Opposing Roles of FoxA1 and FoxA3 in Intrahepatic Cholangiocarcinoma Progression

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
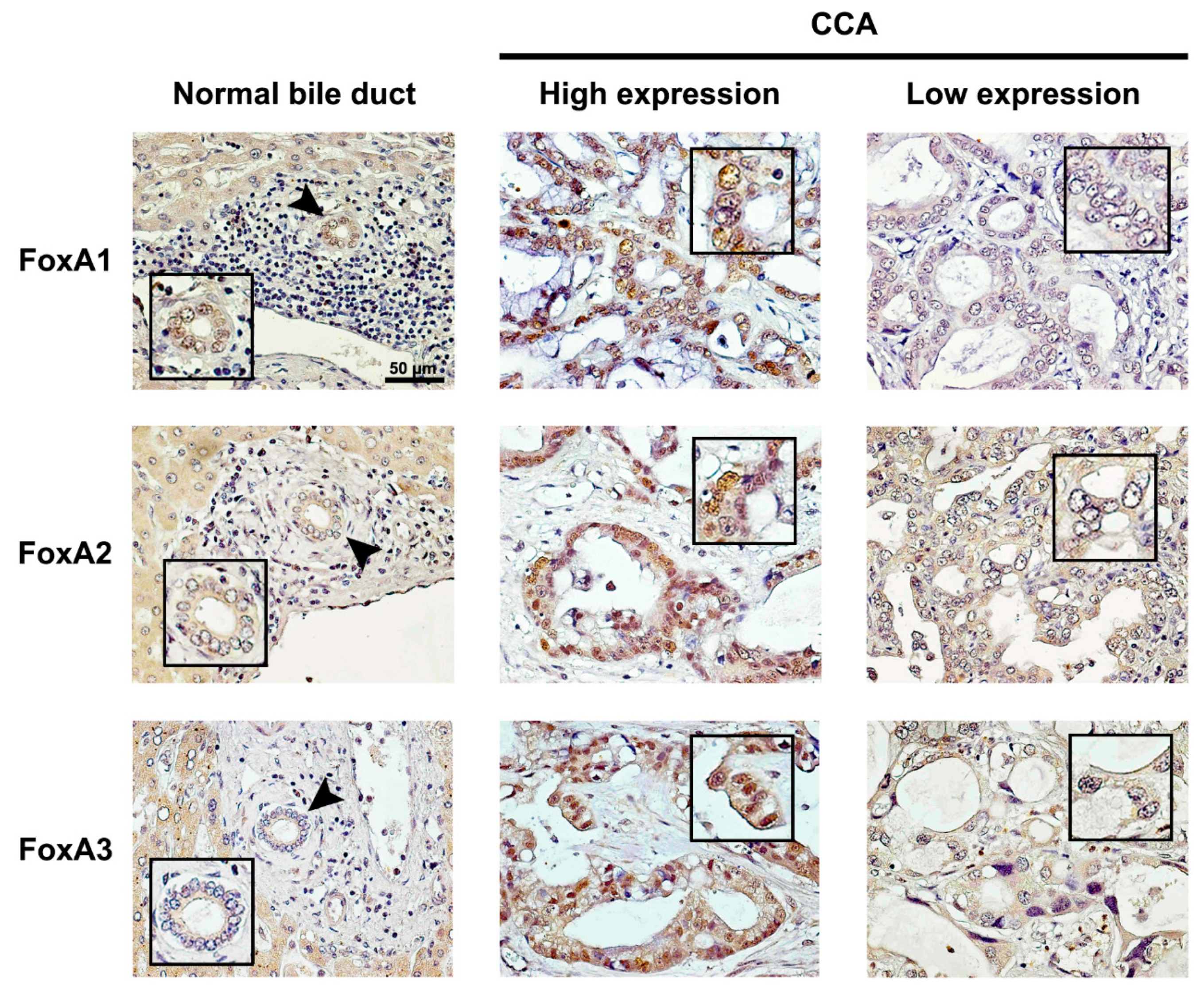
2.1. Expression of FoxAs in Intrahepatic CCA Tissues
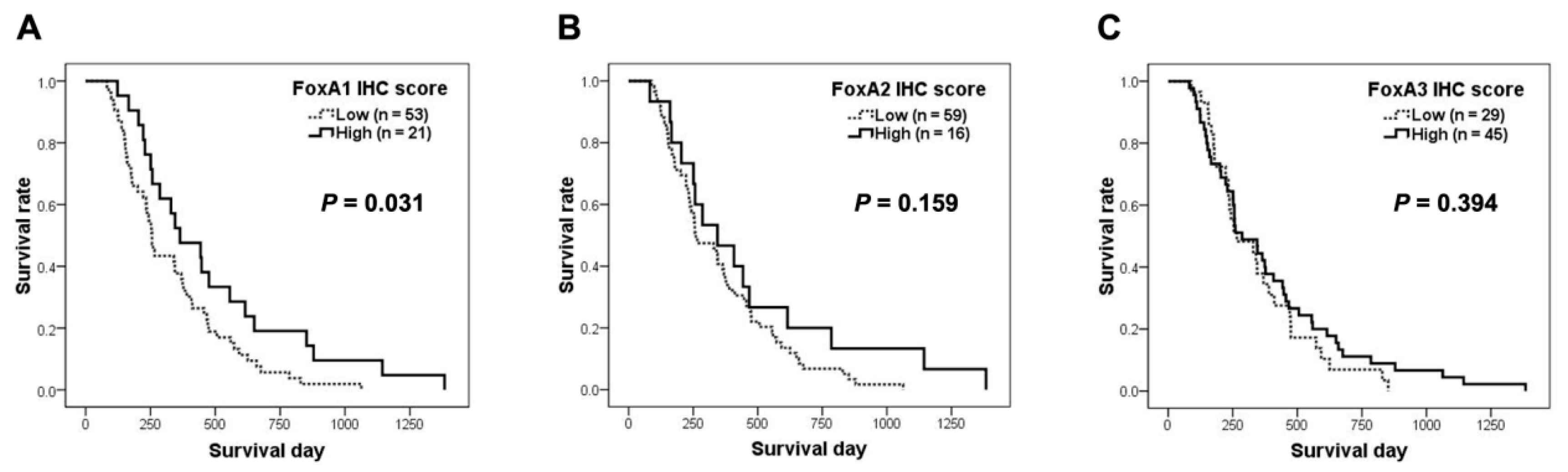
2.2. Correlations of FoxAs Expression in Intrahepatic CCA Tissues and Clinicopathological Data
2.3. Pan-Cancer Gene Expression Analysis
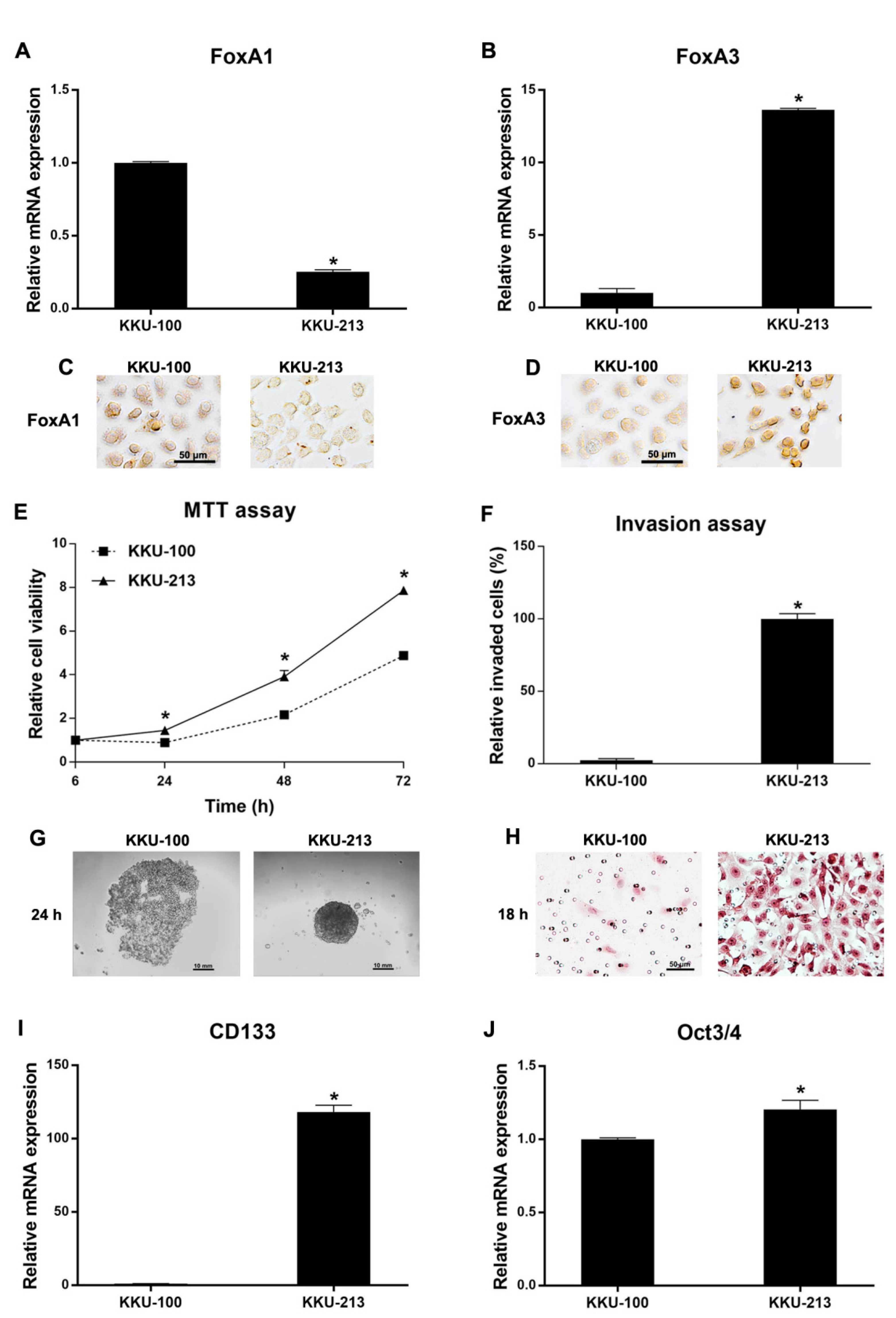
2.4. Functional Analysis of CCA Cell Lines Related to FoxA1 and FoxA3 Baseline Expression Levels
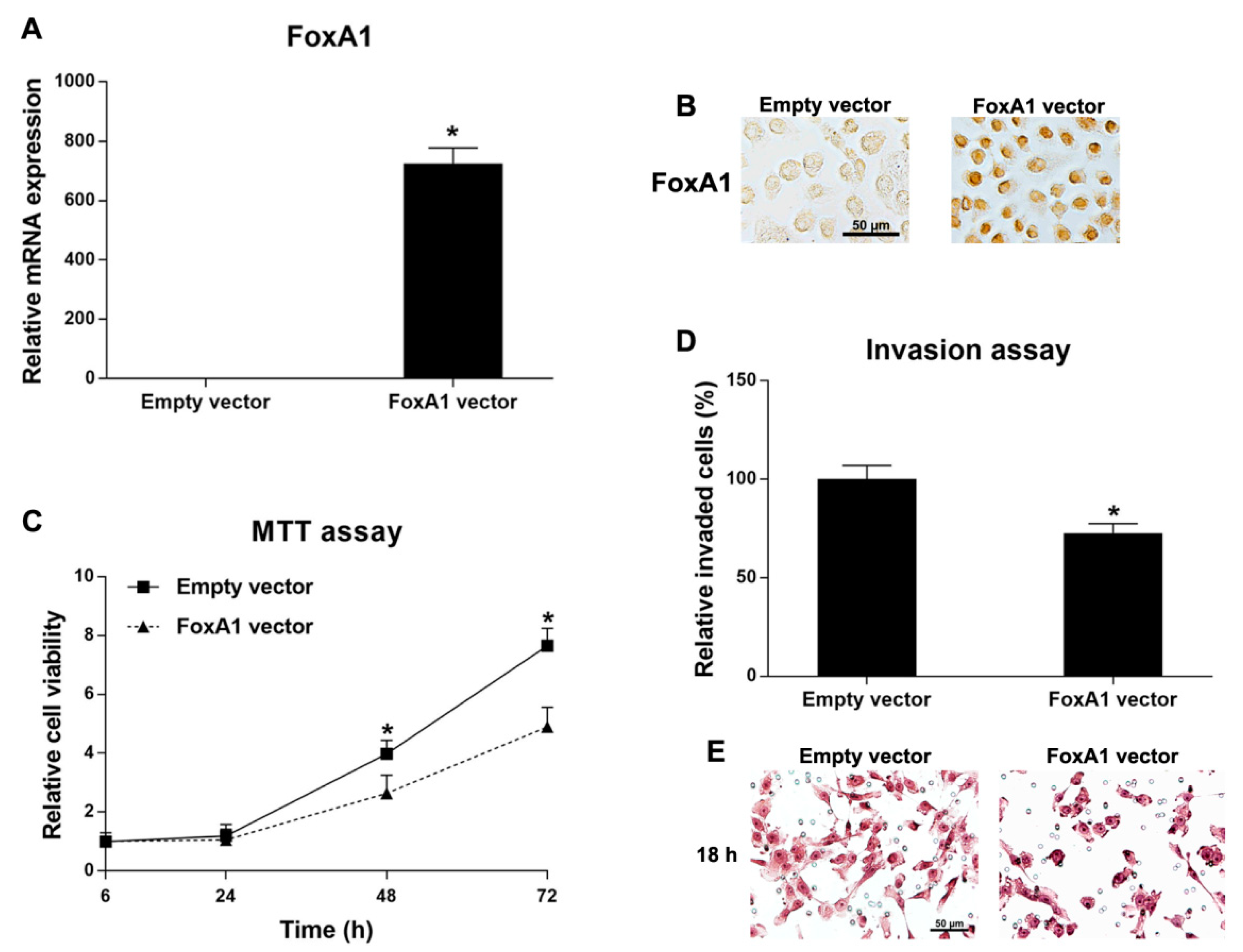
2.5. Roles of FoxA1 in CCA Cell Line
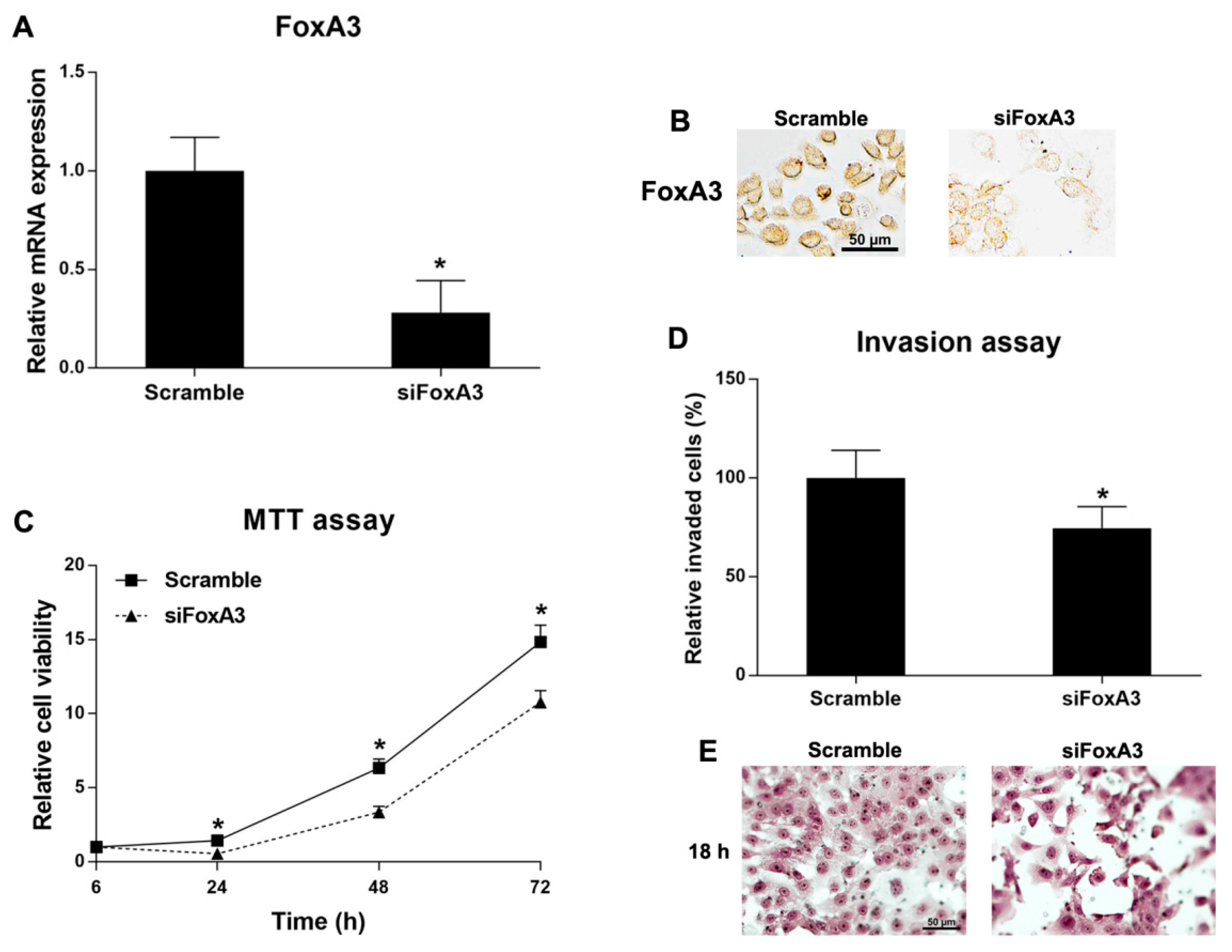
2.6. Roles of FoxA3 in CCA Cell Line
3. Discussion
4. Materials and Methods
4.1. Intrahepatic CCA Tissues
4.2. Immunohistochemistry
4.3. Pan-Cancer Gene Expression Analysis
4.4. Cell Lines
4.5. Immunocytochemistry
4.6. RNA Extraction and Real-Time PCR
4.7. FoxA1 Overexpression in CCA Cell Line
4.8. FoxA3 Knockdown Using Specific siRNA
4.9. Cell Proliferation Assay
4.10. Cell Invasion Assay
4.11. Spheroid Formation
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Golson, M.L.; Kaestner, K.H. Fox transcription factors: From development to disease. Development 2016, 143, 4558–4570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H. Targeting forkhead box transcription factors FOXM1 and FOXO in