Transcriptome Analysis of Larval Segment Formation and Secondary Loss in the Echiuran Worm Urechis unicinctus

 , , and
, , and
Abstract
:1. Introduction
2. Results
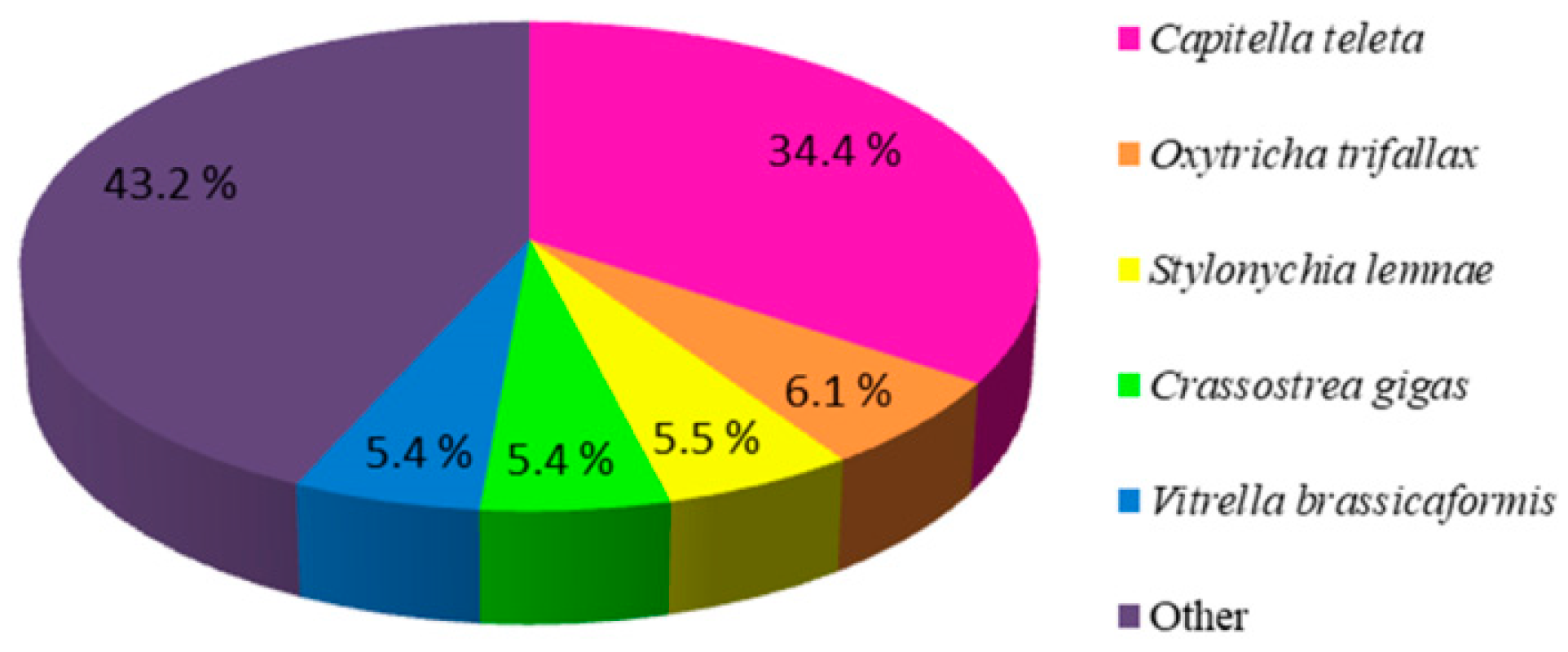
2.1. Illumina Sequencing, De Novo Assembly and Functional Annotation
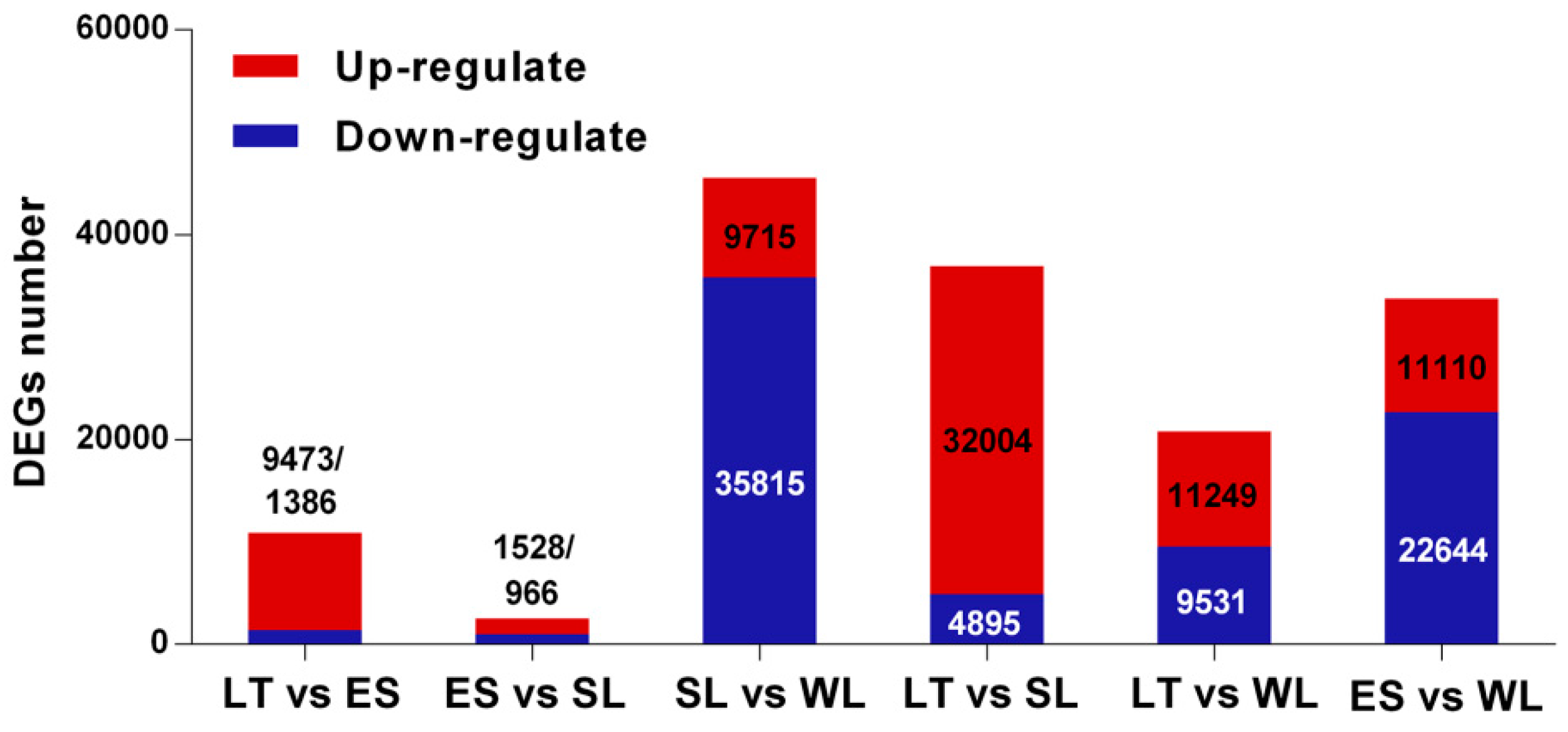
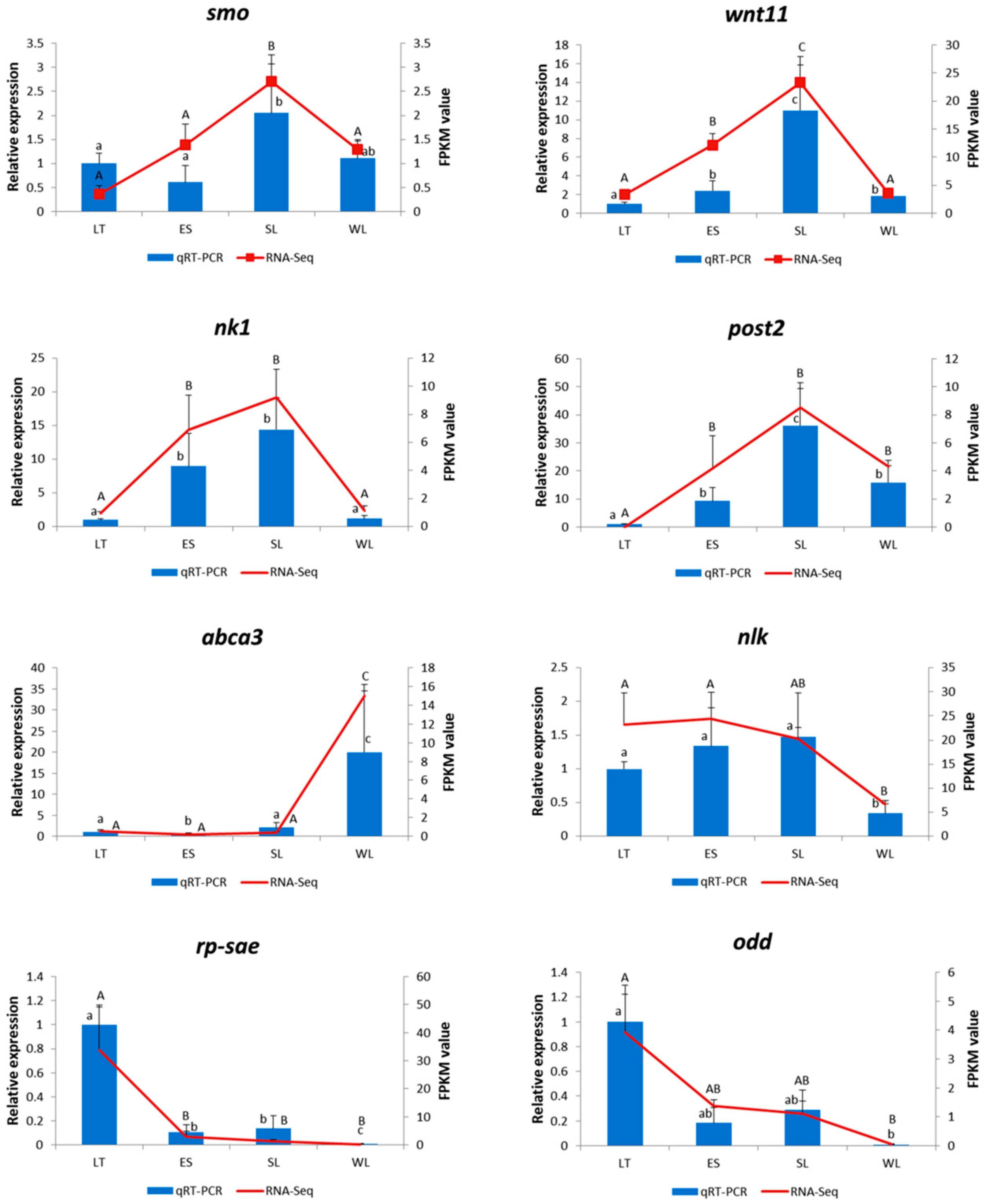
2.2. Identification, Classification, and Validation of the Differentially Expressed Genes (DEGs)
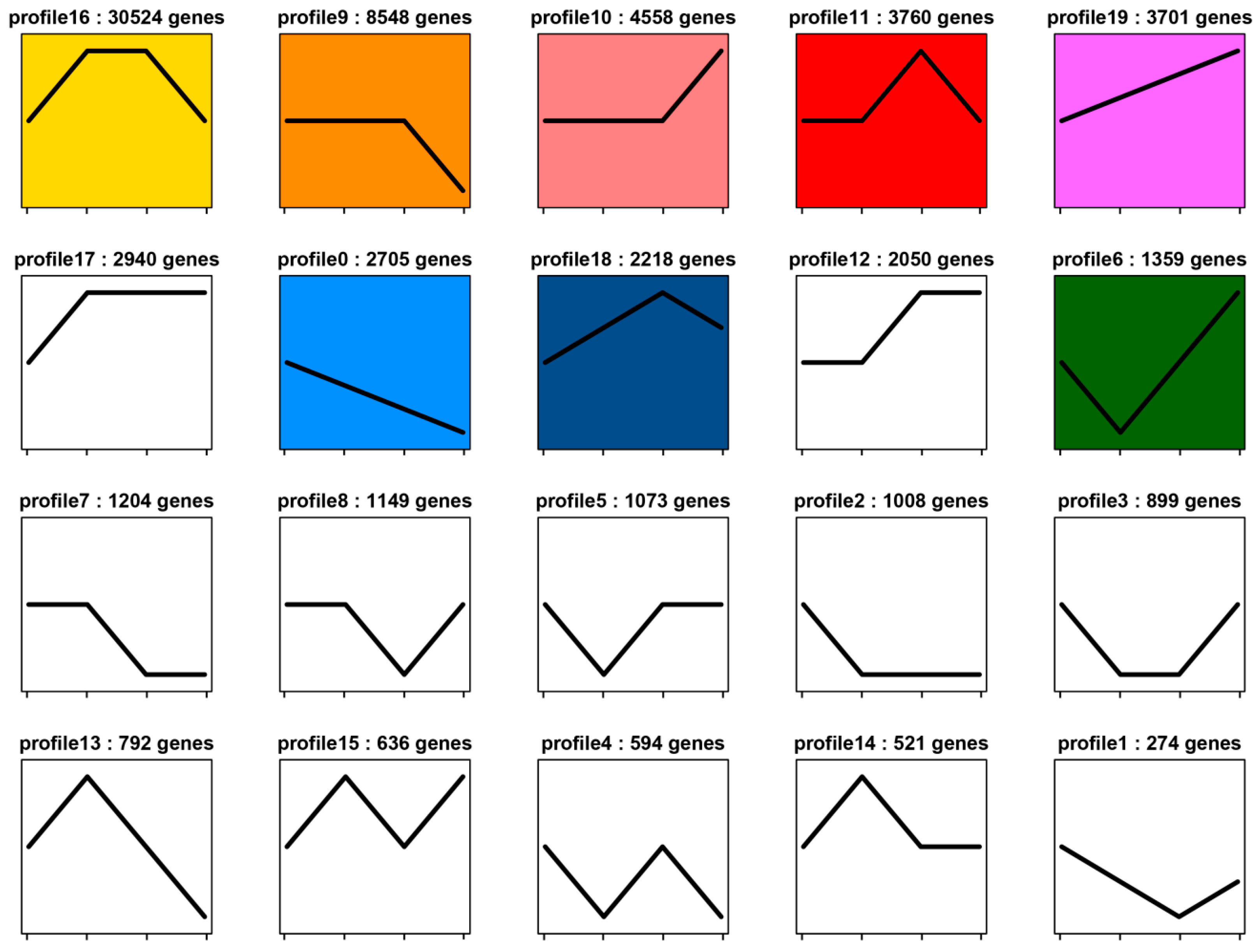
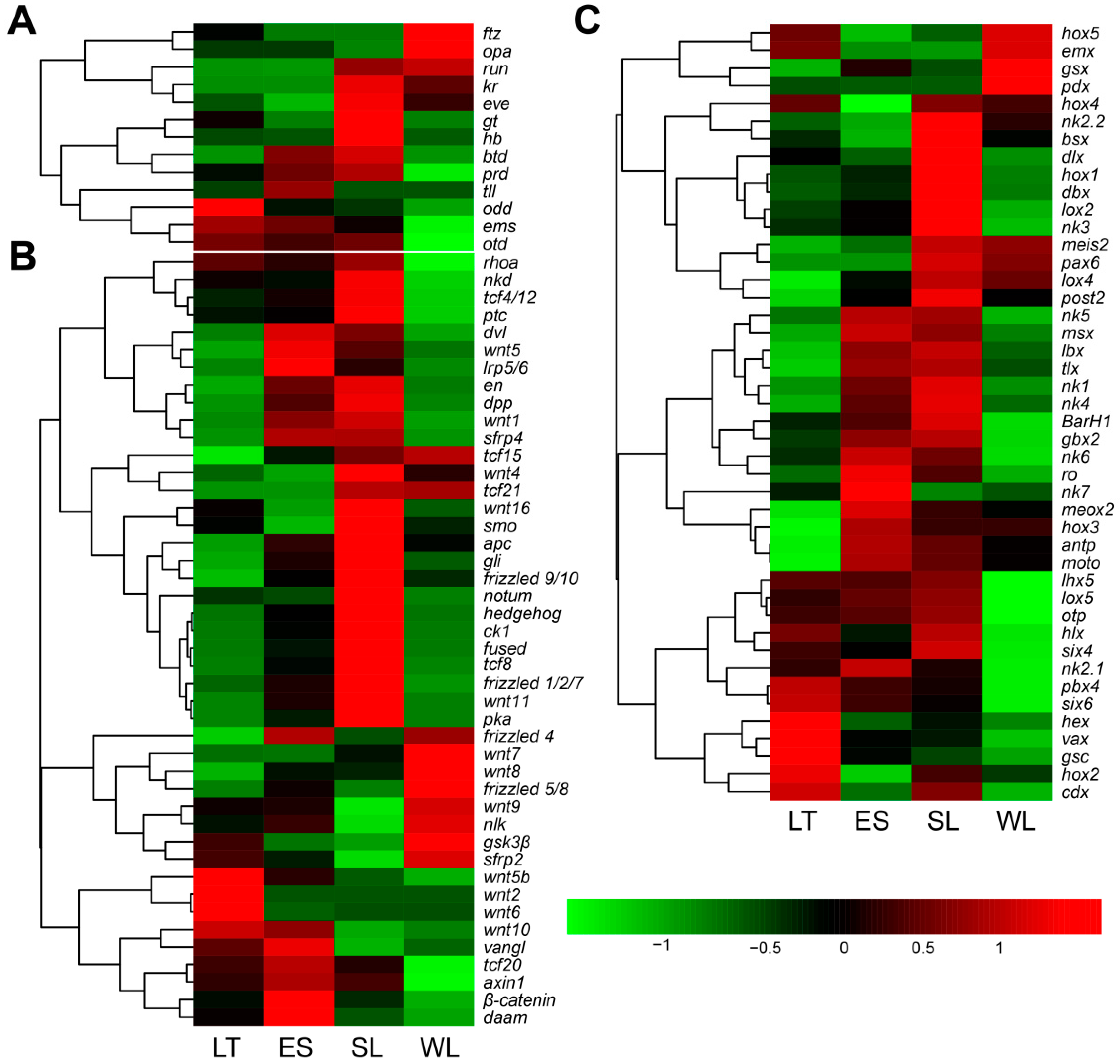
2.3. Expression Profiles of Genes Related to Segment Formation
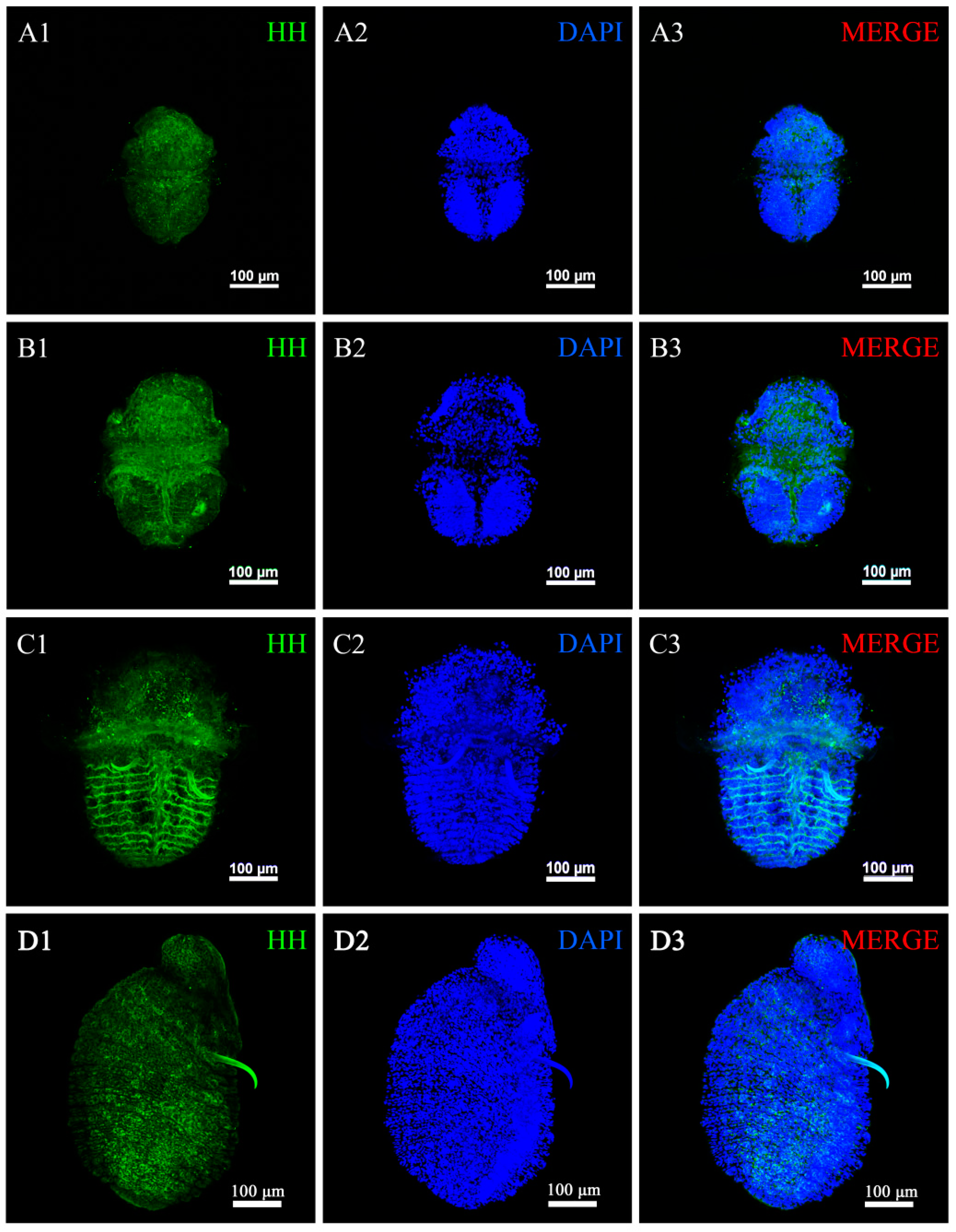
2.4. Expression Characteristics of Hedgehog (HH) in U. unicinctus Larvae
3. Discussion
3.1. Function of Gap and Pair-Rule Genes in U. unicinctus Was Not Consistent with Drosophila during Segmentation
3.2. Key Genes and Pathways Involved in U. unicinctus Larval Segment Formation and Secondary Loss
3.3. Hedgehog Is a Conservative Gene for the Larval Segment Boundary Definition in U. unicinctus
4. Materials and Methods
4.1. Ethics Statement
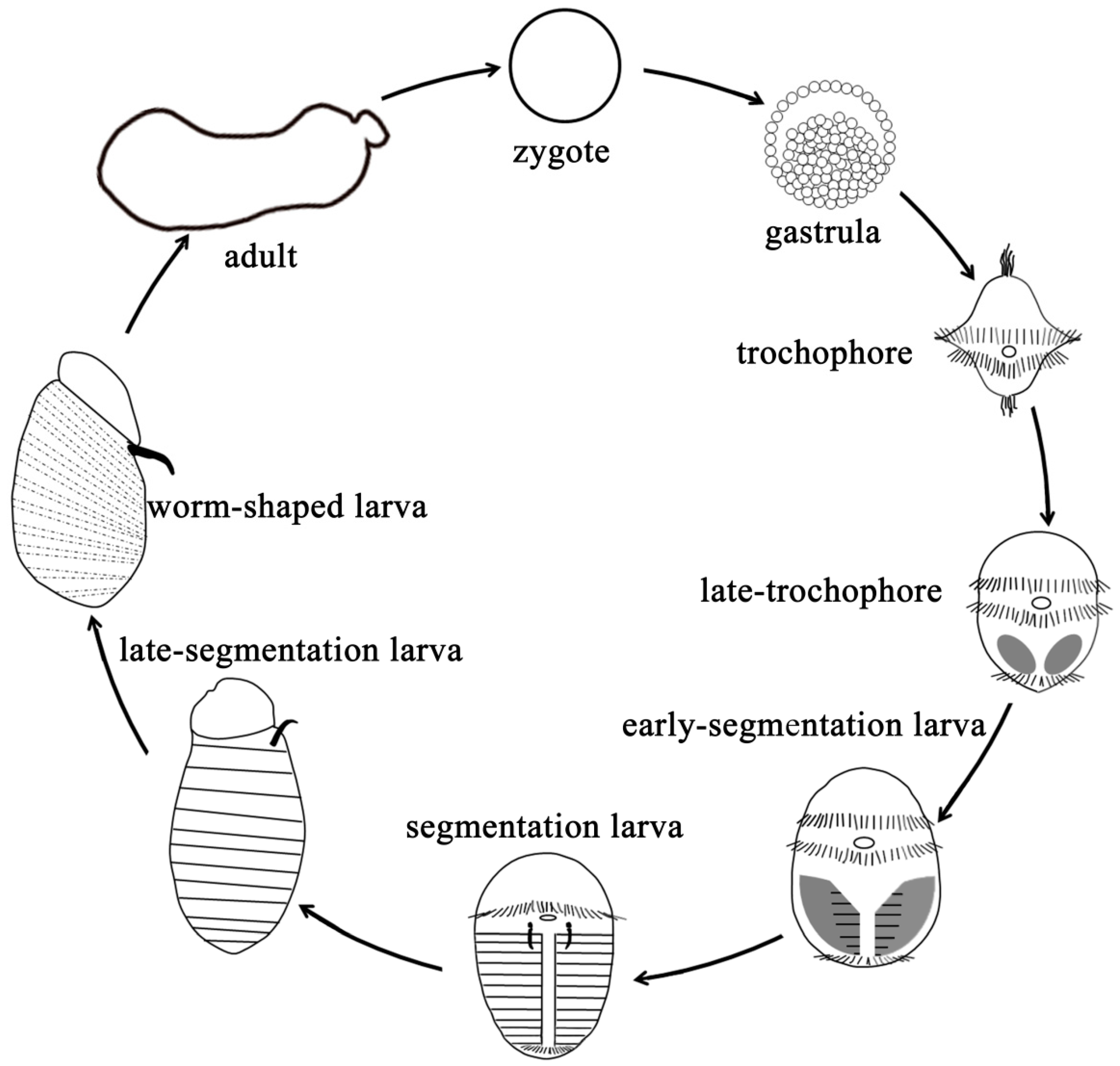
4.2. Animals and Sampling
4.3. RNA Extraction, RNA-Seq Library Construction, and Sequencing
4.4. De Novo Assembly and Functional Annotation
4.5. Enrichment and Dynamic Expression Profile of Differentially Expressed Genes (DEGs)
4.6. Validation of RNA-Seq Data with Quantitative RT-PCR (qRT-PCR)
4.7. Immunofluorescence Histochemistry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| LT | late-trochophore |
| ES | early-segmentation larva |
| SL | segmentation larva |
| WL | worm-shaped larva |
| DEG | Differentially expressed gene |
| FDR | False discovery rate |
| STEM | Short time-series expression miner |
| qRT-PCR | Quantitative real-time PCR |
| FPKM | Fragments per kilobase of transcript sequence per million base pairs sequenced |
| HH | Hedgehog protein |
References
