Modeling R0 for Pathogens with Environmental Transmission: Animal Movements, Pathogen Populations, and Local Infectious Zones

 , , ,
, , ,
Abstract
:1. Introduction
1.1. A Brief History of R0
1.2. Challenges for R0 for Environmentally Maintained Pathogens
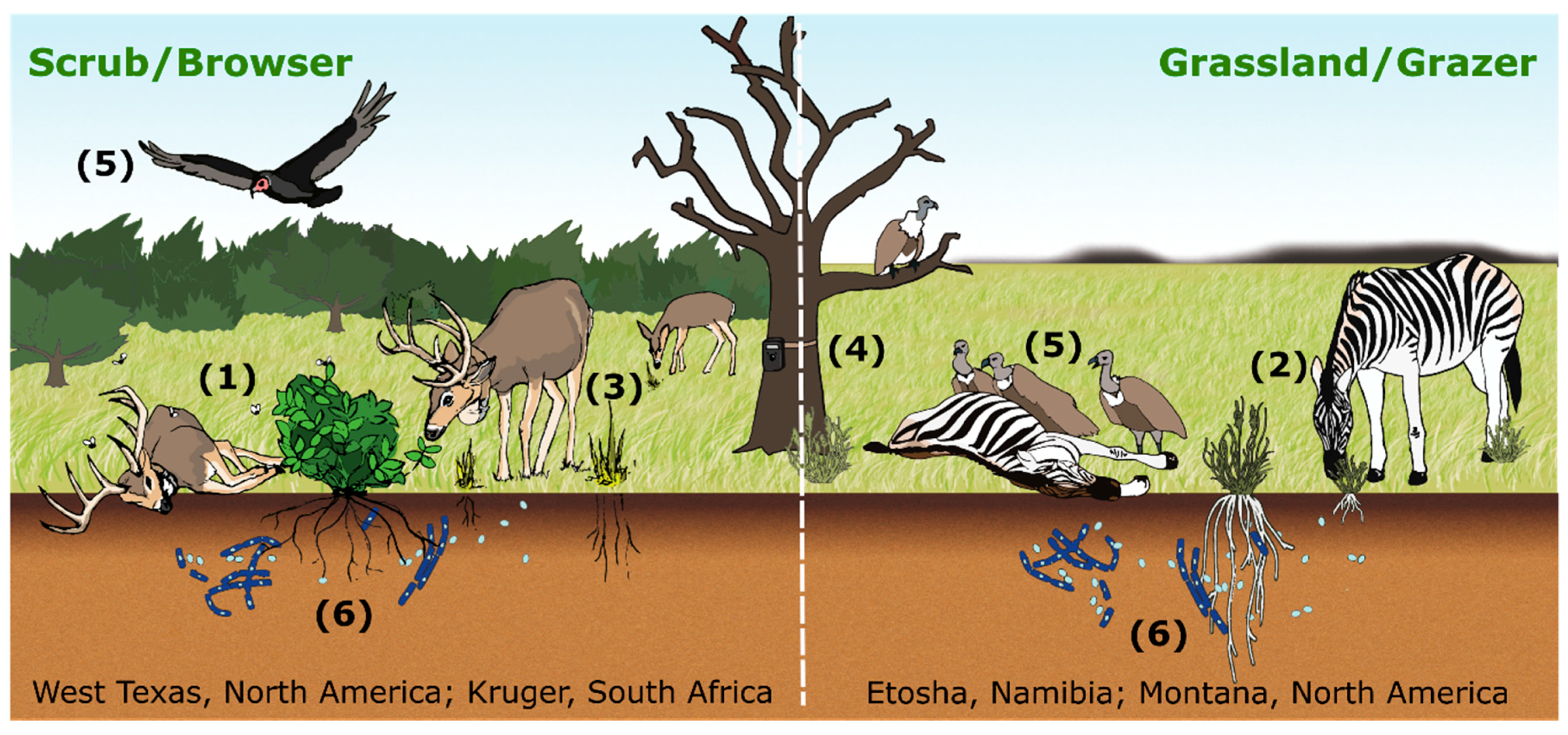
2. Host Interaction with Local Infectious Zones (LIZs) at the Landscape Level
2.1. Local Infectious Zone (LIZ) Ecology
2.2. Host Movement Ecology and Transmission
2.3. Dynamic Thresholds and the Joint Modeling of Reservoir (LIZ) and Host Dynamics
3. Discrete, Self-Decomposable (DSD) Parameter Estimation
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dixon, M.G.; Schafer, I.J. Ebola viral disease outbreak—West Africa, 2014. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 548–551. [Google Scholar]
- Bowman, C.; Gumel, A.; Van den Driessche, P.; Wu, J.; Zhu, H. A mathematical model for assessing control strategies against West Nile virus. Bull. Math. Biol. 2005, 67, 1107–1133. [Google Scholar] [CrossRef] [PubMed]
- Heesterbeek, J. A brief history of R0 and a recipe for its calculation. Acta Biotheor. 2002, 50, 189–204. [Google Scholar] [CrossRef] [PubMed]
- Dublin, L.I.; Lotka, A.J. On the true rate of natural increase: As exemplified by the population of the United States, 1920. J. Am. Stat. Assoc. 1925, 20, 305–339. [Google Scholar] [CrossRef]
- Dietz, K. Transmission and Control of Arbovirus Diseases. In Proceedings of the SIMS Conference on Epidemiology, Alta, UT, USA, 8–12 July 1974; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 1975; pp. 104–121. [Google Scholar]
- Diekmann, O.; Heesterbeek, J.; Metz, J.A. On the definition and the computation of the basic reproduction ratio R0 in models for infectious diseases in heterogeneous populations. J. Math. Biol. 1990, 28, 365–382. [Google Scholar] [CrossRef] [PubMed]
- Van den Driessche, P.; Watmough, J. Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef]
- Li, J.; Blakeley, D.; Smith, R.J. The Failure of R0. Comput. Math. Methods Med. 2011, 2011, 527610. [Google Scholar] [CrossRef]
- Hugh-Jones, M.; Blackburn, J. The ecology of Bacillus anthracis. Mol. Aspects Med. 2009, 30, 356–367. [Google Scholar] [CrossRef]
- Almberg, E.S.; Cross, P.C.; Johnson, C.J.; Heisey, D.M.; Richards, B.J. Modeling routes of chronic wasting disease transmission: Environmental prion persistence promotes deer population decline and extinction. PLoS ONE 2011, 6, e19896. [Google Scholar] [CrossRef]
- Turner, W.C.; Kausrud, K.L.; Krishnappa, Y.S.; Cromsigt, J.P.G.M.; Ganz, H.H.; Mapaure, I.; Cloete, C.C.; Havarua, Z.; Küsters, M.; Getz, W.M.; et al. Fatal attraction: Vegetation responses to nutrient inputs attract foraging herbivores to infectious anthrax carcass sites. Proc. R. Soc. B Biol. Sci. 2014, 281. [Google Scholar] [CrossRef]
- Turner, W.C.; Imologhome, P.; Havarua, Z.; Kaaya, G.P.; Mfune, J.K.; Mpofu, I.D.; Getz, W.M. Soil ingestion, nutrition and the seasonality of anthrax in herbivores of Etosha National Park. Ecosphere 2013, 4, 1–19. [Google Scholar] [CrossRef]