leukemia (Review). Oncol. Rep. 2014, 32, 1327–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardo, G.M.; Keri, R.A. FOXA1: A transcription factor with parallel functions in development and cancer. Biosci. Rep. 2012, 32, 113–130. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.R.; Kaestner, K.H. The Foxa family of transcription factors in development and metabolism. Cell Mol. Life Sci. 2006, 63, 2317–2328. [Google Scholar] [CrossRef]
- Chaya, D.; Hayamizu, T.; Bustin, M.; Zaret, K.S. Transcription factor FoxA (HNF3) on a nucleosome at an enhancer complex in liver chromatin. J. Biol. Chem. 2001, 276, 44385–44389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.S.; Friedman, J.R.; Fulmer, J.T.; Kaestner, K.H. The initiation of liver development is dependent on Foxa transcription factors. Nature 2005, 435, 944–947. [Google Scholar] [CrossRef]
- Duncan, A.W.; Dorrell, C.; Grompe, M. Stem cells and liver regeneration. Gastroenterology 2009, 137, 466–481. [Google Scholar] [CrossRef] [Green Version]
- Thanan, R.; Pairojkul, C.; Pinlaor, S.; Khuntikeo, N.; Wongkham, C.; Sripa, B.; Ma, N.; Vaeteewoottacharn, K.; Furukawa, A.; Kobayashi, H.; et al. Inflammation-related DNA damage and expression of CD133 and Oct3/4 in cholangiocarcinoma patients with poor prognosis. Free Radic. Biol. Med. 2013, 65, 1464–1472. [Google Scholar] [CrossRef]
