Cadmium Complexed with β2-Microglubulin, Albumin and Lipocalin-2 rather than Metallothionein Cause Megalin:Cubilin Dependent Toxicity of the Renal Proximal Tubule

 , , and
, , and
Abstract
:1. Introduction
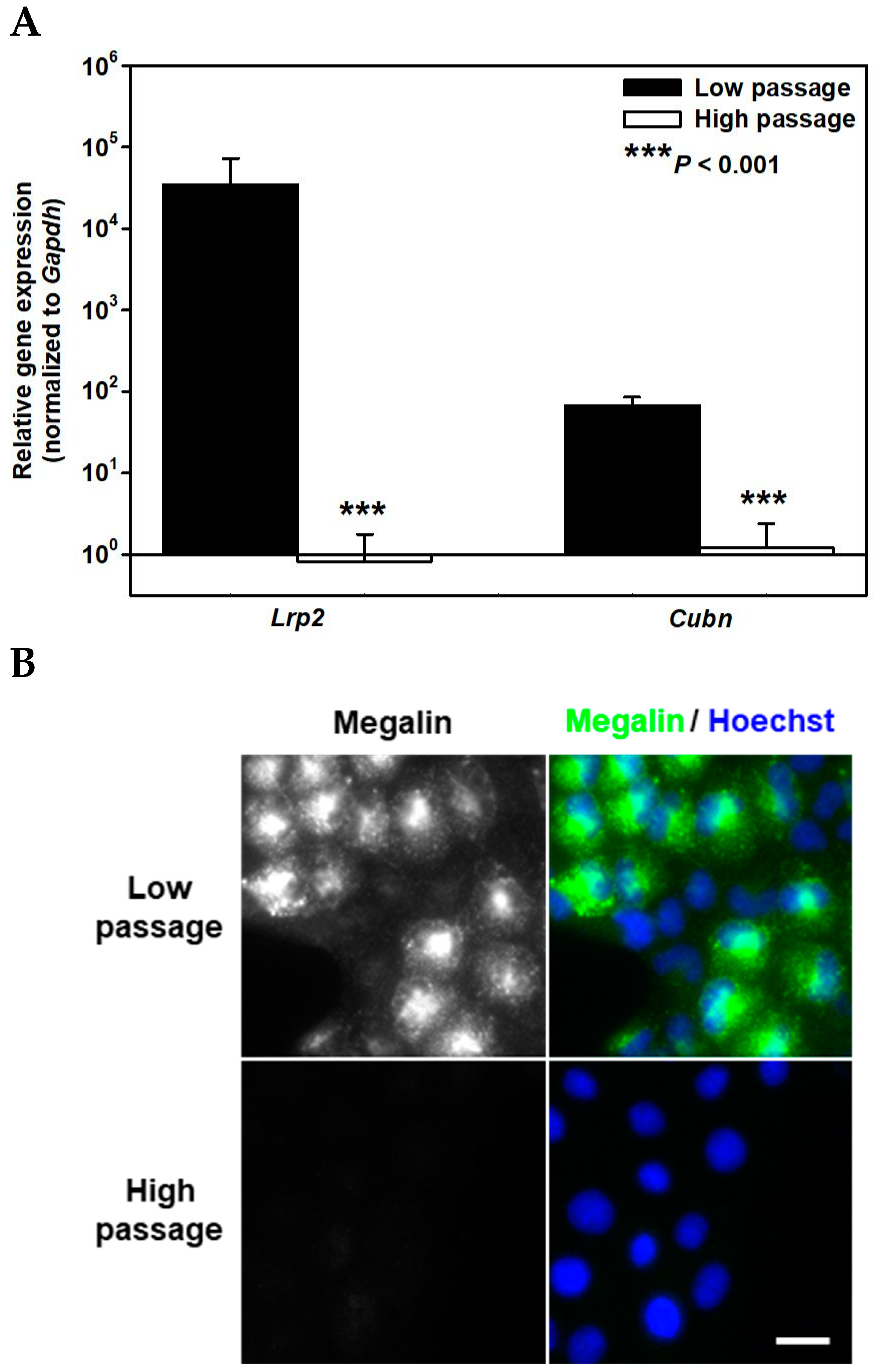
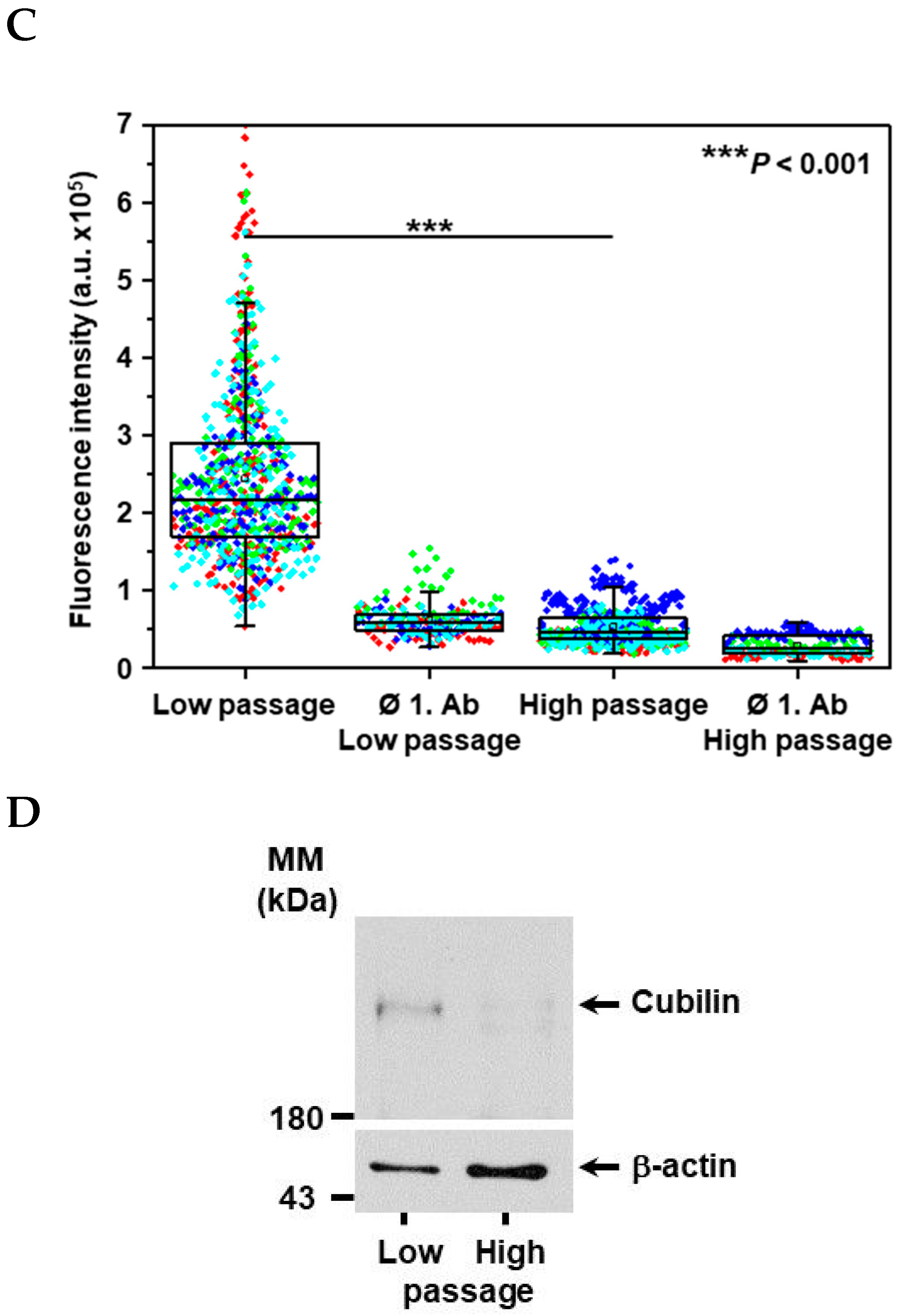
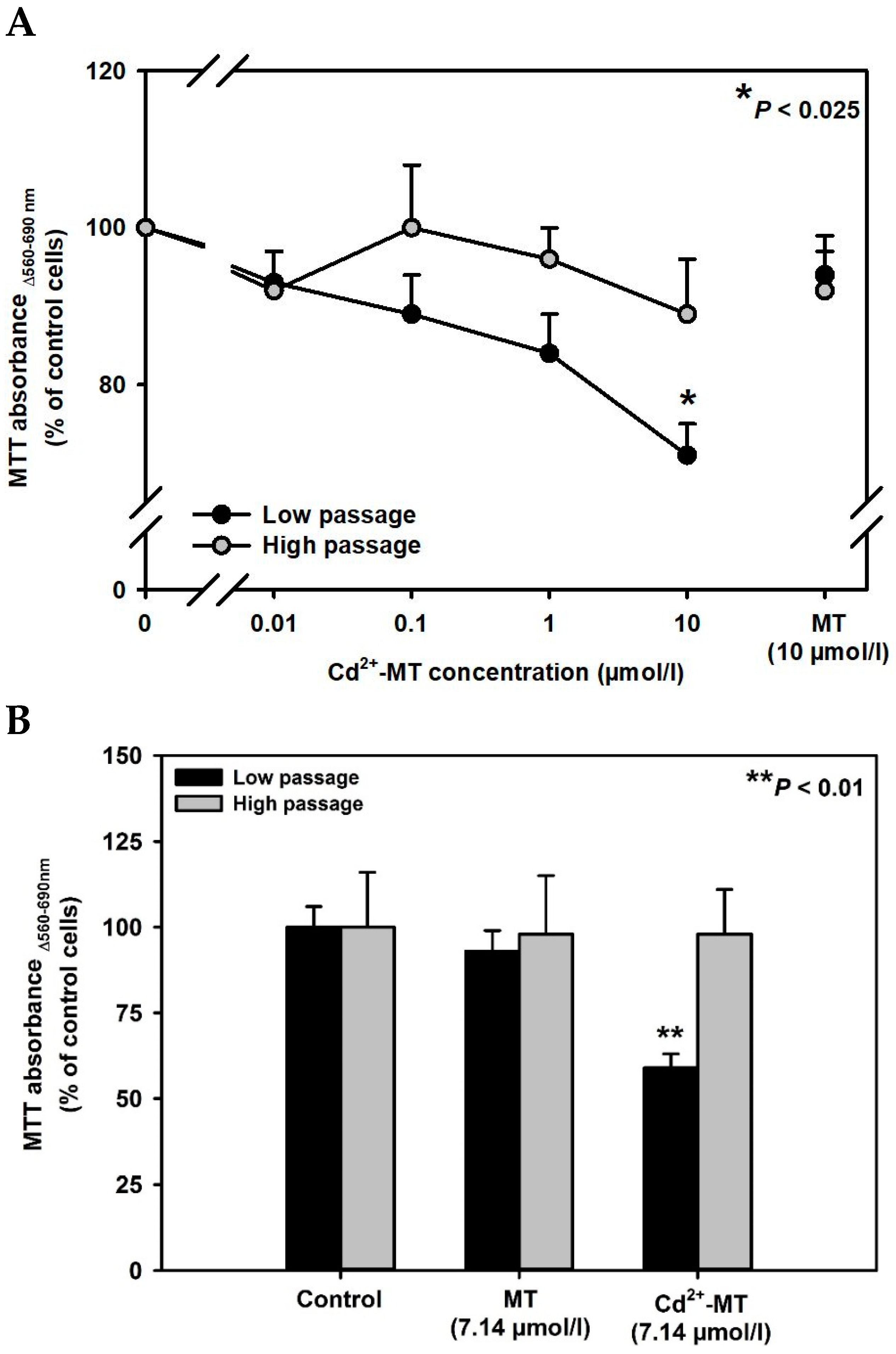
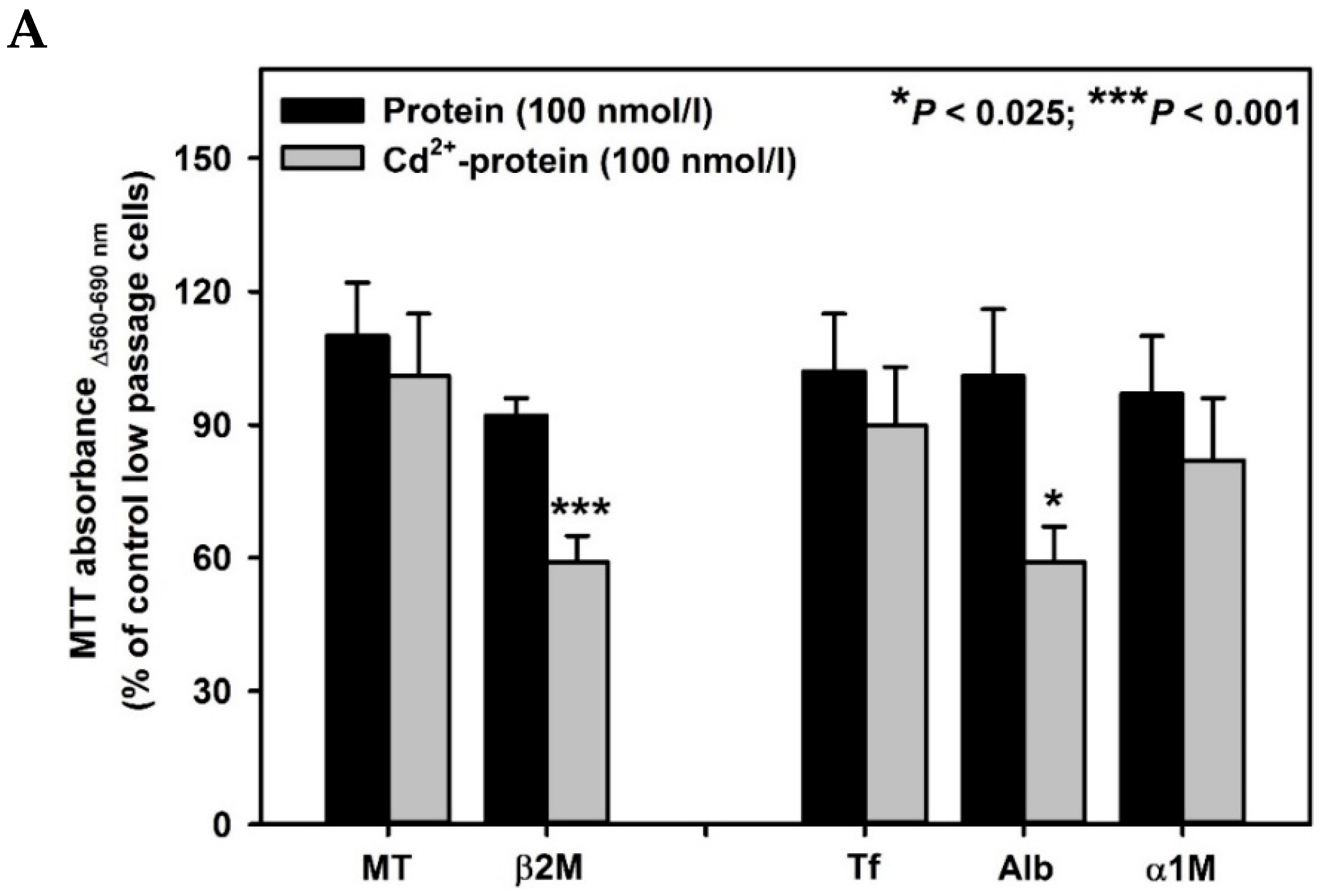
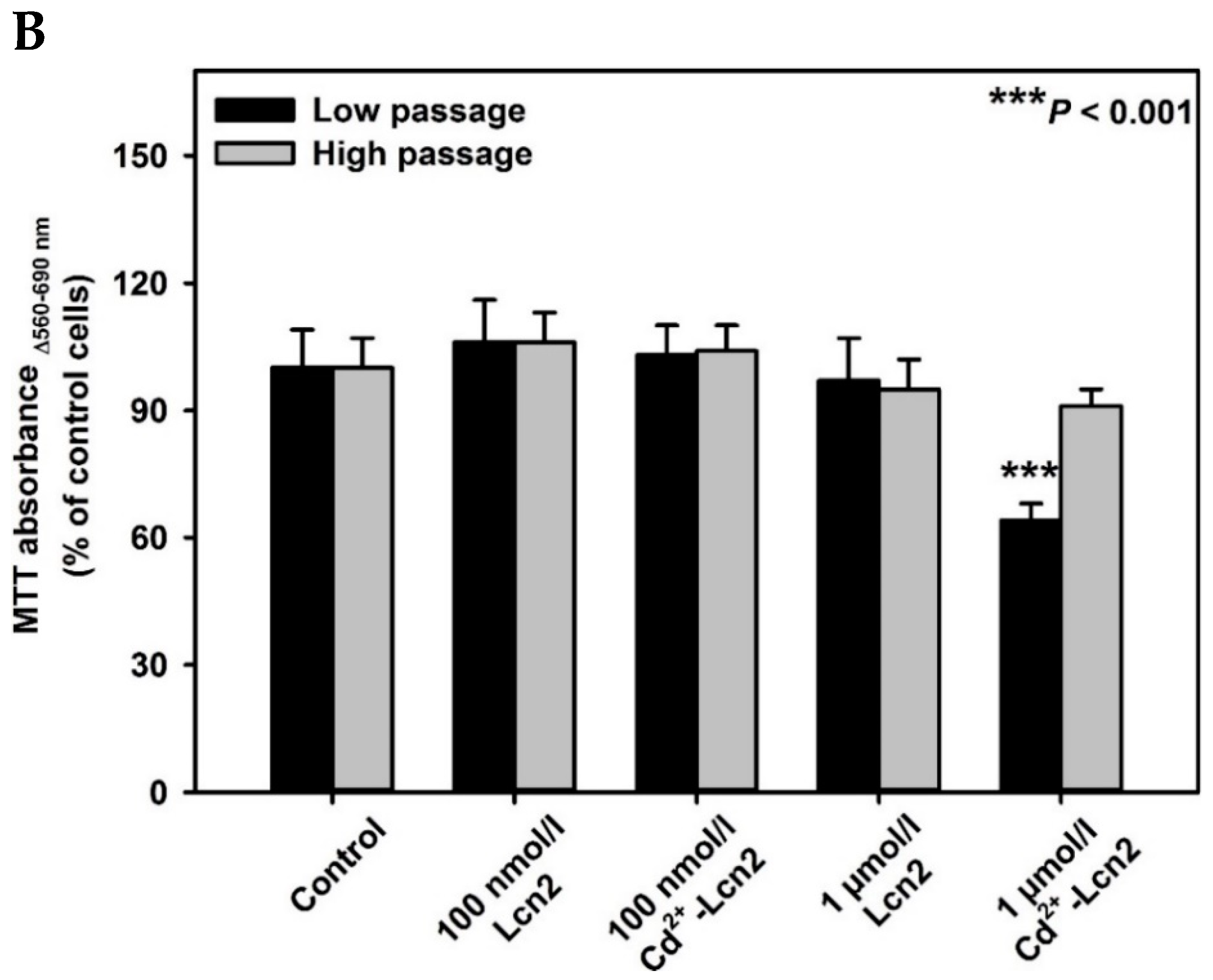
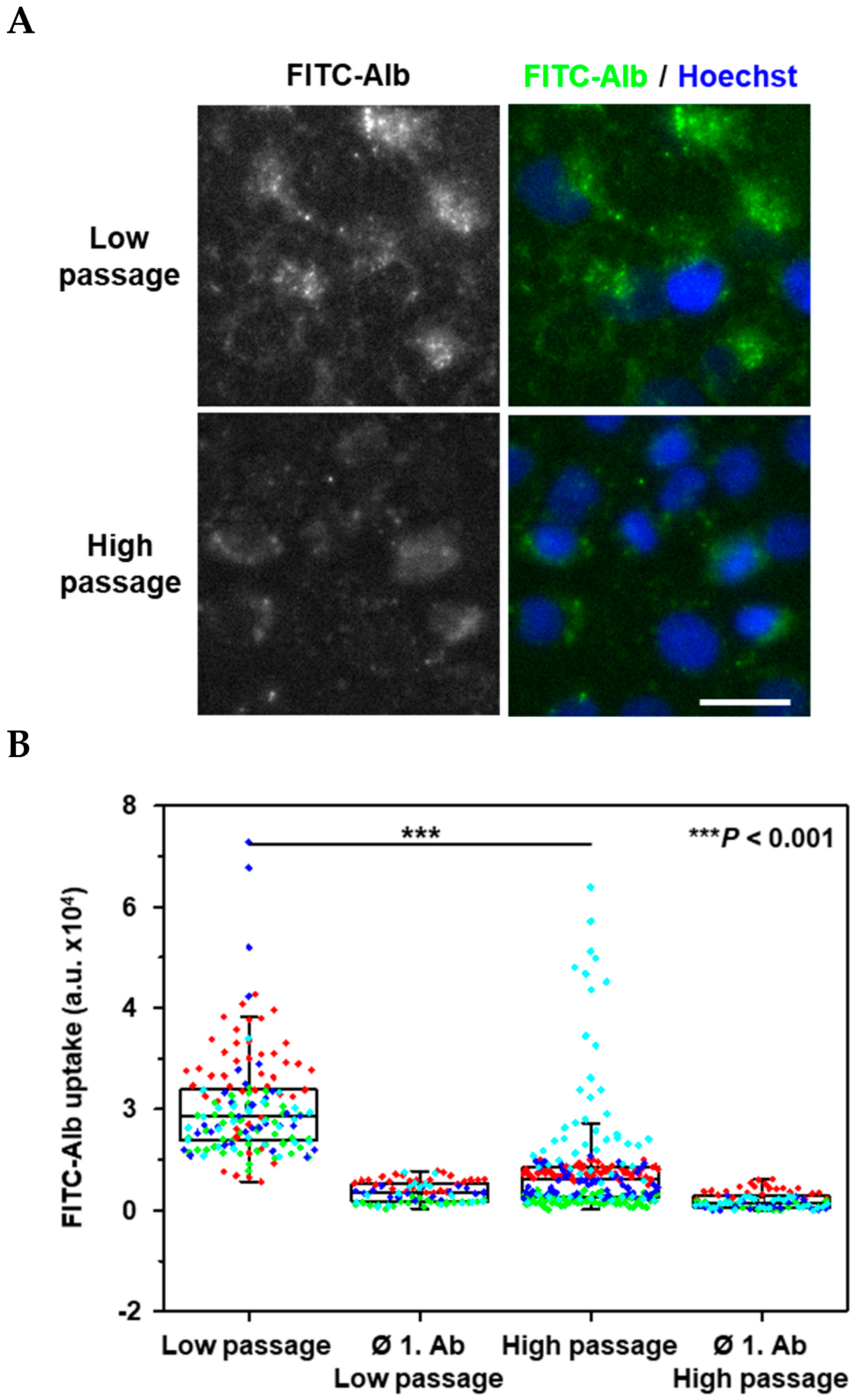
2. Results
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Cell Culture
4.2.2. Coupling of Cd2+-Protein Complexes
4.2.3. Cell Viability Assay of WKPT-0293 Cl.2 Cells
4.2.4. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.2.5. Immunofluorescence Staining of Megalin
4.2.6. Immunoblotting
4.2.7. Detection of Alb and β2M Uptake
4.2.8. Fluorescence Imaging
4.2.9. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization (WHO). Exposure to Cadmium: A Major Public Health Concern. 2010. Available online: https://www.who.int/ipcs/assessment/public_health/cadmium/en/ (accessed on 1 March 2019).
- Thévenod, F.; Lee, W.K. Toxicology of cadmium and its damage to mammalian organs. Metal. Ions Life Sci. 2013, 11, 415–490. [Google Scholar]
- Freisinger, E.; Vasak, M. Cadmium in metallothioneins. Met. Ions Life Sci. 2013, 11, 339–371. [Google Scholar]
- Liu, J.; Goyer, R.A.; Waalkes, M.P. Toxic effects of metals. In Casarett & Doull’s Toxicology: The Basic Science of Poisons, 7th ed.; Klaasen, C.D., Ed.; McGraw -Hill: New York, NY, USA, 2008; pp. 931–979. [Google Scholar]
- Sabolic, I.; Breljak, D.; Skarica, M.; Herak-Kramberger, C.M. Role of metallothionein in cadmium traffic and toxicity in kidneys and other mammalian organs. Biometals 2010, 23, 897–926. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, M.; Nordberg, G.F. Distribution of metallothionein-bound cadmium and cadmium chloride in mice: Preliminary studies. Environ. Health Perspect. 1975, 12, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Dudley, R.E.; Gammal, L.M.; Klaassen, C.D. Cadmium-induced hepatic and renal injury in chronically exposed rats: Likely role of hepatic cadmium-metallothionein in nephrotoxicity. Toxicol Appl. Pharmacol. 1985, 77, 414–426. [Google Scholar] [CrossRef]