- Palmer, M.V.; Waters, W.R.; Whipple, D.L. Aerosol exposure of white-tailed deer (Odocoileus virginianus) to Mycobacterium bovis. J. Wildl. Dis. 2003, 39, 817–823. [Google Scholar] [CrossRef]
- Piercy, T.; Smither, S.; Steward, J.; Eastaugh, L.; Lever, M. The survival of filoviruses in liquids, on solid substrates and in a dynamic aerosol. J. Appl. Microbiol. 2010, 109, 1531–1539. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Whipple, D.L. Survival of Mycobacterium bovis on feedstuffs commonly used as supplemental feed for white-tailed deer (Odocoileus virginianus). J. Wildl. Dis. 2006, 42, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Aune, K.; Rhyan, J.C.; Russell, R.; Roffe, T.J.; Corso, B. Environmental persistence of Brucella abortus in the Greater Yellowstone Area. J. Wildl. Manag. 2012, 76, 253–261. [Google Scholar] [CrossRef]
- Mock, M.; Fouet, A. Anthrax. Annu. Rev. Microbiol. 2001, 55, 647–671. [Google Scholar] [CrossRef]
- Li, M.Y.; Muldowney, J.S. Global stability for the SEIR model in epidemiology. Math. Biosci. 1995, 125, 155–164. [Google Scholar] [CrossRef]
- Blackburn, J.K.; Goodin, D.G. Differentiation of Springtime Vegetation Indices Associated with Summer Anthrax Epizootics in West Texas, USA Deer. J. Wildl. Dis. 2013, 49, 699–703. [Google Scholar] [CrossRef]
- Colwell, R.R. Infectious disease and environment: Cholera as a paradigm for waterborne disease. Int. Microbiol. 2004, 7, 285–289. [Google Scholar]
- Nevas, M.; Lindström, M.; Hörman, A.; Keto-Timonen, R.; Korkeala, H. Contamination routes of Clostridium botulinum in the honey production environment. Environ. Microbiol. 2006, 8, 1085–1094. [Google Scholar] [CrossRef]
- Justice-Allen, A.; Trujillo, J.; Corbett, R.; Harding, R.; Goodell, G.; Wilson, D. Survival and replication of Mycoplasma species in recycled bedding sand and association with mastitis on dairy farms in Utah. J. Dairy Sci. 2010, 93, 192–202. [Google Scholar] [CrossRef]
- Colwell, R.R.; Spira, W.M. The ecology of Vibrio cholerae. Cholera 1992, 3, 107–127. [Google Scholar]
- Lipp, E.K.; Huq, A.; Colwell, R.R. Effects of global climate on infectious disease: The cholera model. Clin. Microbiol. Rev. 2002, 15, 757–770. [Google Scholar] [CrossRef]
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R.; Gotuzzo, E. Leptospirosis: A zoonotic disease of global importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef]
- Saunders, S.E.; Shikiya, R.A.; Langenfeld, K.; Bartelt-Hunt, S.L.; Bartz, J.C. Replication efficiency of soil-bound prions varies with soil type. J. Virol. 2011, 85, 5476–5482. [Google Scholar] [CrossRef]
- Saunders, S.E.; Bartelt-Hunt, S.L.; Bartz, J.C. Occurrence, transmission, and zoonotic potential of chronic wasting disease. Emerg. Infect. Dis. 2012, 18, 369–376. [Google Scholar] [CrossRef]