- Tyson, G.L.; El-Serag, H.B. Risk factors for cholangiocarcinoma. Hepatology 2011, 54, 173–184. [Google Scholar] [CrossRef]
- Songserm, N.; Promthet, S.; Sithithaworn, P.; Pientong, C.; Ekalaksananan, T.; Chopjitt, P.; Parkin, D.M. Risk factors for cholangiocarcinoma in high-risk area of Thailand: Role of lifestyle, diet and methylenetetrahydrofolate reductase polymorphisms. Cancer Epidemiol. 2012, 36, e89–e94. [Google Scholar] [CrossRef]
- Greenbaum, L.E.; Wells, R.G. The role of stem cells in liver repair and fibrosis. Int. J. Biochem. Cell Biol. 2011, 43, 222–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armartmuntree, N.; Murata, M.; Techasen, A.; Yongvanit, P.; Loilome, W.; Namwat, N.; Pairojkul, C.; Sakonsinsiri, C.; Pinlaor, S.; Thanan, R. Prolonged oxidative stress down-regulates Early B cell factor 1 with inhibition of its tumor suppressive function against cholangiocarcinoma genesis. Redox Biol. 2018, 14, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Thanan, R.; Techasen, A.; Hou, B.; Jamnongkan, W.; Armartmuntree, N.; Yongvanit, P.; Murata, M. Development and characterization of a hydrogen peroxide-resistant cholangiocyte cell line: A novel model of oxidative stress-related cholangiocarcinoma genesis. Biochem. Biophys. Res. Commun. 