- Nomiyama, K.; Nomiyama, H.; Kameda, N. Plasma cadmium-metallothionein, a biological exposure index for cadmium-induced renal dysfunction, based on the mechanism of its action. Toxicology 1998, 129, 157–168. [Google Scholar] [CrossRef]
- Thijssen, S.; Maringwa, J.; Faes, C.; Lambrichts, I.; Van Kerkhove, E. Chronic exposure of mice to environmentally relevant, low doses of cadmium leads to early renal damage, not predicted by blood or urine cadmium levels. Toxicology 2007, 229, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Klotz, K.; Weistenhofer, W.; Drexler, H. Determination of cadmium in biological samples. Met. Ions Life Sci. 2013, 11, 85–98. [Google Scholar] [PubMed]
- Milnerowicz, H.; Bizon, A. Determination of metallothionein in biological fluids using enzyme-linked immunoassay with commercial antibody. Acta Biochim. Pol. 2010, 57, 99–104. [Google Scholar] [CrossRef]
- Akintola, D.F.; Sampson, B.; Fleck, A. Development of an enzyme-linked immunosorbent assay for human metallothionein-1 in plasma and urine. J. Lab. Clin. Med. 1995, 126, 119–127. [Google Scholar] [PubMed]
- Thévenod, F. Nephrotoxicity and the proximal tubule. Insights from cadmium. Nephron Physiol. 2003, 93, 87–93. [Google Scholar] [CrossRef]
- Schuh, C.D.; Polesel, M.; Platonova, E.; Haenni, D.; Gassama, A.; Tokonami, N.; Ghazi, S.; Bugarski, M.; Devuyst, O.; Ziegler, U.; et al. Combined structural and functional imaging of the kidney reveals major axial differences in proximal tubule endocytosis. J. Am. Soc. Nephrol. 2018, 29, 2696–2712. [Google Scholar] [CrossRef]
- Christensen, E.I.; Birn, H.; Storm, T.; Weyer, K.; Nielsen, R. Endocytic receptors in the renal proximal tubule. Physiology 2012, 27, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Abouhamed, M.; Wolff, N.A.; Lee, W.K.; Smith, C.P.; Thévenod, F. Knockdown of endosomal/lysosomal divalent metal transporter 1 by rna interference prevents cadmium-metallothionein-1 cytotoxicity in renal proximal tubule cells. Am. J. Physiol. Renal Physiol. 2007, 293, F705–F712. [Google Scholar] [CrossRef]
