
Heavy Metal Induced Antibiotic Resistance in Bacterium LSJC7

Abstract
:1. Introduction
2. Results and Discussion
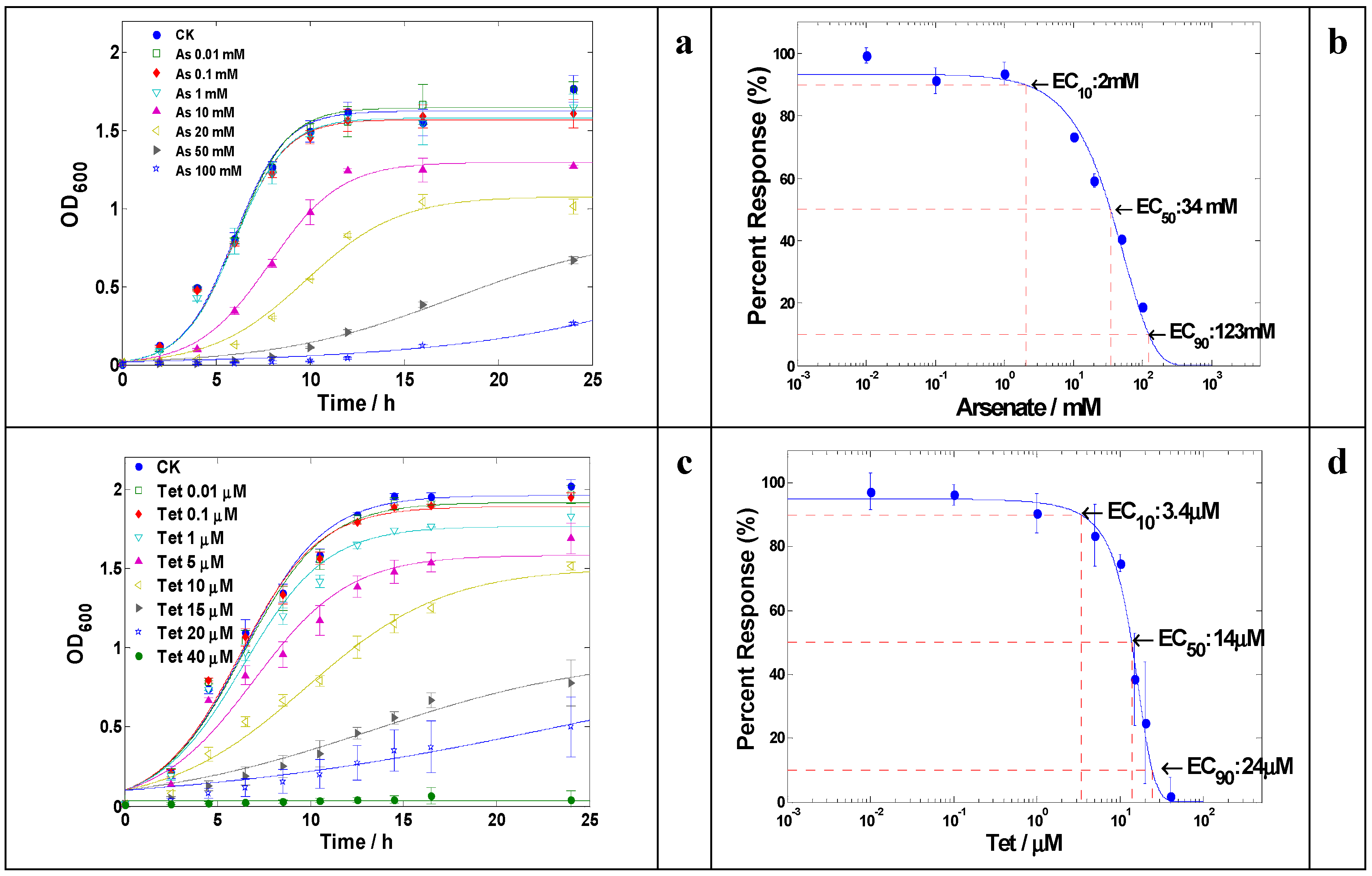
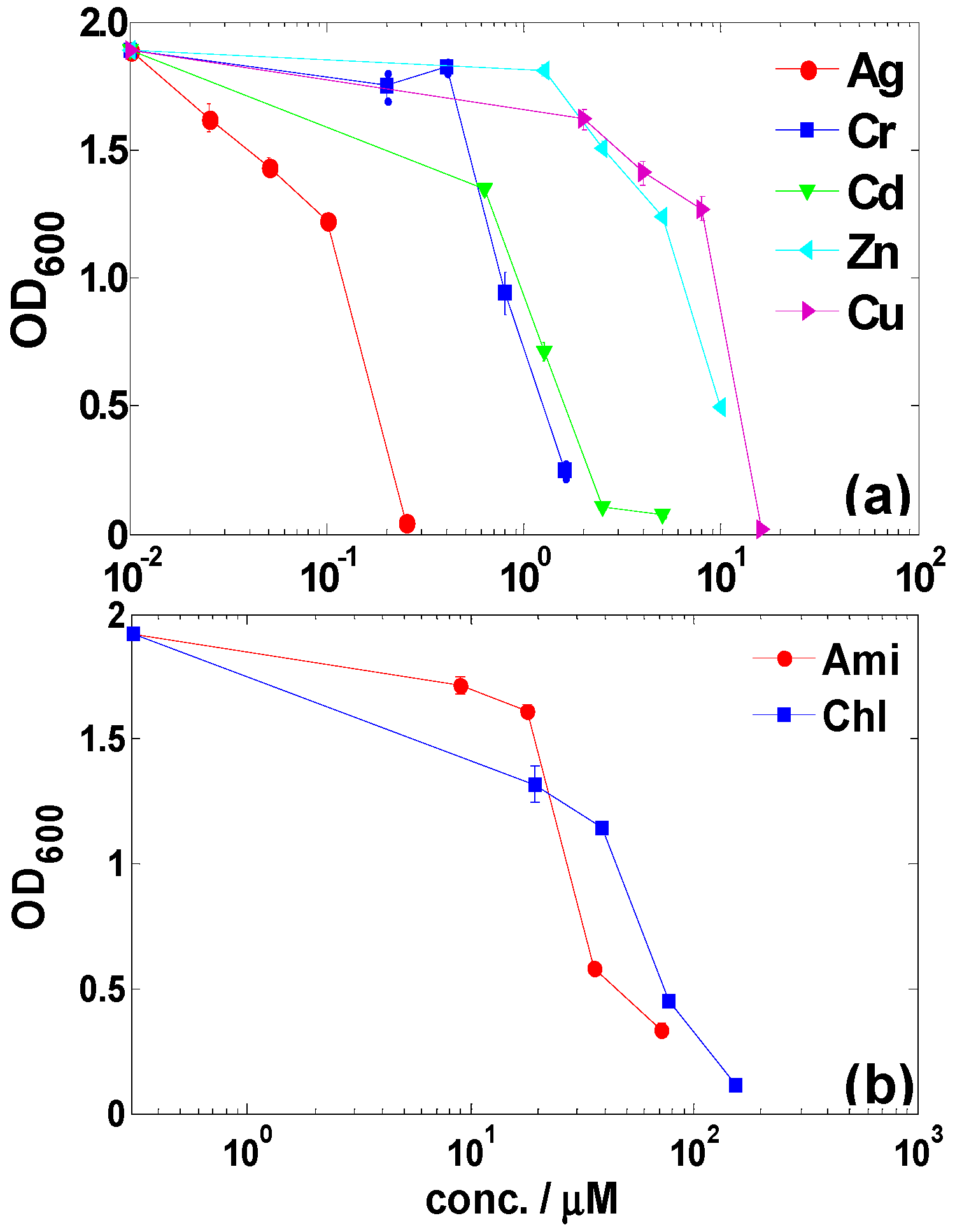
2.1. Metal(loid) Resistance of LSJC7 and Identification of Putative Resistance Genes


2.2. Antibiotic Resistance of LSJC7 and Identification of Putative Resistance Genes
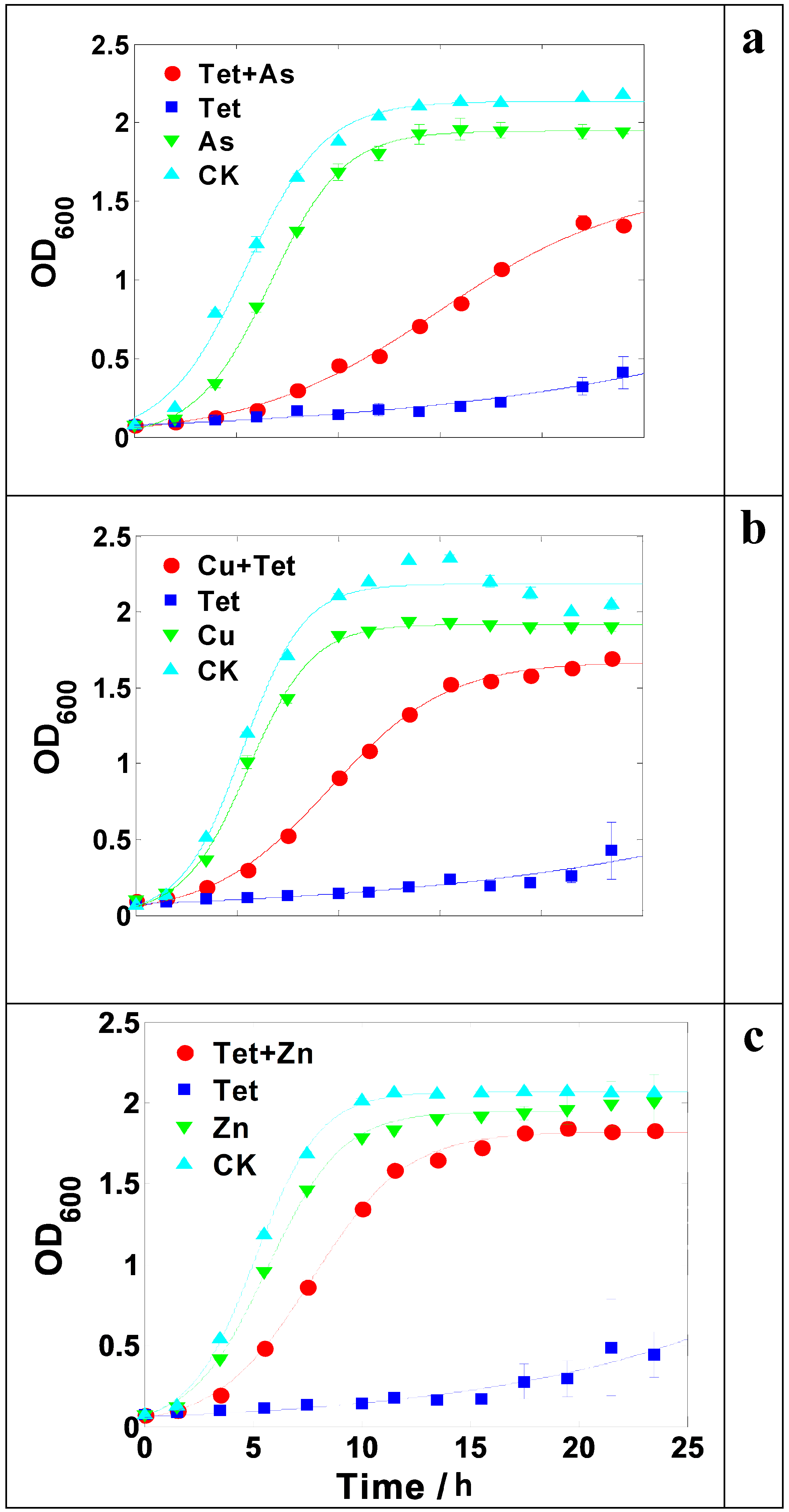
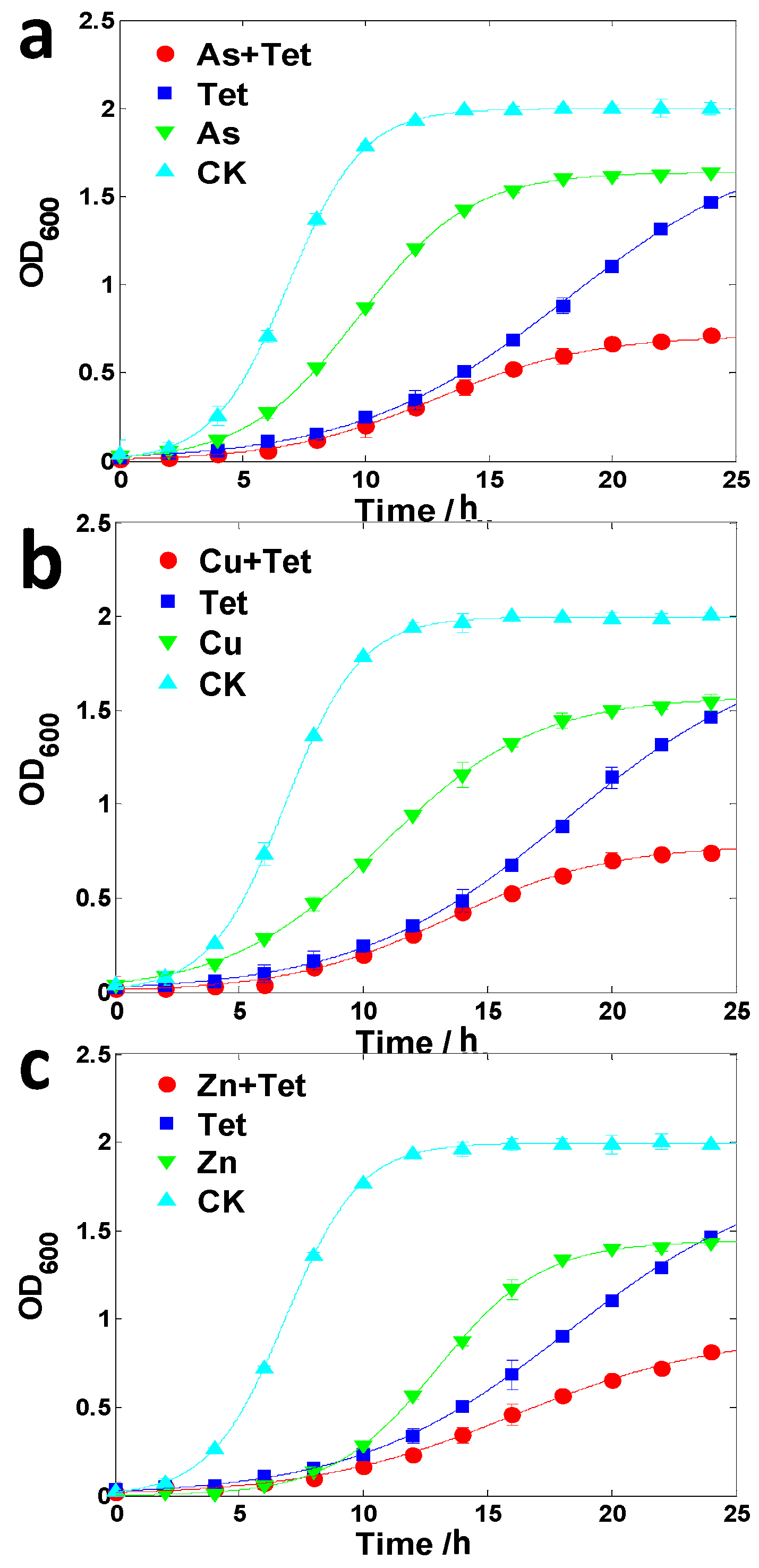
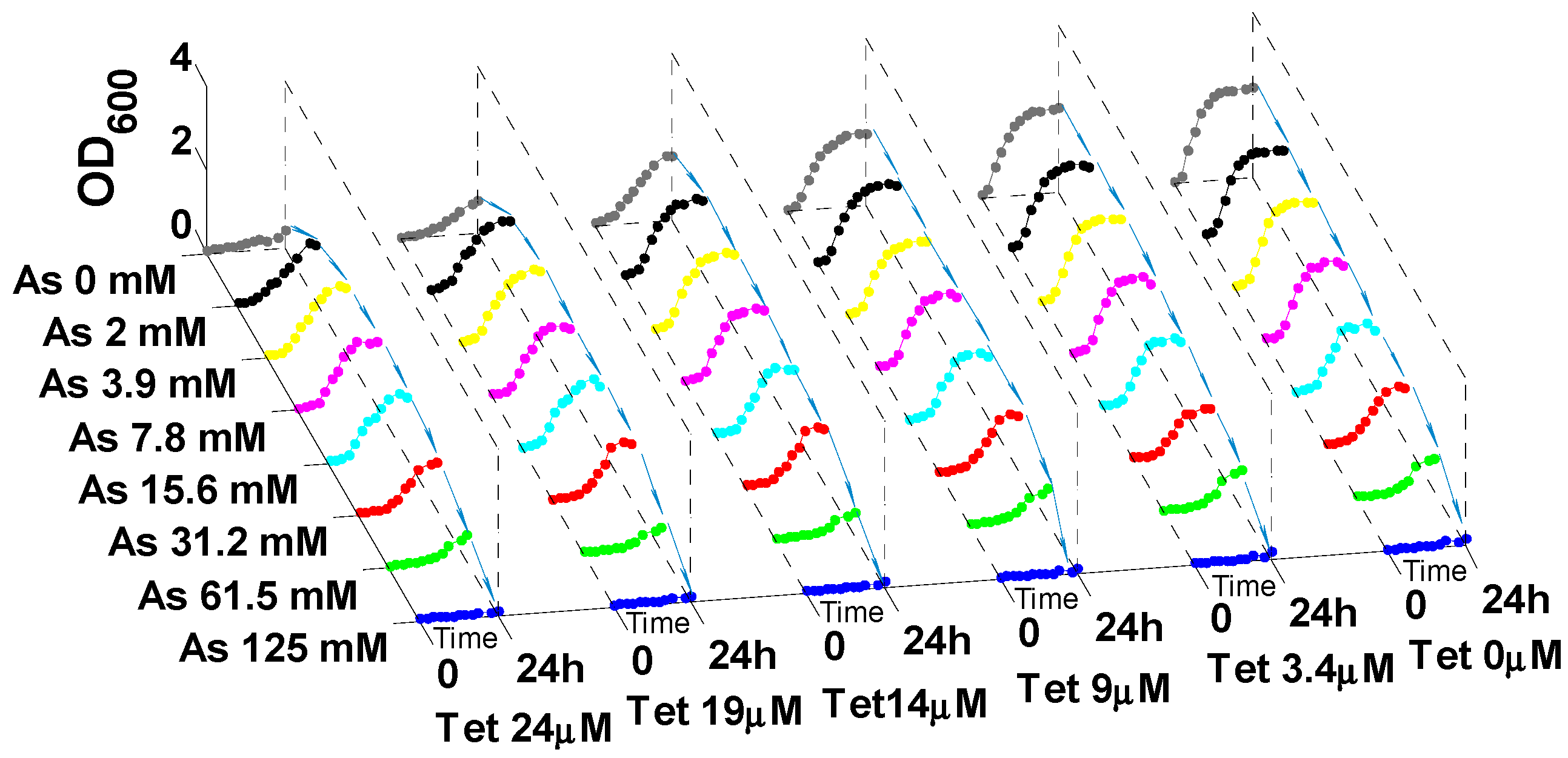
2.3. Enhancement of Antibiotic Resistance in the Presence of Heavy Metals




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | LSJC7 | E. coli DH5α | ||||
|---|---|---|---|---|---|---|
| Combination | Tet | Amp | Chl | Tet | Amp | Chl |
| As | * | / | * | / | / | / |
| Cu | * | / | / | * | / | / |
| Zn | * | / | / | * | / | / |
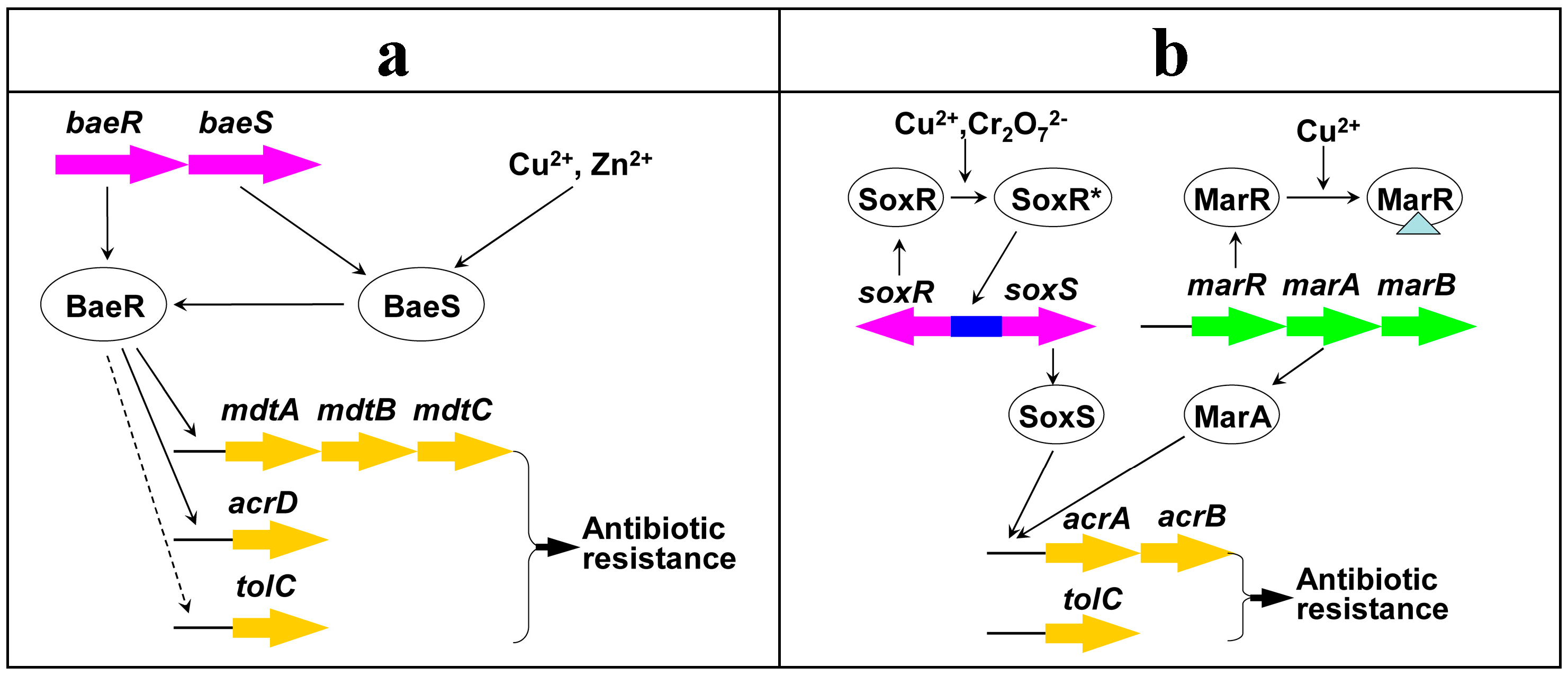
2.4. Possible Mechanisms of Heavy Metal Induced Antibiotic Resistance

3. Experimental Section
3.1. Bacterial Strains and Growth Medium
3.2. Dose-Response Inhibition of Growth for Heavy Metals
3.3. Dose–Response Inhibition of Growth for Antibiotics
3.4. Growth Kinetics of Bacteria
3.5. Metal and Antibiotic Analysis
3.6. Data Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Martinez, J.L.; Rojo, F. Metabolic regulation of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 768–789. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Johnson, T.A.; Su, J.Q.; Qiao, M.; Guo, G.X.; Stedtfeld, R.D.; Hashsham, S.A.; Tiedje, J.M. Diverse and abundant antibiotic resistance genes in Chinese swine farms. Proc. Natl. Acad. Sci. USA 2013, 110, 3435–3440. [Google Scholar] [CrossRef] [PubMed]