2015, 464, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Rangel, N.; Fortunati, N.; Osella-Abate, S.; Annaratone, L.; Isella, C.; Catalano, M.G.; Rinella, L.; Metovic, J.; Boldorini, R.; Balmativola, D.; et al. FOXA1 and AR in invasive breast cancer: New findings on their co-expression and impact on prognosis in ER-positive patients. BMC Cancer 2018, 18, 703. [Google Scholar] [CrossRef]
- Wang, B.; Liu, G.; Ding, L.; Zhao, J.; Lu, Y. FOXA2 promotes the proliferation, migration and invasion, and epithelial mesenchymal transition in colon cancer. Exp. Ther. Med. 2018, 16, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Liu, J.; Xiong, B.; Yonemura, Y.; Yang, X. Expression and prognosis analyses of forkhead box A (FOXA) family in human lung cancer. Gene 2019, 685, 202–210. [Google Scholar] [CrossRef]
- Chen, B.; Yu, J.; Lu, L.; Dong, F.; Zhou, F.; Tao, X.; Sun, E. Upregulated forkhead-box A3 elevates the expression of forkhead-box A1 and forkhead-box A2 to promote metastasis in esophageal cancer. Oncol. Lett. 2019, 17, 4351–4360. [Google Scholar] [CrossRef] [Green Version]
- Kaewlert, W.; Sakonsinsiri, C.; Namwat, N.; Sawanyawisuth, K.; Ungarreevittaya, P.; Khuntikeo, N.; Armartmuntree, N.; Thanan, R. The Importance of CYP19A1 in Estrogen Receptor-Positive Cholangiocarcinoma. Horm. Cancer 2018, 9, 408–419. [Google Scholar] [CrossRef]
