
Falsochrobactrum tianjinense sp. nov., a New Petroleum-Degrading Bacteria Isolated from Oily Soils
Abstract
:1. Introduction
2. Materials and Methodologies
2.1. Chemicals and Media
- KH2PO4 1.00 g L−1,
- K2HPO4 1.00 g L−1,
- NaCl 10.00 g L−1,
- (NH4)2SO4 1.50 g L−1,
- Anhydrous CaCl2 0.10 g L−1,
- FeSO4·7H2O 0.01 g L−1; and,
- MgSO4 0.20 g L−1.
2.2. Enrichment and Isolation of Oil-Bearing Soils Degrading Microorganisms
2.3. Identification and Genome Features of Strains
2.4. Physiological Characterization of the Isolated Strain
2.5. TPH Biodegradation Ability of Falsochrobactrum sp. TDYN1
- M0 is the weight of petroleum initially added into the medium; and,
- M is the weight of residual petroleum after extraction.
3. Result and Discussion
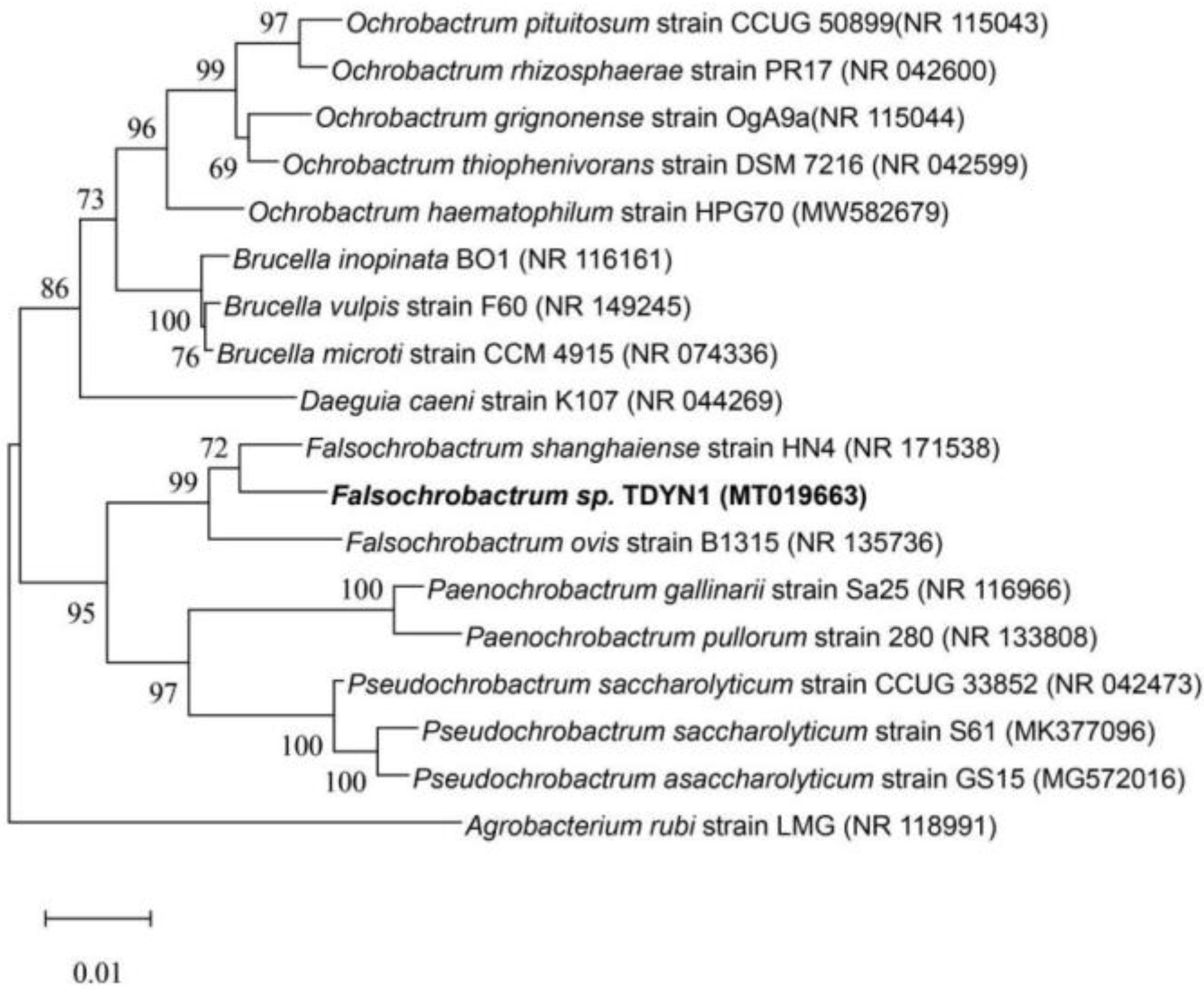
3.1. Isolation and Identification of the Falsochrobactrum TDYN1
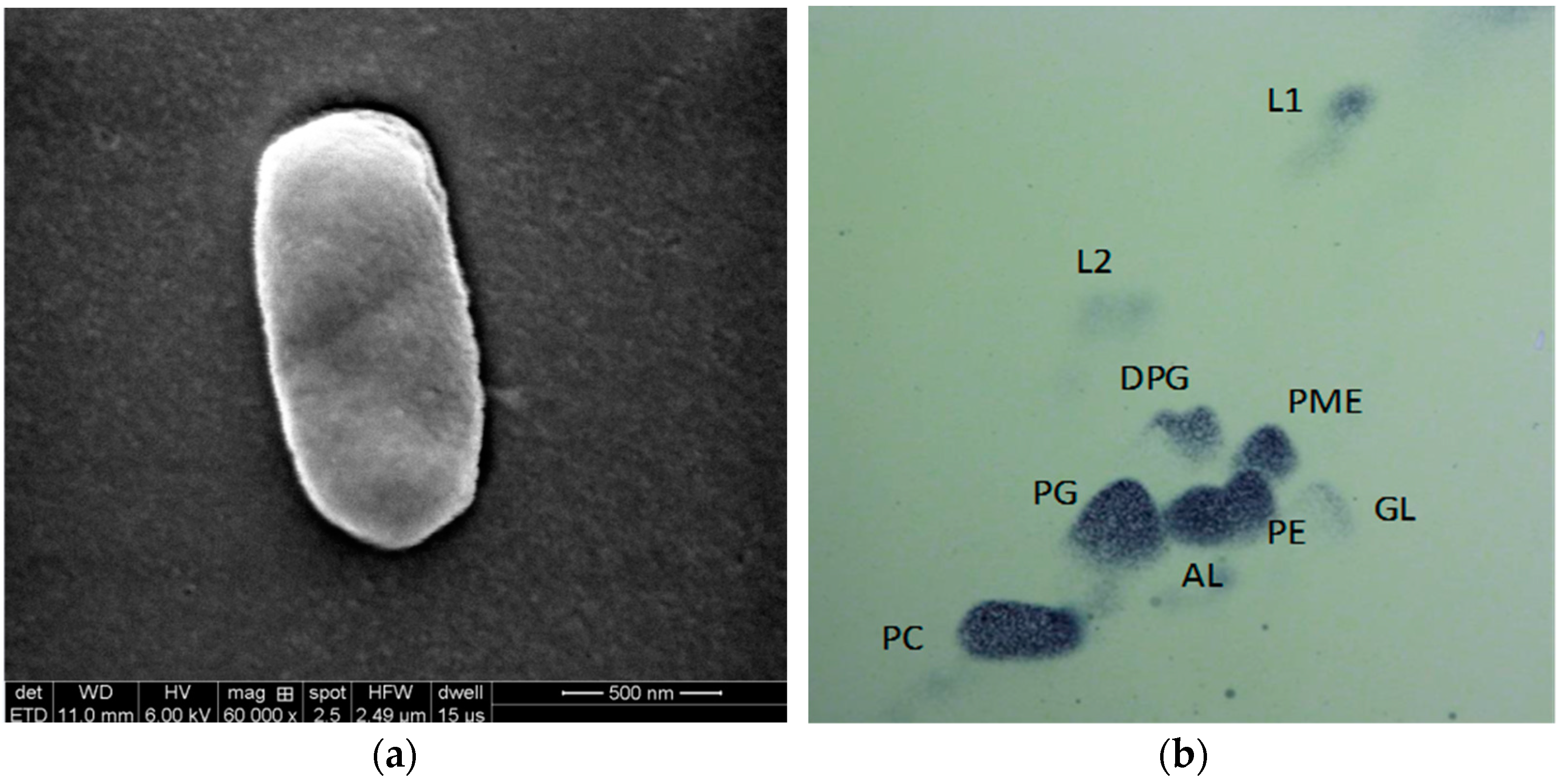
3.2. Physiological Characterization of the Isolated Strain
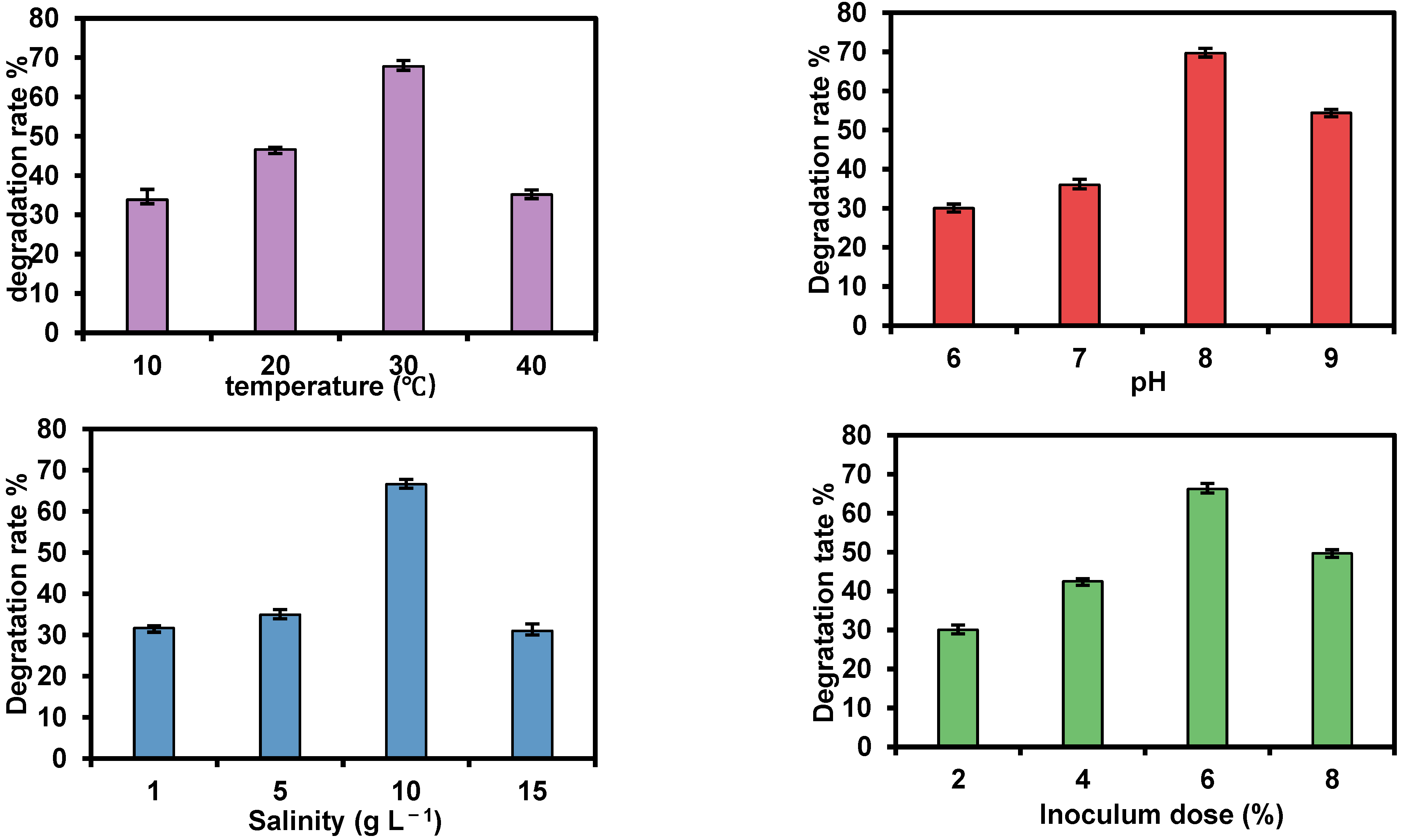
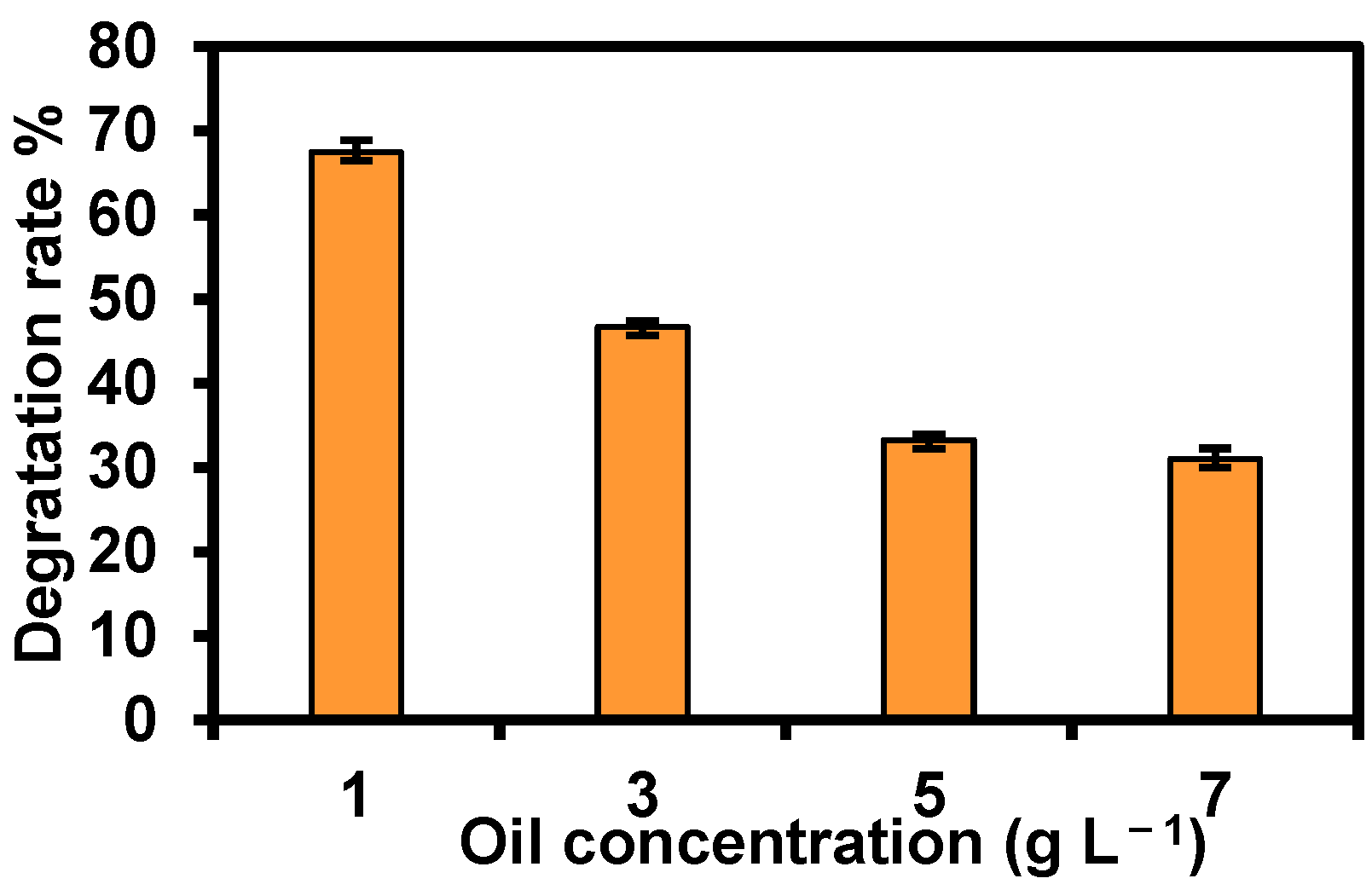
3.3. TPH Biodegradation Ability and Optimal Degradation Conditions of Strain TDYN1
- temperature = 30 °C,
- initial pH = 8,
- salinity = 10 g L−1,
- petroleum concentration = 1 g L−1; and,
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Saadoun, I. Isolation and characterization of bacteria from crude petroleum oil contaminated soil and their potential to degrade diesel fuel. J. Basic Microbiol. Int. J. Biochem. Physiol. Genet. Morphol. Ecol. Microorg. 2002, 42, 420–428. [Google Scholar] [CrossRef]
- Chandra, S.; Sharma, R.; Singh, K.; Sharma, A. Application of bioremediation technology in the environment contaminated with petroleum hydrocarbon. Ann. Microbiol. 2013, 63, 417–431. [Google Scholar] [CrossRef]
- Ron, E.Z.; Rosenberg, E. Enhanced bioremediation of oil spills in the sea. Curr Opin Biotechnol 2014, 27, 191–194. [Google Scholar] [CrossRef]
- Al-Hawash, A.B.; Dragh, M.A.; Li, S.; Alhujaily, A.; Abbood, H.A.; Zhang, X.; Ma, F. Principles of microbial degradation of petroleum hydrocarbons in the environment. Egypt. J. Aquat. Res. 2018, 44, 71–76. [Google Scholar] [CrossRef]