- Thévenod, F.; Wolff, N.A. Iron transport in the kidney: Implications for physiology and cadmium nephrotoxicity. Met. Integr. Biometal Sci. 2016, 8, 17–42. [Google Scholar] [CrossRef]
- Harris, W.R.; Madsen, L.J. Equilibrium studies on the binding of cadmium(ii) to human serum transferrin. Biochemistry 1988, 27, 284–288. [Google Scholar] [CrossRef]
- Goumakos, W.; Laussac, J.P.; Sarkar, B. Binding of cadmium(ii) and zinc(ii) to human and dog serum albumins. An equilibrium dialysis and 113cd-nmr study. Biochem. Cell Biol. 1991, 69, 809–820. [Google Scholar] [CrossRef]
- Eakin, C.M.; Knight, J.D.; Morgan, C.J.; Gelfand, M.A.; Miranker, A.D. Formation of a copper specific binding site in non-native states of beta-2-microglobulin. Biochemistry 2002, 41, 10646–10656. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.I.; Birn, H. Megalin and cubilin: Synergistic endocytic receptors in renal proximal tubule. Am. J. Physiol. Renal Physiol. 2001, 280, F562–F573. [Google Scholar] [CrossRef]
- Klassen, R.B.; Crenshaw, K.; Kozyraki, R.; Verroust, P.J.; Tio, L.; Atrian, S.; Allen, P.L.; Hammond, T.G. Megalin mediates renal uptake of heavy metal metallothionein complexes. Am. J. Physiol. Renal Physiol. 2004, 287, F393–F403. [Google Scholar] [CrossRef] [PubMed]
- Wolff, N.A.; Abouhamed, M.; Verroust, P.J.; Thévenod, F. Megalin-dependent internalization of cadmium-metallothionein and cytotoxicity in cultured renal proximal tubule cells. J. Pharmacol. Exp. Ther. 2006, 318, 782–791. [Google Scholar] [CrossRef]
- Woost, P.G.; Orosz, D.E.; Jin, W.; Frisa, P.S.; Jacobberger, J.W.; Douglas, J.G.; Hopfer, U. Immortalization and characterization of proximal tubule cells derived from kidneys of spontaneously hypertensive and normotensive rats. Kidney Int. 1996, 50, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, R.; Birn, H.; Moestrup, S.K.; Nielsen, M.; Verroust, P.; Christensen, E.I. Characterization of a kidney proximal tubule cell line, llc-pk1, expressing endocytotic active megalin. J. Am. Soc. Nephrol. 1998, 9, 1767–1776. [Google Scholar] [PubMed]
