
| AT5G50000 | Raf33 | Protein kinase superfamily protein | PM | Protein modification | LLDWGEEGHRS[+80]EAEIVSLR | S | 120 | C * |
| AT5G58950 | Raf36 | Protein kinase superfamily protein | PM | Protein modification | SVS[+80]PSPQMAVPDVFK | S | 101 | C * |
| AT3G58760 | Raf47 | Integrin-linked protein kinase family | PM | Protein modification | SSGS[+80]FNR | S | 468 | A * |
| AT4G35310 | CPK5 | Calcium-dependent protein kinase 5 | PM | Protein modification | NSLNIS[+80]MRDA | S | 552 | A * |
| AT1G49580 | CRK8 | Calcium-dependent protein kinase (CDPK) family protein | PM | Protein modification | TES[+80]GIFR | S | 360 | A * |
| AT4G04720 | CPK21 | calcium-dependent protein kinase 21 | PM | Protein modification | T[+80]MFANIDTDK | T | 387 | A * |
| AT2G17290 | CPK6 | Calcium-dependent protein kinase family protein | C,PM | Protein modification | NSLNIS[+80]MRDV | S | 540 | E,D * |
| AT5G12480 | CPK7 | calmodulin-domain protein kinase 7 | PM | Protein modification | FNSLS[+80]LKLMR | S | 520 | E,C * |
| AT3G10660 | CPK2 | calmodulin-domain protein kinase cdpk isoform 2 | ER,EM | Protein modification | VSSAGLRT[+80]ESVLQRK | T | 171 | C * |
| AT3G19100 | CRK2 | Protein kinase superfamily protein | PM | Protein modification | DAVLQNDDSTPAHPGKS[+80]PVR | S | 37 | D * |
| AT5G58380 | CIPK10 | SOS3-interacting protein 1 | C,PM | Protein modification | KS[+80]NGDTLEYQK | S | 421 | B * |
| AT3G09830 | PCRK1 | Protein kinase superfamily protein | PM | Protein modification | IVEASSGNGS[+80]PQLVPLNSVK | S | 377 | A * |
| AT2G01820 | TMK3 | Leucine-rich repeat protein kinase family protein | PM | Protein modification | LAPDGKYS[+80]IETR | S | 745 | A * |
| AT5G18610 | PBL27 | Protein kinase superfamily protein | PM | Protein modification | LGPVGDKTHVS[+80]TR | S | 244 | B * |
| AT1G66150 | TMK1 | transmembrane kinase 1 | PM | Protein modification | EASFKKAIDT[+80]T[+80]IDLDEET[+80]LASVHTVAELAGHCCAR | T;T;T | 825;826;833 | C * |
| AT2G39660 | BIK1 | botrytis-induced kinase1 | PM | Protein modification | LDTQYLPEEAVRMASVAVQCLS[+80]FEPKSRPTMDQVVR | S | 333 | C * |
| AT3G53380 | LECRK81 | Concanavalin A-like lectin protein kinase family protein | PM | Protein modification | QIEHDKS[+80]PEATVAAGTMGYLAPEYLLTGR | S | 530 | C * |
| AT4G23250 | EMB1290 | cysteine-rich receptor-like protein kinase 17 | PM | Protein modification | IFGVDQTVANT[+80]AR | T | 518 | D * |
| AT2G37710 | LECRK41 | receptor lectin kinase | PM | Protein modification | LYDHGSDPQTT[+80]HVVGTLGYLAPEHTR | T | 506 | D * |
| AT5G16590 | LRR1 | Leucine-rich repeat protein kinase family protein | PM | Protein modification | WVSSITEQQS[+80]PSDVFDPELTR | S | 562 | E,B * |
| AT5G41210 | GSTT1 | glutathione S-transferase THETA 1 | Per | Protein modification | REMGTLSKPGLQS[+80]KI | S | 243 | E,C * |
| AT1G72710 | CKL2 | casein kinase 1-like protein 2 | PM | Protein modification | NSGQIFNS[+80]GSLAK | S | 353 | C * |
| AT4G26540 | RGI3 | Leucine-rich repeat receptor-like protein kinase family protein | PM | Phytohormone action | NLTS[+80]ANVIGTGSSGVVYR | S | 762 | B * |
| AT2G13790 | SERK4 | somatic embryogenesis receptor-like kinase 4 | PM | Phytohormone action | LMNYNDS[+80]HVTTAVR | S | 451 | B * |
| AT5G38990 | AT5G38990 | Malectin/receptor-like protein kinase family protein | PM | Phytohormone action | VGPTSAS[+80]QTHVSTVVK | S | 683 | C * |
| AT5G59700 | AT5G59700 | Protein kinase superfamily protein | PM | Phytohormone action | ANPKSQQGLAEFRT[+80]EIEMLSQFR | T | 525 | D * |
| AT1G06390 | ASK9 | GSK3/SHAGGY-like protein kinase 1 | PM | Phytohormone action | VLVKGEPNIS[+80]YICSR | S | 229 | B * |
| AT1G01740 | BSK4 | kinase with tetratricopeptide repeat domain-containing protein | PM | Phytohormone action | SYS[+80]TNLAFTPPEYLR | S | 210 | E,C * |
| AT4G18710 | ASK7 | Protein kinase superfamily protein | PM | Phytohormone action | QLVKGEANISY[+80]ICSR | Y | 200 | D * |
| AT1G70940 | PIN3 | Auxin efflux carrier family protein | PM | Phytohormone action | ELHMFVWSSNGS[+80]PVSDR | S | 366 | D * |