- Luo, Y.R.; Tian, Y.; Huang, X.; Kwon, K.; Yang, S.H.; Seo, H.S.; Kim, S.J.; Zheng, T.L. Sphingomonas polyaromaticivorans sp. nov., a polycyclic aromatic hydrocarbon-degrading bacterium from an oil port water sample. Int. J. Syst. Evol. Microbiol. 2012, 62 Pt 6, 1223–1227. [Google Scholar] [CrossRef]
- Ma, J.; Xu, L.; Jia, L. Degradation of polycyclic aromatic hydrocarbons by Pseudomonas sp. JM2 isolated from active sewage sludge of chemical plant. J. Environ. Sci. 2012, 24, 2141–2148. [Google Scholar] [CrossRef]
- Liu, B.; Ju, M.; Liu, J.; Wu, W.; Li, X. Isolation, identification, and crude oil degradation characteristics of a high-temperature, hydrocarbon-degrading strain. Mar. Pollut. Bull. 2016, 106, 301–307. [Google Scholar] [CrossRef]
- Das, K.; Mukherjee, A.K. Crude petroleum-oil biodegradation efficiency of Bacillus subtilis and Pseudomonas aeruginosa strains isolated from a petroleum-oil contaminated soil from North-East India. Bioresour. Technol. 2007, 98, 1339–1345. [Google Scholar] [CrossRef]
- Baumann, P.; Doudoroff, M.; Stanier, R. A study of the Moraxella group II. Oxidative-negative species (genus Acinetobacter). J. Bacteriol. 1968, 95, 1520–1541. [Google Scholar] [CrossRef]
- Essien, J.; Ebong, G.; Asuquo, J.; Olajire, A. Hydrocarbons contamination and microbial degradation in mangrove sediments of the Niger Delta region (Nigeria). Chem. Ecol. 2012, 28, 421–434. [Google Scholar] [CrossRef]
- Oyetibo, G.O.; Ilori, M.O.; Obayori, O.S.; Amund, O.O. Biodegradation of petroleum hydrocarbons in the presence of nickel and cobalt. J. Basic Microbiol. 2013, 53, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Wang, Y.; Lin, T.; Liu, L.; Bin, M.; Du, H. Isolation and characterization of a hydrocarbon-degrading strain of Compostibacillus. Microbiol. China 2020, 47, 691–698. [Google Scholar]
- Panda, S.; Kar, R.; Panda, C. Isolation and identification of petroleum hydrocarbon degrading microorganisms from oil contaminated environment. Int. J. Environ. Sci. 2013, 3, 1314. [Google Scholar]
