3D Printing PLA/Gingival Stem Cells/ EVs Upregulate miR-2861 and -210 during Osteoangiogenesis Commitment

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. 3D-PLA Evaluation
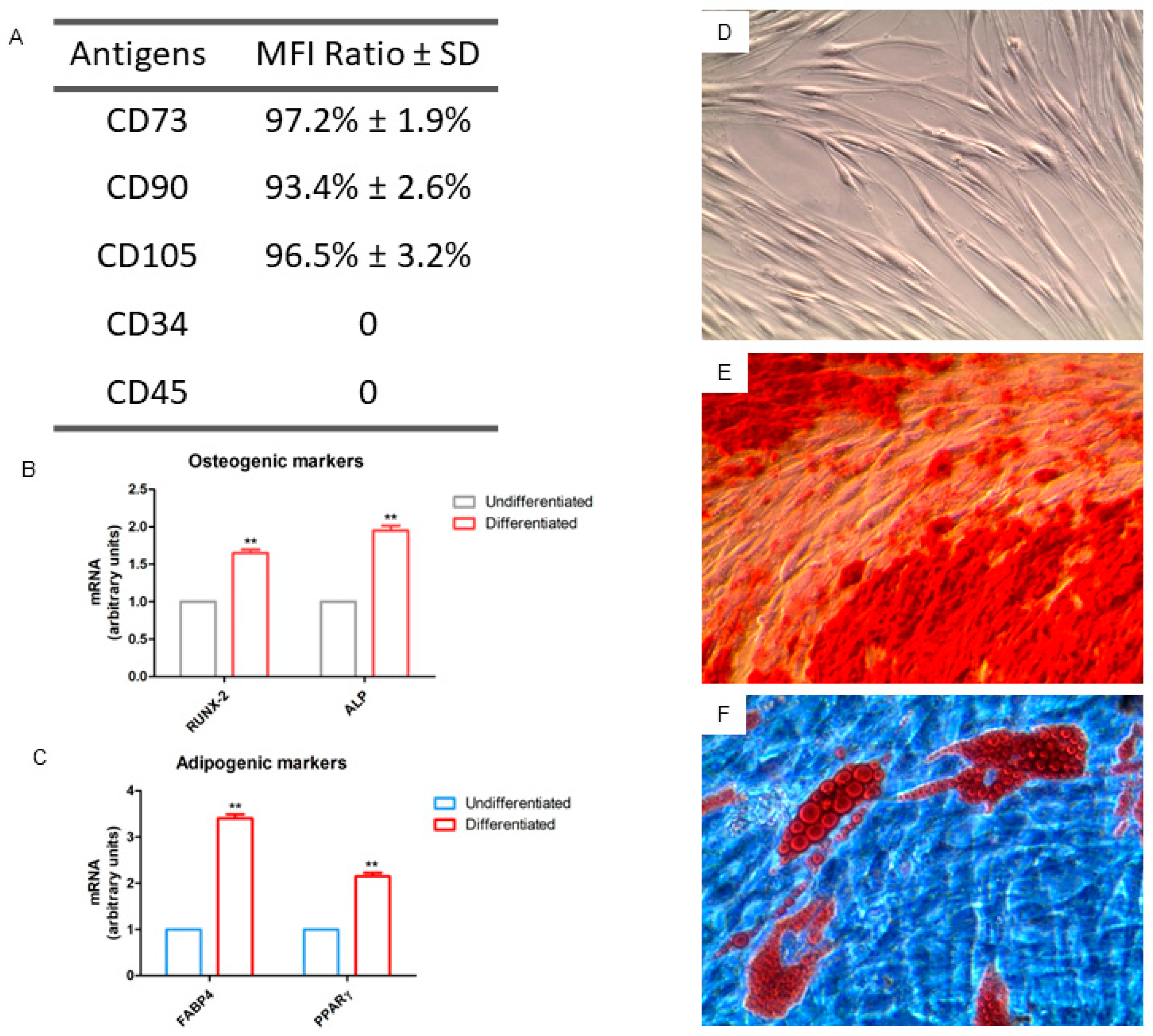
2.2. Cell Characterization
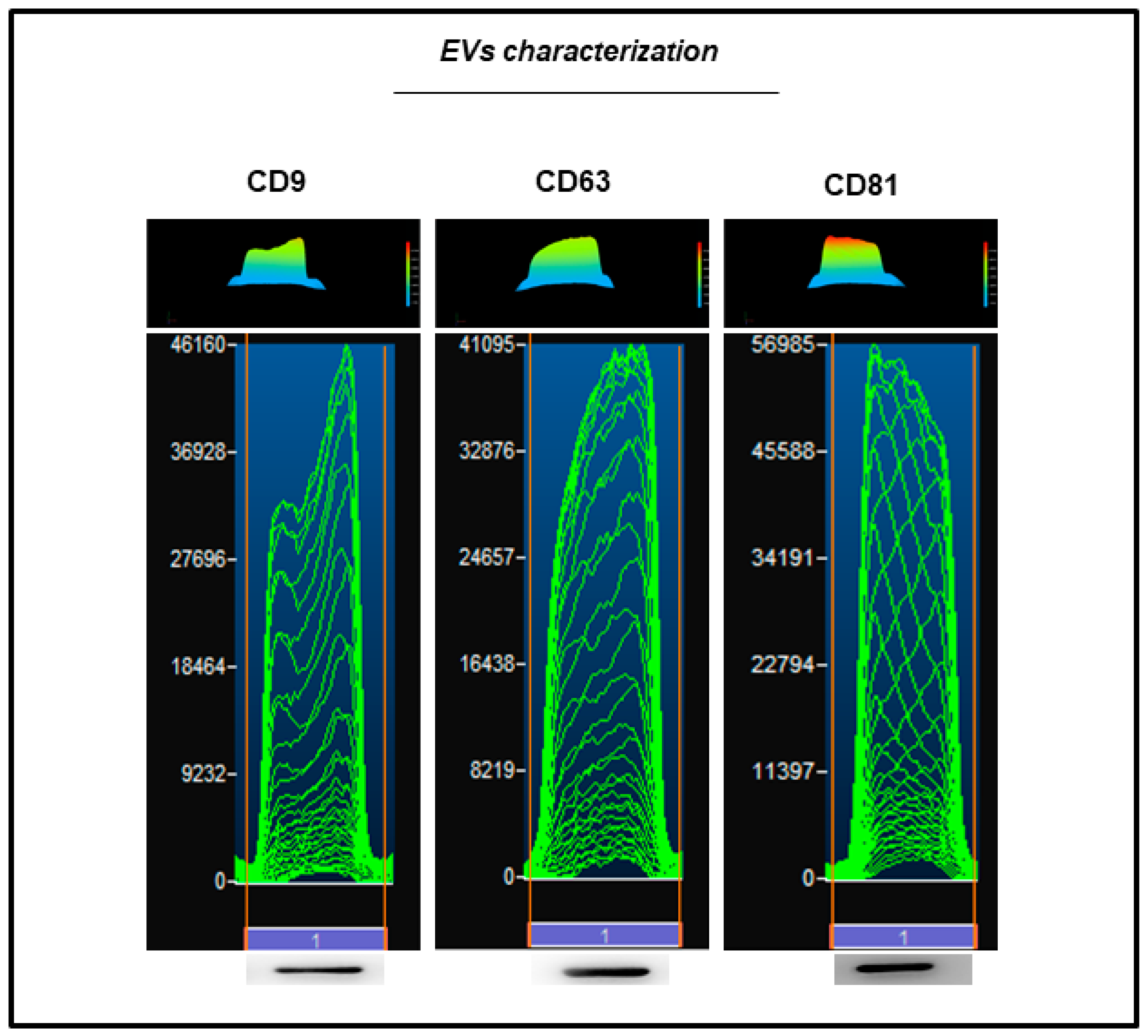
2.3. EVs Characterization
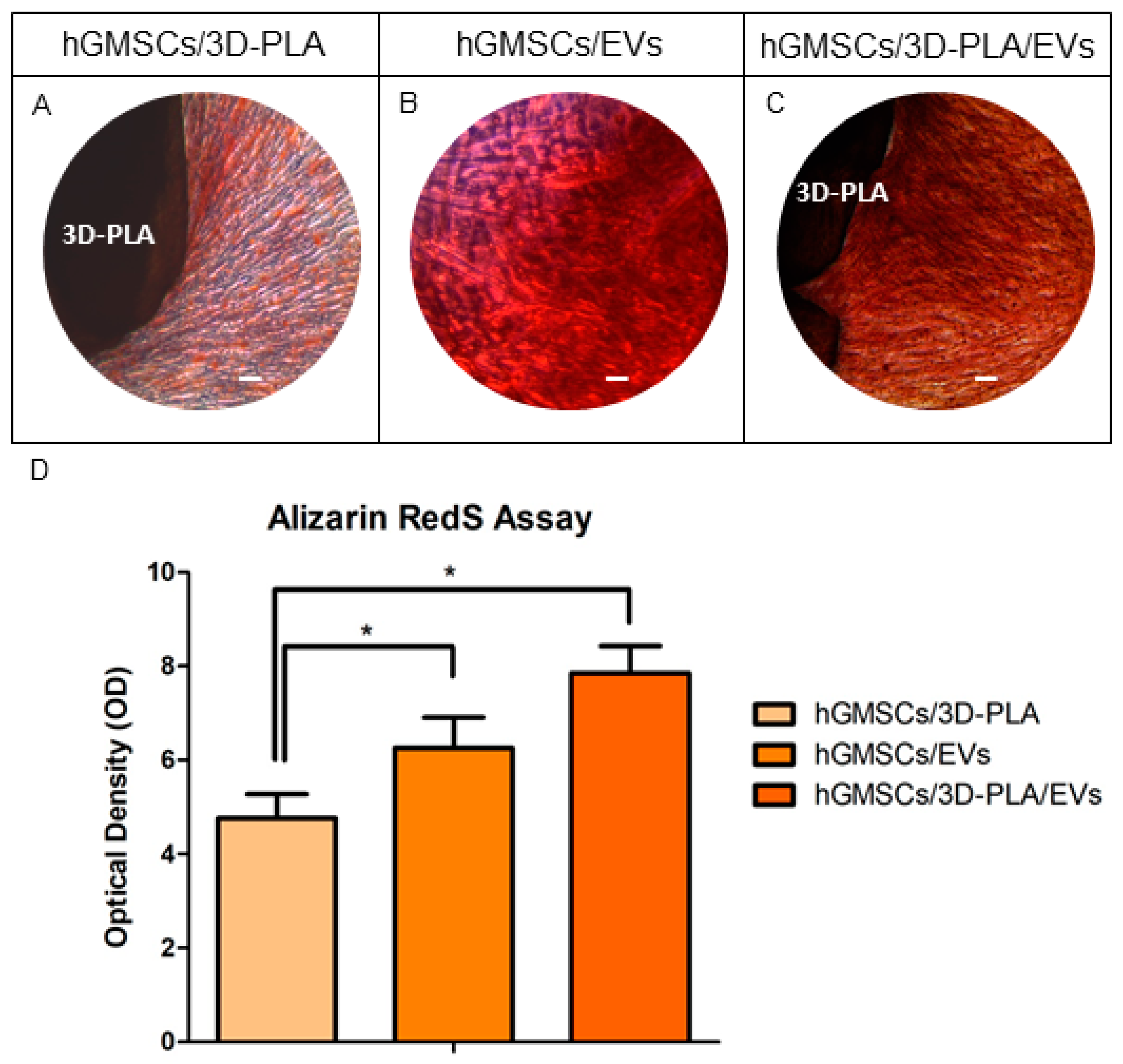
2.4. In Vitro Osteogenic Characterization
2.5. Gene Expression of Osteogenic Markers In Vitro
2.6. Western Blot Analysis of RUNX2 and VEGFA
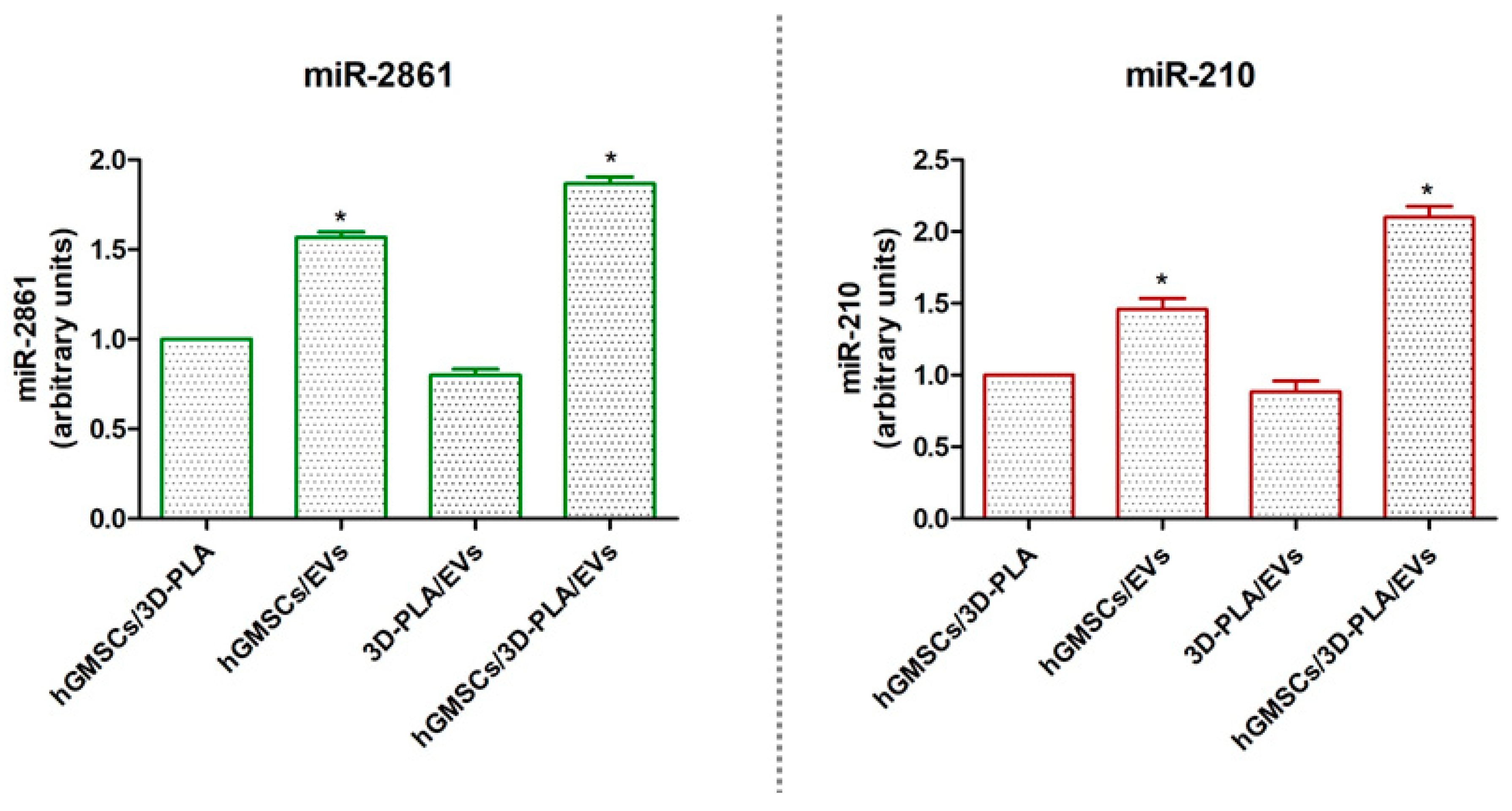
2.7. Micro-RNAs Expression
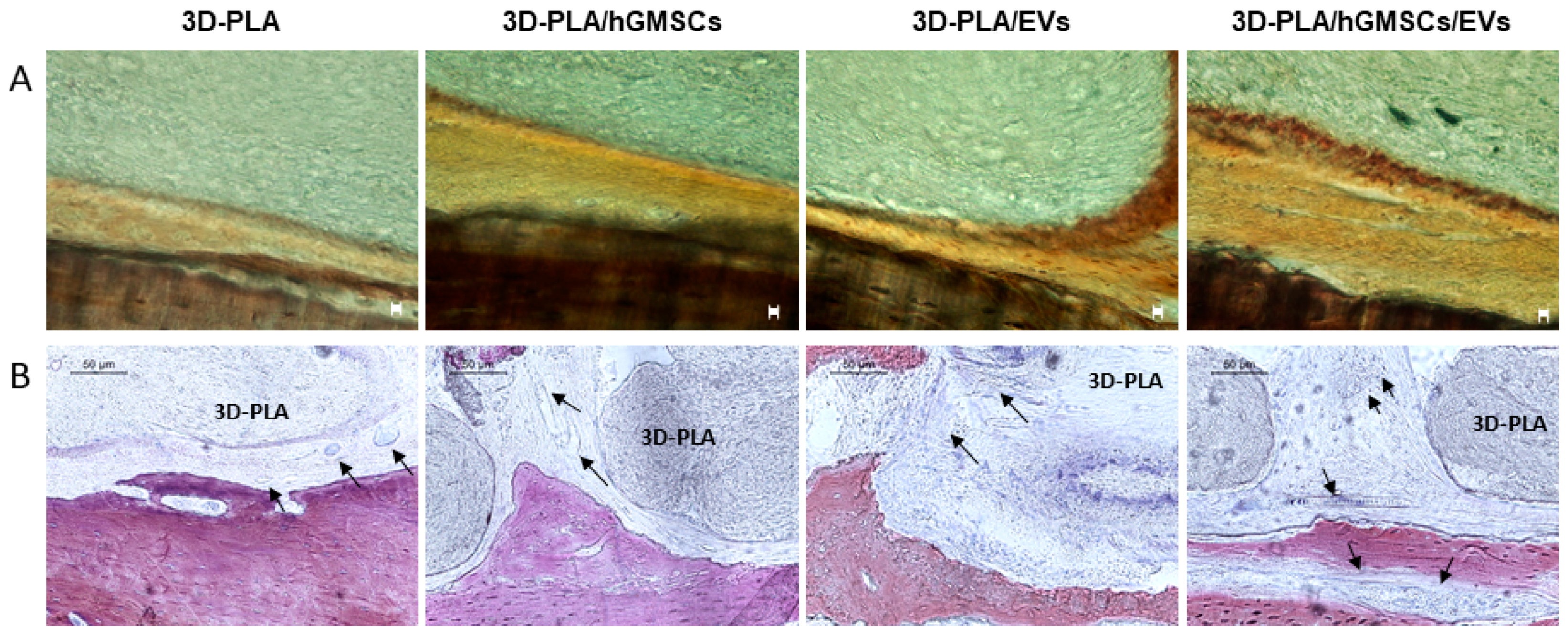
2.8. Histological Evaluation
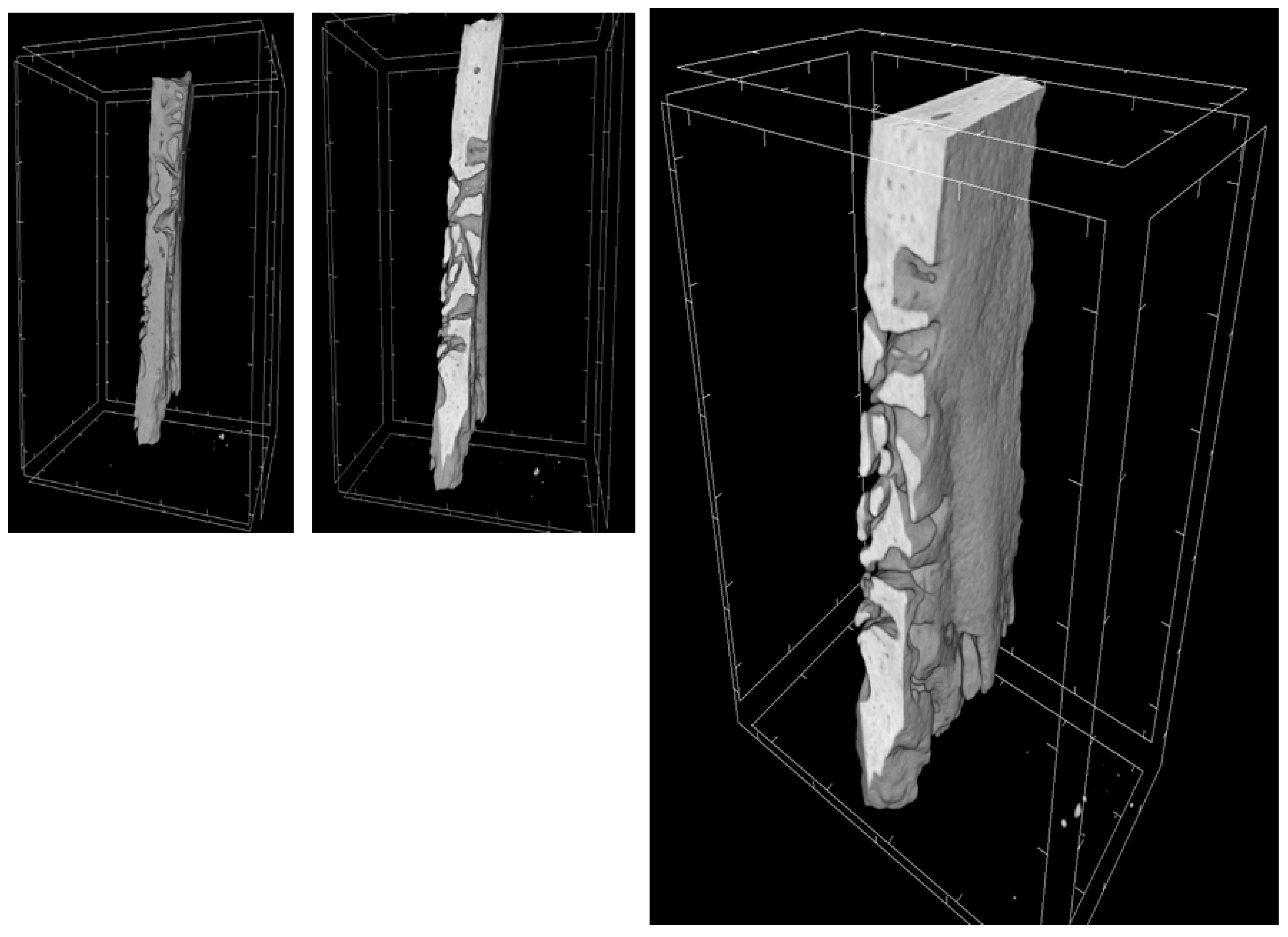
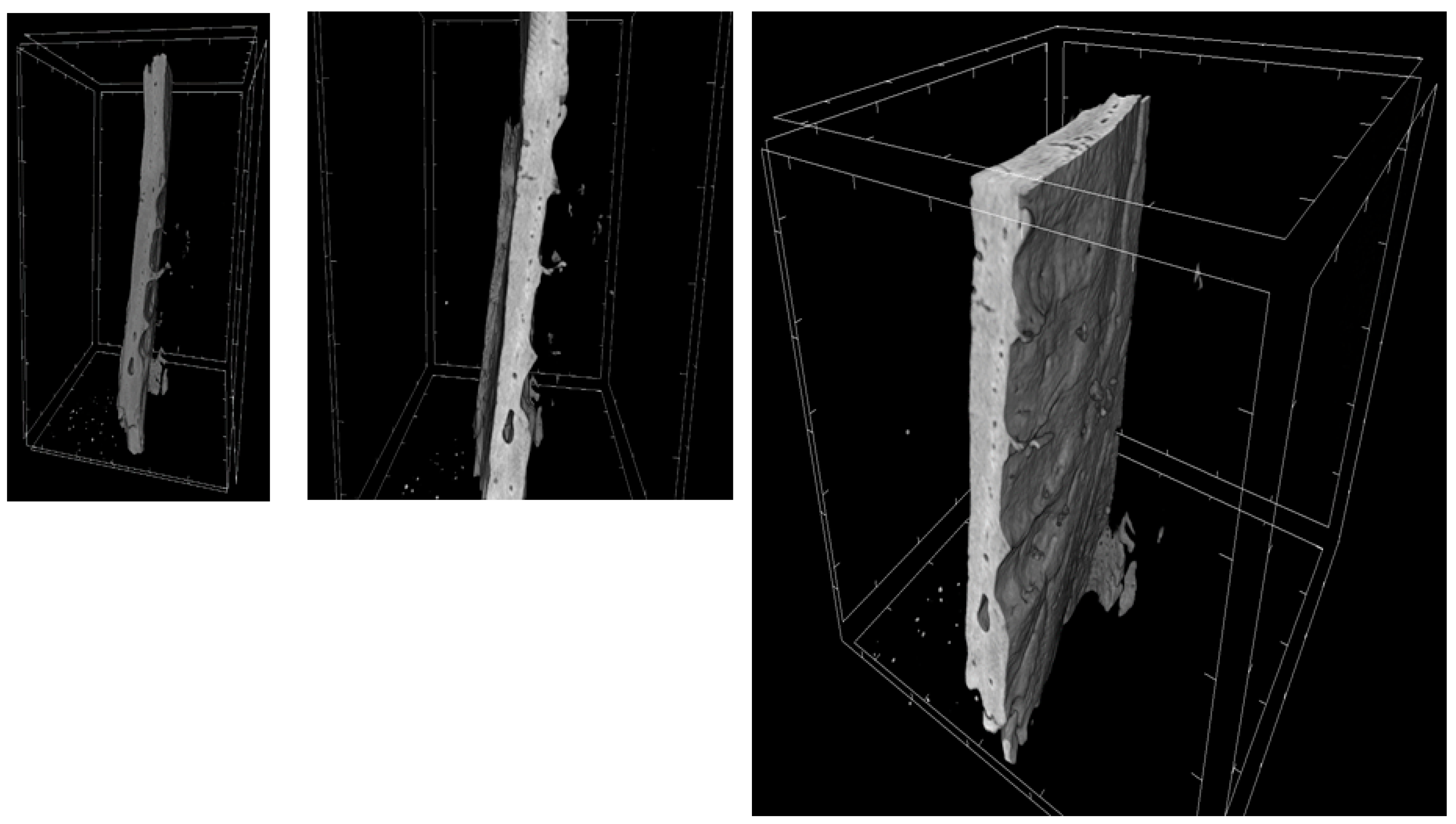
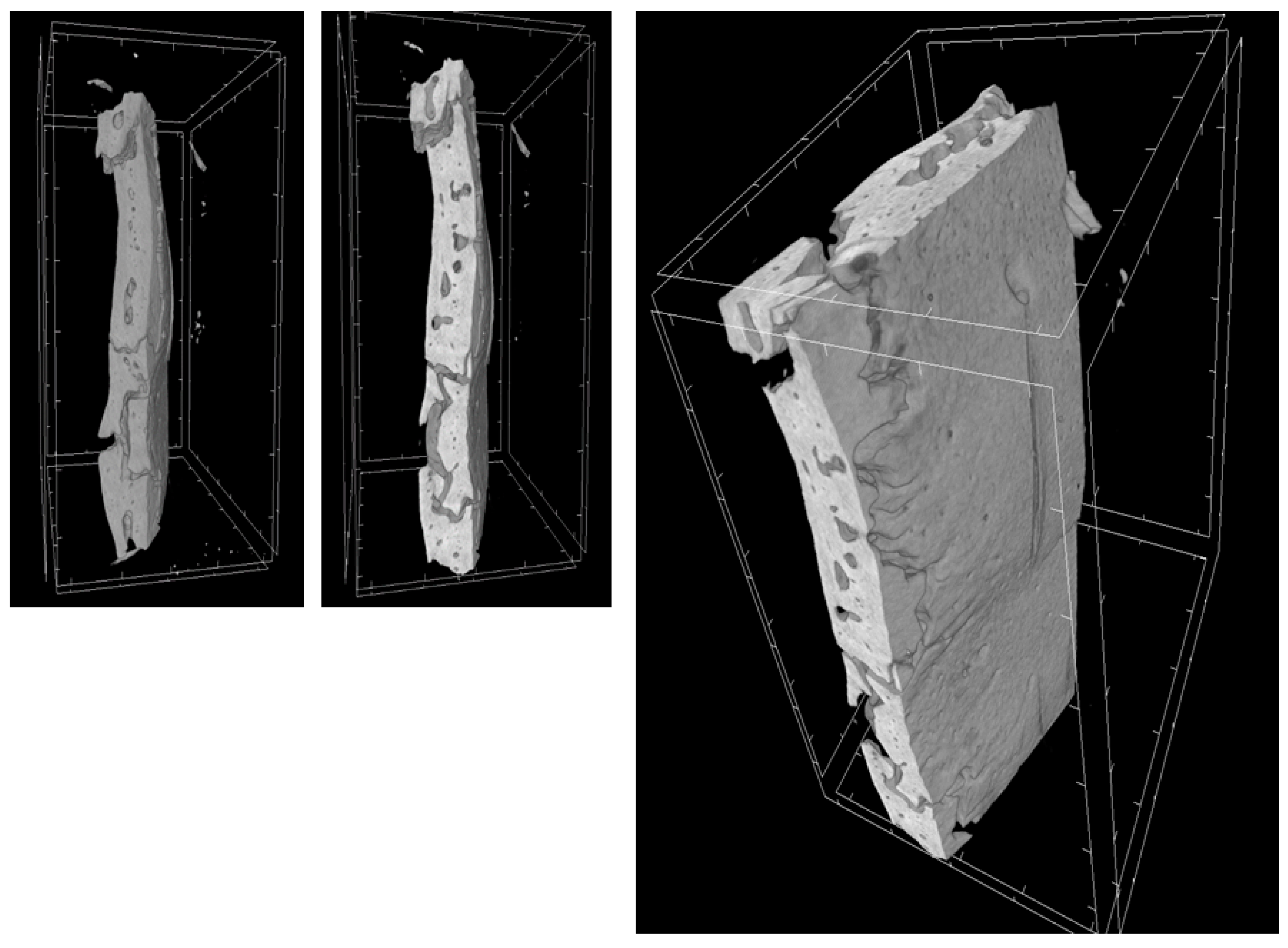
2.9. MicroCT
3. Discussion
4. Materials and Methods
4.1. Scaffold Material
4.2. In Vitro Study
4.2.1. Ethics Statement for In Vitro Experiments
4.2.2. Cell Culture Establishment and Characterization
4.2.3. Scanning Electron Microscopy Analysis
4.2.4. Extracellular Vesicles (EVs) Isolation
4.2.5. Scaffold Preparation
4.2.6. In Vitro Osteogenesis Performance
4.2.7. RNA Isolation and Real Time-PCR Analysis
4.2.8. Western Blot Analysis
4.2.9. MicroRNAs Quantization
4.3. In Vivo Study
4.3.1. Animals
4.3.2. Ethics Statement for Animal Use
4.3.3. Scaffold Grafting
4.3.4. Experimental Design
- 3D-PLA (N = 4): rats subjected to calvaria bone damage and grafted with 3D-PLA;
- 3D-PLA/hGMSCs (N =4): rats subjected to calvaria bone damage and grafted with 3D-COL enriched with hGMSCs;
- 3D-PLA/EVs (N = 4): rats subjected to calvaria bone damage and grafted with 3D-PLA enriched with EVs;
- 3D-PLA/hGMSCs/EVs (N = 4): rats subjected to calvaria bone damage and grafted with 3D-PLA enriched with hGMSCs and EVs;
4.3.5. Histological Evaluation
4.3.6. MicroCT Evaluation
4.4. Data and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MSCs | Mesenchymal stem cells |
