
The Search of a Molecular “Swiss Knife” for Chloroplast Genomic Editing

Institute of Cytology and Genetics, Siberian Branch, Russian Academy of Sciences, Novosibirsk 630090, Russia
*
Author to whom correspondence should be addressed.
Horticulturae 2023, 9(12), 1338; https://doi.org/10.3390/horticulturae9121338
Submission received: 7 November 2023
/
Revised: 7 December 2023
/
Accepted: 12 December 2023
/
Published: 14 December 2023
(This article belongs to the Special Issue Selected Papers from the 3rd International Conference “Genetically Modified Organisms: The Нistory, Achievements, Social and Environmental Risks”)
Abstract
:In recent years, genome editing methods have become an integral part of the genetic engineering toolset that allows for making targeted changes to plant genomes, both in the case of single-gene mutations and multiplex modifications. These technologies were mostly proven effective for editing nuclear genomes. However, plastids, the best-known example of which is chloroplasts, have their own genome (plastome), which is also available for various genetic manipulations, including editing. Despite the fact that the modification of plastomes represents a very promising task for modern biotechnology, the structure of plastids and the peculiarities of their genome organization require the specific adaptation of genome editing methods. This applies to both the design of genetic constructs and methods of their delivery to plastids. The article provides an overview of the current state of research in the field of plastid genome editing with chloroplasts taken as an example. We consider the possibilities of using programmable genome-editing technologies, analyze their effectiveness, limitations, and problems caused by the structural features of these organelles, and their genome organization. We discuss the results of the first successful experiments in this field and try to assess the prospects for the development of tools and methods for increasing the efficiency and the specificity of this biotechnological platform.
Keywords:
chloroplast; plastome; genome editing; base editing; transformation; signal peptides; nanoparticles1. Introduction
Genome editing methods that allow for the intended modification and regulation of gene expression are the most in-demand in modern plant biotechnology [1,2]. Technologies based on the use of programmable nucleases, CRISPR/Cas9 in particular, have become an effective tool for targeted mutagenesis, including single- and multiple-nucleotide substitutions, gene knockouts and knockdowns, and multiplex genome modifications [3,4]. For over a decade, these technologies have been applied for editing the nuclear genome and developing new varieties with desired traits, starting from model plants and ending with agricultural crops and bioproducers [5,6,7]. Over the past few years, these methods have begun to be adapted for the genetic engineering of chloroplasts [8,9].
Compared to the nuclear genome, the chloroplast genome’s organization has a number of distinct biotechnological advantages. The system of homologous recombination, multiple copies of the chloroplast DNA, and the absence of gene silencing made it possible to improve the accuracy of transgene construct integration, increase its expression level, and accumulate significant volumes of heterologous proteins [10]. In addition, chloroplast genes are organized into operons, which allows transgenes to be combined into artificial operons within a single transformation event [11,12]. These characteristics provide advantages in the use of chloroplasts as bioproducers of recombinant proteins for various purposes, but cause problems for editing their genome.
One of the main tasks of the genome editing strategy is associated with achieving site-specific point mutations, base pair substitutions, and insertions or deletions of one or several nucleotides. This result is possible, owing to non-homologous recombination—an error-prone DNA repair mechanism mediated by non-homologous end joining (NHEJ). This mechanism prevails in the nucleus, but for chloroplasts, the typical mechanism is homologous recombination, which restores the original DNA sequence, thus the preventing occurrence of modifications [8,13].
Multiple copies of the chloroplast genome also cause difficulties for the genome editing of these organelles, as it is necessary to obtain and select homoplastomic lines and prevent the reverse mutation of the modified genetic sequence [8]. Achieving homoplasmy under high polyploidy requires the development of additional techniques to increase editing efficiency. All of this explains why genome editing is not yet effective enough in its application to chloroplasts.