- Goto, R.; Okamoto, T.; Ishikawa, H.; Hamamura, Y.; Kato, M. Molecular phylogeny of echiuran worms (Phylum: Annelida) reveals evolutionary pattern of feeding mode and sexual dimorphism. PLoS ONE 2013, 8, e56809. [Google Scholar] [CrossRef] [PubMed]
- Hessling, R.; Westheide, W. Are Echiura derived from a segmented ancestor? Immunohistochemical analysis of the nervous system in developmental stages of Bonellia viridis. J. Morphol. 2002, 252, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Q.; Fan, Q.H.; Pesic, V.; Smit, H.; Bochkov, A.V.; Khaustov, A.A.; Baker, A.; Wohltmann, A.; Wen, T.; Amrine, J.W. Animal Biodiversity: An Outline of Higher-Level Classification and Survey of Taxonomic Richness; Magnolia Press: Auckland, New Zealand, 2011; pp. 10–100. [Google Scholar]
- Hessling, R. Metameric organisation of the nervous system in developmental stages of Urechis caupo (Echiura) and its phylogenetic implications. Zoomorphology 2002, 121, 221–234. [Google Scholar] [CrossRef]
- Nielsen, C. Spezielle Zoologie. Teil 1: Einzeller und Wirbellose Tiere. Acta. Zool. 2008, 89, 179–180. [Google Scholar] [CrossRef]
- Purschke, G.; Hessling, R.; Westheide, W. The phylogenetic position of the Clitellata and the Echiura—On the problematic assessment of absent characters. J. Zool. Syst. Evol. Res. 2010, 38, 165–173. [Google Scholar] [CrossRef]
- Struck, T.H.; Paul, C.; Hill, N.; Hartmann, S.; Hösel, C.; Kube, M.; Lieb, B.; Meyer, A.; Tiedemann, R.; Purschke, G. Phylogenomic analyses unravel annelid evolution. Nature 2011, 471, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Struck, T.H.; Schult, N.; Kusen, T.; Hickman, E.; Bleidorn, C.; Mchugh, D.; Halanych, K.M. Annelid phylogeny and the status of Sipuncula and Echiura. BMC Evol. Biol. 2007, 7, 57. [Google Scholar] [CrossRef]
- Wu, Z.; Shen, X.; Sun, M.; Ren, J.; Wang, Y.; Huang, Y.; Liu, B. Phylogenetic analyses of complete mitochondrial genome of Urechis unicinctus (Echiura) support that echiurans are derived annelids. Mol. Phylogenet. Evol. 2009, 52, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Barneswriter, R. Invertebrate Zoology, 5th ed.; Saunders College Publishing: Eastbourne, UK, 1987; pp. 25–79. [Google Scholar]