| PLA | poly(lactide) |
| EVs | Extracellular vesicles |
| miRNA | microRNA |
| CM | conditioned medium |
| VEGF | Vascular endothelial growth factor |
| Hoxa2 | Homeobox A2 |
| HDACs | Histone deacetylases |
| RUNX2 | Runt-related transcription factor-2 |
| SEM | Scanning electron microscopy |
| ECM | Extracellular matrix |
| DLS | Dynamic light scattering |
| ALP | Alkaline phosphatase |
| OSX | Osterix |
| PPARγ | Peroxisome Proliferator-Activated Receptor γ |
| BMP-4 | Bone morphogenetic protein 4 |
| PBS | Phosphate-buffered saline |
| UPL | Universal ProbeLibrary |
References
- Wiese, A.; Pape, H.C. Bone Defects Caused by High-energy Injuries, Bone Loss, Infected Nonunions, and Nonunions. Orthop. Clin. N. Am. 2010, 41, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.P.; Leong, K.W. Scaffolding in tissue engineering: general approaches and tissue-specific considerations. Eur. Spine J. 2008, 17, S467–S479. [Google Scholar] [CrossRef] [PubMed]
- Nasonova, M.V.; Glushkova, T.V.; Borisov, V.V.; Velikanova, E.A.; Burago, A.Y.; Kudryavtseva, Y.A. Biocompatibility and Structural Features of Biodegradable Polymer Scaffolds. B. Exp. Biol. Med. 2015, 160, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Shum, A.W.T.; Mak, A.F.T. Morphological and biomechanical characterization of poly(glycolic acid) scaffolds after in vitro degradation. Polym. Degrad. Stabil. 2003, 81, 141–149. [Google Scholar] [CrossRef]
- Lopes, M.S.; Jardini, A.L.; Maciel, R. Poly (lactic acid) production for tissue engineering applications. Procedia Engineer. 2012, 42, 1402–1413. [Google Scholar] [CrossRef]