- Miller, M.W.; Williams, E.S.; Hobbs, N.T.; Wolfe, L.L. Environmental sources of prion transmission in mule deer. Emerg. Infect. Dis. 2004, 10, 1003–1006. [Google Scholar] [CrossRef]
- Lindner, D.L.; Gargas, A.; Lorch, J.M.; Banik, M.T.; Glaeser, J.; Kunz, T.H.; Blehert, D.S. DNA-based detection of the fungal pathogen Geomyces destructans in soils from bat hibernacula. Mycologia 2011, 103, 241–246. [Google Scholar] [CrossRef]
- Lorch, J.M.; Lindner, D.L.; Gargas, A.; Muller, L.K.; Minnis, A.M.; Blehert, D.S. A culture-based survey of fungi in soil from bat hibernacula in the eastern United States and its implications for detection of Geomyces destructans, the causal agent of bat white-nose syndrome. Mycologia 2013, 105, 237–252. [Google Scholar] [CrossRef]
- Miller, M.; Gardner, I.; Kreuder, C.; Paradies, D.; Worcester, K.; Jessup, D.; Dodd, E.; Harris, M.; Ames, J.; Packham, A. Coastal freshwater runoff is a risk factor for Toxoplasma gondii infection of southern sea otters (Enhydra lutris nereis). Int. J. Parasitol. 2002, 32, 997–1006. [Google Scholar] [CrossRef]
- Miller, M.; Miller, W.; Conrad, P.; James, E.; Melli, A.; Leutenegger, C.; Dabritz, H.; Packham, A.; Paradies, D.; Harris, M. Type X Toxoplasma gondii in a wild mussel and terrestrial carnivores from coastal California: New linkages between terrestrial mammals, runoff and toxoplasmosis of sea otters. Int. J. Parasitol. 2008, 38, 1319–1328. [Google Scholar] [CrossRef]
- Blackburn, J.K.; Mullins, J.C.; Van Ert, M.; Hadfield, T.; O’Shea, B.; Hugh-Jones, M.E. The necrophagous fly anthrax transmission pathway: Empirical and genetic evidence from a wildlife epizootic in West Texas 2010. Vector Borne Zoonotic Dis. 2014, 14, 576–583. [Google Scholar] [CrossRef]
- Blackburn, J.K.; Hadfield, T.L.; Curtis, A.J.; Hugh-Jones, M.E. Spatial and temporal patterns of anthrax in white-tailed deer, Odocoileus virginianus, and hematophagous flies in West Texas during the summertime anthrax risk period. Ann. Assoc. Am. Geogr. 2014, 104, 939–958. [Google Scholar] [CrossRef]
- Rohani, P.; Breban, R.; Stallknecht, D.E.; Drake, J.M. Environmental transmission of low pathogenicity avian influenza viruses and its implications for pathogen invasion. Proc. Natl. Acad. Sci. USA 2009, 106, 10365–10369. [Google Scholar] [CrossRef]
- Brown, V.L.; Drake, J.M.; Stallknecht, D.E.; Brown, J.D.; Pedersen, K.; Rohani, P. Dissecting a wildlife disease hotspot: The impact of multiple host species, environmental transmission and seasonality in migration, breeding and mortality. J. R. Soc. Interface 2013, 10. [Google Scholar] [CrossRef]
- Blackburn, J.K.; McNyset, K.M.; Curtis, A.; Hugh-Jones, M.E. Modeling the geographic distribution of Bacillus anthracis, the causative agent of anthrax disease, for the contiguous United States using predictive ecologic niche modeling. Am. J. Trop. Med. Hyg. 2007, 77, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Mullins, J.; Lukhnova, L.; Aikimbayev, A.; Pazilov, Y.; Van Ert, M.; Blackburn, J.K. Ecological Niche Modelling of the Bacillus anthracis A1.a sub-lineage in Kazakhstan. BMC Ecol. 2011, 11, 32. [Google Scholar] [CrossRef]
