1. Introduction
Prematurity (delivery before or at 37 weeks gestation) is the leading cause of death in newborn infants, and one in nine babies in the United States are born prematurely [
1]. Infants who survive preterm birth often face serious and lifelong health problems, including lung diseases, vision loss, and neurodevelopmental disorders. An Institute of Medicine report in 2007 estimated that premature birth costs the American healthcare system at least USD 26 billion a year [
2]. Despite its major impact on infant health, spontaneous preterm birth (SPTB) remains a significant and poorly understood perinatal complication. SPTB includes preterm labor, preterm spontaneous rupture of membranes, preterm premature rupture of membranes and cervical weakness; it does not include indicated preterm delivery for maternal or fetal conditions. While the exact etiology of SPTB remains unknown, there are likely many factors that contribute, including uterine distension, cervical insufficiency, vascular disorders, chorioamnionitis, and placental dysfunction [
3,
4]. Placental dysfunction is classically associated with fetal growth restriction and preeclampsia; however, emerging evidence supports the concept that placental insufficiency is also associated with a significant proportion of preterm births, especially early preterm births as well as those complicated by chorioamnionitis [
5]. Therefore, obtaining a better understanding of factors that contribute to placental dysfunction may lead to a deeper understanding of the etiology of prematurity and open the door for preventative treatment.
During gestation, the placenta supports growth and development of the fetus by facilitating nutrient transport and gas exchange. It produces and releases hormones into both the maternal and fetal circulation to affect uterine function, maternal metabolism, fetal growth, and development. A wide variety of metabolites are produced by the placenta, many of which are involved in energy production [
6,
7]. The placenta also functions to protect the fetus against xenobiotic molecules, infections, toxins, and maternal diseases [
8]. Therefore, a well-functioning placenta is crucial for normal gestation. Healthy term birth is associated with a growing fetus with metabolic demands that gradually begin to exceed the supply capacity of an aging placenta [
9]. We hypothesize that deficits in the capacity of the placenta to maintain bioenergetic and metabolic stability throughout the course of pregnancy may ultimately result in SPTB.
To test this hypothesis, we performed a metabolomic analysis of placenta samples obtained from women with spontaneous preterm deliveries to identify metabolic pathways that may be altered in preterm birth. The aim of our study is to identify novel metabolites and metabolic pathways in the placenta that are altered in the setting of SPTB that will provide insight into the underlying mechanisms driving this process.
3. Discussion