- Gugliandolo, A.; Diomede, F.; Cardelli, P.; Bramanti, A.; Scionti, D.; Bramanti, P.; Trubiani, O.; Mazzon, E. Transcriptomic analysis of gingival mesenchymal stem cells cultured on 3D bioprinted scaffold: A promising strategy for neuroregeneration. J. Biomed. Mater. Res. A 2018, 106, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Manescu, A.; Giuliani, A.; Mohammadi, S.; Tromba, G.; Mazzoni, S.; Diomede, F.; Zini, N.; Piattelli, A.; Trubiani, O. Osteogenic potential of dualblocks cultured with human periodontal ligament stem cells: in vitro and synchrotron microtomography study. J. Period. Res. 2016, 51, 112–124. [Google Scholar] [CrossRef]
- Matsushita, K. Mesenchymal Stem Cells and Metabolic Syndrome: Current understanding and Potential Clinical Implications. Stem Cells Int. 2016, 2016, 2892840. [Google Scholar] [CrossRef]
- Caplan, A.I.; Dennis, J.E. Mesenchymal stem cells as trophic mediators. J. cell. Biochem. 2006, 98, 1076–1084. [Google Scholar] [CrossRef]
- Konala, V.B.R.; Mamidi, M.K.; Bhonde, R.; Das, A.K.; Pochampally, R.; Pal, R. The current landscape of the mesenchymal stromal cell secretome: A new paradigm for cell-free regeneration. Cytotherapy 2016, 18, 13–24. [Google Scholar] [CrossRef]
- Baraniak, P.R.; McDevitt, T.C. Stem cell paracrine actions and tissue regeneration. Regen. Med. 2010, 5, 121–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammana, S.; Gugliandolo, A.; Cavalli, E.; Diomede, F.; Iori, R.; Zappacosta, R.; Bramanti, P.; Conti, P.; Fontana, A.; Pizzicannella, J.; et al. Human Gingival Mesenchymal Stem Cells (GMSCs) pre-treated with vesicular Moringin nanostructures as a new therapeutic approach in a mouse model of Spinal Cord Injury. J. Tissue Eng. Regen. Med. 2019. [Google Scholar] [CrossRef] [PubMed]
- Roura, S.; Vives, J. Extracellular vesicles: squeezing every drop of regenerative potential of umbilical cord blood. Metab. Clin. Experim. 2019. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Zhang, X.M.; Li, X.R. Exosomes Derived from Mesenchymal Stem Cells. Int. J. Mol. Sci 2014, 15, 4142–4157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahabipour, F.; Banach, M.; Sahebkar, A. Exosomes as nanocarriers for siRNA delivery: Paradigms and challenges. Arch. Med. Sci 2016, 12, 1324–1326. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.H.; Wang, L.; Gao, Z.L.; Chen, G.Y.; Zhang, C.Q. Bone marrow stromal/stem cell-derived extracellular vesicles regulate osteoblast activity and differentiation in vitro and promote bone regeneration in vivo. Sci Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Miyaki, S.; Ishitobi, H.; Ogura, T.; Kato, Y.; Kamei, N.; Miyado, K.; Higashi, Y.; Ochi, M. Mesenchymal Stem Cell-Derived Exosomes Promote Fracture Healing in a Mouse Model. Stem Cell Transl. Med. 2016, 5, 1620–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, H.; Wang, Z.X.; Zhang, L.M.; Lei, Q.; Zhao, A.Q.; Wang, H.X.; Li, Q.B.; Cao, Y.L.; Zhang, W.J.; Chen, Z.C. Extracellular Vesicle-functionalized Decalcified Bone Matrix Scaffolds with Enhanced Pro-angiogenic and Pro-bone Regeneration Activities. Sci Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Casap, N.; Venezia, N.B.; Wilensky, A.; Samuni, Y. VEGF facilitates periosteal distraction-induced osteogenesis in rabbits: A micro-computerized tomography study. Tissue Eng. Part A 2008, 14, 247–253. [Google Scholar] [CrossRef]
- Ferguson, C.; Alpern, E.; Miclau, T.; Helms, J.A. Does adult fracture repair recapitulate embryonic skeletal formation? Mech. Develop. 1999, 87, 57–66. [Google Scholar] [CrossRef]
- Hu, R.; Liu, W.; Li, H.; Yang, L.; Chen, C.; Xia, Z.Y.; Guo, L.J.; Xie, H.; Zhou, H.D.; Wu, X.P.; et al. A Runx2/miR-3960/miR-2861 regulatory feedback loop during mouse osteoblast differentiation. J. Biolog. Chem. 2011, 286, 12328–12339. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.O.; Vorwald, C.E.; Dreher, M.L.; Mott, E.J.; Cheng, M.H.; Cinar, A.; Mehdizadeh, H.; Somo, S.; Dean, D.; Brey, E.M.; et al. Evaluating 3D-Printed Biomaterials as Scaffolds for Vascularized Bone Tissue Engineering. Adv. Mater. 