Mapping and Identifying a Candidate Gene Plr4, a Recessive Gene Regulating Purple Leaf in Rice, by Using Bulked Segregant and Transcriptome Analysis with Next-Generation Sequencing

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Statistical Analysis of Phenotypes and Genetics of Rice’s Purple Leaf Trait
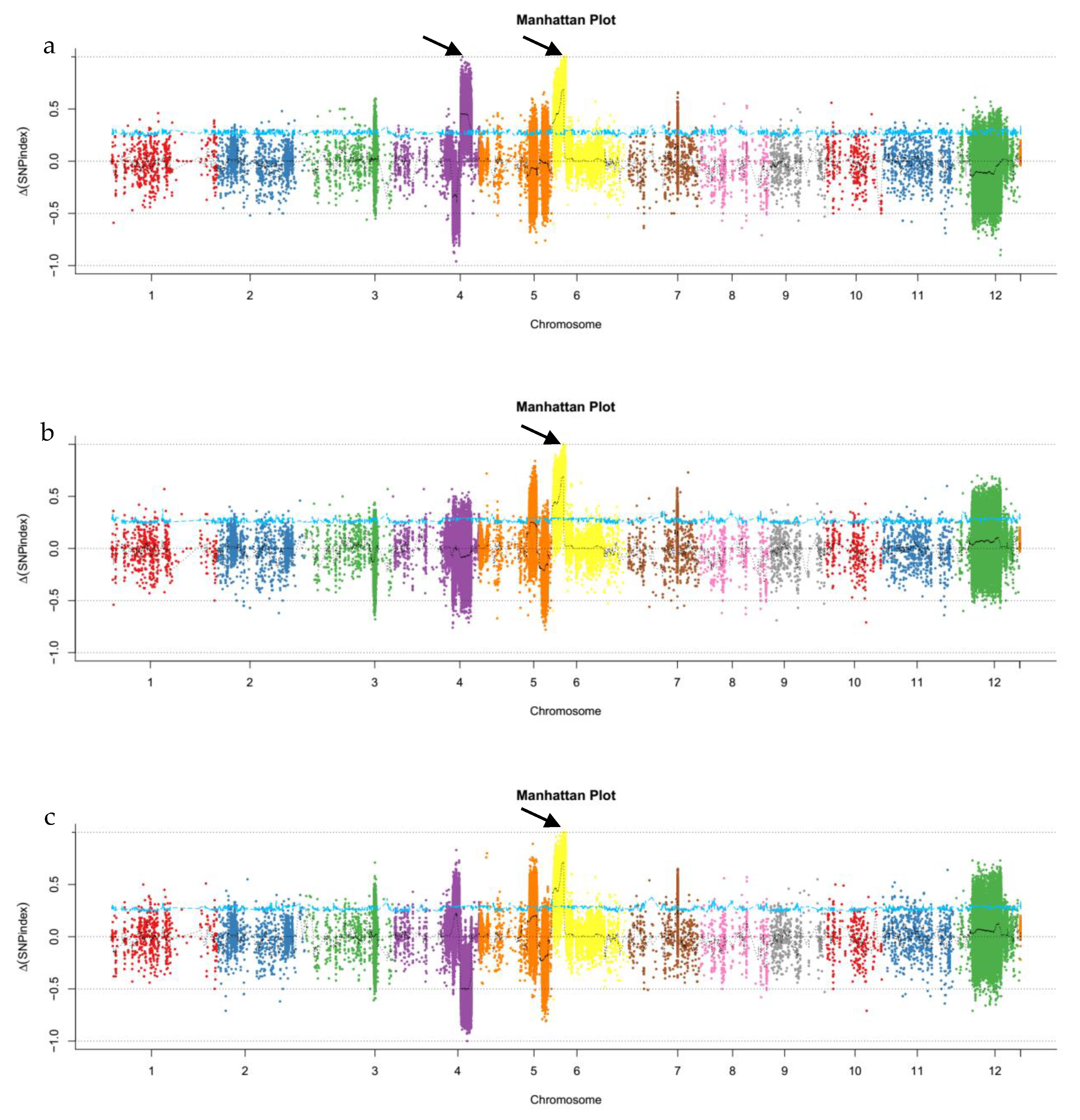
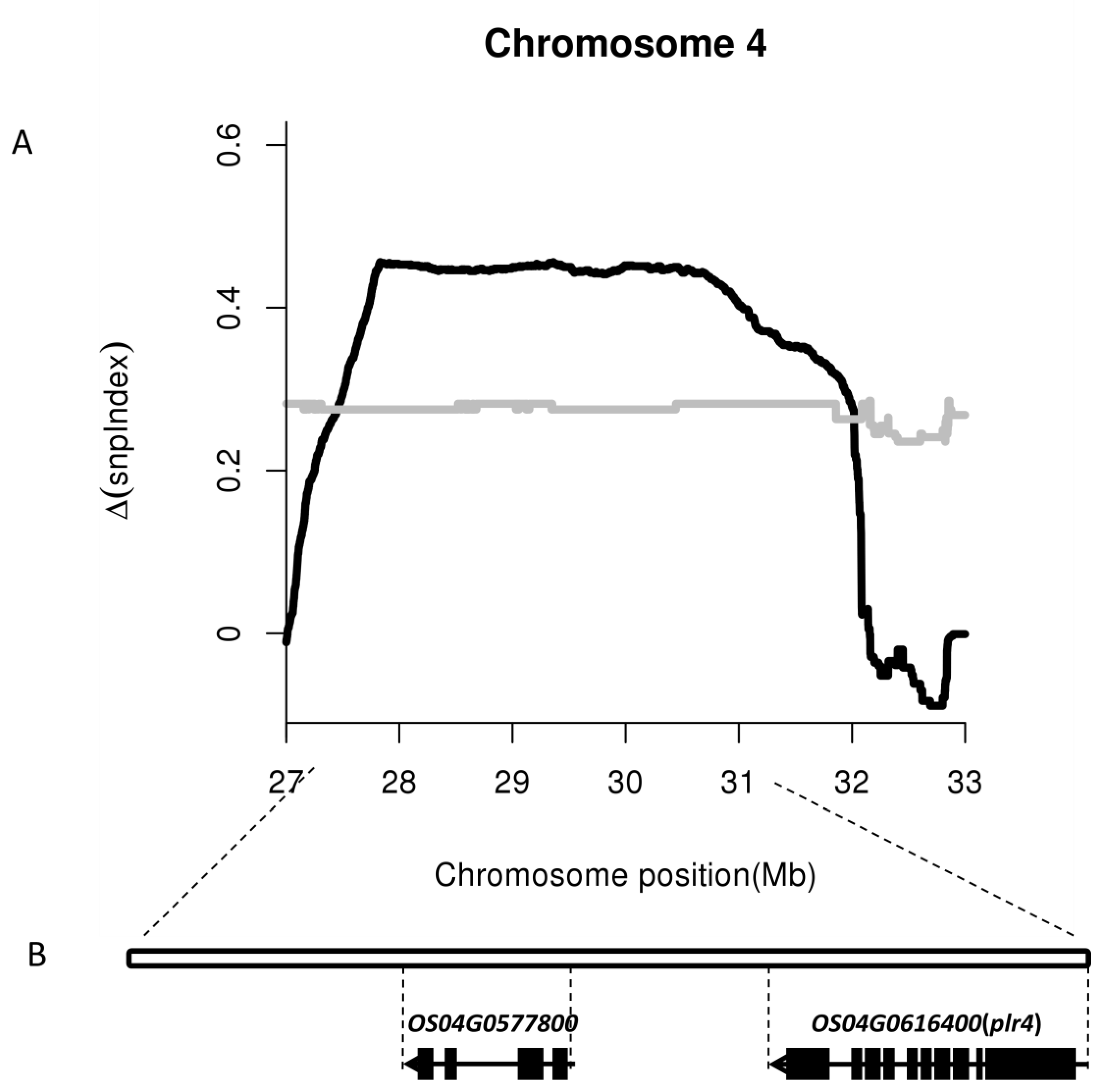
2.2. Identification of Candidate Intervals via Whole-Genome Resequencing Analysis
2.3. Identification of Expressed Genes in a Single Candidate Interval via Transcriptome Sequencing
2.4. Verification of DEGs via qRT-PCR
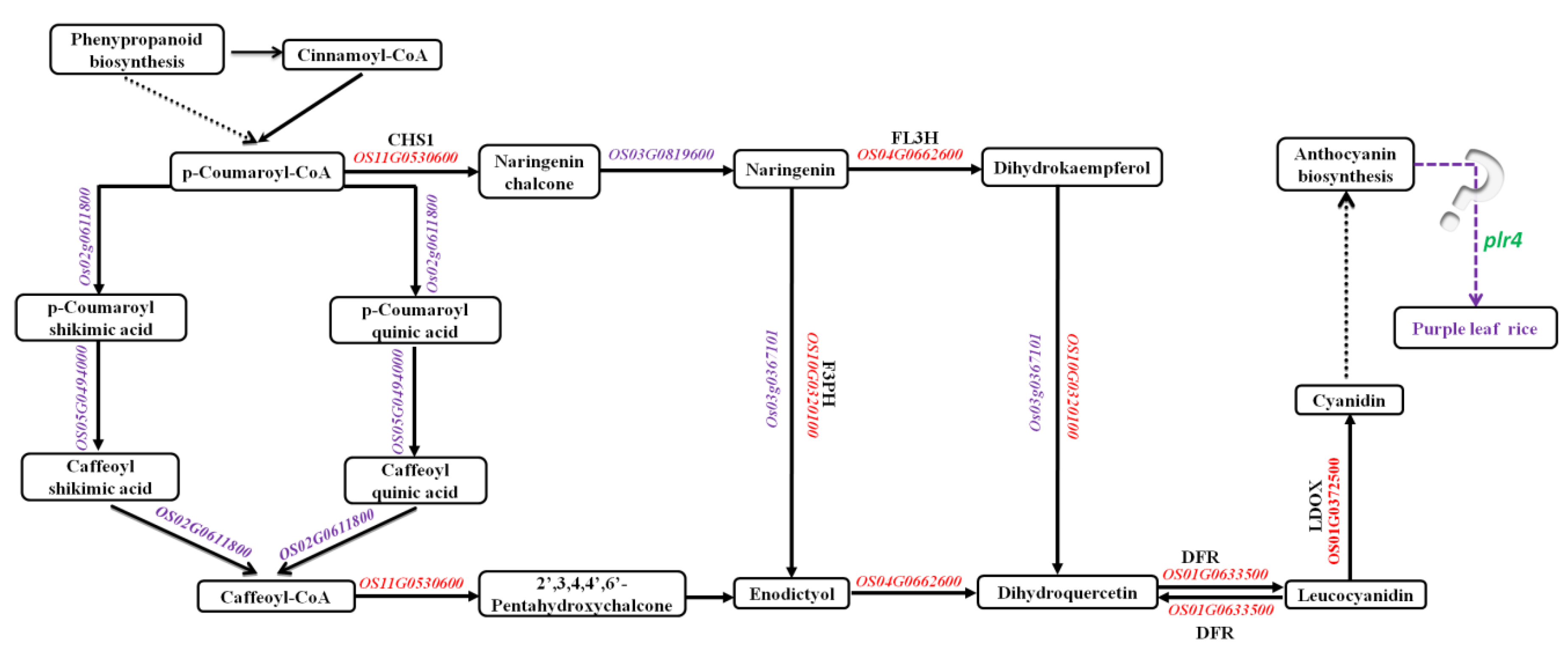
2.5. Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment of DEGs in the Anthocyanin Biosynthesis Pathway
3. Discussion
4. Materials and Methods
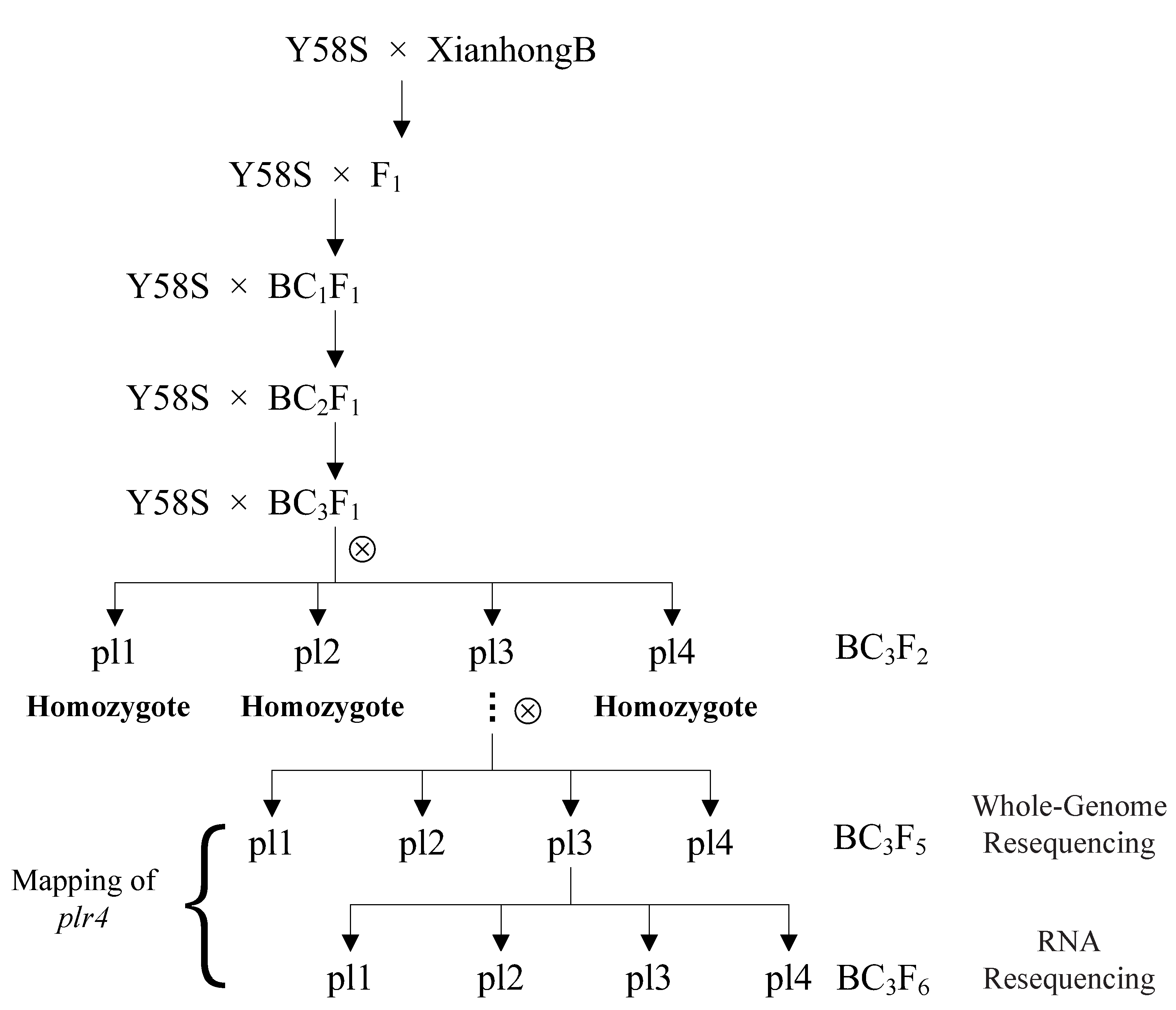
4.1. Materials and Population Construction
4.2. DNA Isolation and Analysis of WGS Data
4.3. RNA Extraction and Illumina Sequencing
4.4. Quantification of Gene Expression Level and Differential Expression Analysis
4.5. GO and KEGG Enrichment Analysis of DEGs
4.6. qRT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| qRT-PCR | Quantitative real time-PCR |
| NILs | Near isogeniclines |
| BSA-Seq | Bulked segregant analysis with a next-generation sequencing |
| RNA-Seq | Transcriptome sequencing |
| FDR | False discovery rate |
| DEGs | Differentially expressed genes |
References
- Austin, M.B.; Noel, J.P. The chalcone synthase superfamily of type III polyketide synthases. Nat. Prod. Rep. 2003, 20, 79–110. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, M.; Nakajima, J.; Yamanashi, M.; Sugiyama, M.; Makita, Y.; Springob, K.; Awazuhara, M.; Saito, K. Metabolomics and differential gene expression in anthocyanin chemo-varietal forms of Perilla frutescens. Phytochemistry 2003, 62, 987–995. [Google Scholar] [CrossRef]