- Vaquero, J.; Guedj, N.; Claperon, A.; Nguyen Ho-Bouldoires, T.H.; Paradis, V.; Fouassier, L. Epithelial-mesenchymal transition in cholangiocarcinoma: From clinical evidence to regulatory networks. J. Hepatol. 2017, 66, 424–441. [Google Scholar] [CrossRef] [Green Version]
- Vij, K.; Wang, H.L. Aberrant expression of alpha-fetoprotein in intrahepatic cholangiocarcinoma: An exceptional occurrence. Int. J. Surg. Pathol. 2008, 16, 194–198. [Google Scholar] [CrossRef]
- Zhou, Y.M.; Yang, J.M.; Li, B.; Yin, Z.F.; Xu, F.; Wang, B.; Liu, P.; Li, Z.M. Clinicopathologic characteristics of intrahepatic cholangiocarcinoma in patients with positive serum a-fetoprotein. World J. Gastroenterol 2008, 14, 2251–2254. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research, N.; Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic. Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulouarn, C.; Factor, V.M.; Andersen, J.B.; Durkin, M.E.; Thorgeirsson, S.S. Loss of miR-122 expression in liver cancer correlates with suppression of the hepatic phenotype and gain of metastatic properties. Oncogene 2009, 28, 3526–3536. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Washington, M.K.; Crawford, H.C. Loss of FOXA1/2 is essential for the epithelial-to-mesenchymal transition in pancreatic cancer. Cancer Res. 2010, 70, 2115–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neben, K.; Schnittger, S.; Brors, B.; Tews, B.; Kokocinski, F.; Haferlach, T.; Muller, J.; Hahn, M.; Hiddemann, W.; Eils, R.; et