2015, 27, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Gugliandolo, A.; Scionti, D.; Merciaro, I.; Cavalcanti, M.F.; Mazzon, E.; Trubiani, O. Biotherapeutic Effect of Gingival Stem Cells Conditioned Medium in Bone Tissue Restoration. Int. J. Mol. Sci 2018, 19, 329. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Gugliandolo, A.; Cardelli, P.; Merciaro, I.; Ettorre, V.; Traini, T.; Bedini, R.; Scionti, D.; Bramanti, A.; Nanci, A.; et al. Three-dimensional printed PLA scaffold and human gingival stem cell-derived extracellular vesicles: a new tool for bone defect repair. Stem Cell Res. Ther. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Rajan, T.S.; Giacoppo, S.; Diomede, F.; Ballerini, P.; Paolantonio, M.; Marchisio, M.; Piattelli, A.; Bramanti, P.; Mazzon, E.; Trubiani, O. The secretome of periodontal ligament stem cells from MS patients protects against EAE. Sci Rep. 2016, 6, 38743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyorgy, B.; Szabo, T.G.; Pasztoi, M.; Pal, Z.; Misjak, P.; Aradi, B.; Laszlo, V.; Pallinger, E.; Pap, E.; Kittel, A.; et al. Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles. Cell. Mol. Life Sci 2011, 68, 2667–2688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarkin, C.E.; Gerstenfeld, L.C. VEGF and bone cell signalling: an essential vessel for communication? Cell Bioche. Func. 2013, 31, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Olsen, B.R. Osteoblast-derived VEGF regulates osteoblast differentiation and bone formation during bone repair. J. Clinic. Invest. 2016, 126, 509–526. [Google Scholar] [CrossRef] [Green Version]
- Wiszniak, S.; Mackenzie, F.E.; Anderson, P.; Kabbara, S.; Ruhrberg, C.; Schwarz, Q. Neural crest cell-derived VEGF promotes embryonic jaw extension. P. Natl. Acad. Sci USA 2015, 112, 6086–6091. [Google Scholar] [CrossRef] [Green Version]
