1. Introduction
Malignant melanoma is a skin cancer that develops from the abnormal growth and differentiation of melanocytes with hyperpigmentation; the incidence of melanoma cases has been increasing, and this particular skin cancer is associated with a high rate of mortality caused by early and rapid metastasis [
1]. Significant therapeutic advances have been made using small molecule inhibitors that target melanoma, but challenges to eradicate these solid tumors still persist.
Given the fact that a hypoxic microenvironment is a major feature in multiple types of solid cancers including melanoma, hypoxia-inducible factor-1 (HIF-1) composed of α and β subunits is a pivotal transcription factor in the adaptation of cells to low oxygen conditions. HIF-1α protein is tightly regulated by oxygen levels despite the constitutive expression of HIF-1α mRNA. Under normoxic conditions, HIF-1α gets degraded before it can be translocated to the nucleus, associate with HIF-1β, and begin the hypoxic response that is conducive to tumor formation. For this degradation to occur, HIF-1α is hydroxylated by the prolyl 4-hydroxylase (P4H) enzyme into proline 402 and 564 residues, which cause HIF-1α to bind to the von Hippel-Lindau protein (VHL) E3-ubiquitin ligase complex, leading to ubiquitination of this complex and subsequent signaling for proteasomal degradation. Conversely, hypoxic conditions arrest this oxygen-dependent reaction, resulting in the dimerization of α and β subunits in the nucleus. Additionally, oncogenic signaling pathways such as the PI3K (phosphoinositide 3-kinase)-AKT-mTOR (mammalian target of rapamycin )axis and mitogen-activated protein kinase (MAPK) pathway are involved in de novo HIF-1α protein synthesis via 5′-cap-dependent translation initiation [
2,
3], and various transcription factors including signal transducer and activator of transcription 3 (STAT3) and nuclear factor-κB (NF-κB) are critical for regulating HIF-1α mRNA expression with promoter occupancy at the proximal region of
HIF1A [
4,
5,
6,
7]. The HIF-1α and HIF-1β complex binds to the hypoxia-response element (HRE), thereby affecting various downstream genes associated with cancer cell angiogenesis, migration, and metastasis. During cancer metastasis, HIF-1α in hypoxic microenvironments transcriptionally increases various transcripts related to stimulation of cell migration and invasion, including fibronectin 1 (
FN1), urokinase plasminogen activator receptor (
uPAR), lysyl oxidase-like 2 (
LOXL2), and matrix metalloproteinases (
MMPs) [
8].
Vanillin is a major component of vanilla bean extract and is widely used as a flavoring in foods. Because of its antioxidant properties, many biological activities of vanillin have been studied. Vanillin inhibited mutagen induced-DNA damage or spontaneous mutation in bacteria and human cells by eliciting DNA repair [
9,
10,
11]. Recent reports have shown that vanillin has anti-cancer effects through increased apoptosis and cell cycle arrest in melanoma, colon, and cervical cancer cells [
12,
13,
14]. In addition, vanillin has also been reported to exhibit anti-invasive and anti-metastatic activities by suppressing the phosphoinositide 3-kinase (PI3K) and NF-κB signaling pathways in lung, breast, and liver cancer cells [
15,
16,
17]. Nevertheless, the precise molecular mechanism by which vanillin suppresses cancer growth and metastatic potential has not yet been elucidated. Taking into consideration all of the above facts, this study focused on the role of vanillin in the suppression of cancer cell motility and the mechanism of HIF-1α inhibition under hypoxic environments in A2058 and A375 malignant melanoma cells.
In the present study, we evaluated the inhibitory effects of vanillin on hypoxia-inducible factor (HIF)-1α accumulation and cancer cell motility under hypoxia by abrogating STAT3-mediated HIF1A mRNA expression. Our results suggest that vanillin is a potential therapeutic compound that can be used to develop anti-metastatic agents or preventive functional foods for malignant melanoma.