- Zhou, L.; Li, H.; Zhang, Y.; Han, S.; Xu, H. Sphingomonas from petroleum-contaminated soils in Shenfu, China and their PAHs degradation abilities. Braz. J. Microbiol. 2016, 47, 271–278. [Google Scholar] [CrossRef]
- Liu, Q.Y.; Li, L.; Zhao, D.F.; Zhao, C.C. Screening of a Thermophilic Hydrocarbon-Degrading Bacterial Consortium and Community Structure Analysis. Adv. Mater. Res. 2012, 610-613, 225–228. [Google Scholar] [CrossRef]
- Kshirsagar, S.D.; Mattam, A.J.; Jose, S.; Ramachandrarao, B.; Velankar, H.R. Heavy hydrocarbons as selective substrates for isolation of asphaltene degraders: A substrate-based bacterial isolation strategy for petroleum hydrocarbon biodegradation. Environ. Technol. Innov. 2020, 19, 100832. [Google Scholar] [CrossRef]
- Liu, Q.; Tang, J.; Bai, Z.; Hecker, M.; Giesy, J.P. Distribution of petroleum degrading genes and factor analysis of petroleum contaminated soil from the Dagang Oilfield, China. Sci. Rep. 2015, 5, 11068. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-Q.; Xiao-Jing, L.; Khan, M.A.; Gul, B. Relationship between soil characteristics and halophytic vegetation in coastal region of North China. Pak. J. Bot. 2008, 40, 1081–1090. [Google Scholar]
- Mishra, S.; Jyot, J.; Kuhad, R.C.; Lal, B. In situ bioremediation potential of an oily sludge-degrading bacterial consortium. Curr. Microbiol. 2001, 43, 328–335. [Google Scholar] [CrossRef]
- Déziel, É.; Paquette, G.; Villemur, R.; Lépine, F.; Bisaillon, J. Biosurfactant production by a soil Pseudomonas strain growing on polycyclic aromatic hydrocarbons. Appl. Environ. Microbiol. 1996, 62, 1908–1912. [Google Scholar] [CrossRef]
- Cai, Y.; Wang, R.; Rao, P.; Wu, B.; Yan, L.; Hu, L.; Park, S.; Ryu, M.; Zhou, X. Bioremediation of Petroleum Hydrocarbons Using Acinetobacter sp. SCYY-5 Isolated from Contaminated Oil Sludge: Strategy and Effectiveness Study. Int. J. Environ. Res. Public Health 2021, 18, 819. [Google Scholar] [CrossRef] [PubMed]