| AT2G01420 | PIN4 | Auxin efflux carrier family protein | PM | Phytohormone action | MVVSDQPRKS[+80]NAR | S | 395 | E,C * |
| AT2G01420 | PIN4 | Auxin efflux carrier family protein | PM | Phytohormone action | KS[+80]GGDDIGGLDSGEGEREIEK | S | 395 | C * |
| AT1G23080 | PIN7 | Auxin efflux carrier family protein | PM | Phytohormone action | LRCNS[+80]TAELNPK | S | 431 | E,C * |
| AT1G23080 | PIN7 | Auxin efflux carrier family protein | PM | Phytohormone action | SFYGGGGTNMTPRPSNLTGAEIYSLNTT[+80]PR | T | 242 | C * |
| AT4G33240 | FAB1A | 1-phosphatidylinositol-3-phosphate 5-kinase FAB1A | G,EM | Multi-process regulation | NVS[+80]LEKLSDEKVK | S | 1143 | C * |
| AT1G71010 | FAB1C | FORMS APLOID AND BINUCLEATE CELLS 1C | C,EM | Multi-process regulation | VQS[+80]FDSAIR | S | 1184 | A * |
| AT1G49340 | PI4KA1 | Phosphatidylinositol 3- and 4-kinase family protein | PM | Multi-process regulation | LIS[+80]GAFSQAPQPEDDSFNEMLIAR | S | 1111 | D * |
| AT1G60890 | AT1G60890 | Phosphatidylinositol-4-phosphate 5-kinase family protein | PM | Multi-process regulation | AFS[+80]VGEKEVDLILPGTAR | S | 662 | C * |
| AT3G01310 | AT3G01310 | Phosphoglycerate mutase-like family protein | G,PM | Multi-process regulation | QGS[+80]GIIGTFGQSEELR | S | 358 | B * |
| AT1G22620 | SAC1 | Phosphoinositide phosphatase family protein | G,EM | Multi-process regulation | ASQLSHANTAREPS[+80]LRDLR | S | 456 | A * |
| AT2G18730 | DGK3 | diacylglycerol kinase 3 | PM | Lipid metabolism | FVAS[+80]RPSTADSKTMR | S | 18 | A * |
| AT3G08510 | PLC2 | phospholipase C 2 | PM | Lipid metabolism | RLS[+80]LSEEQLEK | S | 346 | D |
| AT5G60900 | RLK1 | receptor-like protein kinase 1 | PM | Enzyme classification | GAFGIVYKGYLEVAGGSEVT[+80]VAVKK | T | 559 | D * |
| AT1G17580 | XI-1 | myosin 1 | C,CT | Cytoskeleton organization | QQTLTIS[+80]PTTR | S | 1052 | E,D * |
| AT3G19960 | ATM1 | myosin 1 | PM | Cytoskeleton organization | S[+80]LPADYRFDGSPVSDRLENSSGASVR | S | 14 | E,C * |
| AT5G20490 | XI-K | myosin family protein with Dil | G,EM | Cytoskeleton organization | ENS[+80]GFGFLLTRK | S | 1516 | C * |
| AT1G08730 | XI-C | Myosin family protein with Dil domain-containing protein | C,CT | Cytoskeleton organization | KLHVASLVVQT[+80]GLR | T | 823 | C * |
| AT1G24764 | ATMAP70-2 | microtubule-associated proteins 70-2 | MT | Cytoskeleton organization | GTSKS[+80]FDGGTR | S | 464 | C * |
| AT4G27060 | TOR1 | ARM repeat superfamily protein | C,MT | Cytoskeleton organization | EASDGSTLS[+80]PDSASKGK | S | 370 | E,A * |
| AT2G01750 | MAP70-3 | microtubule-associated proteins 70-3 | PM | Cytoskeleton organization | MS[+80]EKLKLTENLLDS[+80]K | S;S | 125;137 | E,C * |
| AT4G29810 | ATMKK2 | MAP kinase kinase 2 | PM | Cell cycle organization | IISQLEPEVLS[+80]PIKPADDQLSLSDLDMVK | S | 56 | D * |
| AT5G55230 | ATMAP65-1 | microtubule-associated proteins 65-1 | MT | Cell cycle organization | RLS[+80]LNANQNGSR | S | 532 | C * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamal, M.M.; Ishikawa, S.; Takahashi, F.; Suzuki, K.; Kamo, M.; Umezawa, T.; Shinozaki, K.; Kawamura, Y.; Uemura, M. Large-Scale Phosphoproteomic Study of Arabidopsis Membrane Proteins Reveals Early Signaling Events in Response to Cold. Int. J. Mol. Sci. 2020, 21, 8631. https://doi.org/10.3390/ijms21228631
Kamal MM, Ishikawa S, Takahashi F, Suzuki K, Kamo M, Umezawa T, Shinozaki K, Kawamura Y, Uemura M. Large-Scale Phosphoproteomic Study of Arabidopsis Membrane Proteins Reveals Early Signaling Events in Response to Cold. International Journal of Molecular Sciences. 2020; 21(22):8631. https://doi.org/10.3390/ijms21228631
Chicago/Turabian StyleKamal, Md Mostafa, Shinnosuke Ishikawa, Fuminori Takahashi, Ko Suzuki, Masaharu Kamo, Taishi Umezawa, Kazuo Shinozaki, Yukio Kawamura, and Matsuo Uemura. 2020. "Large-Scale Phosphoproteomic Study of Arabidopsis Membrane Proteins Reveals Early Signaling Events in Response to Cold" International Journal of Molecular Sciences 21, no. 22: 8631. https://doi.org/10.3390/ijms21228631