We report significant and biologically relevant alterations in the metabolome of SPTB placenta compared to term controls. A major limitation in spontaneous preterm birth research is the lack of human gestational controls. We attempted to address this limitation by investigating acylcarnitine metabolites in Rhesus macaque preterm and term placenta. While this is not a perfect control, our results show that at least for acylcarnitine metabolism, gestational age is not a factor determining differences between SPTB and term placentas.
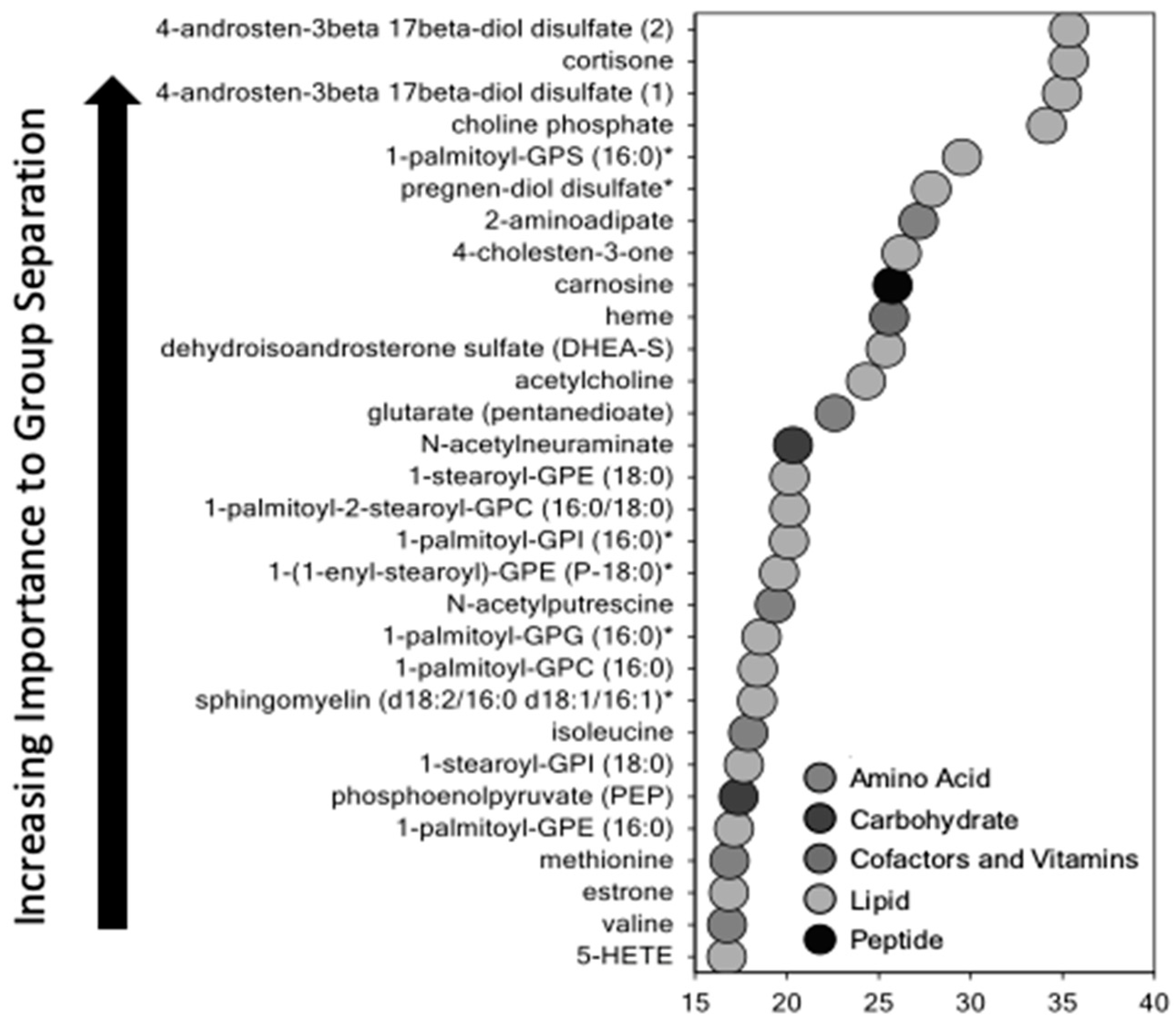
Using untargeted metabolomic analysis, we identified multiple pathways that were altered in the SPTB placenta. Levels of sphingolipids, prostaglandins, progesterone, amino acids, and metabolites involved in fatty acid oxidation were significantly different between SPTB and term placenta. Each class of metabolites that were altered may play a role in placental dysfunction through energy failure, inflammation, changes in vasculogenesis, early senescence, and maternal-fetal tolerance. All warrant further investigation to determine whether these changes are causal or merely associated with spontaneous preterm birth.
Unexpectedly, SPTB placentae had higher levels of progesterone and lower levels of estrogen and estrogen precursors than controls. The biochemical basis for these differences is not known but could reflect alterations in placental and/or fetal adrenal mitochondrial function discussed below that affect cholesterol side-chain cleavage or secretion of steroid hormones. Cholesterol side-chain cleavage, the first committed step in steroidogenesis, takes place in mitochondria. Steroid hormone production is dependent on precursors provided by the fetus and mother to form an integrated fetal–placental–maternal steroidogenic unit [
16]. Our observations suggest that the changes in fatty acid oxidation in the SPTB placenta could increase cholesterol side-chain cleavage, thereby increasing progesterone production. The role of placenta estrogens in human SPTB is unclear and remains to be determined.
The most significant changes in SPTB placenta were observed in lipid metabolites. Fatty acids are actively transported into the placenta and are key for normal fetal development [
17]. They serve as an energy-yielding substrate and play an important role in fetal energy utilization and placental function. However, when elevated, fatty acids can induce inflammation leading to preterm labor or premature rupture of membranes. Excess inflammation had been proposed as a major cause of SPTB. Of note, was our finding that multiple 2-hydroxy long chain fatty acids were elevated in SPTB placenta. These fatty acids are potent uncouplers of oxidative phosphorylation and have been shown to impair energy homeostasis [
18] as well as induce the mitochondria permeability transition pore [
19], resulting in significant mitochondrial dysfunction. This finding is in keeping with previous studies showing an altered redox state and oxidative stress in SPTB placenta [
20].