- Onur, G.; Yilmaz, F.; Icgen, B. Diesel Oil Degradation Potential of a Bacterium Inhabiting Petroleum Hydrocarbon Contaminated Surface Waters and Characterization of Its Emulsification Ability. J. Surfactants Deterg. 2015, 18, 707–717. [Google Scholar] [CrossRef]
- Bernardet, J.F.; Nakagawa, Y.; Holmes, B.; Subcommittee on the Taxonomy of Flavobacterium; Cytophaga-like Bacteria of the International Committee on Systematics of Prokaryotes. Proposed minimal standards for describing new taxa of the family Flavobacteriaceae and emended description of the family. Int. J. Syst. Evol. Microbiol. 2002, 52 Pt 3, 1049–1070. [Google Scholar] [PubMed]
- Claus, D. A standardized Gram staining procedure. World J. Microbiol. Biotechnol. 1992, 8, 451–452. [Google Scholar] [CrossRef]
- Ohta, H.; Hattori, T. Agromonas oligotrophica gen. nov., sp. nov., a nitrogen-fixing oligotrophic bacterium. Antonie Van Leeuwenhoek 1983, 49, 429–446. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, R.; Xiao, Y.; Liu, C.; Fu, H. Isolation and identification of a chlorimuron-ethyl-degrading bacterium and optimization of its degradation conditions. Chin. J. Biotechnol. 2020, 36, 560–568. [Google Scholar]
- Sun, L.; Yao, L.; Gao, X.; Huang, K.; Bai, N.; Lyu, W.; Chen, W. Falsochrobactrum shanghaiense sp. nov., isolated from paddy soil and emended description of the genus Falsochrobactrum. Int. J. Syst. Evol. Microbiol. 2019, 69, 778–782. [Google Scholar] [CrossRef]
- Kampfer, P.; Glaeser, S.; Busse, H.J.; Eisenberg, T.; Scholz, H. Falsochrobactrum ovis gen. nov., sp. nov., isolated from a sheep. Int. J. Syst. Evol. Microbiol. 2013, 63 Pt 10, 3841–3847. [Google Scholar] [CrossRef]
- Isaac, P.; Martínez, F.L.; Bourguignon, N.; Sánchez, L.A.; Ferrero, M.A. Improved PAHs removal performance by a defined bacterial consortium of indigenous Pseudomonas and actinobacteria from Patagonia, Argentina. Int. Biodeterior. Biodegrad. 2015, 101, 23–31. [Google Scholar] [CrossRef]
- Ortega-González, D.K.; Cristiani-Urbina, E.; Flores-Ortíz, C.M.; Cruz-Maya, J.A.; Cancino-Díaz, J.C.; Jan-Roblero, J. Evaluation of the Removal of Pyrene and Fluoranthene by Ochrobactrum anthropi, Fusarium sp. and Their Coculture. Appl. Biochem. Biotechnol. 2015, 175, 1123–1138. [Google Scholar] [CrossRef]