- Weilner, S.; Schraml, E.; Wieser, M.; Messner, P.; Schneider, K.; Wassermann, K.; Micutkova, L.; Fortschegger, K.; Maier, A.B.; Westendorp, R.; et al. Secreted microvesicular miR-31 inhibits osteogenic differentiation of mesenchymal stem cells. Aging Cell. 2016, 15, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Marin-Muller, C.; Bharadwaj, U.; Chow, K.H.; Yao, Q.Z.; Chen, C.Y. MicroRNAs: Control and Loss of Control in Human Physiology and Disease. World J. Surg. 2009, 33, 667–684. [Google Scholar] [CrossRef] [PubMed]
- Suarez, Y.; Sessa, W.C. MicroRNAs As Novel Regulators of Angiogenesis. Circ. Res. 2009, 104, 442–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Chen, C.; Yang, D.; Liao, Q.; Luo, H.; Wang, X.; Zhou, F.; Yang, X.; Yang, J.; Zeng, C.; et al. Mesenchymal stem cells-derived extracellular vesicles, via miR-210, improve infarcted cardiac function by promotion of angiogenesis. Biochim. Bioph. Acta Mol. Basis Dis. 2017, 1863, 2085–2092. [Google Scholar] [CrossRef] [PubMed]
- Pizzicannella, J.; Cavalcanti, M.; Trubiani, O.; Diomede, F. MicroRNA 210 Mediates VEGF Upregulation in Human Periodontal Ligament Stem Cells Cultured on 3DHydroxyapatite Ceramic Scaffold. Int. J. Mol. Sci 2018, 19, 3916. [Google Scholar] [CrossRef] [PubMed]
- Luft, F.C. Merely miR210 in mesenchymal stem cells-one size fits all. J. Mol. Med. 2012, 90, 983–985. [Google Scholar] [CrossRef] [PubMed]
- Ivan, M.; Harris, A.L.; Martelli, F.; Kulshreshtha, R. Hypoxia response and microRNAs: no longer two separate worlds. J. Cell. Mol. Med. 2008, 12, 1426–1431. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.D.; Cai, F.; Liu, L.; Zhang, Y.; Yang, A.L. microRNA-210 is involved in the regulation of postmenopausal osteoporosis through promotion of VEGF expression and osteoblast differentiation. Biol. Chem. 2015, 396, 339–347. [Google Scholar] [CrossRef]