- Bogs, J.; Ebadi, A.; McDavid, D.; Robinson, S.P. Identification of the flavonoid hydroxylases from grapevine and their regulation during fruit development. Plant Physiol. 2006, 140, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.T.; Goto-Yamamoto, N.; Hashizume, K.; Esaka, M. Expression of the flavonoid 3′-hydroxylase and flavonoid 3′,5′-hydroxylase genes and flavonoid composition in grape (Vitis vinifera). Plant Sci. 2006, 170, 61–69. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Di Gaspero, G.; Marconi, R.; Nonis, A.; Peterlunger, E.; Paillard, S.; Adam-Blondon, A.F.; Testolin, R. Colour variation in red grapevines (Vitis vinifera L.): Genomic organisation, expression of flavonoid 3′-hydroxylase, flavonoid 3′,5′-hydroxylase genes and related metabolite profiling of red cyanidin-/blue delphinidin-based anthocyanins in berry skin. BMC Genom. 2006, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Shimada, S.; Inoue, Y.T.; Sakuta, M. Anthocyanidin synthase in non-anthocyanin-producing Caryophyllales species. Plant J. 2005, 44, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuka, T.; Nishihara, M.; Mishiba, K.; Yamamura, S. Temporal expression of flavonoid biosynthesis-related genes regulates flower pigmentation in gentian plants. Plant Sci. 2005, 168, 1309–1318. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. It takes a garden. How work on diverse plant species has contributed to an understanding of flavonoid metabolism. Plant Physiol. 2001, 127, 1399–1404. [Google Scholar] [CrossRef]