- Tilic, E.; Lehrke, J.; Bartolomaeus, T. Homology and evolution of the chaetae in Echiura (Annelida). PLoS ONE 2015, 10, e0120002. [Google Scholar] [CrossRef] [PubMed]
- Mito, T.; Shinmyo, Y.; Kurita, K.; Nakamura, T.; Ohuchi, H.; Noji, S. Ancestral functions of Delta/Notch signaling in the formation of body and leg segments in the cricket Gryllus bimaculatus. Development 2011, 138, 3823–3833. [Google Scholar] [CrossRef]
- Balavoine, G. Segment formation in Annelids: Patterns, processes and evolution. Int. J. Dev. Biol. 2014, 58, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Dill, K.K.; Thamm, K.; Seaver, E.C. Characterization of twist and snail gene expression during mesoderm and nervous system development in the polychaete annelid Capitella sp. I. Dev. Genes. Evol. 2007, 217, 435–447. [Google Scholar] [CrossRef]
- Franke, H.D. Reproduction of the Syllidae (Annelida: Polychaeta). Hydrobiologia 1999, 402, 39–55. [Google Scholar] [CrossRef]
- Xie, Y.Y.; Wei, M.K.; Li, Q.; Qin, Z.K.; Zhang, Z.F. Organogenesis of body wall and formation of larval segment in Urechis unicinctus. Can. J. Zool. 2019. under review. [Google Scholar]
- Pourquié, O. The segmentation clock: Converting embryonic time into spatial pattern. Science 2003, 301, 328–330. [Google Scholar] [CrossRef]
- Gibb, S.; Maroto, M.; Dale, J.K. The segmentation clock mechanism moves up a notch. Trends Cell Biol. 2010, 20, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Dray, N.; Tessmar-Raible, K.; Le Gouar, M.; Vibert, L.; Christodoulou, F.; Schipany, K.; Guillou, A.; Zantke, J.; Snyman, H.; Behague, J.; et al. Hedgehog signaling regulates segment formation in the annelid Platynereis. Science 2010, 329, 339–342. [Google Scholar] [CrossRef]