al. Distinct gene expression patterns associated with FLT3- and NRAS-activating mutations in acute myeloid leukemia with normal karyotype. Oncogene 2005, 24, 1580–1588. [Google Scholar] [CrossRef] [Green Version]
- Badve, S.; Turbin, D.; Thorat, M.A.; Morimiya, A.; Nielsen, T.O.; Perou, C.M.; Dunn, S.; Huntsman, D.G.; Nakshatri, H. FOXA1 expression in breast cancer--correlation with luminal subtype A and survival. Clin. Cancer Res. 2007, 13, 4415–4421. [Google Scholar] [CrossRef] [Green Version]
- Jain, R.K.; Mehta, R.J.; Nakshatri, H.; Idrees, M.T.; Badve, S.S. High-level expression of forkhead-box protein A1 in metastatic prostate cancer. Histopathology 2011, 58, 766–772. [Google Scholar] [CrossRef]
- Lin, L.; Miller, C.T.; Contreras, J.I.; Prescott, M.S.; Dagenais, S.L.; Wu, R.; Yee, J.; Orringer, M.B.; Misek, D.E.; Hanash, S.M.; et al. The hepatocyte nuclear factor 3 alpha gene, HNF3alpha (FOXA1), on chromosome band 14q13 is amplified and overexpressed in esophageal and lung adenocarcinomas. Cancer Res. 2002, 62, 5273–5279. [Google Scholar]
- Xu, Z.; Liu, G.; Zhang, M.; Zhang, Z.; Jia, Y.; Peng, L.; Zhu, Y.; Hu, J.; Huang, R.; Sun, X. miR-122-5p Inhibits the Proliferation, Invasion and Growth of Bile Duct Carcinoma Cells by Targeting ALDOA. Cell Physiol. Biochem. 2018, 48, 2596–2606. [Google Scholar] [CrossRef]
