Genome Mining Revealed a High Biosynthetic Potential for Antifungal Streptomyces sp. S-2 Isolated from Black Soot




Abstract
:1. Introduction
2. Results
2.1. The Isolation of the Streptomyces sp. S-2 Strain From Black Soot
2.2. Phenotypic Fingerprint of Streptomyces sp. S-2
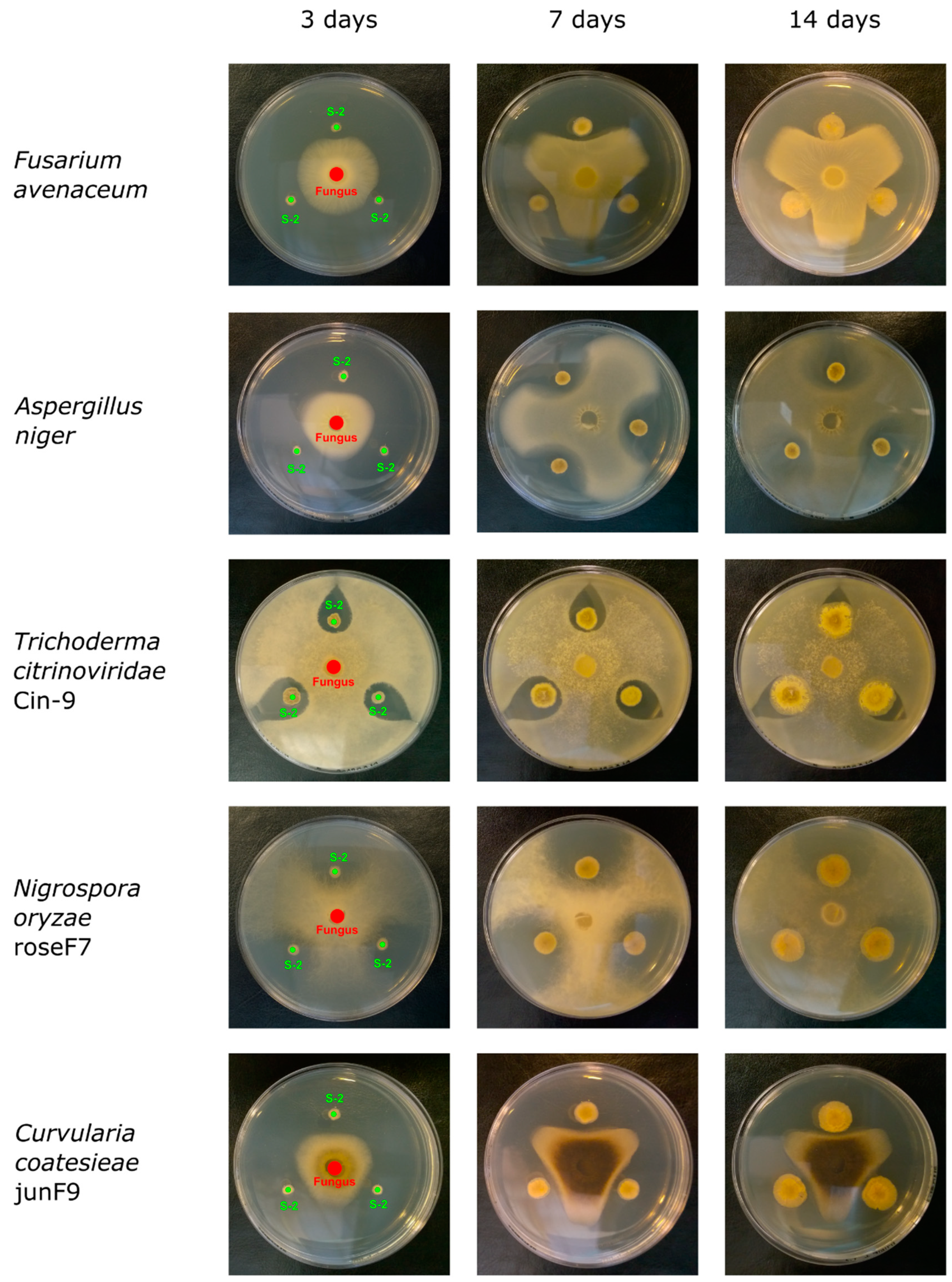
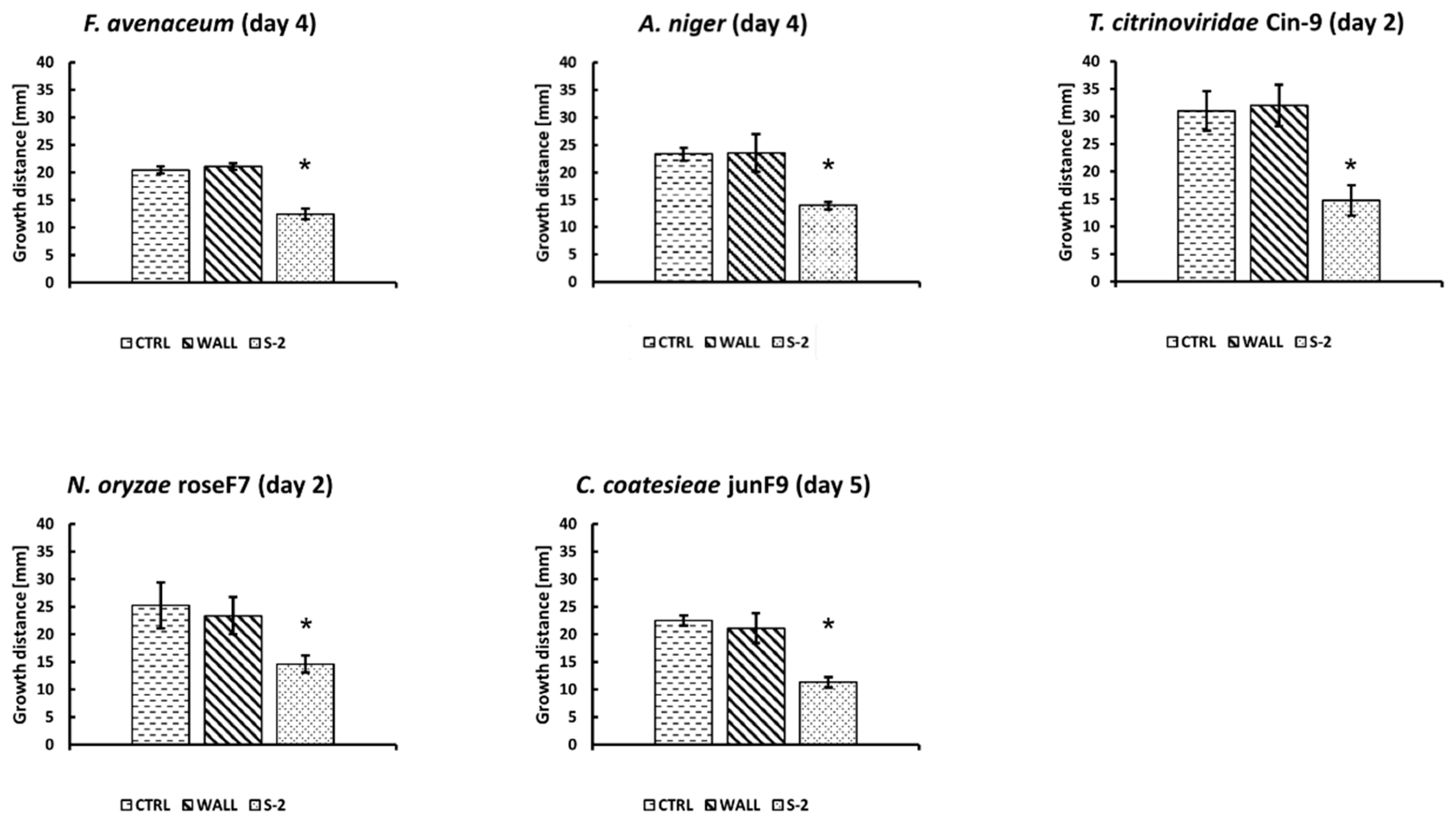
2.3. Antifungal Activity of Streptomyces sp. S-2
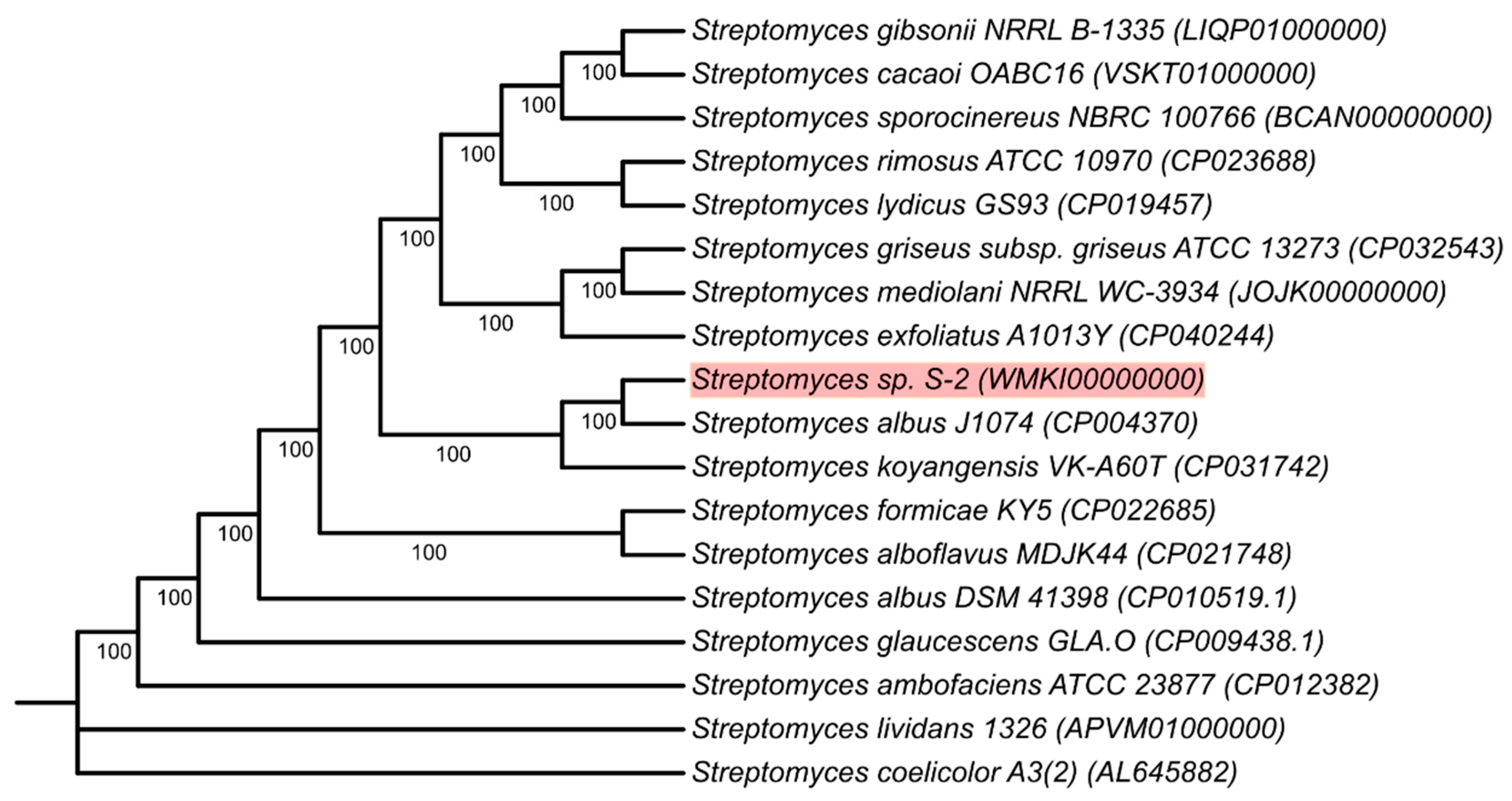
2.4. Genome Sequencing and Phylogeny of Streptomyces sp. S-2
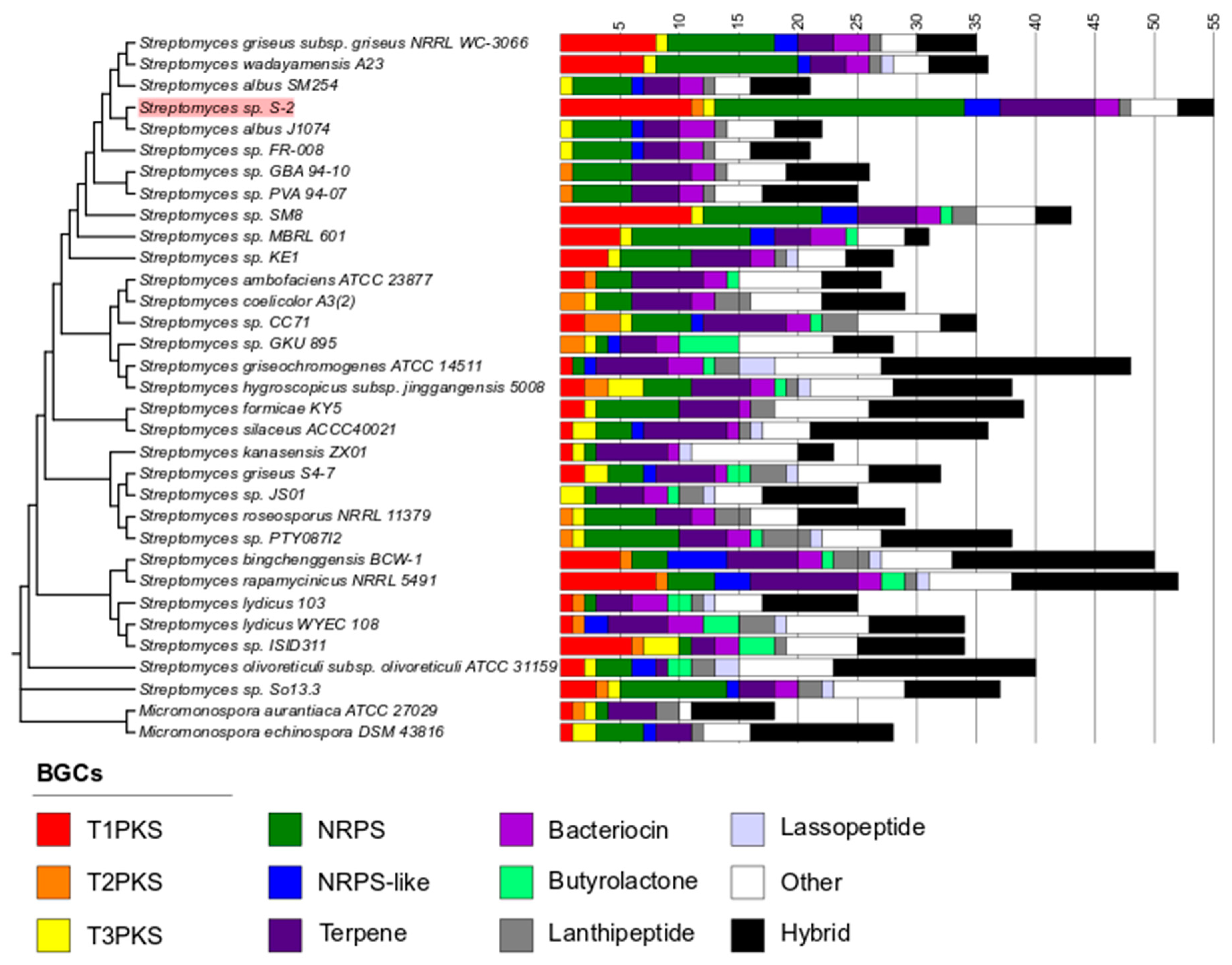
2.5. Prediction of Secondary Metabolites Biosynthetic Gene Clusters (BGCs) in the Streptomyces sp. S-2 Genome
3. Discussion
4. Materials and Methods
4.1. Microbial Culture Media
4.2. Sampling and Strains Isolation
4.3. Microorganism Strains
4.4. DNA Isolation and Sequencing
4.5. Phylogenetic Analysis of Streptomyces sp. S-2
4.6. Secondary Metabolite Gene Clusters Prediction
4.7. Phenotype Fingerprinting
4.8. Antifungal Activity Tests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BGCs | Biosynthetic gene clusters |
| NRPS | Non-ribosomal peptide synthase |
| T1PKS | Type 1 polyketide synthase |
References
- Kim, M.K.; Kang, H.J.; Roh, S.G.; Park, J.S.; Kim, S.B. Streptomyces fodineus sp. nov., an actinobacterium with antifungal activity isolated from mine area soil. Int. J. Syst. Evol. Microbiol. 2019, 69, 1350–1354. [Google Scholar] [CrossRef] [PubMed]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J.P.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, physiology, and natural products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Chater, K.F.; Biró, S.; Lee, K.J.; Palmer, T.; Schrempf, H. The complex extracellular biology of Streptomyces: Review article. FEMS Microbiol. Rev. 2010, 34, 171–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.; Tan, H. Molecular regulation of antibiotic biosynthesis in Streptomyces. Microbiol. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stulberg, E.R.; Lozano, G.L.; Morin, J.B.; Park, H.; Baraban, E.G.; Mlot, C.; Heffelfinger, C.; Phillips, G.M.; Rush, J.S.; Phillips, A.J.; et al. Genomic and secondary metabolite analyses of Streptomyces sp. 2AW provide insight into the evolution of the cycloheximide pathway. Front. Microbiol. 2016, 7, 573. [Google Scholar] [CrossRef] [Green Version]
- Bérdy, J. Thoughts and facts about antibiotics: Where we are now and where we are heading. J. Antibiot. (Tokyo) 2012, 65, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Watve, M.G.; Tickoo, R.; Jog, M.M.; Bhole, B.D. How many antibiotics are produced by the genus streptomyces? Arch. Microbiol. 2001, 176, 386–390. [Google Scholar] [CrossRef]
- Nakashima, T.; Takahashi, Y.; Ōmura, S. Search for new compounds from Kitasato microbial library by physicochemical screening. Biochem. Pharmacol. 2017, 134, 42–55. [Google Scholar] [CrossRef]
- Tiwari, K.; Gupta, R.K. Rare actinomycetes: A potential storehouse for novel antibiotics. Crit. Rev. Biotechnol. 2012, 32, 108–132. [Google Scholar] [CrossRef]