- Farzana, L.; Brown, S.J. Hedgehog signaling pathway function conserved in Tribolium segmentation. Dev. Genes Evol. 2008, 218, 181–192. [Google Scholar] [CrossRef]
- Aulehla, A.; Pourquié, O. Oscillating signaling pathways during embryonic development. Curr. Opin. Cell Biol. 2008, 20, 632–635. [Google Scholar] [CrossRef] [PubMed]
- Peel, A.D.; Chipman, A.D.; Akam, M. Arthropod segmentation: Beyond the Drosophila paradigm. Nat. Rev. Genet. 2005, 6, 905–916. [Google Scholar] [CrossRef]
- Seaver, E.C.; Yamaguchi, E.; Richards, G.S.; Meyer, N.P. Expression of the pair-rule gene homologs runt, Pax3/7, even-skipped-1 and even-skipped-2 during larval and juvenile development of the polychaete annelid Capitella teleta does not support a role in segmentation. EvoDevo 2012, 3, 8. [Google Scholar] [CrossRef]
- Hui, J.H.; Raible, F.; Korchagina, N.; Dray, N.; Samain, S.; Magdelenat, G.; Jubin, C.; Segurens, B.; Balavoine, G.; Arendt, D. Features of the ancestral bilaterian inferred from Platynereis dumerilii ParaHox genes. BMC Biol. 2009, 7, 43. [Google Scholar] [CrossRef]
- Saudemont, A.; Dray, N.; Hudry, B.; Gouar, M.L.; Vervoort, M.; Balavoine, G. Complementary striped expression patterns of NK homeobox genes during segment formation in the annelid Platynereis. Dev. Biol. 2008, 317, 430–443. [Google Scholar] [CrossRef]
- Seaver, E.C.; Kaneshige, L.M. Expression of ‘segmentation’ genes during larval and juvenile development in the polychaetes Capitella sp. I and H. elegans. Dev. Biol. 2006, 289, 179–194. [Google Scholar] [CrossRef]
- Steinmetz, P.R.; Kostyuchenko, R.P.; Fischer, A.; Arendt, D. The segmental pattern of otx, gbx, and Hox genes in the annelid Platynereis dumerilii. Evol. Dev. 2011, 13, 72–79. [Google Scholar] [CrossRef]
- Zhang, L.T.; Liu, X.L.; Liu, J.G.; Zhang, Z.F. Characteristics and function of sulfur dioxygenase in Echiuran worm Urechis unicinctus. PLoS ONE 2013, 8, e81885. [Google Scholar] [CrossRef]