It should also be taken into account that the chloroplast DNA contains just 100–250 genes encoding only 5% of proteins necessary for these organelles. These are mainly proteins involved in photosynthesis, RNA binding, ribosomal proteins, and proteins of the transcription/translation system [14]. This circumstance minimizes the number of problems that could be solved via the genome editing of the plastome. However, over the past 3–4 years, a whole series of studies have appeared, focusing on the development of this particular area in plant genetic engineering. There were experiments in the modification of genes responsible for photosynthesis; new methods were developed that allow for the use of genome-editing tools to enhance plastome transformation efficiency, and new technological solutions were found for delivering genome editing components into chloroplasts [9,15]. The relevance of these studies is determined by the need to develop editing methods and tools applicable to photosynthesis genes and, especially, the gene encoding the most important enzyme of the Calvin cycle, RuBisCO (Ribulose-1,5-bisphosphate carboxylase/oxygenase) [16]. Increasing the catalytic activity of the photosystem proteins, especially RuBisCo, will enable a more effective use of absorbed light energy and an improvement in the properties and yield of agricultural crops.
In this review, we summarize information about the development of technologies for plastid genome editing. We provide an overview of the existing tools and approaches, discuss the current technological limitations, emphasizing the issue of why editing tools successfully used for the nuclear genome turned out to be ineffective for chloroplasts. We are also considering opportunities for and ways of further developing a universal toolset to find a molecular “Swiss army knife” for solving various problems of plastid genome modification.
2. Genome Editing Systems Applicable to Chloroplasts
For chloroplast genome editing, researchers can use the now universally applicable systems developed on the basis of programmable site-specific nucleases—ZFN (zinc finger nucleases), TALENs (transcription activator-like effector nucleases) and CRISPR/Cas9 (clustered regularly interspaced short palindromic repeats) [9,10,17,18]. ZFN and TALENs are synthetic heterodimer proteins, each subunit of which contains a DNA-recognizing protein domain that binds specific DNA sequences and an endonuclease domain that induces double-strand breaks (DSB). The practical application of the ZFN-based technique proved to be too labor-consuming and often provided unpredictable results due to off-target effects [19]. There are few examples of applying ZFN in plant genetic engineering, and this technology was ineffective for chloroplast genome editing.
The use of the TALEN nuclease complex significantly increased the efficiency of genome editing in different groups of organisms, including plants [20]. The DNA-binding TALE domain consists of a certain set of tandem repeats, each of which allows for the recognition of one nucleotide in the target DNA. The distinctive feature of this domain is the presence of two highly variable amino acid residues, RVD (repeat-variable di-residues), capable of binding only to a certain nucleotide, which imparts high specificity to the entire construct. The second main domain was originally a catalytic subunit of the bacterial endonuclease FokI [3,20,21]. However, experiments using TALEN to cause a site-specific double-strand breakage (DSB) of chloroplast DNA (cpDNA) in plants cannot be called successful. Although transgenic plants showed the appearance of point mutations in the target gene, most DNA breaks were quickly repaired due to homologous recombination without retaining any modifications [13,22]. Therefore, in this form, the TALEN system did not find applications in editing chloroplast genomes. The TALE-based editing strategies became effective for plastomes after the development of a tool, in which the TALE DNA-binding domain was used in combination with the deaminase of nucleotide bases. Deaminase replaces the endonuclease complex in the catalytic domain (TALED instead of TALEN). Two enzymes were used for base substitution. One of the enzymes, cytosine deaminase, catalyzes the deamination of cytosine and converts it into uracil, which, as a result of transcription and replication, is converted into thymine [23,24]. The other enzyme is adenine deaminase, which converts adenine into guanine [25]. The import of TALEDs into chloroplasts was achieved by adding a targeting sequence such as a chloroplast transit peptide (CTP) to the N-terminus of TALEs.
The approach using the chloroplast-targeted TALE base editor was effective for lettuce, rapeseed, arabidopsis, and rice [22,25,26,27,28]. The TALED not only optimized the technology for the genome editing of chloroplasts, but also made it possible to obtain successful results in an extremely short time (in 2–3 years). This tool is now effectively applied when working with plastomes, despite the existing limitations associated with the complexity of the design, assembly methods and bulky engineered construct of TALED itself. It should be noted that currently, it is possible to overcome the difficulties of assembling such structures using commercially available kits and tools. For example, the Golden Gate cloning-based TALEN construction kit and Platinum Gate TALEN Kit can significantly simplify the technical procedures.