- Zhai, X.Y.; Nielsen, R.; Birn, H.; Drumm, K.; Mildenberger, S.; Freudinger, R.; Moestrup, S.K.; Verroust, P.J.; Christensen, E.I.; Gekle, M. Cubilin- and megalin-mediated uptake of albumin in cultured proximal tubule cells of opossum kidney. Kidney Int. 2000, 58, 1523–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozyraki, R.; Fyfe, J.; Verroust, P.J.; Jacobsen, C.; Dautry-Varsat, A.; Gburek, J.; Willnow, T.E.; Christensen, E.I.; Moestrup, S.K. Megalin-dependent cubilin-mediated endocytosis is a major pathway for the apical uptake of transferrin in polarized epithelia. Proc. Natl. Acad. Sci. USA 2001, 98, 12491–12496. [Google Scholar] [CrossRef] [Green Version]
- Birn, H.; Fyfe, J.C.; Jacobsen, C.; Mounier, F.; Verroust, P.J.; Orskov, H.; Willnow, T.E.; Moestrup, S.K.; Christensen, E.I. Cubilin is an albumin binding protein important for renal tubular albumin reabsorption. J. Clin. Invest. 2000, 105, 1353–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storm, T.; Emma, F.; Verroust, P.J.; Hertz, J.M.; Nielsen, R.; Christensen, E.I. A patient with cubilin deficiency. N. Engl. J. Med. 2011, 364, 89–91. [Google Scholar] [CrossRef] [PubMed]
- Leheste, J.R.; Rolinski, B.; Vorum, H.; Hilpert, J.; Nykjaer, A.; Jacobsen, C.; Aucouturier, P.; Moskaug, J.O.; Otto, A.; Christensen, E.I.; et al. Megalin knockout mice as an animal model of low molecular weight proteinuria. Am. J. Pathol. 1999, 155, 1361–1370. [Google Scholar] [CrossRef]
- Hvidberg, V.; Jacobsen, C.; Strong, R.K.; Cowland, J.B.; Moestrup, S.K.; Borregaard, N. The endocytic receptor megalin binds the iron transporting neutrophil-gelatinase-associated lipocalin with high affinity and mediates its cellular uptake. FEBS Lett. 2005, 579, 773–777. [Google Scholar] [CrossRef]
- Norden, A.G.; Lapsley, M.; Lee, P.J.; Pusey, C.D.; Scheinman, S.J.; Tam, F.W.; Thakker, R.V.; Unwin, R.J.; Wrong, O. Glomerular protein sieving and implications for renal failure in fanconi syndrome. Kidney Int. 2001, 60, 1885–1892. [Google Scholar] [CrossRef]