- Mullins, J.C.; Garofolo, G.; Van Ert, M.; Fasanella, A.; Lukhnova, L.; Hugh-Jones, M.E.; Blackburn, J.K. Ecological Niche Modeling of Bacillus anthracis on Three Continents: Evidence for Genetic-Ecological Divergence? PLoS ONE 2013, 8, e72451. [Google Scholar] [CrossRef]
- Morris, L.R.; Proffitt, K.M.; Asher, V.; Blackburn, J.K. Elk resource selection and implications for anthrax management in Montana. J. Wildl. Manag. 2016, 80, 235–244. [Google Scholar] [CrossRef]
- Havarua, Z.; Turner, W.C.; Mfune, J.K. Seasonal variation in foraging behaviour of plains zebra (Equus quagga) may alter contact with the anthrax bacterium (Bacillus anthracis). Can. J. Zool. 2014, 92, 331–337. [Google Scholar] [CrossRef]
- Cizauskas, C.A.; Bellan, S.E.; Turner, W.C.; Vance, R.E.; Getz, W.M. Frequent and seasonally variable sublethal anthrax infections are accompanied by short-lived immunity in an endemic system. J. Anim. Ecol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Saile, E.; Koehler, T. Bacillus anthracis multiplication, persistence, and genetic exchange in the rhizosphere of grass plants. Appl. Environ. Microbiol. 2006, 72, 3168–3174. [Google Scholar] [CrossRef]
- Hendriksen, N.B.; Hansen, B.M. Long-term survival and germination of Bacillus thuringiensis var. kurstaki in a field trial. Can. J. Microbiol. 2002, 48, 256–261. [Google Scholar] [CrossRef]
- Gu, G.; Hu, J.; Cevallos-Cevallos, J.M.; Richardson, S.M.; Bartz, J.A.; van Bruggen, A.H. Internal colonization of Salmonella enterica serovar Typhimurium in tomato plants. PLoS ONE 2011, 6, e27340. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.B.; Yaron, S.; Matthews, K.R. Transmission of Escherichia coli O157: H7 from contaminated manure and irrigation water to lettuce plant tissue and its subsequent internalization. Appl. Environ. Microbiol. 2002, 68, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Debré, P. Louis Pasteur; Johns Hopkins University Press: Baltimore, MD, USA, 1998; ISBN 0801858089. [Google Scholar]
- Van Ness, G.B. Ecology of Anthrax. Science 1971, 172, 1303–1307. [Google Scholar] [CrossRef] [PubMed]
- Lindeque, P.M.; Turnbull, P.C.B. Ecology and Epidemiology of Anthrax in the Etosha-National-Park, Namibia. Onderstepoort J. Vet. Res. 1994, 61, 71–83. [Google Scholar] [PubMed]
- Ganz, H.H.; Karaoz, U.; Getz, W.M.; Versfeld, W.; Brodie, E.L. Diversity and structure of soil bacterial communities associated with vultures in an African savanna. Ecosphere 2012, 3, art47. [Google Scholar] [CrossRef]
- Manchee, R.J.; Broster, M.G.; Melling, J.; Henstridge, R.M.; Stagg, A.J. Bacillus-Anthracis on Gruinard Island. Nature 1981, 294, 254–255. [Google Scholar] [CrossRef]
- Williams, G.; Linley, E.; Nicholas, R.; Baillie, L. The role of the exosporium in the environmental distribution of anthrax. J. Appl. Microbiol. 2013, 114, 396–403. [Google Scholar] [CrossRef]