The CRISPR/Cas editing system is based on the immune system of bacteria and archaea, which protects them from mobile genetic elements and bacteriophages [29]. It is a complex formed by a guide RNA (gRNA) and an endonuclease (Cas9). The gRNA is a single-stranded RNA that has a Cas nuclease-binding site and a sequence measuring approximately 20 bases long that is complementary to the DNA target sequence. The choice of a unique sequence in the genome ensures the high specificity of gRNA recognition and, accordingly, increases the accuracy of editing. The Cas9 enzymes contain an HNH domain, which cleaves the DNA strand complementary to the gRNA sequence (target strand), and a RuvC nuclease domain, which is necessary for cleaving the non-complementary strand (non-target strand), and thus are capable of producing double-strand breaks [30,31]. Subsequently, the double-strand breaks can be repaired via various DNA repair mechanisms: the use of non-homologous and homologous end joining. As a result, several types of mutations, such as deletions, insertions, substitutions, the integration of a specific DNA sequence at a desired locus, and site-specific corrections can occur with some frequency.
Non-homologous recombination is the main DNA repair pathway in the cell, but it is prone to significant errors because it involves the direct joining of ends without the participation of a homologous template. This mechanism is effective for the formation of nucleotide substitutions or deletions in target sites to obtain gene knockouts, and is actively used in genetic engineering to suppress or eliminate genes that negatively affect plant growth and productivity [32,33].
Homologous recombination is a high-precision mechanism that requires a homologous template to mediate repair and can be used to achieve accurate edits such as gene insertion and replacement. At the same time, to integrate new exogenous fragments into target sites, it is necessary to introduce a donor (template) DNA together with the CRISPR-Cas9 complex. Such DNA has at its ends nucleotide sequences identical to those at the sections of genomic DNA flanking the insertion region. The process of inserting sequences into the genome is based on recombination between a region of the genome and an exogenous DNA molecule [33,34].
As a genetic engineering platform, CRISPR-Cas9 came into use in 2012, and for plants it came into use in 2013. This approach proved to be easy to develop, cost-effective, and versatile in generating targeted gene mutations and alterations for a variety of plant species, from model plants to some economically important crops [35]. The implementation of this approach in editing chloroplast genomes has encountered some problems. First, the CRISPR/Cas systems currently in use for genome editing are critically dependent on guide RNAs. At present, there is no reliable method available to deliver RNAs into plastids. Secondly, as we wrote above, upon the creation of DSBs by the endonuclease Cas, the repair machinery often creates small insertions or deletions via NHEJ. However, in chloroplasts, DNA repair occurs predominantly via HR, which prevents the persistence of errors in the organelle genome. Because of these limitations, the CRISPR/Cas systems cannot yet be used effectively in editing chloroplast genomes. So far, only initial results have been obtained, allowing us to draw a conclusion about the potential applications of this system and further lines of research to optimize its use in this area. The primary tasks to be solved is the selection and optimization of methods for delivering genome-editing tools into plant cells and plastids, as well as bypassing the homologous recombination mechanism.
3. Delivery of Genome-Editing Tools into Chloroplasts
In the practice of plastid transformation, preference is given to two methods: the transfection of protoplasts in the presence of polyethylene glycol and bioballistics, which involves the delivery of expression constructs with the help of microparticles [36,37]. However, the choice of the method for delivering plastome-editing tools to chloroplasts primarily depends on the system in use and is not limited only to direct methods of delivering expression vectors to chloroplasts.
3.1. Delivery of DNA Plasmids into Chloroplasts Using Bioballistics
This direct delivery method involves immobilizing DNA on gold or tungsten microparticles, which then deliver this DNA into the plant cell, and thus the chloroplasts, using helium gas under high pressure [11]. A significant drawback of the method is that it requires special expensive equipment and has low productivity. Its availability is largely limited to those plant species for which the regeneration system has been developed. In addition, this method can lead to off-target mutations both in the nucleus and chloroplasts and can sometimes cause damage to chromosomes and cellular structures [38]. The frequency of achieved target modifications in chloroplast DNA also remains low [12].