3. Discussion
Because intratumoral hypoxia causes HIF-1α overexpression, genetic alterations of HIF-1α are commonly observed in malignant solid cancers and closely associated with treatment failure and increased mortality; therefore, it is important to identify HIF-1α inhibitors and test their efficacy as anticancer therapeutics [
8]. A growing number of HIF-1α inhibitors derived from natural products, low molecular weight secondary metabolites produced by plants and microbes, have recently been identified as HIF-1α inhibitors [
18]. For example, it has been reported that apigenin (4′,5,7-trihydroxyflavone) and resveratrol (
trans-3,4,5′-trihydroxystilbene) promote HIF-1α protein degradation in a manner that is independent of the microenvironment oxygen levels [
19,
20]. Pleurotin and genistein (4′,5,7-Trihydroxyisoflavone) inhibit the accumulation of HIF-1α protein by suppressing protein synthesis under both normoxic and hypoxic conditions [
21,
22]. Many of the currently identified HIF-1α inhibitors derived from natural products affect protein accumulation or degradation. Interestingly, we propose that vanillin may be a promising HIF-1α inhibitor that acts to reduce HIF-1α levels by suppressing HIF-1α mRNA expression.
Vanillin, a widely used flavoring agent from vanilla, has been shown to exhibit multiple biological effects, including anti-cancer, anti-mutagenic, and anti-bacterial activity in mammalian cells [
9,
14,
23]. In this study, we demonstrate that vanillin effectively decreases HIF-1α protein levels and the expression of its target genes related to cell motility, angiogenesis, and glycolytic metabolism in A2058 and A375 malignant melanoma cells.
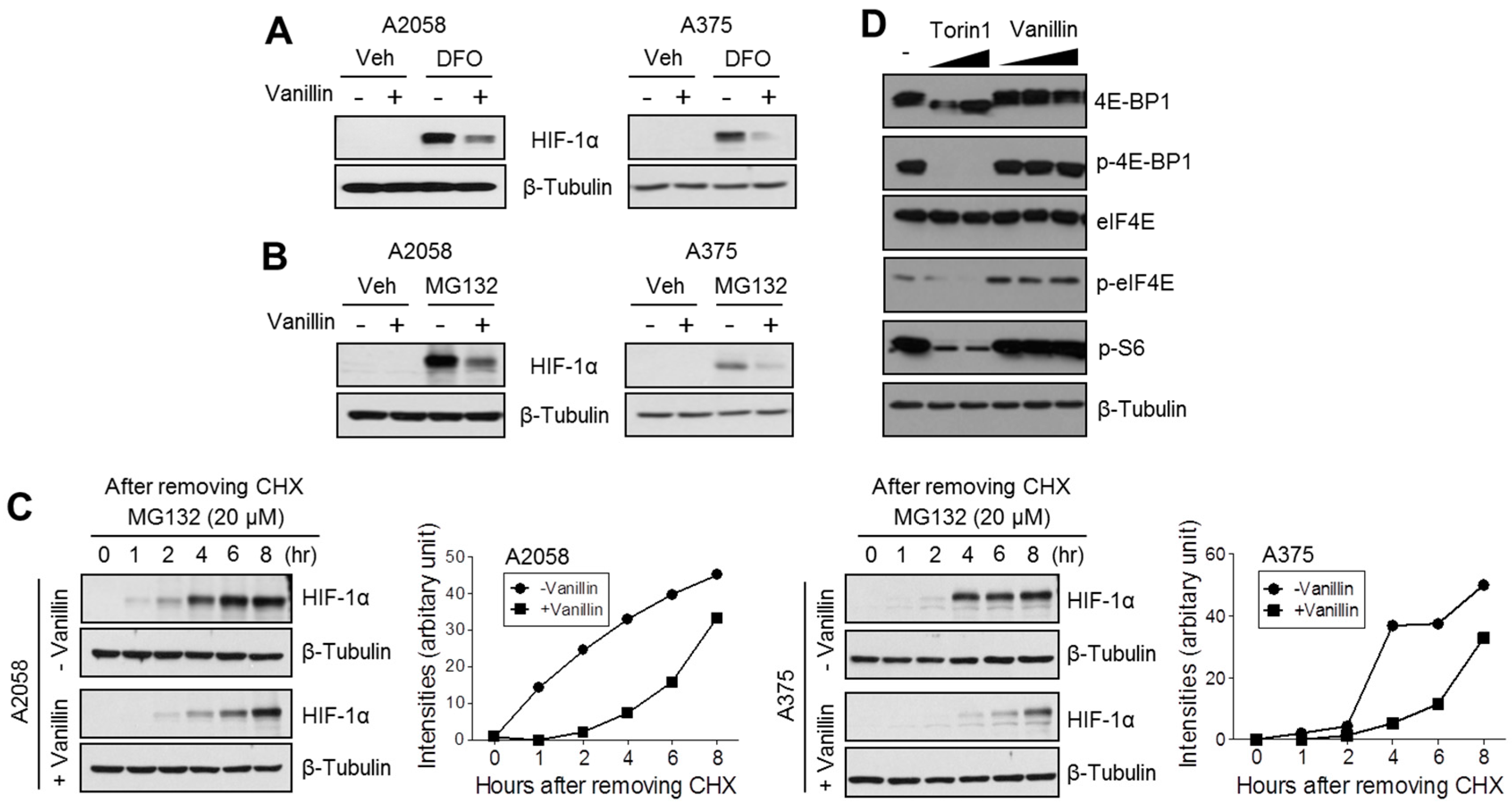
To understand the precise molecular mechanism by which vanillin decreases HIF-1α protein levels, we investigated whether vanillin attenuates HIF-1α protein synthesis or promotes proteasomal degradation. We found that vanillin dramatically attenuates de novo HIF-1α protein synthesis. In addition, MG132, a 26S proteasome inhibitor, did not rescue vanillin-mediated HIF-1α reduction in melanoma cells, suggesting that vanillin reduces HIF-1α using a mechanism that is independent of proteasomal degradation. Because the growth factor-mediated PI3K-mTOR signaling pathway activates HIF-1α protein synthesis via 4E-BP1, eIF4E, and S6 kinase linked 5′-cap-dependent translation initiation [
2,