- Shaikh, Z.A. Metallothionein as a storage protein for cadmium: Its toxicological implications. Dev. Toxicol. Environ. Sci. 1982, 9, 69–76. [Google Scholar]
- Liu, J.; Liu, Y.; Habeebu, S.S.; Klaassen, C.D. Susceptibility of mt-null mice to chronic cdcl2-induced nephrotoxicity indicates that renal injury is not mediated by the cdmt complex. Toxicol. Sci. 1998, 46, 197–203. [Google Scholar]
- Cherian, M.G.; Shaikh, Z.A. Metabolism of intravenously injected cadmium-binding protein. Biochem. Biophys. Res. Commun. 1975, 65, 863–869. [Google Scholar] [CrossRef]
- Cherian, M.G.; Goyer, R.A.; Delaquerriere-Richardson, L. Cadmium-metallothionein-induced nephropathy. Toxicol. Appl. Pharmacol. 1976, 38, 399–408. [Google Scholar]
- Foulkes, E.C. Renal tubular transport of cadmium-metallothionein. Toxicol. Appl. Pharmacol. 1978, 45, 505–512. [Google Scholar] [CrossRef]
- Bernard, A.M.; Ouled Amor, A.; Lauwerys, R.R. The effects of low doses of cadmium-metallothionein on the renal uptake of beta 2-microglobulin in rats. Toxicol. Appl. Pharmacol. 1987, 87, 440–445. [Google Scholar] [CrossRef]
- Tang, W.; Shaikh, Z.A. Renal cortical mitochondrial dysfunction upon cadmium metallothionein administration to sprague-dawley rats. J. Toxicol. Environ. Health A 2001, 63, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Habeebu, S.S.; Liu, Y.; Klaassen, C.D. Acute cdmt injection is not a good model to study chronic cd nephropathy: Comparison of chronic cdcl2 and cdmt exposure with acute cdmt injection in rats. Toxicol. Appl. Pharmacol. 1998, 153, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Cain, K.; Griffiths, B. Transfer of liver cadmium to the kidney after aflatoxin induced liver damage. Biochem. Pharmacol. 1980, 29, 1852–1855. [Google Scholar] [CrossRef]
- Tanaka, K.; Nomura, H.; Onosaka, S.; Min, K. Release of hepatic cadmium by carbon tetrachloride treatment. Toxicol. Appl. Pharmacol. 1981, 59, 535–539. [Google Scholar] [CrossRef]