- Liu, X.L.; Zhang, L.T.; Zhang, Z.F.; Ma, X.Y.; Liu, J.G. Transcriptional response to sulfide in the Echiuran Worm Urechis unicinctus by digital gene expression analysis. BMC Genom. 2015, 16, 829. [Google Scholar] [CrossRef]
- Ernst, J.; Bar-Joseph, Z. STEM: A tool for the analysis of short time series gene expression data. BMC Bioinform. 2006, 7, 191. [Google Scholar] [CrossRef]
- Chen, Z.T. Primary Study on the Early Development in Urechis uniconctus. Master’s Thesis, Ocean University of China, Qingdao, China, 2005. (In Chinese). [Google Scholar]
- Larsen, C.W.; Hirst, E.; Alexandre, C.; Vincent, J.P. Segment boundary formation in Drosophila embryos. Development 2003, 130, 5625–5635. [Google Scholar] [CrossRef]
- Treffkorn, S.; Kahnke, L.; Hering, L.; Mayer, G. Expression of NK cluster genes in the onychophoran Euperipatoides rowelli: Implications for the evolution of NK family genes in nephrozoans. EvoDevo 2018, 9, 17. [Google Scholar] [CrossRef]
- Perrimon, N.; Mahowald, A.P. Multiple functions of segment polarity genes in Drosophila. Dev. Biol. 1987, 119, 587–600. [Google Scholar] [CrossRef]
- Wallingford, J.B.; Habas, R. The developmental biology of Dishevelled: An enigmatic protein governing cell fate and cell polarity. Development 2005, 132, 4421–4432. [Google Scholar] [CrossRef]
- Alcedo, J.; Ayzenzon, M.; Von, T.O.; Noll, M.; Hooper, J.E. The Drosophila smoothened gene encodes a seven-pass membrane protein, a putative receptor for the hedgehog signal. Cell 1996, 86, 221–232. [Google Scholar] [CrossRef]
- Mimeault, M.; Batra, S.K. Frequent deregulations in the hedgehog signaling network and cross-talks with the epidermal growth factor receptor pathway involved in cancer progression and targeted therapies. Pharmacol. Rev. 2010, 62, 497–524. [Google Scholar] [CrossRef]
- Peifer, M.; Wieschaus, E. The segment polarity gene armadillo encodes a functionally modular protein that is the Drosophila homolog of human plakoglobin. Cell 1990, 63, 1167–1178. [Google Scholar] [CrossRef]