- Kumar, P.S.; Al-Dhabi, N.A.; Duraipandiyan, V.; Balachandran, C.; Kumar, P.P.; Ignacimuthu, S. In vitro antimicrobial, antioxidant and cytotoxic properties of Streptomyces lavendulae strain SCA5. BMC Microbiol. 2014, 14, 291. [Google Scholar]
- Dastager, S.G.; Li, W.J.; Dayanand, A.; Tang, S.K.; Tian, X.P.; Zhi, X.Y.; Xu, L.H.; Jiang, C.L. Seperation, identification and analysis of pigment (melanin) production in Streptomyces. African J. Biotechnol. 2006, 5, 1131–1134. [Google Scholar]
- Fajardo, A.; Martínez, J.L. Antibiotics as signals that trigger specific bacterial responses. Curr. Opin. Microbiol. 2008, 11, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Montero, K.; Lamilla, C.; Abanto, M.; Maruyama, F.; Jorquera, M.A.; Santos, A.; Martinez-Urtaza, J.; Barrientos, L. Antarctic Streptomyces fildesensis So13.3 strain as a promising source for antimicrobials discovery. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lo Giudice, A.; Bruni, V.; Michaud, L. Characterization of Antarctic psychrotrophic bacteria with antibacterial activities against terrestrial microorganisms. J. Basic Microbiol. 2007, 47, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Gulder, T.A.; Moore, B.S. Chasing the treasures of the sea—Bacterial marine natural products. Curr. Opin. Microbiol. 2009, 12, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Yu, D.; Sang, B.; Feng, J.; Han, L.; Zhang, X. Genome-wide analysis reveals the secondary metabolome in Streptomyces kanasensis ZX01. Genes 2017, 8, 346. [Google Scholar] [CrossRef] [Green Version]
- Nakaew, N.; Lumyong, S.; Sloan, W.T.; Sungthong, R. Bioactivities and genome insights of a thermotolerant antibiotics-producing Streptomyces sp. TM32 reveal its potentials for novel drug discovery. Microbiologyopen 2019, 8, e842. [Google Scholar] [CrossRef] [Green Version]
- Low, Z.J.; Pang, L.M.; Ding, Y.; Cheang, Q.W.; Hoang, K.L.M.; Tran, H.T.; Li, J.; Liu, X.W.; Kanagasundaram, Y.; Yang, L.; et al. Identification of a biosynthetic gene cluster for the polyene macrolactam sceliphrolactam in a Streptomyces strain isolated from mangrove sediment. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Ikeda, H.; Shin-Ya, K.; Omura, S. Genome mining of the Streptomyces avermitilis genome and development of genome-minimized hosts for heterologous expression of biosynthetic gene clusters. J. Ind. Microbiol. Biotechnol. 2014, 41, 233–250. [Google Scholar] [CrossRef]
- Chevrette, M.G.; Currie, C.R. Emerging evolutionary paradigms in antibiotic discovery. J. Ind. Microbiol. Biotechnol. 2019, 46, 257–271. [Google Scholar] [CrossRef]
- Rodríguez, H.; Rico, S.; Díaz, M.; Santamaría, R.I. Two-component systems in Streptomyces: Key regulators of antibiotic complex pathways. Microb. Cell Fact. 2013, 12, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, S.; Onaka, H.; Abe, I. Activation of silent biosynthetic pathways and discovery of novel secondary metabolites in actinomycetes by co-culture with mycolic acid-containing bacteria. J. Ind. Microbiol. Biotechnol. 