- Looft, T.; Johnson, T.A.; Allen, H.K.; Bayles, D.O.; Alt, D.P.; Stedtfeld, R.D.; Sul, W.J.; Stedtfeld, T.M.; Chai, B.L.; Cole, J.R.; et al. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. USA 2012, 109, 1691–1696. [Google Scholar] [CrossRef] [PubMed]
- Hvistendahl, M. China takes aim at rampant antibiotic resistance. Science 2012, 336, 795–795. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Qiang, Z.M.; Ben, W.W.; Chen, M.X. Residual veterinary antibiotics in swine manure from concentrated animal feeding operations in Shandong Province, China. Chemosphere 2011, 84, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.L.; Zhao, G.L.; Tian, J.; Su, X.O. High-performance liquid chromatography inductively coupled plasma mass spectrometry based method for the determination of organic arsenic feed additives and speciation of anionic arsenics in animal feed. J. Agric. Food Chem. 2010, 58, 5263–5270. [Google Scholar] [CrossRef] [PubMed]
- Binh, C.T.T.; Heuer, H.; Kaupenjohann, M.; Smalla, K. Piggery manure used for soil fertilization is a reservoir for transferable antibiotic resistance plasmids. FEMS Microbiol. Ecol. 2008, 66, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Gao, Y.; Luo, J.; Yan, S.H.; Rengel, Z.; Zhang, Z.H. Interaction of veterinary antibiotic tetracyclines and copper on their fates in water and water hyacinth (Eichhornia crassipes). J. Hazard. Mater. 2014, 280, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Hernando, M.D.; Mezcua, M.; Fernandez-Alba, A.R.; Barcelo, D. Environmental risk assessment of pharmaceutical residues in wastewater effluents, surface waters and sediments. Talanta 2006, 69, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Skurnik, D.; Ruimy, R.; Ready, D.; Ruppe, E.; Bernede-Bauduin, C.; Djossou, F.; Guillemot, D.; Pier, G.B.; Andremont, A. Is exposure to mercury a driving force for the carriage of antibiotic resistance genes? J. Med. Microbiol. 