- Dassow, G.V.; Meir, E.; Munro, E.M.; Odell, G.M. The segment polarity network is a robust developmental module. Nature 2000, 406, 188–192. [Google Scholar] [CrossRef]
- Ingham, P.W. Hedgehog signaling. Cold. Spring. Harb. Perspect. Biol. 2012, 4, 1–2. [Google Scholar] [CrossRef]
- Pechmann, M.; Mcgregor, A.P.; Schwager, E.E.; Feitosa, N.M.; Damen, W.G. Dynamic gene expression is required for anterior regionalization in a spider. Proc. Natl. Acad. Sci. USA 2009, 106, 1468–1472. [Google Scholar] [CrossRef] [Green Version]
- Simonnet, F.; Deutsch, J.; Quéinnec, E. Hedgehog is a segment polarity gene in a crustacean and a chelicerate. Dev. Genes. Evol. 2004, 214, 537–545. [Google Scholar] [CrossRef]
- Prud’homme, B.; Rosa, R.D.; Arendt, D.; Julien, J.F.; Pajaziti, R.; Dorresteijn, A.W.; Adoutte, A.; Wittbrodt, J.; Balavoine, G. Arthropod-like expression patterns of engrailed and wingless in the annelid Platynereis dumerilii suggest a role in segment formation. Curr. Biol. 2003, 13, 1876–1881. [Google Scholar] [CrossRef]
- Janssen, R.; Gouar, M.L.; Pechmann, M.; Poulin, F.; Bolognesi, R.; Schwager, E.E.; Hopfen, C.; Colbourne, J.K.; Budd, G.E.; Brown, S.J. Conservation, loss, and redeployment of Wnt ligands in protostomes: Implications for understanding the evolution of segment formation. BMC Evol. Biol. 2010, 10, 374. [Google Scholar] [CrossRef]
- Damen, W.G.M. Evolutionary conservation and divergence of the segmentation process in arthropods. Dev. Dyn. 2010, 236, 1379–1391. [Google Scholar] [CrossRef]
- Jiang, Y.J.; Aerne, B.L.; Smithers, L.; Haddon, C.; Ish-Horowicz, D.; Lewis, J. Notch signalling and the synchronization of the somite segmentation clock. Nature 2000, 408, 475–479. [Google Scholar] [CrossRef]
- Beisner, J.; Buck, M.B.; Fritz, P.; Dippon, J.; Schwab, M.; Brauch, H.; Zugmaier, G.; Pfizenmaier, K.; Knabbe, C. A Novel Functional Polymorphism in the Transforming Growth Factor-β2 Gene Promoter and Tumor Progression in Breast Cancer. Cancer Res. 2006, 66, 7554–7561. [Google Scholar] [CrossRef]