- Pugazhendi, A.; Qari, H.; Basahi, J.M.A.-B.; Godon, J.J.; Dhavamani, J. Role of a halothermophilic bacterial consortium for the biodegradation of PAHs and the treatment of petroleum wastewater at extreme conditions. Int. Biodeterior. Biodegrad. 2017, 121, 44–54. [Google Scholar] [CrossRef]
- Imran, A.; Hafeez, F.Y.; Frühling, A.; Schumann, P.; Malik, K.; Stackebrandt, E. Ochrobactrum ciceri sp. nov., isolated from nodules of Cicer arietinum. Int. J. Syst. Evol. Microbiol. 2010, 60, 1548–1553. [Google Scholar] [CrossRef] [PubMed]
- Abou-Shanab, R.A.; Eraky, M.; Haddad, A.M.; Abdel-Gaffar, A.-R.B.; Salem, A.M. Characterization of crude oil degrading bacteria isolated from contaminated soils surrounding gas stations. Bull. Environ. Contam. Toxicol. 2016, 97, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Gao, W.; Li, Q.; Cui, Z.; Zhang, K.; Zheng, L. Screen and analysis of degradation ability for crude oil degradation strains from South China Sea. Acta Sci. Circumstantiae 2012, 32, 1572–1578. [Google Scholar]
- Qin, X.; Tang, J.C.; Li, D.S.; Zhang, Q.M. Effect of salinity on the bioremediation of petroleum hydrocarbons in a saline-alkaline soil. Lett. Appl. Microbiol. 2012, 55, 210–217. [Google Scholar] [CrossRef]
- Marsh, W.S.; Heise, B.W.; Krzmarzick, M.J.; Murdoch, R.W.; Fathepure, B.Z. Isolation and characterization of a halophilic Modicisalibacter sp. strain Wilcox from produced water. Sci. Rep. 2021, 11, 6943. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Mi, S.; Peng, X.; Han, Y. The mutual influence between corrosion and the surrounding soil microbial communities of buried petroleum pipelines. RSC Adv. 2019, 9, 18930–18940. [Google Scholar] [CrossRef]
- Cui, H.; Yang, X.; Xu, L.; Fan, Y.; Yi, Q.; Li, R.; Zhou, J. Effects of goethite on the fractions of Cu, Cd, Pb, P and soil enzyme activity with hydroxyapatite in heavy metal-contaminated soil. RSC Adv. 2017, 7, 45869–45877. [Google Scholar] [CrossRef]