2010, 59, 804–807. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.D.; Zhu, Y.G.; Fu, B.J.; Marschner, P.; He, J.Z. The veterinary antibiotic oxytetracycline and Cu influence functional diversity of the soil microbial community. Environ. Pollut. 2006, 143, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Peltier, E.; Vincent, J.; Finn, C.; Graham, D.W. Zinc-induced antibiotic resistance in activated sludge bioreactors. Water Res. 2010, 44, 3829–3836. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.; Tom-Petersen, A.; Nybroe, O. Copper amendment of agricultural soil selects for bacterial antibiotic resistance in the field. Lett. Appl. Microbiol. 2005, 40, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Stepanauskas, R.; Glenn, T.C.; Jagoe, C.H.; Tuckfield, R.C.; Lindell, A.H.; King, C.J.; McArthur, J.V. Coselection for microbial resistance to metals and antibiotics in freshwater microcosms. Environ. Microbiol. 2006, 8, 1510–1514. [Google Scholar] [CrossRef] [PubMed]
- Knapp, C.W.; McCluskey, S.M.; Singh, B.K.; Campbell, C.D.; Hudson, G.; Graham, D.W. Antibiotic resistance gene abundances correlate with metal and geochemical conditions in archived Scottish soils. PLoS ONE 2011, 6, e27300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.Q.; Ye, J.; Zhu, Y.G. Draft genome sequence of a novel bacterial strain, LSJC7, belonging to the family Enterobacteriaceae with dual resistance to arsenic and tetracycline. J. Bacteriol. 2012, 194, 7005–7006. [Google Scholar] [CrossRef] [PubMed]