3], we tested whether vanillin regulates the phosphorylation status of 4E-BP1, eIF4E, and S6 kinase using torin1, a selective mTOR inhibitor, as a positive control. Unlike torin1, vanillin did not alter the phosphorylation status of 4E-BP1, eIF4E, and S6 kinase, suggesting that vanillin does not participate in PI3K-mTOR-mediated protein synthesis. Therefore, we further investigated the inhibitory effect of vanillin on HIF-1α mRNA expression. Interestingly, HIF-1α mRNA was significantly decreased by vanillin treatment in A2058 melanoma cells.
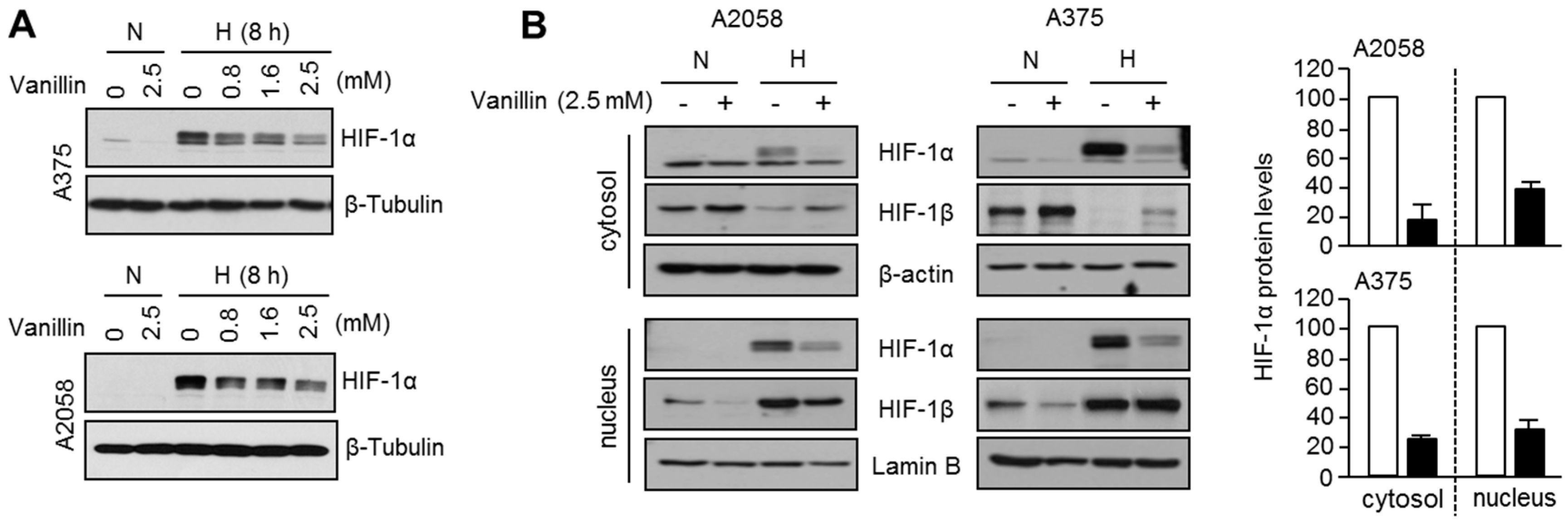
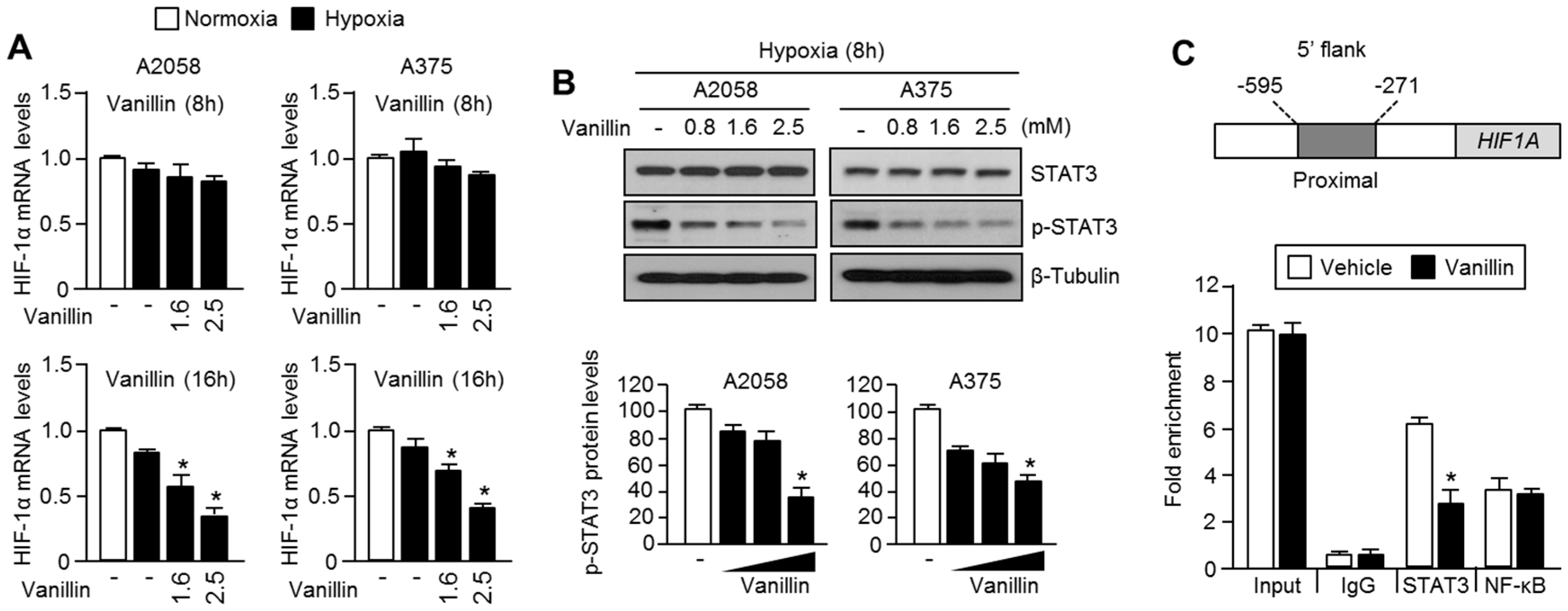
How does vanillin decrease HIF-1α mRNA expression? To answer this question, we investigated the inhibitory effect of vanillin on STAT3-mediated HIF-1α mRNA expression, because STAT3 is one of the transcription factors that is associated with the proximal promoter of
HIF1A [
5,
6]. Vanillin reduces STAT3 phosphorylation and promoter occupancy on the 5′-flank of
HIF1A. These results suggest that vanillin decreases HIF-1α by suppressing STAT3-mediated transcription. Nevertheless, we did not provide a precise molecular mechanism to explain how vanillin inhibits STAT3 phosphorylation and proximal promoter occupancy on the 5′ flanking region of
HIF1A. Therefore, how vanillin suppresses STAT3 phosphorylation and transcriptional activity should be further investigated.