2019, 46, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Alberti, F.; Leng, D.J.; Wilkening, I.; Song, L.; Tosin, M.; Corre, C. Triggering the expression of a silent gene cluster from genetically intractable bacteria results in scleric acid discovery. Chem. Sci. 2019, 10, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Gosse, J.T.; Ghosh, S.; Sproule, A.; Overy, D.; Cheeptham, N.; Boddy, C.N. Whole genome sequencing and metabolomic study of cave Streptomyces isolates ICC1 and ICC4. Front. Microbiol. 2019, 10, 1020. [Google Scholar] [CrossRef] [PubMed]
- Chevrette, M.G.; Carlson, C.M.; Ortega, H.E.; Thomas, C.; Ananiev, G.E.; Barns, K.J.; Book, A.J.; Cagnazzo, J.; Carlos, C.; Flanigan, W.; et al. The antimicrobial potential of Streptomyces from insect microbiomes. Nat. Commun. 2019, 10, 516. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, J.; Baker, P.; Piper, C.; Cotter, P.D.; Walsh, M.; Mooij, M.J.; Bourke, M.B.; Rea, M.C.; O’Connor, P.M.; Ross, R.P.; et al. Isolation and analysis of bacteria with antimicrobial activities from the marine sponge haliclona simulans collected from irish waters. Mar. Biotechnol. 2009, 11, 384–396. [Google Scholar] [CrossRef]
- Terra, L.; Dyson, P.J.; Hitchings, M.D.; Thomas, L.; Abdelhameed, A.; Banat, I.M.; Gazze, S.A.; Vujaklija, D.; Facey, P.D.; Francis, L.W.; et al. A novel alkaliphilic Streptomyces inhibits ESKAPE pathogens. Front. Microbiol. 2018, 9, 2458. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, M.; Gao, J.; Pu, T.; Bilal, M.; Wang, Y.; Zhang, X. Antifungal activity screening of soil actinobacteria isolated from Inner Mongolia, China. Biol. Control 2018, 127, 78–84. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Z.; Qiao, X.; Li, Z.; Li, F.; Chen, M.; Wang, Y.; Huang, Y.; Cui, H. Antifungal activity of volatile organic compounds from Streptomyces alboflavus TD-1. FEMS Microbiol. Lett. 2013, 341, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, A.; Takahashi, Y.; Omura, S. Takanawaenes, novel antifungal antibiotics produced by Streptomyces sp. K99-5278. J. Antibiot. (Tokyo) 2003, 56, 448–453. [Google Scholar]
- Yang, M.; Lu, L.; Pang, J.; Hu, Y.; Guo, Q.; Li, Z.; Wu, S.; Liu, H.; Wang, C. Biocontrol activity of volatile organic compounds from Streptomyces alboflavus TD-1 against Aspergillus flavus growth and aflatoxin production. J. Microbiol. 2019, 57, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.; Zou, L.; Zhou, D.; Chen, Y.; Gao, Z.; Feng, R.; Zhang, M.; Li, K.; Xie, J.; Wang, W. Taxonomy and broad-spectrum antifungal activity of Streptomyces sp. SCA3-4 isolated from rhizosphere soil of opuntia stricta. Front. Microbiol. 2019, 10, 1390. [Google Scholar] [CrossRef] [PubMed]
- Tchize Ndejouong, B.L.S.; Sattler, I.; Maier, A.; Kelter, G.; Menzel, K.D.; Fiebig, H.H.; Hertweck, C. Hygrobafilomycin, a cytotoxic and antifungal macrolide bearing a unique monoalkylmaleic anhydride moiety, from Streptomyces varsoviensis. J. Antibiot. (Tokyo) 2010, 63, 359–363. [Google Scholar] [CrossRef] [Green Version]
- Bubici, G.; Kaushal, M.; Prigigallo, M.I.; Cabanás, C.G.L.; Mercado-Blanco, J. Biological control agents against Fusarium wilt of banana. Front. Microbiol. 2019, 10, 616. [Google Scholar] [CrossRef] [Green Version]