- Cuebas, M.; Sannino, D.; Bini, E. Isolation and characterization of an arsenic resistant Geobacillus kaustophilus strain from geothermal soils. J. Basic Microb. 2011, 51, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.F.; Yang, J.B.; Rosen, B.P. ArsD: An As(III) metallochaperone for the ArsAB As(III)-translocating ATPase. J. Bioenerg. Biomembr. 2007, 39, 453–458. [Google Scholar] [CrossRef]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Editorial: Assessing the antimicrobial susceptibility of bacteria obtained from animals. J. Antimicrob. Chemother. 2010, 65, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Hill, G.M.; Cromwell, G.L.; Crenshaw, T.D.; Dove, C.R.; Ewan, R.C.; Knabe, D.A.; Lewis, A.J.; Libal, G.W.; Mahan, D.C.; Shurson, G.C.; et al. Growth promotion effects and plasma changes from feeding high dietary concentrations of zinc and copper to weanling pigs (regional study). J. Anim. Sci. 2000, 78, 1010–1016. [Google Scholar] [PubMed]
- Katouli, M.; Melin, L.; Jensen-Waern, M.; Wallgren, P.; Mollby, R. The effect of zinc oxide supplementation on the stability of the intestinal flora with special reference to composition of coliforms in weaned pigs. J. Appl. Microbiol. 1999, 87, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.P.; Zhang, W.F.; Hu, Y.N.; Hu, E.D.; Xie, X.D.; Wang, L.L.; Cheng, H.F. Arsenic pollution of agricultural soils by concentrated China animal feeding operations (CAFOs). Chemosphere 2015, 119, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Granados-Chinchilla, F.; Rodriguez, C. Bioavailability of in-feed tetracyclines is influenced to a greater extent by crude protein rather than calcium. Anim. Feed Sci. Technol. 2014, 198, 323–332. [Google Scholar] [CrossRef]
- Webber, M.A.; Piddock, L.J.V. The importance of efflux pumps in bacterial antibiotic resistance. J. Antimicrob. Chemother. 2003, 51, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Qiao, M.; Chen, W.D.; Su, J.Q.; Zhang, B.; Zhang, C. Fate of tetracyclines in swine manure of three selected swine farms in China. J. Environ. Sci. 2012, 24, 1047–1052. [Google Scholar] [CrossRef]