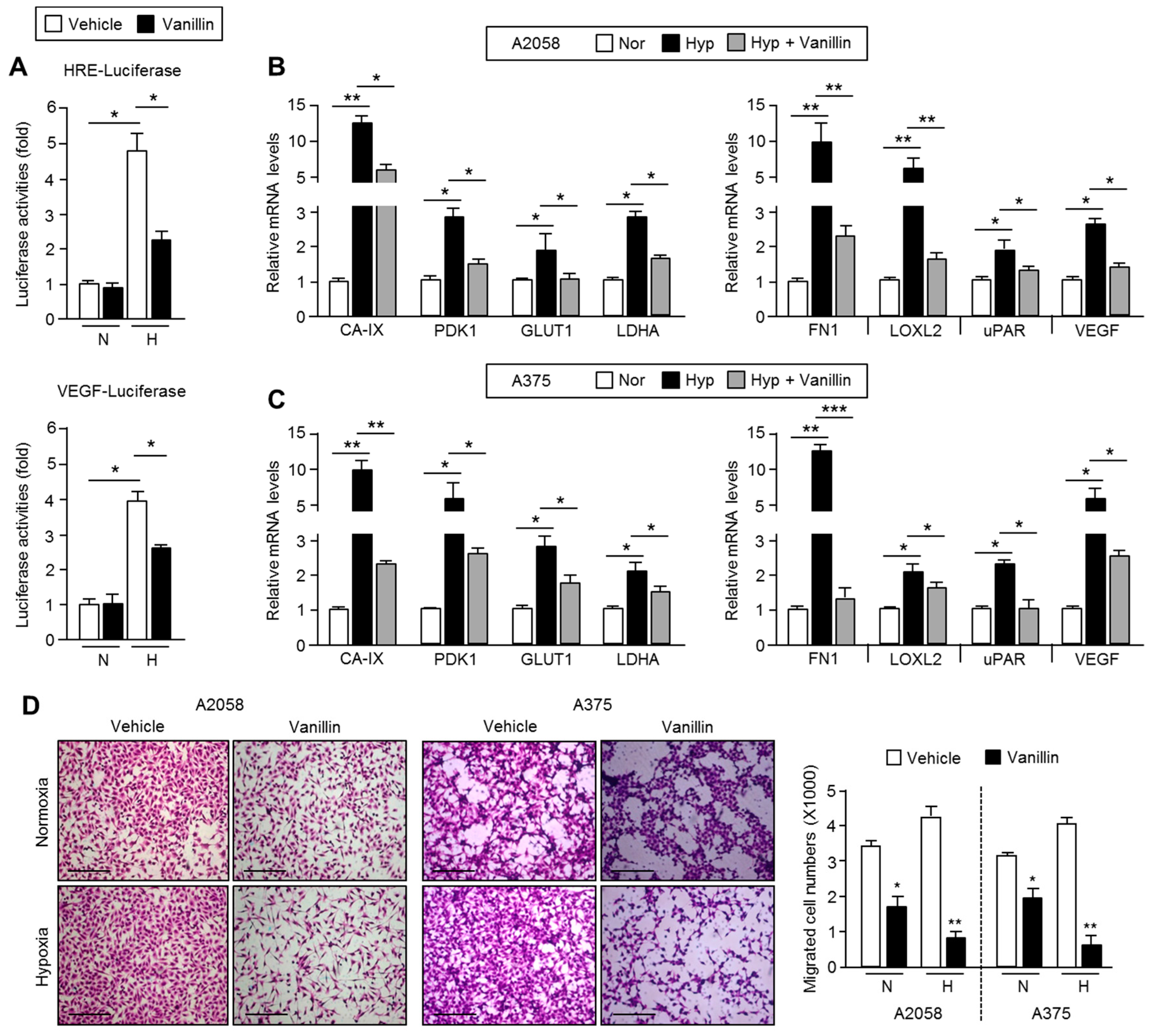
Although the anti-metastatic effect of vanillin by inhibiting MMP-9 expression in breast and hepatocellular carcinoma cells has recently been reported, the molecular mechanism by which vanillin attenuates migration and invasion in cancer cells is not fully demonstrated [
15,
16,
17]. In the present study, we provide insight into some part of the mechanism that involves the STAT3-HIF-1α axis on the vanillin-mediated suppression of cancer cell migration and invasion. Indeed, cell migration was significantly decreased by vanillin by approximately 50% under normoxic condition. Under hypoxia, vanillin suppressed cell migration by approximately 75%, suggesting that vanillin could sensitively block cell motility in malignant tumors with hypoxic microenvironments.