However, in the genome editing of plastids, according to the literature data, it was exactly the bioballistic delivery of specially designed DNA-editing plasmids into the chloroplasts of the alga Chlamydomonas reinhardtii that brought positive results. These plasmids were shown to be able to autonomously replicate, express and produce Cas9, gRNA and donor DNA within the chloroplasts [39]. By using bioballistics, plasmids that encode editing tools were also successfully delivered into the chloroplasts of tobacco leaves, where they were effectively expressed, increasing the frequency of transgene integration [40].
3.2. Nuclear Transformation: Vector-Mediated Transfer and Direct Delivery
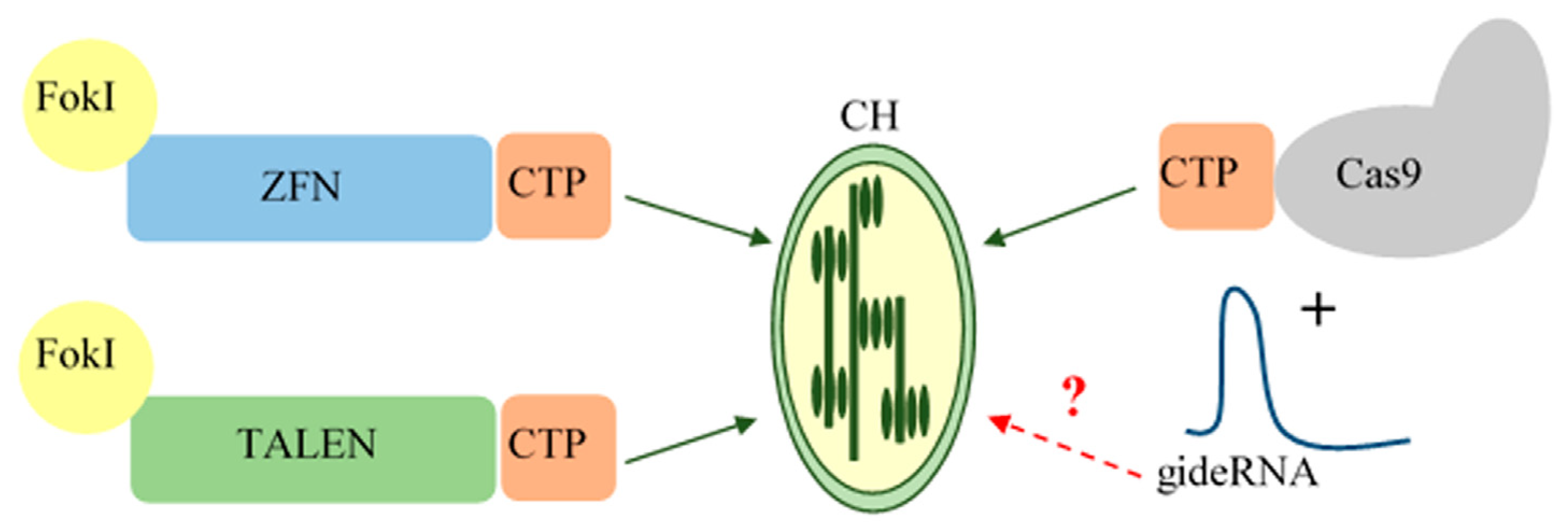
The expression products of genetically engineered constructs encoding ZFN- and TALE-based editing tools are synthetic heterodimer proteins. In this case, any delivery methods that are optimal for the nuclear transformation of plants are suitable, including direct delivery via bioballistics or vector delivery as part of T-DNA using agrobacterium-mediated transfer [22,24,27]. These methods can provide both the transient expression of genetically engineered constructs and stable nuclear transformation of plants with the subsequent synthesis of the editing complex. However, both in the case of bioballistics and agrobacterium-mediated transfer, with respect to nuclear transformation, it is an indispensable condition that the signal peptide (CTP—chloroplast transit peptide) must be present, and encoded in the expression cassette, which is to ensure the import of the editing complex into chloroplasts (Figure 1).