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324. [Google Scholar] [CrossRef]
- Law, J.W.F.; Ser, H.L.; Khan, T.M.; Chuah, L.H.; Pusparajah, P.; Chan, K.G.; Goh, B.H.; Lee, L.H. The potential of Streptomyces as biocontrol agents against the rice blast fungus, Magnaporthe oryzae (Pyricularia oryzae). Front. Microbiol. 2017, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Laitila, A. Toxigenic Fungi and Mycotoxins in the Barley-to-Beer Chain; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; ISBN 9781782423492. [Google Scholar]
- Feng, J.; Hwang, R.; Chang, K.F.; Hwang, S.F.; Strelkov, S.E.; Gossen, B.D.; Conner, R.L.; Turnbull, G.D. Genetic variation in Fusarium avenaceum causing root rot on field pea. Plant Pathol. 2010, 59, 845–852. [Google Scholar] [CrossRef]
- Hershenhorn, J.; Park, S.H.; Stierle, A.; Strobel, G.A. Fusarium avenaceum as a novel pathogen of spotted knapweed and its phytotoxins, acetamido-butenolide and enniatin B. Plant Sci. 1992, 86, 155–160. [Google Scholar] [CrossRef]
- Satyaprasad, K.; Bateman, G.L.; Ward, E. Comparisons of isolates of Fusarium avenaceum from white lupin and other crops by pathogenicity tests, DNA analyses and vegetative compatibility tests. J. Phytopathol. 2000, 148, 211–219. [Google Scholar] [CrossRef]
- Abass, M.H.; Hameed, M.A.; Ahmed, A.N. First report of Nigrospora sphaerica (Sacc.) Mason as a potential pathogen on date palm (Phoenix dactylifera L.). Can. J. Plant Pathol. 2013, 35, 75–80. [Google Scholar] [CrossRef]
- Wang, M.; Liu, F.; Crous, P.W.; Cai, L. Phylogenetic reassessment of Nigrospora: Ubiquitous endophytes, plant and human pathogens. Pers. Mol. Phylogeny Evol. Fungi 2017, 39, 118–142. [Google Scholar] [CrossRef] [PubMed]
- Park, G.J.; Kim, J.C.; Shon, M.J.; Kim, H.T.; Cho, K.Y. Phytotoxin production of nigrospora sphaerica pathogenic on turfgrasses. Plant Pathol. J. 2000, 16, 137–141. [Google Scholar]
- Fisher, P.J.; Petrini, O. Fungal saprobes and pathogens as endophytes of rice (Oryza sativa L.). New Phytol. 1992, 120, 137–143. [Google Scholar] [CrossRef]
- Beom, J.Y.; Jung, J.A.; Lee, K.T.; Hwangbo, A.; Song, M.C.; Lee, Y.; Lee, S.J.; Oh, J.H.; Ha, S.J.; Nam, S.J.; et al. Biosynthesis of nonimmunosuppressive FK506 analogues with antifungal activity. J. Nat. Prod. 2019, 82, 2078–2086. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.W.; Kim, D.R.; Kwon, Y.S.; Kwak, Y.S. Genome-wide screening antifungal genes in Streptomyces griseus S4-7, a Fusarium wilt disease suppressive microbial agent. FEMS Microbiol. Lett. 2019, 366, fnz133. [Google Scholar] [CrossRef]
- Getha, K.; Vikineswary, S.; Wong, W.H.; Seki, T.; Ward, A.; Goodfellow, M. Evaluation of Streptomyces sp. strain g10 for suppression of Fusarium wilt and rhizosphere colonization in pot-grown banana plantlets. J. Ind. Microbiol. Biotechnol. 2005, 32, 24–32. [Google Scholar] [CrossRef]
- Mahadevan, B.; Crawford, D.L. Properties of the chitinase of the antifungal biocontrol agent Streptomyces lydicus WYEC108. Enzym. Microb. Technol. 1997, 20, 489–493. [Google Scholar] [CrossRef]