Acylcarnitine pathways were markedly disrupted in SPTB placenta. Acylcarnitines are intermediate oxidative metabolites consisting of a fatty acyl group esterified to a carnitine moiety that facilitates its transport across the mitochondrial membrane for β-oxidation [
21]. In general, acylcarnitines transport long-chain fatty acids into mitochondria and generate acetyl CoA and ultimately ATP but their direct function in the placenta has not been well characterized. Previous studies have demonstrated that the human placenta expresses high levels of enzymes involved in fatty acid oxidation [
7]. Accumulating data suggest that acylcarnitines can activate classical proinflammatory signaling pathways, engage pattern recognition receptor (PRR)-associated pathways, and induce the expression of cyclooxygenase-2, resulting in uncontrolled inflammation [
22]. Acylcarnitines can also induce mitochondrial dysfunction [
21]. This ongoing inflammation and oxidative stress can ultimately lead to severe placental dysfunction, and or disruption of fetal membranes, resulting in preterm birth. Further, elevated circulating levels of free carnitine and several short-chain, medium-chain, and long-chain acylcarnitines have been observed in the setting of adverse pregnancy complications such as gestational diabetes and preeclampsia, both of which are associated with placental dysfunction [
21,
23].
It has been hypothesized that impaired mitochondrial ß-oxidation is the underlying mechanism leading to accumulation of acylcarnitine species [
24,
25], which is consistent with our finding of impaired fatty acid oxidation in SPTB placentas. Fatty acid oxidation is a multi-step process in which a fatty acid acyl-CoA is oxidized to yield an acyl-CoA that is 2 carbons shorter, yielding NADH and FADH2 to enter the electron transport chain, as well as an acetyl-CoA molecule that enters the Krebs cycle. Defects in this process can result in the build-up of metabolic intermediaries of the pathways involved.
Our finding that acylcarnitine metabolites were elevated in SPTB placentas in both the global metabolomic analysis and in a separate set of placenta samples from CRIB that underwent a more sensitive assay for acyl carnitine profiling was a key finding. Furthermore, the lack of acylcarnitine metabolite elevation in the Rhesus monkey placenta suggests that the findings in the SPTB human placentas are not a result of a gestational age effect. However, we acknowledge that Rhesus monkey placenta is not the ideal control, and this is a major limiting factor in our study. Not surprisingly, one of the many struggles of SPTB research is determining whether significant differences between SPTB samples and control samples are due to pathology or are merely the result normal gestation. While some studies have used placentae from medically-indicated preterm births as gestational controls, almost all of these pregnancies in our population are complicated by pre-eclampsia and the metabolic profile of these placentas overlap significantly with SPTB, making their use as controls problematic. Given the obvious difficulty in obtaining preterm gestation age-matched normal placenta samples from humans, we opted to use Rhesus monkey placentas to further evaluate the effect of gestational age on placenta acylcarnitine concentrations as this was the most significantly affected pathway. A study by Eidem et al. compared the transcriptome of human SPTB to term human transcriptome and then overlaid results comparing gestational age-matched controls to term Rhesus macaque placentas after cesarean section without labor [
26]. They identified 37 GA specific and 29 SPTB specific candidate genes. While interspecies transcriptome comparison requires careful interpretation, without a better alternative for GA controls, the use of macaque placenta can help to address this major limitation of preterm birth research. We adapted this approach to evaluate our own finding of altered acylcarnitine metabolites.
A second important finding was the marked elevation of sphingolipids in SPTB placenta. Sphingolipids are a large class of bioactive membrane lipids, and in the placenta, they are critically important in regulating angiogenesis, blood vessel stability, and decidualization [
27]. Additionally, their metabolism plays an important role in maintaining maternal–fetal tolerance during pregnancy [
27,
28]. Animal studies have demonstrated that mice with a defect in sphingosine kinase have increased chemokine production, which promotes neutrophil infiltration and decreased decidual NK cells, leading to feto-maternal intolerance and pregnancy loss [
28]. Thus, an altered balance in sphingolipid metabolites may induce placental dysfunction, leading to early labor.
Prostaglandins were also markedly elevated in SPTB compared to term placenta. Prostaglandins are derived from arachidonic acid through a pathway catalyzed by COX2. Activation of the prostaglandin pathway in the fetal placental unit plays a key role in the initiation of labor by increasing sensitivity to oxytocin and triggering myometrial contractions, cervical remodeling, and extracellular matrix degradation through a paracrine effect, leading to rupture of membranes [
29].
The finding of elevated progesterone in the preterm birth placenta was unexpected since typical levels of progesterone increase with gestation. Of note, 4 of the 12 of the cases received 17-hydroxyprogesterone caproate as a prematurity prevention treatment during pregnancy, and none of the controls received this treatment.
We observed an overall increase in all classes of amino acid metabolites and peptides in SPTB placenta. Amino acids (AA) are actively transported across the placenta from mother to fetus through a number of well-characterized AA transporters. Impaired mitochondrial metabolism of some amino acids, in particular the branched chain amino acids, can also result in the accumulation of short-chain acylcarnitines. Amino acids are also metabolized in the placenta and the fetus. Thus, while elevated levels of AA in the placenta may be a result of an increase in transport activity, it may also reflect increased catabolism within the placenta.