- Raymond, B.; Wyres, K.L.; Sheppard, S.K.; Ellis, R.J.; Bonsall, M.B. Environmental factors determining the epidemiology and population genetic structure of the Bacillus cereus group in the field. PLoS Pathog. 2010, 6, e1000905. [Google Scholar] [CrossRef]
- Towne, E.G. Prairie vegetation and soil nutrient responses to ungulate carcasses. Oecologia 2000, 122, 232–239. [Google Scholar] [CrossRef]
- Danell, K.; Berteaux, D.; Bråthen, K.A. Effect of muskox carcasses on nitrogen concentration in tundra vegetation. Arctic 2002, 389–392. [Google Scholar] [CrossRef]
- Bump, J.K.; Peterson, R.O.; Vucetich, J.A. Wolves modulate soil nutrient heterogeneity and foliar nitrogen by configuring the distribution of ungulate carcasses. Ecology 2009, 90, 3159–3167. [Google Scholar] [CrossRef]
- Melis, C.; Selva, N.; Teurlings, I.; Skarpe, C.; Linnell, J.D.C.; Andersen, R. Soil and vegetation nutrient response to bison carcasses in Białowieża Primeval Forest, Poland. Ecol. Res. 2007, 22, 807–813. [Google Scholar] [CrossRef]
- Kukielka, E.; Barasona, J.; Cowie, C.; Drewe, J.; Gortazar, C.; Cotarelo, I.; Vicente, J. Spatial and temporal interactions between livestock and wildlife in South Central Spain assessed by camera traps. Prev. Vet. Med. 2013, 112, 213–221. [Google Scholar] [CrossRef]
- VerCauteren, K.C.; Burke, P.W.; Phillips, G.E.; Fischer, J.W.; Seward, N.W.; Wunder, B.A.; Lavelle, M.J. Elk use of wallows and potential chronic wasting disease transmission. J. Wildl. Dis. 2007, 43, 784–788. [Google Scholar] [CrossRef]
- Joyner, T.; Lukhnova, L.; Pazilov, Y.; Temiralyeva, G.; Hugh-Jones, M.; Aikimbayev, A.; Blackburn, J. Modeling the potential distribution of Bacillus anthracis under multiple climate change scenarios for Kazakhstan. PLoS ONE 2010, 5, e9596. [Google Scholar] [CrossRef]
- Blackburn, J. Integrating geographic information systems and ecological niche modeling into disease ecology: A case study of Bacillus anthracis in the United States and Mexico. In Emerging and Endemic Pathogens: Advances in Surveillance, Detection, and Identification; O’Connell, K.P., Skowronski, E.W., Sulakvelidze, A., Bakanidze, L., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 59–88. [Google Scholar]
- Steenkamp, P.J. Ecological Suitability Modelling for Anthrax in the Kruger National Park, South Africa. Master’s Thesis, University of Pretoria, Pretoria, South Africa, January 2013. [Google Scholar]
- Escobar, L.E.; Ryan, S.J.; Stewart-Ibarra, A.M.; Finkelstein, J.L.; King, C.A.; Qiao, H.; Polhemus, M.E. A global map of suitability for coastal Vibrio cholerae under current and future climate conditions. Acta Trop. 2015, 149, 202–211. [Google Scholar] [CrossRef]
- Jia, P.; Joyner, A. Human brucellosis occurrences in inner mongolia, China: A spatio-temporal distribution and ecological niche modeling approach. BMC Infect. Dis. 2015, 15, 36. [Google Scholar] [CrossRef]