- Diomede, F.; Merciaro, I.; Martinotti, S.; Cavalcanti, M.F.X.B.; Caputi, S.; Mazzon, E.; Trubiani, O. miR-2861 IS INVOLVED IN OSTEOGENIC COMMITMENT OF HUMAN PERIODONTAL LIGAMENT STEM CELLS GROWN ONTO 3D SCAFFOLD. J. Biol. Reg. Homeos. Ag. 2016, 30, 1009–1018. [Google Scholar]
- Xia, Z.Y.; Hu, Y.; Xie, P.L.; Tang, S.Y.; Luo, X.H.; Liao, E.Y.; Chen, F.; Xie, H. Runx2/miR-3960/miR-2861 Positive Feedback Loop Is Responsible for Osteogenic Transdifferentiation of Vascular Smooth Muscle Cells. Biomed. Res. Int. 2015, 2015, 624037. [Google Scholar] [CrossRef]
- Filipowska, J.; Tomaszewski, K.A.; Niedzwiedzki, L.; Walocha, J.A.; Niedzwiedzki, T. The role of vasculature in bone development, regeneration and proper systemic functioning. Angiogenesis 2017, 20, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Pizzicannella, J.; Diomede, F.; Merciaro, I.; Caputi, S.; Tartaro, A.; Guarnieri, S.; Trubiani, O. Endothelial committed oral stem cells as modelling in the relationship between periodontal and cardiovascular disease. J. Cell. Physiol. 2018, 233, 6734–6747. [Google Scholar] [CrossRef] [PubMed]
- Pizzicannella, J.; Gugliandolo, A.; Orsini, T.; Fontana, A.; Ventrella, A.; Mazzon, E.; Bramanti, P.; Diomede, F.; Trubiani, O. Engineered Extracellular Vesicles From Human Periodontal-Ligament Stem Cells Increase VEGF/VEGFR2 Expression During Bone Regeneration. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Zini, N.; Pizzicannella, J.; Merciaro, I.; Pizzicannella, G.; D’Orazio, M.; Piattelli, A.; Trubiani, O. 5-Aza Exposure Improves Reprogramming Process Through Embryoid Body Formation in Human Gingival Stem Cells. Front. Genet. 2018, 9, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzicannella, J.; Rabozzi, R.; Trubiani, O.; Di Giammarco, G. HTK solution helps to preserve endothelial integrity of saphenous vein: an immunohistochemical and ultrastructural analysis. J. Biol. Regul. Homeost. Agents 2011, 25, 93–99. [Google Scholar] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fiber diameter | 2.245 × 102 µm |