- Alcock, R.E.; Sweetman, A.; Jones, K.C. Assessment of organic contaminant fate in waste water treatment plants I: Selected compounds and physicochemical properties. Chemosphere 1999, 38, 2247–2262. [Google Scholar] [CrossRef]
- Ji, X.L.; Shen, Q.H.; Liu, F.; Ma, J.; Xu, G.; Wang, Y.L.; Wu, M.H. Antibiotic resistance gene abundances associated with antibiotics and heavy metals in animal manures and agricultural soils adjacent to feedlots in Shanghai; China. J. Hazard. Mater. 2012, 235, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Garbarino, J.R.; Bednar, A.J.; Rutherford, D.W.; Beyer, R.S.; Wershaw, R.L. Environmental fate of roxarsone in poultry litter. I. Degradation of roxarsone during composting. Environ. Sci. Technol. 2003, 37, 1509–1514. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; LaPara, T.M. The effects of subtherapeutic antibiotic use in farm animals on the proliferation and persistence of antibiotic resistance among soil bacteria. ISME J. 2007, 1, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Pruden, A.; Arabi, M.; Storteboom, H.N. Correlation between upstream human activities and riverine antibiotic resistance genes. Environ. Sci. Technol. 2012, 46, 11541–11549. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.G.; Yang, L.S. A review of heavy metal contaminations in urban soils, urban road dusts and agricultural soils from China. Microchem. J. 2010, 94, 99–107. [Google Scholar] [CrossRef]