- Tanaka, K. Effects of hepatic disorder on the fate of cadmium in rats. Dev. Toxicol. Environ. Sci. 1982, 9, 237–249. [Google Scholar]
- Chan, H.M.; Zhu, L.F.; Zhong, R.; Grant, D.; Goyer, R.A.; Cherian, M.G. Nephrotoxicity in rats following liver transplantation from cadmium-exposed rats. Toxicol. Appl. Pharmacol. 1993, 123, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Buchet, J.P.; Roels, H.; Masson, P.; Lauwerys, R. Renal excretion of proteins and enzymes in workers exposed to cadmium. Eur. J. Clin. Invest. 1979, 9, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Viau, C.; Lauwerys, R. Renal handling of human beta 2-microglobulin in normal and cadmium-poisoned rats. Arch. Toxicol. 1983, 53, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Prozialeck, W.C.; Edwards, J.R. Early biomarkers of cadmium exposure and nephrotoxicity. Biometals 2010, 23, 793–809. [Google Scholar] [CrossRef] [PubMed]
- Dickson, L.E.; Wagner, M.C.; Sandoval, R.M.; Molitoris, B.A. The proximal tubule and albuminuria: Really! J. Am. Soc. Nephrol. 2014, 25, 443–453. [Google Scholar] [CrossRef]
- Norden, A.G.; Sharratt, P.; Cutillas, P.R.; Cramer, R.; Gardner, S.C.; Unwin, R.J. Quantitative amino acid and proteomic analysis: Very low excretion of polypeptides >750 da in normal urine. Kidney Int. 2004, 66, 1994–2003. [Google Scholar] [CrossRef]
- Torra, M.; To-Figueras, J.; Brunet, M.; Rodamilans, M.; Corbella, J. Total and metallothionein-bound cadmium in the liver and the kidney of a population in barcelona (spain). Bull. Environ. Contam. Toxicol. 1994, 53, 509–515. [Google Scholar] [CrossRef]
- Yoshida, M.; Ohta, H.; Yamauchi, Y.; Seki, Y.; Sagi, M.; Yamazaki, K.; Sumi, Y. Age-dependent changes in metallothionein levels in liver and kidney of the japanese. Biol. Trace Elem. Res. 1998, 63, 167–175. [Google Scholar] [CrossRef]
- Garrett, S.H.; Sens, M.A.; Todd, J.H.; Somji, S.; Sens, D.A. Expression of mt-3 protein in the human kidney. Toxicol. Lett. 1999, 105, 207–214. [Google Scholar] [CrossRef]