- Xu, J.L.; Zhang, J.; Huang, T.L.; Xi, A.L. Comparative Bioremediation of Oil Contaminated Soil by Natural Attenuation, Biostimulation and Bioaugmentation, Advanced Materials Research; Trans Tech Publ: Zurich, Switzerland, 2013; pp. 258–262. [Google Scholar]
- Zhang, B.; Zhang, L.; Zhang, X. Bioremediation of petroleum hydrocarbon-contaminated soil by petroleum-degrading bacteria immobilized on biochar. RSC Adv. 2019, 9, 35304–35311. [Google Scholar] [CrossRef]
- Saum, L.; Jimenez, M.B.; Crowley, D. Influence of biochar and compost on phytoremediation of oil-contaminated soil. Int. J. Phytoremediation 2018, 20, 54–60. [Google Scholar] [CrossRef]
- McCarty, N.S. Ledesma-Amaro, R. Synthetic biology tools to engineer microbial communities for biotechnology. Trends Biotechnol. 2019, 37, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Levels | Factor | ||||
|---|---|---|---|---|---|
| (A) Temperature | (B) pH | (C) Salinity | (D) Inoculum Dose | (E) Petroleum Concentration | |
| 1 | 10 °C | 6 | 1 g L−1 | 2% | 1 g L−1 |
| 2 | 20 °C | 7 | 5 g L−1 | 4% | 3 g L−1 |
| 3 | 30 °C | 8 | 10 g L−1 | 6% | 5 g L−1 |
| 4 | 40 °C | 9 | 15 g L−1 | 8% | 7 g L−1 |
| Number | A | B | C | D | E |
|---|---|---|---|---|---|
| 1 | 1 | 1 | 1 | 1 | 1 |
| 2 | 1 | 2 | 2 | 2 | 2 |
| 3 | 1 | 3 | 3 | 3 | 3 |
| 4 | 1 | 4 | 4 | 4 | 4 |
| 5 | 2 | 1 | 2 | 3 | 4 |
| 6 | 2 | 2 | 1 | 4 | 3 |
| 7 | 2 | 3 | 4 | 2 | 2 |
| 8 | 2 | 4 | 3 | 1 | 1 |
| 9 | 3 | 1 | 3 | 4 | 2 |
| 10 | 3 | 2 | 1 | 3 | 1 |
| 11 | 3 | 3 | 4 | 2 | 4 |
| 12 | 3 | 4 | 2 | 1 | 3 |
| 13 | 4 | 1 | 4 | 2 | 3 |
| 14 | 4 | 2 | 3 | 1 | 4 |
| 15 | 4 | 3 | 2 | 4 | 1 |
| 16 | 4 | 4 | 1 | 3 | 2 |
| Characteristic | 1 | 2 | 3 | |
|---|---|---|---|---|
| Gram stain | − | − | − | |
| Shape | rod | rod | Short−rod | |
| Motile | − | − | + | |
| Spore | + | − | − | |
| Capsule | + | ND | ND | |
| Glucose | Acid production | + | ND | ND |
| Aerogenesis | − | ND | ND | |
| Lactose | Acid production | + | ND | ND |
| Aerogenesis | − | ND | ND | |
| Methyl red | − | ND | ND | |
| Methyl acetyl alcohol | − | ND | ND | |
| Benzpyrole | + | ND | ND | |