- Kracalik, I.T.; Blackburn, J.K.; Lukhnova, L.; Pazilov, Y.; Hugh-Jones, M.E.; Aikimbayev, A. Analysing the spatial patterns of livestock anthrax in Kazakhstan in relation to environmental factors: A comparison of local (Gi*) and morphology cluster statistics. Geospatial Health 2012, 7, 111–126. [Google Scholar] [CrossRef]
- Kracalik, I.T.; Malania, L.; Tsertsvadze, N.; Manvelyan, J.; Bakanidze, L.; Imnadze, P.; Tsanava, S.; Blackburn, J.K. Evidence of Local Persistence of Human Anthrax in the Country of Georgia Associated with Environmental and Anthropogenic Factors. PLoS Negl. Trop. Dis. 2013, 7, e2388. [Google Scholar] [CrossRef]
- Osnas, E.E.; Heisey, D.M.; Rolley, R.E.; Samuel, M.D. Spatial and temporal patterns of chronic wasting disease: Fine-scale mapping of a wildlife epidemic in Wisconsin. Ecol. Appl. 2009, 19, 1311–1322. [Google Scholar] [CrossRef]
- Parkinson, R.; Rajic, A.; Jenson, C. Investigation of an anthrax outbreak in Alberta in 1999 using a geographic information system. Can. Vet. J. 2003, 44, 315–318. [Google Scholar]
- Turner, A.; Galvin, J.; Rubira, R.; Miller, G. Anthrax explodes in an Australian summer. J. Appl. Microbiol. 1999, 87, 196–199. [Google Scholar] [CrossRef]
- Ganz, H.H.; Turner, W.C.; Brodie, E.L.; Kusters, M.; Shi, Y.; Sibanda, H.; Torok, T.; Getz, W.M. Interactions between Bacillus anthracis and Plants May Promote Anthrax Transmission. PLoS Negl. Trop. Dis. 2014, 8, e2903. [Google Scholar] [CrossRef]
- Proffitt, K.M.; Gude, J.A.; Hamlin, K.L.; Garrott, R.A.; Cunningham, J.A.; Grigg, J.L. Elk distribution and spatial overlap with livestock during the brucellosis transmission risk period. J. Appl. Ecol. 2011, 48, 471–478. [Google Scholar] [CrossRef]
- O’Brien, D.J.; Schmitt, S.M.; Fitzgerald, S.D.; Berry, D.E.; Hickling, G.J. Managing the wildlife reservoir of Mycobacterium bovis: The Michigan, USA, experience. Vet. Microbiol. 2006, 112, 313–323. [Google Scholar] [CrossRef]
- Johnson, D.H. The comparison of usage and availability measurements for evaluating resource preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
- Nekorchuk, D.M.; Morris, L.R.; Asher, V.; Hunter, D.L.; Ryan, S.J.; Blackburn, J.K. Potential Bacillus anthracis Risk Zones for Male Plains Bison (Bison bison bison) in Southwestern Montana, USA. J. Wildl. Dis. 2019, 55, 136–141. [Google Scholar] [CrossRef]
- Getz, W.M.; Wilmers, C.C. a local nearest-neighbor convex-hull construction of home ranges and ultization distributions. Ecography 2004, 27, 489–505. [Google Scholar] [CrossRef]
- Jennrich, R.; Turner, F. Measurement of non-circular home range. J. Theor. Biol. 1969, 22, 227–237. [Google Scholar] [CrossRef]
- Lyons, A.J.; Turner, W.C.; Getz, W.M. Home Range Plus: A Space-Time Characterization of Movement over Real Landscapes. Mov. Ecol. 2013, 1, 2. [Google Scholar] [CrossRef]
- Dougherty, E.R.; Carlson, C.J.; Blackburn, J.K.; Getz, W.M. A cross-validation-based approach for delimiting reliable home range estimates. Mov. Ecol. 2017, 5, 19. [Google Scholar] [CrossRef]