- Ramsay, N.A.; Glover, B.J. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends Plant Sci. 2005, 10, 63–70. [Google Scholar] [CrossRef]
- Xie, D.Y.; Sharma, S.B.; Wright, E.; Wang, Z.Y.; Dixon, R.A. Metabolic engineering of proanthocyanidins through co-expression of anthocyanidin reductase and the PAP1 MYB transcription factor. Plant J. 2006, 45, 895–907. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, S.R.; Wessler, S.R. Maize R gene family: Tissue-specific helix-loop-helix proteins. Cell 1990, 62, 849–851. [Google Scholar] [CrossRef]
- Reddy, A.R.; Scheffler, B.; Madhuri, G.; Srivastava, M.N.; Kumar, A.; Sathyanarayanan, P.V.; Nair, S.; Mohan, M. Chalcone synthase in rice (Oryza sativa L.): Detection of the CHS protein in seedlings and molecular mapping of the chs locus. Plant Mol. Biol. 1996, 32, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Druka, A.; Kudrna, D.; Rostoks, N.; Brueggeman, R.; von Wettstein, D.; Kleinhofs, A. Chalcone isomerase gene from rice (Oryza sativa) and barley (Hordeum vulgare): Physical, genetic and mutation mapping. Gene 2003, 302, 171–178. [Google Scholar] [CrossRef]
- Reddy, A.M.; Reddy, V.S.; Scheffler, B.E.; Wienand, U.; Reddy, A.R. Novel transgenic rice overexpressing anthocyanidin synthase accumulates a mixture of flavonoids leading to an increased antioxidant potential. Metab. Eng. 2007, 9, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Tanksley, S.D. Comparative linkage maps of the rice and maize genomes. Proc. Natl. Acad Sci. USA 1993, 90, 7980–7984. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.J.; Fan, Y.Y.; Du, J.H.; Zhuang, J.Y. Fine Mapping of C (Chromogen for Anthocyanin) Gene in Rice. Rice Sci. 2008, 15, 1–6. [Google Scholar] [CrossRef]