| Oxidase | + | + | + | |
| Catalase | + | ND | ND | |
| Production of ammonia | + | ND | ND | |
| DNA G + C content (mol%) | 56.0% | 49.1% | 56.9% | |
| pH | 7.0–7.4 | ND | 7.0 | |
| Temperature | 30 ± 1 °C | 30 °C | 30–35 °C | |
| NaCl | 1% | ND | 1% |
| Fatty Acid | 1 | 2 | 3 |
|---|---|---|---|
| C8.0 | 3.6 | ND | ND |
| C10.0 | 3.6 | ND | ND |
| C12.0 | 20.1 | ND | ND |
| C13.0 | 5.6 | ND | ND |
| C14.0 | 318.2 | ND | ND |
| C15.0 | 179.3 | ND | ND |
| C15.1 | 8.5 | 12 | ND |
| C16.0 | 10,733.9 | ND | 8.6 |
| C16.1 | 1896.5 | ND | ND |
| C17.0 | 1195.3 | ND | ND |
| C17.1 | 99.7 | 9.6 | ND |
| C18.0 | 7282.6 | ND | 13.5 |
| C18.1N9C | 16,543.4 | ND | ND |
| C18.1N9T | 4.5 | ND | ND |
| C 18:1 2-OH | ND | ND | 3.2 |
| C18.2N6C | 48.1 | ND | ND |
| C18:1ω7c | ND | 32.1 | ND |
| C19:0cycloω8c | ND | 44.4 | 30.5 |
| C20.0 | 14.6 | ND | ND |
| C20.1 | 65.4 | ND | ND |
| C20:2ω6.9c | ND | 1.9 | ND |
| C20.4N6 | 4.1 | ND | ND |
| C22.0 | 7.5 | ND | ND |
| C22.1N9 | 349.2 | ND | ND |
| C22.2 | 8.9 | ND | ND |
| Polar Lipids | 1 | 2 | 3 |
|---|---|---|---|
| Diphosphatidylglycerol | + | + | + |
| Phosphatidylglycerol | + | + | + |
| Phosphatidylethanolamine | + | + | + |
| Phosphatidylmonomthylethanolamine | + | + | ND |
| Phosphatidylcholine | + | + | + |
| Unidentified aminolipid AL | + | ND | ND |
| Unidentified aminolipid AL1 | ND | + | ND |
| Unidentified glycolipid GL | + | ND | ND |
| Unidentified glycolipid APL1, 2 | ND | ND | + |
| Unidentified phospholipid PL7 | ND | + | ND |
| Unidentified phospholipid GL1, 2 | ND | ND | + |
| Unidentified polar lipids L1-2 | + | ND | ND |
| Unidentified lipid | ND | ND | + |
| Unidentified aminophospholipid L1, 2, 3 | ND | ND | + |
| Number | A | B | C | D | E | Average Degradation Rate (%) |
|---|---|---|---|---|---|---|
| 1 | 1 | 1 | 1 | 1 | 1 | 6.72 ± 0.24 |
| 2 | 1 | 2 | 2 | 2 | 2 | 29.36 ± 1.21 |
| 3 | 1 | 3 | 3 | 3 | 3 | 59.34 ± 0.51 |
| 4 | 1 | 4 | 4 | 4 | 4 | 39.96 ± 1.54 |
| 5 | 2 | 1 | 2 | 3 | 4 | 28.67 ± 0.23 |
| 6 | 2 | 2 | 1 | 4 | 3 | 23.29 ± 1.18 |
| 7 | 2 | 3 | 4 | 2 | 2 | 42.84 ± 0.56 |