- Ma, X.; Xu, L.; Gavrilova, O.; Mueller, E. Role of forkhead box protein A3 in age-associated metabolic decline. Proc. Natl. Acad Sci. USA 2014, 111, 14289–14294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takashima, Y.; Horisawa, K.; Udono, M.; Ohkawa, Y.; Suzuki, A. Prolonged inhibition of hepatocellular carcinoma cell proliferation by combinatorial expression of defined transcription factors. Cancer Sci. 2018, 109, 3543–3553. [Google Scholar] [CrossRef] [PubMed]
- Sripa, B.; Leungwattanawanit, S.; Nitta, T.; Wongkham, C.; Bhudhisawasdi, V.; Puapairoj, A.; Sripa, C.; Miwa, M. Establishment and characterization of an opisthorchiasis-associated cholangiocarcinoma cell line (KKU-100). World J. Gastroenterol. 2005, 11, 3392–3397. [Google Scholar] [CrossRef] [PubMed]
- Foty, R. A simple hanging drop cell culture protocol for generation of 3D spheroids. J. Vis. Exp. 2011. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Data | FoxA1 | FoxA2 | FoxA3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Low (n = 53) | High (n = 21) | p-value | Low (n = 59) | High (n = 15) | p-value | Low (n = 29) | High (n = 45) | p-value | |
| Age | |||||||||
| <57 | 26 | 10 | 0.911 * | 32 | 4 | 0.056 * | 20 | 16 | 0.005 * |
| ≥57 | 27 | 11 | 27 | 11 | 9 | 29 | |||
| Sex | |||||||||
| Male | 37 | 14 | 0.792 * | 41 | 10 | 0.833 * | 19 | 32 | 0.612 * |
| Female | 16 | 7 | 18 | 5 | 10 | 13 | |||