- Trejo-Estrada, S.R.; Paszczynski, A.; Crawford, D.L. Antibiotics and enzymes produced by the biocontrol agent Streptomyces violaceusniger YCED-9. J. Ind. Microbiol. Biotechnol. 1998, 21, 81–90. [Google Scholar] [CrossRef]
- Castagnoli, E.; Marik, T.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Indoor Trichoderma strains emitting peptaibols in guttation droplets. J. Appl. Microbiol. 2018, 125, 1408–1422. [Google Scholar] [CrossRef] [Green Version]
- Pasqualotto, A.C. Differences in pathogenicity and clinical syndromes due to Aspergillus fumigatus and Aspergillus flavus. Med. Mycol. 2009, 47, 261–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karkowska-Kuleta, J.; Rapala-Kozik, M.; Kozik, A. Fungi pathogenic to humans: Molecular bases of virulence of Candida albicans, Cryptococcus neoformans and Aspergillus fumigatus. Acta Biochim. Pol. 2009, 56, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A global concern for food safety, human health and their management. Front. Microbiol. 2017, 7, 2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, P.E.; Dignani, M.C.; Anaissie, E.J. Taxonomy, biology, and clinical aspects of Fusarium species. Clin. Microbiol. Rev. 1994, 7, 479–504. [Google Scholar] [CrossRef]
- Ballav, S.; Kerkar, S.; Thomas, S.; Augustine, N. Halophilic and halotolerant actinomycetes from a marine saltern of Goa, India producing anti-bacterial metabolites. J. Biosci. Bioeng. 2015, 119, 323–330. [Google Scholar] [CrossRef]
- Zenova, G.M.; Oborotov, G.V.; Norovsuren, Z.; Fedotova, A.V.; Yakovleva, L.V. Halophilic and alkaliphilic streptomycetes in salt-affected soils. Eurasian Soil Sci. 2007, 40, 1203–1207. [Google Scholar] [CrossRef]
- Bibb, M.J. Regulation of secondary metabolism in streptomycetes. Curr. Opin. Microbiol. 2005, 8, 208–215. [Google Scholar] [CrossRef]
- Van Wezel, G.P.; McDowall, K.J. The regulation of the secondary metabolism of Streptomyces: New links and experimental advances. Nat. Prod. Rep. 2011, 28, 1311–1333. [Google Scholar] [CrossRef]
- Vizcaíno, J.A.; Sanz, L.; Basilio, A.; Vicente, F.; Gutiérrez, S.; Hermosa, M.R.; Monte, E. Screening of antimicrobial activities in Trichoderma isolates representing three Trichoderma sections. Mycol. Res. 2005, 109, 1397–1406. [Google Scholar] [CrossRef]
- Sadykova, V.S.; Kurakov, A.V.; Kuvarina, A.E.; Rogozhin, E.A. Antimicrobial activity of fungi strains of Trichoderma from Middle Siberia. Appl. Biochem. Microbiol. 2015, 51, 355–361. [Google Scholar] [CrossRef]
- Shi, M.; Wang, H.N.; Xie, S.T.; Luo, Y.; Sun, C.Y.; Chen, X.L.; Zhang, Y.Z. Antimicrobial peptaibols, novel suppressors of tumor cells, targeted calcium-mediated apoptosis and autophagy in human hepatocellular carcinoma cells. Mol. Cancer 2010, 9, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remali, J.; Sarmin, N.I.M.; Ng, C.L.; Tiong, J.J.L.; Aizat, W.M.; Keong, L.K.; Zin, N.M. Genomic characterization of a new endophytic Streptomyces kebangsaanensis identifies biosynthetic pathway gene clusters for novel phenazine antibiotic production. PeerJ 2017, 2017, e3738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herdini, C.; Mubarika, S.; Hariwiyanto, B.; Wijayanti, N.; Hosoyama, A.; Yamazoe, A.; Nojiri, H.; Widada, J. Secondary bioactive metabolite gene clusters identification of anticandida-producing Streptomyces Sp. GMR22 isolated from Wanagama forest as revealed by Genome mining approach. Indones. J. Pharm. 