- Saitoh, K.; Onishi, K.; Mikami, I.; Thidar, K.; Sano, Y. Allelic diversification at the C (OsC1) locus of wild and cultivated rice: Nucleotide changes associated with phenotypes. Genetics 2004, 168, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; He, B.; Zhou, Y.; Sun, L. Genetic and molecular analysis of a purple sheath somaclonal mutant in japonica rice. Plant Cell Rep. 2011, 30, 901–911. [Google Scholar] [CrossRef]
- Chin, H.S.; Wu, Y.P.; Hour, A.L.; Hong, C.Y.; Lin, Y.R. Genetic and Evolutionary Analysis of Purple Leaf Sheath in Rice. Rice 2016, 9, 8. [Google Scholar] [CrossRef]
- Hu, J.; Anderson, B.; Wessler, S.R. Isolation and characterization of rice R genes: Evidence for distinct evolutionary paths in rice and maize. Genetics 1996, 142, 1021–1031. [Google Scholar]
- Hu, J.; Reddy, V.S.; Wessler, S.R. The rice R gene family: Two distinct subfamilies containing several miniature inverted-repeat transposable elements. Plant Mol. Biol. 2000, 42, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.T.; Thomson, M.J.; Pfeil, B.E.; McCouch, S. Caught red-handed: Rc encodes a basic helix-loop-helix protein conditioning red pericarp in rice. Plant Cell 2006, 18, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, T.; Maekawa, M.; Oki, T.; Suda, I.; Iida, S.; Shimada, H.; Takamure, I.; Kadowaki, K. The Rc and Rd genes are involved in proanthocyanidin synthesis in rice pericarp. Plant J. 2007, 49, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A. Genetic and molecular analysis of the anthocyanin pigmentation pathway in rice. In Rice Genetics III. Proceedings of the Third International Rice Genetics Sympos-ium, GS, K.; *E International Rice Research Institute: Manila, Philippines, 1995; pp. 341–352. [Google Scholar]