- Dougherty, E.R.; de Valpine, P.; Carlson, C.J.; Blackburn, J.K.; Getz, W.M. Commentary to: A cross-validation-based approach for delimiting reliable home range estimates. Mov. Ecol. 2018, 6, 10. [Google Scholar] [CrossRef]
- Bagamian, K.H.; Alexander, K.A.; Hadfield, T.L.; Blackburn, J.K. Ante- and postmortem diagnostic techniques for anthrax: Rethinking pathogen exposure and the geographic extent of the disease in wildlife. J. Wildl. Dis. 2013, 49, 786–801. [Google Scholar] [CrossRef]
- Bellan, S.E.; Gimenez, O.; Choquet, R.; Getz, W.M. A Hierarchical Distance Sampling Approach to Estimating Mortality Rates from Opportunistic Carcass Surveillance Data. Methods Ecol. Evol. 2013. [Google Scholar] [CrossRef]
- Blackburn, J.K.; Asher, V.; Stokke, S.; Hunter, D.L.; Alexander, K.A. Dances with Anthrax: Wolves (Canis lupus) Kill Anthrax Bacteremic Plains Bison (Bison bison bison) in Southwestern Montana. J. Wildl. Dis. 2014, 50, 393–396. [Google Scholar] [CrossRef]
- Turner, W.C.; Getz, W.M. Seasonal and demographic factors influencing gastrointestinal parasitism in ungulates of Etosha National Park. J. Wildl. Dis. 2010, 46, 1108–1119. [Google Scholar] [CrossRef]
- Cizauskas, C.A.; Turner, W.C.; Wagner, B.; Küsters, M.; Vance, R.E.; Getz, W.M. Gastrointestinal helminths may affect host susceptibility to anthrax through seasonal immune trade-offs. BMC Ecol. 2014, 14, 27. [Google Scholar] [CrossRef]
- Gomez, J.P.; Nekorchuk, D.M.; Mao, L.; Ryan, S.J.; Ponciano, J.M.; Blackburn, J.K. Decoupling environmental effects and host population dynamics for anthrax, a classic reservoir-driven disease. PLoS ONE 2018, 13, e0208621. [Google Scholar] [CrossRef] [PubMed]
- Bhadra, A.; Ionides, E.L.; Laneri, K.; Pascual, M.; Bouma, M.; Dhiman, R.C. Malaria in Northwest India: Data analysis via partially observed stochastic differential equation models driven by Lévy noise. J. Am. Stat. Assoc. 2011, 106, 440–451. [Google Scholar] [CrossRef]
- Böttcher, B. Feller processes: The next generation in modeling. Brownian motion, Lévy processes and beyond. PLoS ONE 2010, 5, e15102. [Google Scholar] [CrossRef] [PubMed]
- Kallenberg, O. Foundations of Modern Probability; Springer: New York, NY, USA, 2002; ISBN 0-387-95313-2. [Google Scholar]
- Bertoin, J. On the first exit time of a completely asymmetric stable process from a finite interval. Bull. Lond. Math. Soc. 1996, 28, 514–520. [Google Scholar] [CrossRef]
- Steutel, F.; Van Harn, K. Discrete analogues of self-decomposability and stability. Ann. Probab. 1979, 893–899. [Google Scholar] [CrossRef]
- McKenzie, E. Some ARMA models for dependent sequences of Poisson counts. Adv. Appl. Probab. 1988, 822–835. [Google Scholar] [CrossRef]
- Leonenko, N.; Savani, V.; Zhigljavsky, A. Autoregressive Negative Binomial Processes. Annales de l'Institut de Statistique de l'Universite de Paris. 2007, Volume 51, pp. 25–47. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.332.4844&rep=rep1&type=pdf (accessed on 14 March 2019).