- Finley, R.L.; Collignon, P.; Larsson, D.G.J.; McEwen, S.A.; Li, X.Z.; Gaze, W.H.; Reid-Smith, R.; Timinouni, M.; Graham, D.W.; Topp, E. The scourge of antibiotic resistance: The important role of the environment. Clin. Infect. Dis. 2013, 57, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Nies, D.H. Microbial heavy-metal resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef] [PubMed]
- Gullberg, E.; Albrecht, L.M.; Karlsson, C.; Sandegren, L.; Andersson, D.I. Selection of a multidrug resistance plasmid by sublethal levels of antibiotics and heavy metals. MBio 2014, 5, e01918-14. [Google Scholar] [CrossRef]
- Fang, Y.; Sun, X.Y.; Yang, W.J.; Ma, N.; Xin, Z.H.; Fu, J.; Liu, X.C.; Liu, M.; Mariga, A.M.; Zhu, X.F.; et al. Concentrations and health risks of lead, cadmium, arsenic, and mercury in rice and edible mushrooms in China. Food Chem. 2014, 147, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.H.; Lingas, E.O.; Rahman, M. Contamination of drinking-water by arsenic in Bangladesh: A public health emergency. Bull. World Health Organ. 2000, 78, 1093–1103. [Google Scholar] [PubMed]
- Harris, E.S.J.; Cao, S.G.; Littlefield, B.A.; Craycroft, J.A.; Scholten, R.; Kaptchuk, T.; Fu, Y.L.; Wang, W.Q.; Liu, Y.; Chen, H.B.A.; et al. Heavy metal and pesticide content in commonly prescribed individual raw Chinese Herbal Medicines. Sci. Total Environ. 2011, 409, 4297–4305. [Google Scholar] [CrossRef] [PubMed]
- Mallik, S.; Virdi, J.S.; Johri, A.K. Proteomic analysis of arsenite—Mediated multiple antibiotic resistance in Yersinia enterocolitica biovar 1A. J. Basic Microb. 2012, 52, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Caille, O.; Rossier, C.; Perron, K. A copper-activated two-component system interacts with zinc and imipenem resistance in Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 4561–4568. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.; Mellado, R.P.; Martinez, J.L. Metal accumulation and vanadium-induced multidrug resistance by environmental isolates of Escherichia hermannii and Enterobacter cloacae. Appl. Environ. Microbiol. 1998, 64, 4317–4320. [Google Scholar] [PubMed]