- Maeda, H.; Yamaguchi, T.; Omoteno, M.; Takarada, T.; Fujita, K.; Murata, K.; Iyama, Y.; Kojima, Y.; Morikawa, M.; Ozaki, H.; et al. Genetic dissection of black grain rice by the development of a near isogenic line. Breed. Sci. 2014, 64, 134–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, W.; Ohmori, T.; Kageyama, K.; Miyazaki, C.; Saito, A.; Murata, M.; Noda, K.; Maekawa, M. The Purple leaf (Pl) locus of rice: The Plw allele has a complex organization and includes two genes encoding basic helix-loop-helix proteins involved in anthocyanin biosynthesis. Plant Cell Physiol. 2001, 42, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, T.; Maeda, H.; Oguchi, T.; Yamaguchi, T.; Tanabe, N.; Ebana, K.; Yano, M.; Ebitani, T.; Izawa, T. The Birth of a Black Rice Gene and Its Local Spread by Introgression. Plant Cell 2015, 27, 2401–2414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Zhang, Z.; Chen, C.; Wu, W.; Ren, N.; Jiang, C.; Yu, J.; Zhao, Y.; Zheng, X.; Yang, Q.; et al. The C-S-A gene system regulates hull pigmentation and reveals evolution of anthocyanin biosynthesis pathway in rice. J. Exp. Bot. 2018, 69, 1485–1498. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tao, Y.; Ali, A.; Zhuang, Z.; Guo, D.; Guo, Q.; Riaz, A.; Zhang, H.; Xu, P.; Liao, Y.; et al. Transcriptome and Proteome Profiling of Different Colored Rice Reveals Physiological Dynamics Involved in the Flavonoid Pathway. Int. J. Mol. Sci. 2019, 20, 2463. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.; Du, D.; Xiao, L.; Yu, Q.; Shang, G.; Zhao, Z. Mapping and Identifying a Candidate Gene (Bnmfs) for Female-Male Sterility through Whole-Genome Resequencing and RNA-Seq in Rapeseed (Brassica napus L.). Front. Plant Sci. 2017, 8, 2086. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Li, J. Molecular genetic dissection of quantitative trait loci regulating rice grain size. Annu. Rev. Genet. 2014, 48, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Chen, K.; Mi, X.; Chen, T.; Ali, J.; Ye, G.; Xu, J.; Li, Z. Identification and Fine Mapping of a Stably Expressed QTL for Cold Tolerance at the Booting Stage Using an Interconnected Breeding Population in Rice. PLoS ONE 2015, 10, e0145704. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.L.; Grondin, A.; Courtois, B.; Gantet, P. Next-Generation Sequencing Accelerates Crop Gene Discovery. Trends Plant Sci. 2019, 24, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Lachagari, V.; Gupta, R.; Lekkala, S.P.; Mahadevan, L.; Kuriakose, B.; Chakravartty, N.; Mohan, K.A.; Santhosh, S.; Reddy, A.R.; Thomas, G. Whole Genome Sequencing and Comparative Genomic Analysis Reveal Allelic Variations Unique to a Purple Colored Rice Landrace (Oryza sativa ssp. indica cv. Purpleputtu). Front. Plant Sci. 2019, 10, 513. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Kersey, P.J.; Lawson, D.; Birney, E.; Derwent, P.S.; Haim el, M.; Herrero, J.; Keenan, S.; Kerhornou, A.; Koscielny, G.; Kahari, A.; et al. Genomes: Extending Ensembl across the taxonomic space. Nucleic Acids Res. 2010, 38, D563–D569. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
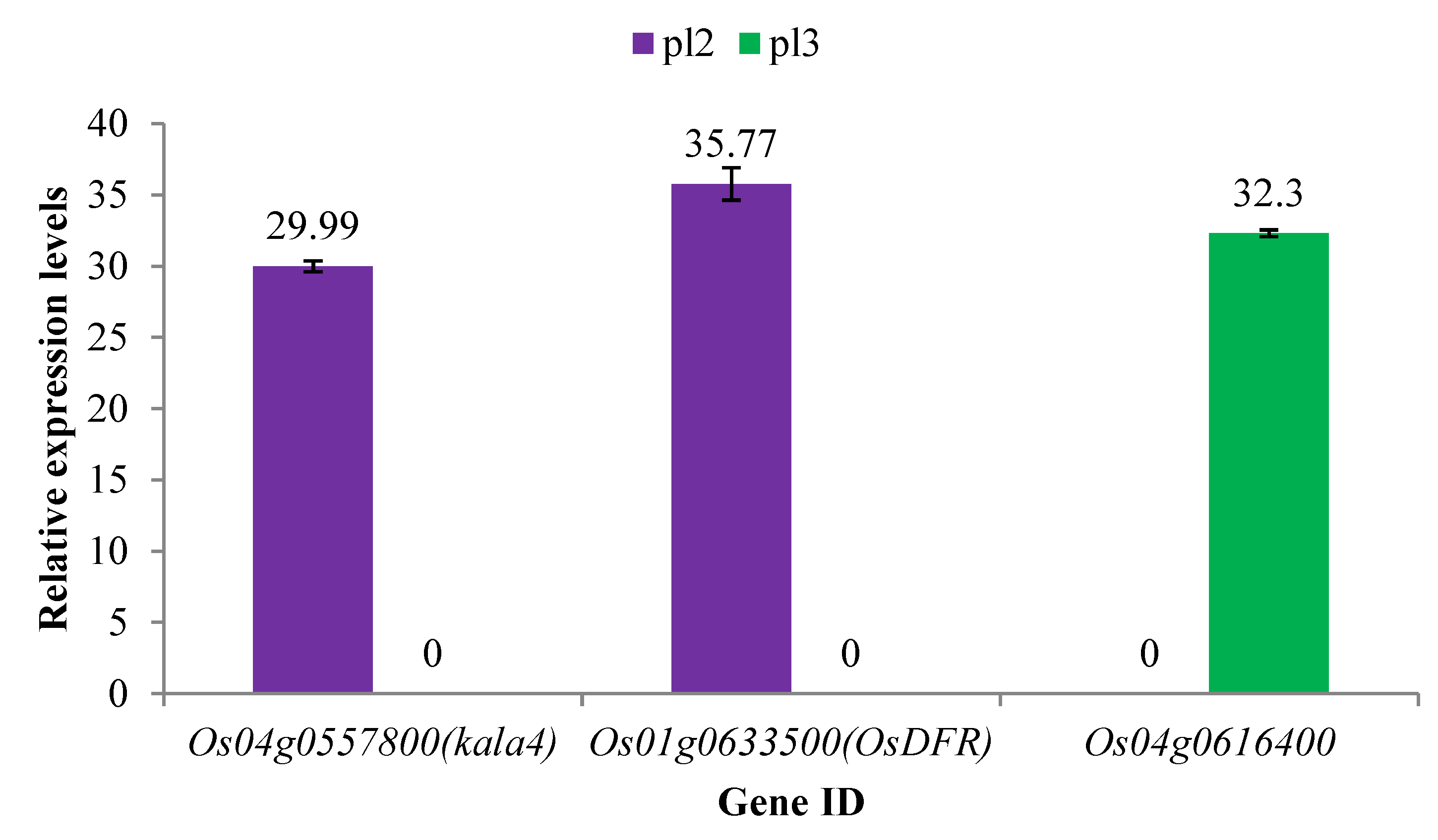{kind=link}
{kind=link}
| Gene_id | Readcount_pl2 | Readcount_pl3 | Log2foldchange | Pval | Padj | Annotation |