Taken together, our data demonstrate that spontaneous preterm birth is associated with significant metabolic changes in the placenta, as reflected in increased catabolism and markers of oxidative stress consistent with early placental senescence. These changes are consistent with placenta insufficiency, which in turn results in an inability to meet the metabolic demands of the fetus.
The metabolomic analysis of this study is hypothesis-generating, identifying pathways that warrant further interrogation. Additional studies need to be performed to determine if and how these pathways contribute to SPTB. A major strength of our study was the use of an unbiased broad approach to identify significant patterns of metabolic changes in SPTB placenta with adjustment for multiple hypothesis testing. As a result, we have established a rich source of data for future studies examining other pathways that may play a role in spontaneous preterm birth. Furthermore, we performed focused pathway-specific validation of the acylcarnitine changes from the metabolomics analysis. The addition of a functional experiment measuring fatty acid oxidation further validates our findings and provides a mechanism to explain the elevated steady state levels that were observed in SPTB placenta.
A challenge of studying the placenta is the heterogeneity of the placental structure. We standardized our approach by sampling and preparing the specimens consistently from the maternal side of the placenta. While different cell types have different metabolic signatures, these cell types interact and function as a placental unit, sampling multiple cell types allows us to examine the placenta as a unit.
There are several limitations to this study. One limitation is the lack of human gestational age control, as noted above. While this is a major challenge that exists in spontaneous preterm birth research, previous work by Eidem et al. showed that Rhesus macaque transcriptome can help to disentangle changes associated with GA differences and those associated with spontaneous birth [
26]. They classified gene candidates as specific to SPTB or GA. Based on this prior work, we used Rhesus macaque placenta to validate our findings, specifically for alterations in the acylcarnitine pathway, the results of which supported the fact that SPTB, and not GA, leads to these changes. In the study by Eidem et al., they identified ACSL3 as an SPTB specific gene. ACSL3 encodes acyl CoA synthetase long chain family member [
23]. The function of this enzyme is to convert free long-chain fatty acids into fatty acyl-CoA esters and thereby plays a key role in lipid biosynthesis and fatty acid degradation.
An additional limitation of our study was the small sample size. We were unable to fully assess the impact of sex of the offspring, among other variables in four cases of IUGR. The increased percentage of IUGR in the cases may reflect underlying placental dysfunction in these cases and may reflect a suboptimal environment with placental dysfunction reflected by poor fetal growth, which is also associated with early labor. In fact, IUGR occurs in a substantial proportion of SPTB, suggesting that the etiology of fetal growth failure and SPTB overlap. Given the major population disparities in preterm birth, larger studies will be necessary to determine whether these findings can be generalized beyond the primarily African American population that was studied. Furthermore, while we evaluated FAO function with palmitate, further studies looking at mitochondrial function and fatty oxidation using other substrates may help characterize the changes in fatty acid oxidation that were observed. SPTB is a broad term that encompasses a variety of etiologies. As we develop better phenotypic classifications, larger studies will also allow for further stratification and can determine whether specific metabolic fingerprints reflect the underlying cause or phenotype of SPTB [
3].
In conclusion, this study provides an overview of metabolic pathways in the placenta that are disrupted in the setting of SPTB. Many of these pathways have been associated with placental dysfunction in the setting of other diseases of pregnancy, including preeclampsia, intrauterine growth restriction, and gestational diabetes. Growth restriction, preeclampsia, preterm birth, and to an extreme, fetal demise likely reflect a continuum of placental dysfunction. In fact, histological features of placental insufficiency occur in SPTB and are similar to preeclampsia and most cases of fetal growth restriction [
30]. How placental failure initiates SPTB remains to be determined, but associated mitochondria dysfunction and oxidative stress result in immune activation which could act on fetal membranes and or the cervix to initiate labor.
As investigators continue to characterize spontaneous preterm birth, we expect to find heterogeneous phenotypes that all lead to a final common pathway of SPTB. While our metabolic findings may not be unique to SPTB, they do support the idea that a dysfunctional placenta with inadequate compensation cannot sustain a healthy pregnancy. Therefore, placental dysfunction represents an important etiology of spontaneous preterm birth. These findings will lead us to further explore how dysfunctional lipid metabolism through fatty acid oxidation in the placenta plays a major role in metabolic disruption in the setting of preterm birth. Finally, it remains to be determined whether the changes in key metabolic pathways are causal or associated with SPTB.

 ,
, 


{kind=link}
{kind=link}