2017, 28, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Challis, G.L. Mining microbial genomes for new natural products and biosynthetic pathways. Microbiology 2008, 154, 1555–1569. [Google Scholar] [CrossRef] [PubMed]
- Samel, S.A.; Marahiel, M.A.; Essen, L.O. How to tailor non-ribosomal peptide products-new clues about the structures and mechanisms of modifying enzymes. Mol. Biosyst. 2008, 4, 387–393. [Google Scholar] [CrossRef]
- Zhao, B.; Moody, S.C.; Hider, R.C.; Lei, L.; Kelly, S.L.; Waterman, M.R.; Lamb, D.C. Structural analysis of cytochrome P450 105N1 involved in the biosynthesis of the zincophore, coelibactin. Int. J. Mol. Sci. 2012, 13, 8500–8513. [Google Scholar] [CrossRef] [Green Version]
- Weissbach, H.; Redfield, B.G.; Beaven, V.; Katz, E. Actinomycin synthesis in washed cells of Streptomyces antibioticus. J. Biol. Chem. 1965, 240, 4377–4381. [Google Scholar]
- Schwecke, T.; Aparicio, J.F.; Molnár, I.; König, A.; Khaw, L.E.; Haydock, S.F.; Oliynyk, M.; Caffrey, P.; Cortés, J.; Lester, J.B.; et al. The biosynthetic gene cluster for the polyketide immunosuppressant rapamycin. Proc. Natl. Acad. Sci. USA 1995, 92, 7839–7843. [Google Scholar] [CrossRef] [Green Version]
- Dholakiya, R.N.; Kumar, R.; Mishra, A.; Mody, K.H.; Jha, B. Antibacterial and antioxidant activities of novel actinobacteria strain isolated from Gulf of Khambhat, Gujarat. Front. Microbiol. 2017, 8, 2420. [Google Scholar] [CrossRef]
- Tan, L.T.H.; Chan, K.G.; Khan, T.M.; Bukhari, S.I.; Saokaew, S.; Duangjai, A.; Pusparajah, P.; Lee, L.H.; Goh, B.H. Streptomyces sp. MUM212 as a source of antioxidants with radical scavenging and metal chelating properties. Front. Pharmacol. 2017, 8, 276. [Google Scholar] [CrossRef]
- Waldron, C.; Matsushima, P.; Rosteck, P.R.; Broughton, M.C.; Turner, J.; Madduri, K.; Crawford, K.P.; Merlo, D.J.; Baltz, R.H. Cloning and analysis of the spinosad biosynthetic gene cluster of Saccharopolyspora spinosa. Chem. Biol. 2001, 8, 487–499. [Google Scholar] [CrossRef] [Green Version]
- Malik, A.Y.; Ali, M.; Jamal, A.; Ali, M.I. Isolation and characterization of coal solubilizing aerobic microorganisms from salt range coal mines, Pakistan. Geomicrobiol. J. 2017, 34, 109–118. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Avram, O.; Rapoport, D.; Portugez, S.; Pupko, T. M1CR0B1AL1Z3R-a user-friendly web server for the analysis of large-scale microbial genomics data. Nucleic Acids Res. 2019, 47, W88–W92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguosly aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Medema, M.H.; Kottmann, R.; Lee, S.Y.; Weber, T. The antiSMASH database, a comprehensive database of microbial secondary metabolite biosynthetic gene clusters. Nucleic Acids Res. 2017, 45, D555–D559. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Pascal Andreu, V.; De Los Santos, E.L.C.; Del Carratore, F.; Lee, S.Y.; Medema, M.H.; Weber, T. The antiSMASH database version 2: A comprehensive resource on secondary metabolite biosynthetic gene clusters. Nucleic Acids Res. 2019, 47, D625–D630. [Google Scholar] [CrossRef] [Green Version]