|---|---|---|---|---|---|---|
| OS02G0207100 | 3.083863443 | 25.98592481 | −3.0749 | 0.00016078 | 0.031904 | UDP-glycosyltransferase |
| OS04G0305700 | 227.9660245 | 60.11418403 | 1.923 | 2.87 × 10−6 | 0.00089272 | UDP-glycosyltransferase |
| OS04G0525100 | 517.0297915 | 55.58533108 | 3.2175 | 7.28 × 10−8 | 3.11 × 10−5 | UDP-glycosyltransferase |
| OS04G0525200 | 192.4804562 | 24.21323097 | 2.9908 | 2.22 × 10−10 | 1.78 × 10−7 | UDP-glycosyltransferase |
| OS06G0187500 | 3107.154031 | 953.1108178 | 1.7049 | 1.28 × 10−5 | 0.0035155 | UDP-glycosyltransferase |
| OS12G0561900 | 11.75742752 | 50.86821711 | −2.1132 | 0.00024505 | 0.045572 | UDP-glycosyltransferase |
| OS01G0735300 | 66.19051183 | 8.444736784 | 2.9705 | 0.00020562 | 0.03935 | anthocyanidin 3-O-glucosyltransferase |
| OS06G0192100 | 3002.334615 | 101.0286376 | 4.8932 | 2.83 × 10−13 | 4.05 × 10−10 | anthocyanidin 4-O-glucosyltransferase |
| OS07G0503500 | 285.5387079 | 19.21306586 | 3.8935 | 2.49 × 10−8 | 1.28 × 10−5 | anthocyanidin 5-O-glucosyltransferase |
| OS11G0461200 | 183.2680426 | 29.44993347 | 2.6376 | 2.74 × 10−5 | 0.0069429 | anthocyanidin 6-O-glucosyltransferase |
| OS04G0557800 | 26651.36063 | 50.70343466 | 9.0379 | 9.20 × 10−72 | 3.03 × 10−67 | anthocyanin regulatory Lc protein |
| OS11G0258700 | 37.48606985 | 0 | Inf | 6.84 × 10−11 | 5.92 × 10−8 | anthocyanin regulatory Lc protein |
| OS01G0372500 | 8756.71214 | 16.44143835 | 9.0569 | 1.39 × 10−20 | 4.15 × 10−17 | Leucoanthocyanidin dioxygenase |
| OS01G0633500 | 4738.917524 | 3.00821781 | 10.621 | 3.50 × 10−30 | 2.88 × 10−26 | Dihydroflavonol−4-reductase |
| OS04G0557500 | 4723.620711 | 2.49424188 | 10.887 | 1.07 × 10−40 | 1.75 × 10−36 | Anthocyanin regulatory R-S protein |