- Nagamine, T.; Nakazato, K.; Suzuki, K.; Kusakabe, T.; Sakai, T.; Oikawa, M.; Satoh, T.; Kamiya, T.; Arakawa, K. Analysis of tissue cadmium distribution in chronic cadmium-exposed mice using in-air micro-pixe. Biol. Trace Elem. Res. 2007, 117, 115–126. [Google Scholar] [CrossRef]
- Wang, L.; Chen, D.; Wang, H.; Liu, Z. Effects of lead and/or cadmium on the expression of metallothionein in the kidney of rats. Biol. Trace Elem. Res. 2009, 129, 190–199. [Google Scholar] [CrossRef]
- Langelueddecke, C.; Roussa, E.; Fenton, R.A.; Wolff, N.A.; Lee, W.K.; Thévenod, F. Lipocalin-2 (24p3/neutrophil gelatinase-associated lipocalin (ngal)) receptor is expressed in distal nephron and mediates protein endocytosis. J. Biol. Chem. 2012, 287, 159–169. [Google Scholar] [CrossRef]
- Smith, H.W. The fate of sodium and water in the renal tubules. Bull. N. Y. Acad. Med. 1959, 35, 293–316. [Google Scholar]
- Ullrich, K.J. Niere. In Kurzgefaßtes Lehrbuch der Physiologie; Keidel, W.D., Ed.; Georg Thieme Verlag: Stuttgart, Germany, 1975. [Google Scholar]
- Girolami, J.P.; Bascands, J.L.; Pecher, C.; Cabos, G.; Moatti, J.P.; Mercier, J.F.; Haguenoer, J.M.; Manuel, Y. Renal kallikrein excretion as a distal nephrotoxicity marker during cadmium exposure in rats. Toxicology 1989, 55, 117–129. [Google Scholar] [CrossRef]
- Oner, G.; Senturk, U.K.; Izgut-Uysal, N. The role of cadmium in the peroxidative response of kidney to stress. Biol. Trace Elem. Res. 1995, 48, 111–117. [Google Scholar] [CrossRef]
- Lauwerys, R.; Bernard, A. Preclinical detection of nephrotoxicity: Description of the tests and appraisal of their health significance. Toxicol. Lett. 1989, 46, 13–29. [Google Scholar] [CrossRef]
- Lee, W.K.; Bork, U.; Gholamrezaei, F.; Thévenod, F. Cd(2+)-induced cytochrome c release in apoptotic proximal tubule cells: Role of mitochondrial permeability transition pore and ca(2+) uniporter. Am. J. Physiol. Renal Physiol. 2005, 288, F27–F39. [Google Scholar] [CrossRef]