- Kerepesi, C.; Bánky, D.; Grolmusz, V. AmphoraNet: The webserver implementation of the AMPHORA2 metagenomic workflow suite. Gene 2014, 533, 538–540. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Source Assay | |||||||
| Compound | OD590 | Compound | OD590 | Compound | OD590 | Compound | OD590 |
| Saccharides | |||||||
| α-D-Glucose | 0.214 ± 0.064 | D-Galactose | 0.089 ± 0.046 | D-Mannose | 0.120 ± 0.063 | D-Fructose | 0.196 ± 0.047 |
| D-Cellobiose | 0.145 ± 0.037 | Gentiobiose | 0.320 ± 0.151 | D-Turanose | 0.036 ± 0.026 | N-Acetylo-D-Glucosamine | 0.082 ± 0.44 |
| Dextrin | 0.155 ± 0.088 | ||||||
| Saccharide derivatives | |||||||
| N-Acetyl-β-D-Mannosamine | 0.077 ± 0.023 | D-Glucose-6-Phosphate | 0.086 ± 0.033 | D-Fructose-6-Phosphate | 0.063 ± 0.053 | N-Acetyl- Neuraminic Acid | 0.183 ± 0.021 |
| Mucic Acid | 0.065 ± 0.041 | ||||||
| Glycosides and polyols | |||||||
| β-D-Methyl-D-Glucoside | 0.098 ± 0.049 | D-Salicin | 0.175 ± 0.030 | Glycerol | 0.112 ± 0.048 | D-Mannitol | 0.067 ± 0.049 |
| Organic Acids | |||||||
| Formic Acid | 0.291 ± 0.082 | Acetic Acid | 0.249 ± 0.029 | Propionic Acid | 0.237 ± 0.021 | Citric Acid | 0.104 ± 0.076 |
| L-Lactic Acid | 0.315 ± 0.224 | D-Gluconic Acid | 0.595 ± 0.127 | D-Malic Acid | 0.133 ± 0.024 | L-Malic Acid | 0.420 ± 0.083 |
| Acetoacetic Acid | 0.114 ± 0.031 | Bromo-Succinic Acid | 0.193 ± 0.084 | Methyl Pyruvate | 0.218 ± 0.021 | α-Hydroxy-Butiric Acid | 0.318 ± 0.082 |
| β-Hydroxy-D,L-Butyric Acid | 0.267 ± 0.024 | α-Keto-Butyric Acid | 0.248 ± 0.042 | γ-Amino-Butyric Acid | 0.102 ± 0.003 | D-Lactic Acid Methyl Ester | 0.176 ± 0.041 |
| Amino Acids | |||||||
| L-Alanine | 0.115 ± 0.087 | L-Histidine | 0.199 ± 0.047 | L-Aspartic Acid | 0.134 ± 0.098 | L-Glutamic Acid | 0.125 ± 0.069 |
| Glycyl-L-Proline | 0.095 ± 0.020 | ||||||
| Others | |||||||
| Gelatin | 0.220 ± 0.009 | Inosine | 0.102 ± 0.026 | Tween 40 | 0.266 ± 0.102 | ||
| CHEMICAL SENSITIVITY ASSAY | |||||||
| pH 6 | 0.487 ± 0.107 | 1% NaCl | 1.018 ± 0.294 | 4% NaCl | 0.636 ± 0.151 | 8% NaCl | 0.349 ± 0.103 |
| 1% Sodium Lactate | 0.508 ± 0.146 | Guanidine HCl | 0.364 ± 0.039 | Nalidixic Acid | 0.456 ± 0.107 | Aztreonam | 0.250 ± 0.085 |
| Sodium Butyrate | 0.467 ± 0.160 | ||||||
| Fungus species | pre-cultured Streptomyces sp. S-2 | ||
|---|---|---|---|
| 3 days | 7 days | 14 days | |
| Fusarium avenaceum | ++ | + | - |
| Aspergillus niger | ++ | + | - |
| Trichoderma citrinoviridae Cin-9 | +++ | ++ | - |
| Nigrospora oryzae RoseF7 | ++ | - | - |
| Curvularia coatesieae JunF9 | ++ | + | - |
| Streptomyces sp. S-2 Draft Genome | |
|---|---|
| Number of contigs | 1799 |
| Contigs sum (bp) | 6.971375 |
| N50 | 6745 |