- Manly, B.; McDonald, L.; Thomas, D.; McDonald, T.; Erickson, W. Resource Selection by Animals: Statistical Analysis and Design for Field Studies; Kluwer: New York, NY, USA, 2002. [Google Scholar]
- Fisher, R.A.; Corbet, A.S.; Williams, C.B. The relation between the number of species and the number of individuals in a random sample of an animal population. J. Anim. Ecol. 1943, 12, 42–58. [Google Scholar] [CrossRef]
- Ponciano, J.M.; Burleigh, J.G.; Braun, E.L.; Taper, M.L. Assessing parameter identifiability in phylogenetic models using Data Cloning. Syst. Biol. 2012, 61, 955–972. [Google Scholar] [CrossRef]
- Getz, W.M. Disease and the dynamics of food webs. PLoS Biol. 2009, 7, e1000209. [Google Scholar] [CrossRef]
- Dragon, D.; Elkin, B.; Nishi, J.; Ellsworth, T. A review of anthrax in Canada and implications for research on the disease in northern bison. J. Appl. Microbiol. 1999, 87, 208–213. [Google Scholar] [CrossRef]
- Kracalik, I.; Malania, L.; Tsertsvadze, N.; Manvelyan, J.; Bakanidze, L.; Imnadze, P.; Tsanava, S.; Blackburn, J.K. Human Cutaneous Anthrax, Georgia 2010–2012. Emerg. Infect. Dis. 2014, 20, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Wint, G.; Robinson, T.; Bourn, D.; Durr, P.; Hay, S.; Randolph, S.; Rogers, D. Mapping bovine tuberculosis in Great Britain using environmental data. Trends Microbiol. 2002, 10, 441–444. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Disease | Pathogen | Host | Environmental Reservoir | Local Infectious Zone (LIZ) | Landscape Characteristics | Survival Time in Environment | References |
|---|---|---|---|---|---|---|---|
| Anthrax | Bacillus anthracis | Wildlife and livestock | Host, bones, soil, water, vegetation | Carcass site, water’s edge | Grasslands, scrub/pothole regions | >1 year | [9] |
| Botulism | Clostridium botulinum | Birds & mammals | Host, honey, soil | Carcass site, honeybee colony | Cosmopolitan | >1 year | [21] |
| Bovine mastitis | Mycoplasma bovis | Bovids | Host, soil and/or animal bedding | Bedding within feedlot | Broad conditions | ~1 year (needs futher study) | [22] |
| Brucellosis | Brucella spp. | Wildlife and livestock | Host, soil and/or birthing tissues, aborted fetuses | birthing tissues and aborted fetuses | ~20–80 days (needs further study) | [16] | |
| Cholera | Vibrio cholerae | Humans | Host, feces, zooplankton, saltwater | Estuaries | Periurban, coastal regions | [23,24] | |
| Leptospirosis | Leptospira spp. | Animals, humans | Host, grass, moist soil, water | Grasslands, streams, rivers, ponds, lakes | Periurban, contaminated lakes | [25] | |
| Chronic wasting disease | Prions | Cervids | Host, some soils | Salt/mineral sites, wallows | Host range & soils overlap | [26,27,28] | |
| White-nosed syndrome | Psuedogymnoascus destructans | Hibernating bats | Host, some soils | Bat hibernacula | Cave system or mountain range | [29,30] | |
| Toxoplasmosis | Toxoplasma gondii | Mammals | Host, feces, soil, invertebrates | Soils, streams, bays, estuaries | Periurban areas, coastal regions | [31,32] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blackburn, J.K.; Ganz, H.H.; Ponciano, J.M.; Turner, W.C.; Ryan, S.J.; Kamath, P.; Cizauskas, C.; Kausrud, K.; Holt, R.D.; Stenseth, N.C.; et al. Modeling R0 for Pathogens with Environmental Transmission: Animal Movements, Pathogen Populations, and Local Infectious Zones. Int. J. Environ. Res. Public Health 2019, 16, 954. https://doi.org/10.3390/ijerph16060954
Blackburn JK, Ganz HH, Ponciano JM, Turner WC, Ryan SJ, Kamath P, Cizauskas C, Kausrud K, Holt RD, Stenseth NC, et al. Modeling R0 for Pathogens with Environmental Transmission: Animal Movements, Pathogen Populations, and Local Infectious Zones. International Journal of Environmental Research and Public Health. 2019; 16(6):954. https://doi.org/10.3390/ijerph16060954
Chicago/Turabian StyleBlackburn, Jason K., Holly H. Ganz, José Miguel Ponciano, Wendy C. Turner, Sadie J. Ryan, Pauline Kamath, Carrie Cizauskas, Kyrre Kausrud, Robert D. Holt, Nils Chr. Stenseth, and et al. 2019. "Modeling R0 for Pathogens with Environmental Transmission: Animal Movements, Pathogen Populations, and Local Infectious Zones" International Journal of Environmental Research and Public Health 16, no. 6: 954. https://doi.org/10.3390/ijerph16060954