- Nishino, K.; Nikaido, E.; Yamaguchi, A. Regulation of multidrug efflux systems involved in multidrug and metal resistance of Salmonella enterica serovar typhimurium. J. Bacteriol. 2007, 189, 9066–9075. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.J.; Tremaroli, V.; Stan, M.A.; Chan, C.S.; Vacchi-Suzzi, C.; Heyne, B.J.; Parsek, M.R.; Ceri, H.; Turner, R.J. Chromosomal antioxidant genes have metal ion-specific roles as determinants of bacterial metal tolerance. Environ. Microbiol. 2009, 11, 2491–2509. [Google Scholar] [PubMed]
- Okusu, H.; Ma, D.; Nikaido, H. AcrAB efflux pump plays a major role in the antibiotic resistance phenotype of Escherichia coli multiple-antibiotic-resistance (Mar) mutants. J. Bacteriol. 1996, 178, 306–308. [Google Scholar] [PubMed]
- Hao, Z.Y.; Lou, H.B.; Zhu, R.F.; Zhu, J.H.; Zhang, D.M.; Zhao, B.X.S.; Zeng, S.Z.; Chen, X.; Chan, J.; He, C.; et al. The multiple antibiotic resistance regulator MarR is a copper sensor in Escherichia coli. Nat. Chem. Biol. 2014, 10, U21–U48. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, Y.B.; Xu, J.X.; Zhang, X.H.; Xu, S.H.; Du, Q.P. Combined toxic effects of heavy metals and antibiotics on a Pseudomonas fluorescens strain ZY2 isolated from swine wastewater. Int. J. Mol. Sci. 2015, 16, 2839–2850. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Sun, K.; Gao, B.; Zhang, G.X.; Liu, X.T.; Zhao, Y. Adsorption of tetracycline on soil and sediment: Effects of pH and the presence of Cu(II). J. Harzad. Mater. 