On the basis of previous studies of the anti-metastatic effects of millimolar-range vanillin used to treat cells in vivo, mice were administered 100 mg/kg/day vanillin. Although 100 mg/kg/day can be regarded as a high concentration, no side effects were observed [
17]. Variable concentrations below 100 mg/kg/day can be considered for animal studies. Several methods can be used to determine the effects low-concentration vanillin. Vanillin derivatives can be developed to improve delivery efficacy at low concentrations, which are more effective for preventing malignant melanoma metastasis. Indeed, a recent report showed that 60 mg/kg/day of
o-vanillin, a vanillin isomer, strongly suppressed tumor growth in mice bearing A375 human malignant melanoma xenografts [
14]. For clinical application, it should be further evaluated whether
o-vanillin has anti-metastatic effects via the inhibition of HIF-1α accumulation. In addition, vanillin may be useful as a functional food and not limited to chemotherapy. Our results provide a foundation for further analysis of vanillin for the prevention and treatment of malignant melanoma.
4. Materials and Methods
4.1. Reagents and Antibodies
Vanillin (V1104), deferoxamine (D9533), MG132 (M7449), dimethyl sulfoxide (DMSO), protease inhibitor cocktail, and cycloheximide (01810) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Antibodies against 4E-BP1 (9452), phospho-4E-BP1 (2855), eIF4E (9742), phospho-eIF4E (9741), phospho-S6 (4857), STAT3 (12640), and phospho-STAT3 (9131) were obtained from Cell Signaling Technology (Danvers, MA, USA). Antibodies against β-tubulin (sc-9104) and β-actin (sc-47778) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Anti-HIF-1α and anti-HIF-1β antibodies were kindly provided by Jong-Wan Park of Seoul National University, Seoul, Korea.
4.2. Cell Culture and Treatment
A2058 and A375 melanoma cell lines were purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum (FBS) (Gibco, Carlsbad, CA, USA) and 25 mM glucose in a humidified atmosphere of 5% CO2 at 37 °C. The oxygen level in the hypoxia incubator chamber was maintained at 1% by continuously injecting N2 gas. Vanillin at various stock concentrations (0.8, 1.6, 2.5 M) was dissolved in dimethyl sulfoxide (DMSO) and diluted into culture medium prior to treatment.
4.3. Cell Viability Assay
To determine the cell viability, we used crystal violet staining in this study. Cells were seeded in 24-well plates, and incubated for 24 h, followed by treatment with vanillin in increasing concentrations for 12 and 24 h at 20% (normoxia) or 1% (hypoxia) oxygen conditions. The cells were fixed with 4% paraformaldehyde for 15 min and stained with 0.05% crystal violet staining solution (HT90132, Sigma-Aldrich) for 15 min. To measure optical density, 1% sodium dodecyl sulfate (SDS) solution was added to the cells, and these were further incubated for 15 min at room temperature. The dissolved solutions were transferred to a 96-well plate and measured at 595 nm using a microplate reader (BioTek, Winooski, VT, USA).