|---|---|
| Pore size | 5.042 × 105 µm2 |
| Interconnectivity | 1877 |
| Surface area | 6 × 103 µm3 |
| MicroCT Parameter | 3D-PLA | 3D-PLA/hGMSCs | 3D-PLA/EVs | 3D-PLA/hGMSCs/EVs |
|---|---|---|---|---|
| BV (µm3) | 3.2 × 109 ± 1.6 × 108 | 4.3 × 109 ± 1.7 × 108 | 3.4 × 109 ± 2.9 × 108 | 6.8 × 109 ± 2.6 × 108 |
| BV/TV (%) | 3.9 ± 0.1 | 5.2 ± 0.2 | 4.1 ± 0.3 | 8.2 ± 0.3 |
| BS (µm2) | 3.1 × 107 ± 2.2 × 106 | 3.2 × 107 ± 2.4 × 106 | 3.2 × 107 ± 2.3 × 106 | 6.6 × 107 ± 1.7 × 106 |
| BS/BV (1/µm) | 9.8 × 10-3 ± 3.8 × 10-4 | 7.4 × 10-3 ± 3.8 × 10-4 | 9.5 × 10-3 ± 3.9 × 10-4 | 9.8 × 10-3 ± 1.6 × 10-4 |
| BS/TV (1/µm) | 3.8 × 10-4 ± 1.3 × 10-5 | 3.9 × 10-4 ± 2.9 × 10-5 | 3.9 × 10-4 ± 2.6 × 10-5 | 8.1 × 10-4 ± 1.3 × 10-5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pizzicannella, J.; Diomede, F.; Gugliandolo, A.; Chiricosta, L.; Bramanti, P.; Merciaro, I.; Orsini, T.; Mazzon, E.; Trubiani, O. 3D Printing PLA/Gingival Stem Cells/ EVs Upregulate miR-2861 and -210 during Osteoangiogenesis Commitment. Int. J. Mol. Sci. 2019, 20, 3256. https://doi.org/10.3390/ijms20133256
Pizzicannella J, Diomede F, Gugliandolo A, Chiricosta L, Bramanti P, Merciaro I, Orsini T, Mazzon E, Trubiani O. 3D Printing PLA/Gingival Stem Cells/ EVs Upregulate miR-2861 and -210 during Osteoangiogenesis Commitment. International Journal of Molecular Sciences. 2019; 20(13):3256. https://doi.org/10.3390/ijms20133256
Chicago/Turabian StylePizzicannella, Jacopo, Francesca Diomede, Agnese Gugliandolo, Luigi Chiricosta, Placido Bramanti, Ilaria Merciaro, Tiziana Orsini, Emanuela Mazzon, and Oriana Trubiani. 2019. "3D Printing PLA/Gingival Stem Cells/ EVs Upregulate miR-2861 and -210 during Osteoangiogenesis Commitment" International Journal of Molecular Sciences 20, no. 13: 3256. https://doi.org/10.3390/ijms20133256