| Metastasis status | |||||||||
| Non-metastasis | 24 | 9 | 0.850 * | 27 | 6 | 0.688 * | 18 | 15 | 0.015 * |
| Metastasis | 29 | 12 | 32 | 9 | 11 | 30 | |||
| Median survival (days) | 256 | 364 | 0.031 # | 260 | 344 | 0.159 # | 266 | 286 | 0.394 # |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thanan, R.; Kaewlert, W.; Sakonsinsiri, C.; Chaiprasert, T.; Armartmuntree, N.; Muengsaen, D.; Techasen, A.; Klanrit, P.; Lert-itthiporn, W.; Pinlaor, S.; et al. Opposing Roles of FoxA1 and FoxA3 in Intrahepatic Cholangiocarcinoma Progression. Int. J. Mol. Sci. 2020, 21, 1796. https://doi.org/10.3390/ijms21051796
Thanan R, Kaewlert W, Sakonsinsiri C, Chaiprasert T, Armartmuntree N, Muengsaen D, Techasen A, Klanrit P, Lert-itthiporn W, Pinlaor S, et al. Opposing Roles of FoxA1 and FoxA3 in Intrahepatic Cholangiocarcinoma Progression. International Journal of Molecular Sciences. 2020; 21(5):1796. https://doi.org/10.3390/ijms21051796
Chicago/Turabian StyleThanan, Raynoo, Waleeporn Kaewlert, Chadamas Sakonsinsiri, Timpika Chaiprasert, Napat Armartmuntree, Duangkamon Muengsaen, Anchalee Techasen, Poramate Klanrit, Worachart Lert-itthiporn, Somchai Pinlaor, and et al. 2020. "Opposing Roles of FoxA1 and FoxA3 in Intrahepatic Cholangiocarcinoma Progression" International Journal of Molecular Sciences 21, no. 5: 1796. https://doi.org/10.3390/ijms21051796