4.4. Cytosolic and Nuclear Extract Preparation
Cells were washed using cold phosphate-buffered saline and harvested by centrifugation at 1000 rpm for 5 min at 4 °C. The harvested cell pellets were resuspended and incubated with ice-cold buffer A (20 mM Tris at pH 7.8, 1.5 mM MgCl2, 10 mM KCl, 0.2 mM ethylenediaminetetraacetic acid (EDTA), 0.5 mM dithiothreitol (DTT), and protease inhibitor cocktail) for 5 min on ice, and then cells were collected by centrifugation at 1000 rpm for 5 min at 4 °C. Next, the cell pellets were lysed using 0.06% NP-40 containing buffer A for 10 min on ice. The cell lysates were centrifuged at 3000 rpm for 5 min, and then the supernatants containing cytosolic proteins were frozen. After obtaining the cytosolic fraction, the pellets were incubated and lysed using buffer B (20 mM Tris-Cl at pH 7.8, 1.5 mM MgCl2, 0.2 mM EDTA, 0.5 mM DTT, and 20% glycerol) containing 400 mM NaCl for 30 min on ice. During incubation, the cells were homogenized with a glass homogenizer. The incubated samples were centrifuged at 14,000 rpm for 30 min at 4 °C, and then supernatants containing the nuclear proteins were transferred into fresh tubes.
4.5. Immunoblotting
Total proteins were extracted using cell lysis buffer (1% NP-40, 150 mM NaCl, 50 mM Tris pH 7.4, 2 mM EDTA, and protease inhibitor cocktail). Cell lysates were separated by 7.5% or 10% SDS-polyacrylamide gel electrophoresis (PAGE). Separated proteins were transferred onto an Immobilon-P membrane (Millipore, Billerica, MA, USA). The transferred membranes were blocked with 5% skim milk in Tris-buffered saline containing 0.05% Tween-20 (TBS-T) for 1 h at room temperature, and then incubated overnight with primary antibodies diluted at 1:1000 or 1:5000 in 5% skim milk in TBS-T at 4 °C. Horseradish peroxidase-conjugated secondary antibodies were incubated for 1 h at room temperature, and then protein levels were visualized using an Enhanced Chemiluminescence Prime kit (GE Healthcare, Little Chalfont, UK).
4.6. Quantitative Real-Time PCR
Total RNA was isolated with TRIzol and 2 μg of this RNA was used to synthesize cDNA using a high capacity cDNA reverse transcription kit (Applied Biosystems). The cDNA was amplified over 40 cycles (95 °C for 15 s, 60 °C for 1 min). Experimental Cq values were normalized to H36B4 and relative mRNA levels were calculated on the basis of H36B4 mRNA levels. The sequences of the PCR primers (5′–3′) were: ATGGAGCCCAGCAGCAA and GGCATTGATGACTCCAGTGTT for GLUT1; CCACTCCAGCAGGGAAGG and GCGACGCAGCCTTTGAAT for CA-IX; TGAACATTCTGGCTGGTGACAGGA and ATGATGTCATTCCCACAATGGCCC for PDK1; CTACCTCCACCATGCCAAGT and AGCTGCGCTGATAGACATCC for VEGF; CCATAAAGGGCAACCAAGAG and ACCTCGGTGTTGTAAGGTGG for FN1; CACTGCGGATCCCTGAAAC and CCTGTCTTCGGGCTGATG for LOXL2; AGCCTTACCGAGGTTGTGTG and AAATGCATTCGAGGTAACGG for uPAR.
4.7. In Vitro Migration Assay
In vitro cell migration assays were performed using a Transwell chamber from Sigma-Aldrich (St. Louis, MO, USA). The underside of the Transwell insert membrane was coated with collagen and incubated at room temperature until it was dry. Cells in 0.1 mL of fetal bovine serum-free medium were seeded into the upper chamber, and the lower chamber was filled with 6% fetal bovine serum-containing medium as a chemotactic source, and then cells were incubated for 16 h at 37 °C. After incubation, the Transwell chambers were quickly washed using phosphate-buffered saline and stained with hematoxylin and eosin. The Transwell insert membranes, containing the migrated cells, were placed on a slide glass and analyzed using a microscope (Olympus, Tokyo, Japan). To quantify the migrated cell numbers, two random fields under 40× magnification were quantified by counting the cell numbers.
4.8. Chromatin Immunoprecipitation
Cultured cells were fixed with 1% formaldehyde to cross-link chromatin and proteins, and soluble chromatin and protein complex samples were incubated overnight at 4 °C with antibodies against STAT3 and NF-κB p65. STAT3 or NF-κB p65 interacting DNA was eluted, and then occupancy of STAT3 on the proximal region of the HIF-1α promoter was measured using quantitative PCR. The sequences of PCR primers for the quantitative ChIP assay (5′–3′) are ATCTGAGCAACGAGACCAAA and CACGTGCTCGTCTGTGTTTA.