| OS04G0630300 | 10.54946251 | 386.0994026 | −5.1937 | 1.24 × 10−9 | 8.16 × 10−7 | Anthocyanidin reductase |
| OS10G0320100 | 938.0204694 | 98.06073341 | 3.2579 | 5.13 × 10−12 | 5.63 × 10−9 | Flavonoid 3′-hydroxylase |
| OS11G0530600 | 814.6837695 | 15.86922443 | 5.6819 | 9.91 × 10−29 | 6.52 × 10−25 | Chalcone synthase 1 |
| OS12G0270900 | 383.6491391 | 8.836561185 | 5.4402 | 8.77 × 10−28 | 4.81 × 10−24 | Flavonol 3-sulfotransferase |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Dai, G.; Zhou, W.; Liang, H.; Huang, J.; Qing, D.; Chen, W.; Wu, H.; Yang, X.; Li, D.; et al. Mapping and Identifying a Candidate Gene Plr4, a Recessive Gene Regulating Purple Leaf in Rice, by Using Bulked Segregant and Transcriptome Analysis with Next-Generation Sequencing. Int. J. Mol. Sci. 2019, 20, 4335. https://doi.org/10.3390/ijms20184335
Gao J, Dai G, Zhou W, Liang H, Huang J, Qing D, Chen W, Wu H, Yang X, Li D, et al. Mapping and Identifying a Candidate Gene Plr4, a Recessive Gene Regulating Purple Leaf in Rice, by Using Bulked Segregant and Transcriptome Analysis with Next-Generation Sequencing. International Journal of Molecular Sciences. 2019; 20(18):4335. https://doi.org/10.3390/ijms20184335
Chicago/Turabian StyleGao, Ju, Gaoxing Dai, Weiyong Zhou, Haifu Liang, Juan Huang, Dongjin Qing, Weiwei Chen, Hao Wu, Xinghai Yang, Danting Li, and et al. 2019. "Mapping and Identifying a Candidate Gene Plr4, a Recessive Gene Regulating Purple Leaf in Rice, by Using Bulked Segregant and Transcriptome Analysis with Next-Generation Sequencing" International Journal of Molecular Sciences 20, no. 18: 4335. https://doi.org/10.3390/ijms20184335