- Morgensztern, D.; Mcleod, H.L. PI3K/Akt/mTOR pathway as a target for cancer therapy. Anti-Cancer Drugs 2005, 16, 797–803. [Google Scholar] [CrossRef]
- Ghiglione, C. The Transmembrane Molecule Kekkon 1 Acts in a Feedback Loop to Negatively Regulate the Activity of the Drosophila EGF Receptor during Oogenesis. Cell 1999, 96, 847–856. [Google Scholar] [CrossRef]
- Luo, W.; Cao, X.; Xu, X.; Huang, S.; Liu, C.; Tomljanovic, T. Developmental transcriptome analysis and identification of genes involved in formation of intestinal air-breathing function of Dojo loach, Misgurnus anguillicaudatus. Sci. Rep. 2016, 6, 31845. [Google Scholar] [CrossRef]
- Ingham, P.W. Localized hedgehog activity controls spatial limits of wingless transcription in the Drosophila embryo. Nature 1993, 366, 560–562. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Clean Bases | Error (%) | Q20 (%) | Q30 (%) | GC (%) |
|---|---|---|---|---|---|---|---|
| LT_1 | 44,405,794 | 43,232,040 | 6.48 Gb | 0.02 | 95.41 | 89.03 | 46.93 |
| LT_2 | 45,768,482 | 44,516,626 | 6.68 Gb | 0.02 | 95.44 | 89.05 | 41.53 |
| LT_3 | 51,280,946 | 49,850,242 | 7.48 Gb | 0.01 | 97.44 | 93.49 | 45 |
| ES_1 | 49,666,198 | 48,375,494 | 7.26 Gb | 0.01 | 97.75 | 94.21 | 45.34 |
| ES_2 | 46,441,732 | 45,203,342 | 6.78 Gb | 0.01 | 97.81 | 94.3 | 47.12 |
| ES_3 | 52,824,412 | 51,511,036 | 7.73 Gb | 0.01 | 97.8 | 94.29 | 45.62 |
| SL_1 | 50,346,424 | 48,909,398 | 7.34 Gb | 0.01 | 97.83 | 94.34 | 47.75 |
| SL_2 | 51,551,170 | 50,039,830 | 7.51 Gb | 0.01 | 97.62 | 93.86 | 49.64 |
| SL_3 | 65,816,438 | 63,885,972 | 9.58 Gb | 0.01 | 97.71 | 94.1 | 46.84 |
| WL_1 | 55,190,790 | 53,346,656 | 8 Gb | 0.01 | 97.6 | 93.8 | 48.91 |
| WL_2 | 50,393,708 | 48,640,910 | 7.3 Gb | 0.01 | 97.58 | 93.78 | 48.88 |
| WL_3 | 45,847,850 | 44,065,776 | 6.61 Gb | 0.01 | 97.64 | 93.9 | 48.68 |
| Min Length | Mean Length | Median Length | Max Length | N50 | N90 | Total Nucleotides | |