2011, 190, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Tong, F.; Zhao, Y.P.; Gu, X.Y.; Gu, C.; Lee, C.C.C. Joint toxicity of tetracycline with copper(II) and cadmium(II) to Vibrio fischeri: Effect of complexation reaction. Ecotoxicology 2015, 24, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Morones-Ramirez, J.R.; Winkler, J.A.; Spina, C.S.; Collins, J.J. Silver enhances antibiotic activity against gram-negative bacteria. Sci. Transl. Med. 2013, 5, 190ra81. [Google Scholar] [CrossRef] [PubMed]
- Vaun McArthur, J.; Tuckfield, R.C.; Baker-Austin, C. Antimicrobial Textiles. In Handbook of Experimental Pharmacology; Anthony, R.M.C., Ed.; Springer: Berlin, Germany; Heidelberg, Germany, 2012; Volume 211, pp. 135–152. [Google Scholar]
- Arias, C.A.; Murray, B.E. Antibiotic-resistant bugs in the 21st century—A clinical super-challenge. N. Engl. J. Med. 2009, 360, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.D.; Lin, L.F.; Luo, Z.X.; Yan, C.Z.; Zhang, X. Occurrence of selected antibiotics in Jiulongjiang River in various seasons, South China. J. Environ. Monit. 2011, 13, 1953–1960. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Li, X.; Sun, G.; Zhang, Y.; Su, J.; Ye, J. Heavy Metal Induced Antibiotic Resistance in Bacterium LSJC7. Int. J. Mol. Sci. 2015, 16, 23390-23404. https://doi.org/10.3390/ijms161023390
Chen S, Li X, Sun G, Zhang Y, Su J, Ye J. Heavy Metal Induced Antibiotic Resistance in Bacterium LSJC7. International Journal of Molecular Sciences. 2015; 16(10):23390-23404. https://doi.org/10.3390/ijms161023390
Chicago/Turabian StyleChen, Songcan, Xiaomin Li, Guoxin Sun, Yingjiao Zhang, Jianqiang Su, and Jun Ye. 2015. "Heavy Metal Induced Antibiotic Resistance in Bacterium LSJC7" International Journal of Molecular Sciences 16, no. 10: 23390-23404. https://doi.org/10.3390/ijms161023390