- Erfurt, C.; Roussa, E.; Thevenod, F. Apoptosis by cd2+ or cdmt in proximal tubule cells: Different uptake routes and permissive role of endo/lysosomal cdmt uptake. Am. J. Physiol. Cell Physiol. 2003, 285, C1367–C1376. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Prabakaran, T.; Christensen, E.I.; Nielsen, R.; Verroust, P.J. Cubilin is expressed in rat and human glomerular podocytes. Nephrol. Dial. Transplant. 2012, 27, 3156–3159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with imagej. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Receptor | KD (nmol/L) | Reference | Concentration in Glomerular Filtrate (nmol/L) * |
|---|---|---|---|---|
| MT | Megalin | 100,000 | [22] | 0.5–5 |
| β2M | Megalin | 420 | [30] | 100 |
| Lcn2 | Megalin | 60 | [31] | 650 |
| Tf | Cubilin | 20 | [27] | 2 |
| Alb | Cubilin | 630 | [28] | 53 |
| α1M | Cubilin | n.d. | Ø | 92 |
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| Rat megalin/Lrp2 (NM_030827.1) a | TGGAATCTCCCTTGATCCTG | TGTTGCTGCCATCAGTCTTC |
| Rat cubilin/Cubn (NM_053332) a | GCACTGGCAATGAACTAGCA | TGATCCAGGAGCACTCTGTG |
| Rat Gapdh (NM_017008) | AGGGCTCATGACCACAGT | TGCAGGGATGATGTTCTG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fels, J.; Scharner, B.; Zarbock, R.; Zavala Guevara, I.P.; Lee, W.-K.; Barbier, O.C.; Thévenod, F. Cadmium Complexed with β2-Microglubulin, Albumin and Lipocalin-2 rather than Metallothionein Cause Megalin:Cubilin Dependent Toxicity of the Renal Proximal Tubule. Int. J. Mol. Sci. 2019, 20, 2379. https://doi.org/10.3390/ijms20102379
Fels J, Scharner B, Zarbock R, Zavala Guevara IP, Lee W-K, Barbier OC, Thévenod F. Cadmium Complexed with β2-Microglubulin, Albumin and Lipocalin-2 rather than Metallothionein Cause Megalin:Cubilin Dependent Toxicity of the Renal Proximal Tubule. International Journal of Molecular Sciences. 2019; 20(10):2379. https://doi.org/10.3390/ijms20102379
Chicago/Turabian StyleFels, Johannes, Bettina Scharner, Ralf Zarbock, Itzel Pamela Zavala Guevara, Wing-Kee Lee, Olivier C. Barbier, and Frank Thévenod. 2019. "Cadmium Complexed with β2-Microglubulin, Albumin and Lipocalin-2 rather than Metallothionein Cause Megalin:Cubilin Dependent Toxicity of the Renal Proximal Tubule" International Journal of Molecular Sciences 20, no. 10: 2379. https://doi.org/10.3390/ijms20102379