|---|---|---|---|---|---|---|---|
| Transcripts | 201 | 640 | 291 | 19,744 | 1206 | 245 | 355,841,139 |
| Unigenes | 201 | 1125 | 654 | 19,744 | 1836 | 464 | 273,911,152 |
| Item | NR | NT | KO | SwissProt | Pfam | GO | KOG | In All Databases | At Least One Database |
|---|---|---|---|---|---|---|---|---|---|
| No. of genes | 122,354 | 22,899 | 33,027 | 94,085 | 109,847 | 111,479 | 71,895 | 7704 | 149,488 |
| Percentage | 50.27% | 9.4% | 13.57% | 38.65% | 45.13% | 45.8% | 29.54% | 3.16% | 61.42% |
| Pathway | Pathway ID | p-Value (profile9) | p-Value (profile16) |
|---|---|---|---|
| Hedgehog signaling pathway | ko04341 | 0.04767 | 0.00608 |
| Wnt signaling pathway | ko04310 | 0.03753 | 0.00201 |
| TGF-beta signaling pathway | ko04350 | 0.03045 | 0.01209 |
| PI3K-AKT signaling pathway | ko04068 | ― | 0.00428 |
| mTOR signaling pathway | ko04150 | ― | 0.00012 |
| MAPK signaling pathway | ko04010 | ― | 0.0085 |
| Notch signaling pathway | ko04330 | 0.04927 | ― |
| Dorso-ventral axis formation | ko04320 | 0.00831 | 0.00909 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, X.; Wei, M.; Li, Q.; Zhang, T.; Zhou, D.; Kong, D.; Xie, Y.; Qin, Z.; Zhang, Z. Transcriptome Analysis of Larval Segment Formation and Secondary Loss in the Echiuran Worm Urechis unicinctus. Int. J. Mol. Sci. 2019, 20, 1806. https://doi.org/10.3390/ijms20081806
Hou X, Wei M, Li Q, Zhang T, Zhou D, Kong D, Xie Y, Qin Z, Zhang Z. Transcriptome Analysis of Larval Segment Formation and Secondary Loss in the Echiuran Worm Urechis unicinctus. International Journal of Molecular Sciences. 2019; 20(8):1806. https://doi.org/10.3390/ijms20081806
Chicago/Turabian StyleHou, Xitan, Maokai Wei, Qi Li, Tingting Zhang, Di Zhou, Dexu Kong, Yueyang Xie, Zhenkui Qin, and Zhifeng Zhang. 2019. "Transcriptome Analysis of Larval Segment Formation and Secondary Loss in the Echiuran Worm Urechis unicinctus" International Journal of Molecular Sciences 20, no. 8: 1806. https://doi.org/10.3390/ijms20081806