4.9. Luciferase Activity Assay
Hypoxia-responsive element (HRE) or VEGF (vascular endothelial growth factor) promoter-luciferase reporter plasmids were a gift from Navdeep Chandel (Addgene plasmid # 26731) and Jong-Wan Park (Seoul National University, Korea). Cells were transfected with reporter plasmids using Polyfect (QIAGEN, Valencia, CA, USA) and then incubated for 48 h for stabilization. After incubation, luciferase activity was measured using a luminometer (Berthold Technologies, Bad Wildbad, Germany) and normalized against β-gal activities to account for transfection efficiency.
4.10. Statistical Analysis
All data were analyzed using an unpaired Student’s t-test for two experimental comparisons and a one-way ANOVA (analysis of variance) followed by Tukey’s post hoc test for multiple comparisons using GraphPad Prism 5.01 (GraphPad Software Inc., La Jolla, CA, USA). Data are represented as means ± standard deviations (SDs). Differences between mean values were considered statistically significant when the associated p-value was less than 0.05.


 and
and 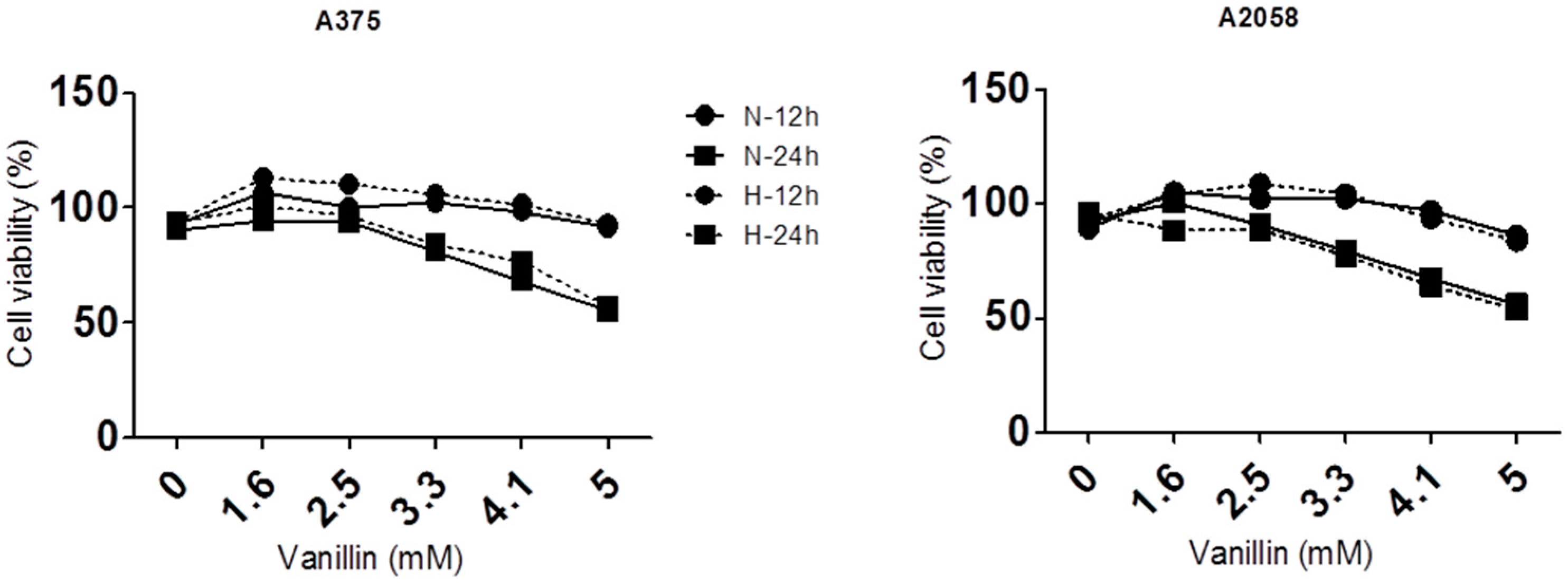{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