| 8 | 2 | 4 | 3 | 1 | 1 | 61.71 ± 0.93 |
| 9 | 3 | 1 | 3 | 4 | 2 | 43.01 ± 1.66 |
| 10 | 3 | 2 | 1 | 3 | 1 | 67.95 ± 0.71 |
| 11 | 3 | 3 | 4 | 2 | 4 | 33.03 ± 1.40 |
| 12 | 3 | 4 | 2 | 1 | 3 | 29.38 ± 0.51 |
| 13 | 4 | 1 | 4 | 2 | 3 | 34.73 ± 0.82 |
| 14 | 4 | 2 | 3 | 1 | 4 | 22.35 ± 0.44 |
| 15 | 4 | 3 | 2 | 4 | 1 | 52.34 ± 0.50 |
| 16 | 4 | 4 | 1 | 3 | 2 | 27.72 ± 1.00 |
| K1 | 135.38 | 113.13 | 125.68 | 120.16 | 188.72 | |
| K2 | 146.51 | 142.95 | 139.75 | 129.96 | 132.93 | |
| K3 | 173.37 | 177.55 | 186.41 | 183.68 | 146.74 | |
| K4 | 137.14 | 158.77 | 140.56 | 158.6 | 124.01 | |
| R | 9.06 | 16.11 | 11.46 | 15.88 | 16.18 | |
| optimal conditions | A3 | B3 | C3 | D3 | E1 |
| Source | Sum of Square | df | Mean Square | F | p | Significant |
|---|---|---|---|---|---|---|
| A | 871.239 | 3 | 290.413 | 160.423 | 0.000 | ** |
| B | 1679.998 | 3 | 559.999 | 309.341 | 0.000 | ** |
| C | 5259.636 | 3 | 1753.212 | 968.466 | 0.000 | ** |
| D | 4868.000 | 3 | 1622.667 | 896.353 | 0.000 | ** |
| E | 5410.477 | 3 | 1803.492 | 996.241 | 0.000 | ** |
| error | 57.930 | 32 | 1.810 | |||
| total | 79,793.420 | 48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, M.; Zhang, F.; Li, G.; Ruan, H.; Li, X.; Zhong, L.; Chen, G.; Rui, Y. Falsochrobactrum tianjinense sp. nov., a New Petroleum-Degrading Bacteria Isolated from Oily Soils. Int. J. Environ. Res. Public Health 2022, 19, 11833. https://doi.org/10.3390/ijerph191811833
Hu M, Zhang F, Li G, Ruan H, Li X, Zhong L, Chen G, Rui Y. Falsochrobactrum tianjinense sp. nov., a New Petroleum-Degrading Bacteria Isolated from Oily Soils. International Journal of Environmental Research and Public Health. 2022; 19(18):11833. https://doi.org/10.3390/ijerph191811833
Chicago/Turabian StyleHu, Mengjie, Feifan Zhang, Gaoyuan Li, Haihua Ruan, Xinhao Li, Lei Zhong, Guanyi Chen, and Yichao Rui. 2022. "Falsochrobactrum tianjinense sp. nov., a New Petroleum-Degrading Bacteria Isolated from Oily Soils" International Journal of Environmental Research and Public Health 19, no. 18: 11833. https://doi.org/10.3390/ijerph191811833