| Longest contig (bp) | 62.211 |
| Shortest contig (bp) | 407 |
| Average length (bp) | 3875 |
| % GC content | 73.1 |
| Number of CDS | 6680 |
| Gene Type | Span [nt] | Most Similar Biosynthetic Gene Cluster | |
|---|---|---|---|
| From | To | ||
| Bacteriocin | 1 | 8813 | - |
| Bacteriocin | 1 | 1875 | - |
| Ectoine | 1 | 4689 | Ectoine (75%) |
| Lanthipeptide | 1 | 6359 | - |
| NRPS | 1 | 14,680 | - |
| NRPS | 1 | 7664 | - |
| NRPS | 1 | 7000 | - |
| NRPS | 1 | 5847 | - |
| NRPS | 1 | 3863 | - |
| NRPS | 1 | 3784 | - |
| NRPS | 1 | 3409 | - |
| NRPS | 1 | 3289 | - |
| NRPS | 1 | 3179 | - |
| NRPS | 1 | 3061 | - |
| NRPS | 1 | 2796 | - |
| NRPS | 1 | 2701 | - |
| NRPS | 1 | 2511 | - |
| NRPS | 1 | 1858 | - |
| NRPS | 8528 | 41,043 | Diisonitrile entibiotic SF2768 (66%) |
| NRPS | 1 | 20,858 | Mannopetimycin (37%) |
| NRPS | 1 | 27,653 | Surugamide A / Surugamide D (19%) |
| NRPS | 1 | 6554 | Curacomycin (18%) |
| NRPS | 1 | 30,250 | Dechlorocuracomycin (16%) |
| NRPS | 1 | 7538 | Desotamide (9%) |
| NRPS | 1 | 10,029 | Mannopeptimycin (7%) |
| NRPS-like | 1 | 7313 | - |
| NRPS-like | 1 | 4785 | - |
| NRPS-like | 1 | 1417 | - |
| Hybrid (NRPS-like, T1PKS) | 1 | 22,016 | Levorin A3/C06690/FR-008-III/candicidin A/UNII-AP5PEF5W7U (66%) |
| siderophore | 4004 | 10,744 | - |
| siderophore | 1 | 1673 | - |
| siderophore | 1 | 10,733 | Desferrioxamine B (100%) |
| T1PKS | 1 | 18,159 | - |
| T1PKS | 1 | 6047 | - |
| T1PKS | 1 | 6025 | - |
| T1PKS | 1 | 5494 | - |
| T1PKS | 1 | 4600 | - |
| T1PKS | 1 | 3778 | - |
| T1PKS | 1 | 1980 | - |
| T1PKS | 1 | 1646 | - |
| T1PKS | 1 | 15,272 | ECO-02301 (32%) |
| T1PKS | 1 | 14,101 | Candicidin (23%) |
| T1PKS | 1 | 16,488 | Reedsmycins (20%) |
| Hybrid (T1PKS, NRPS) | 1 | 13,542 | Antimycin (20%) |
| Hybrid (T1PKS, NRPS) | 1 | 13,498 | Antimycin (13%) |
| T2PKS | 1 | 16,805 | Fredericamycin A (45%) |
| T3PKS | 1 | 8472 | Herboxidiene (7%) |
| Terpene | 1 | 4631 | - |
| Terpene | 1 | 4141 | - |
| Terpene | 1 | 2347 | Geosmin (100%) |
| Terpene | 1 | 9567 | Hopene (53%) |
| Terpene | 1 | 6983 | Isorenieratene (25%) |
| Terpene | 1 | 1604 | Hopene (15%) |
| Terpene | 1 | 1104 | Hopene (15%) |
| Terpene | 3697 | 13,999 | Phoshonoglycans (6%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siupka, P.; Piński, A.; Babicka, D.; Piotrowska-Seget, Z. Genome Mining Revealed a High Biosynthetic Potential for Antifungal Streptomyces sp. S-2 Isolated from Black Soot. Int. J. Mol. Sci. 2020, 21, 2558. https://doi.org/10.3390/ijms21072558
Siupka P, Piński A, Babicka D, Piotrowska-Seget Z. Genome Mining Revealed a High Biosynthetic Potential for Antifungal Streptomyces sp. S-2 Isolated from Black Soot. International Journal of Molecular Sciences. 2020; 21(7):2558. https://doi.org/10.3390/ijms21072558
Chicago/Turabian StyleSiupka, Piotr, Artur Piński, Dagmara Babicka, and Zofia Piotrowska-Seget. 2020. "Genome Mining Revealed a High Biosynthetic Potential for Antifungal Streptomyces sp. S-2 Isolated from Black Soot" International Journal of Molecular Sciences 21, no. 7: 2558. https://doi.org/10.3390/ijms21072558