The major problem with the genome editing of plastomes using the simpler and more convenient CRISPR/Cas system is that the system consists of two components. While the endonuclease Cas supplemented with a transit peptide can be delivered into chloroplasts, in the same way as ZFN and TALE, the gRNA is believed to be unable to penetrate the double membrane of the organelles (Figure 1) due to strong electrochemical potential [13,41]. Consequently, nuclear transformation options in this case will be ineffective and other approaches must be applied.
3.3. gRNA Delivery into Chloroplasts
One of the approaches to using the CRISPR/Cas system for plastome editing is associated with improving the method of gRNA delivery [13]. For this purpose, the findings of studies on RNA import into chloroplasts can be used. Although the molecular mechanisms of this process are not fully known [42], there are experimental designs of RNA molecule delivery into this organelle using signal sequences. In particular, it was shown that the mRNA encoding GFP was delivered into the chloroplast by adding to its 5′UTR-end the signal NcRNA (non-coding RNA) sequence derived from a viroid of the Avsunviroidae family, capable of penetrating the organelle [43]. Similar functions were also found in proteins. The chloroplast protein phosphoglycerate kinase can also mediate the transport of the single-stranded genomic RNA of bamboo mosaic virus [44]. These data could potentially be used to develop more efficient methods for delivering gRNA components into chloroplasts, which is likely to increase the frequency of genetic editing in these organelles.
3.4. Delivery of DNA Plasmids into Chloroplasts Using Nanoparticles
The main barriers to particles and associated DNA while entering the chloroplast genome are the cell wall, the plant cell membrane, the cytosol, and the chloroplast double membrane. Currently, to solve this problem, new technological methods are being developed for the transfer of artificial genetic constructs into chloroplasts. One of these techniques is based on the use of nanoparticles ranging in size from 1 to 100 nanometers, of different shapes, aspect ratios, charges and surface chemistries. These nanoparticles can be composed of various materials, including silicon, gold, carbon and polymers [45,46]. In particular, it was shown that single-walled carbon nanotubes (SWCNTs) coated with a positively charged polymer such as chitosan and loaded with plasmid DNA can penetrate plant cell walls, the plasma membrane and chloroplast double membranes through the lipid exchange envelope penetration (LEEP) mechanism [45,47]. The release of DNA in the chloroplast stroma occurs due to an increase in pH, resulting in a weakening of electrostatic interaction. The creators of this method demonstrated its high efficiency; about 35% of nanoparticles carrying a plasmid with flanking sequences from the Panicum virgatum plastome were delivered into chloroplasts, and the expression of this plasmid was detected in 88% of chloroplasts [10,47]. However, it is unclear whether or not this approach can also be successfully implemented in the genome editing procedure, in particular, CRISPR/Cas. The possibilities of increasing the genome editing efficiency through the use of nanotechnology are being actively discussed, but this platform is currently still under development.
3.5. Delivery of DNA into Chloroplasts Using Signal Peptides
In addition to the methods described above, there are methods currently being developed for the targeted delivery of components of genome-editing systems using signal peptides. They combine, in their structure, functional peptides, such as cell-penetrating peptides (CPP), which can pass through cell membranes; polycationic peptides consisting of positively charged amino acids and binding to nucleic acids; as well as endogenous signal peptides that deliver proteins to organelles [48,49]. Such synthetic peptides can be used for targeted DNA delivery into the cell organelles of various plant tissue types.
For example, hybrid peptide KH-AtOEP34 was developed for plastids, and consists of the KH DNA-binding sequence and the plastid-targeted signal peptide OEP34. It ensures the selective delivery of plasmid DNA into chloroplasts and the expression of the reporter genes [50,51,52]. Such a signal peptide does not require any carriers, such as micro or nanoparticles, for delivery into cells. A suspension containing DNA–protein complexes can be applied to the leaf surface, and then, via endocytosis or direct penetration through stomata, the construct penetrates into the intercellular space and is absorbed by the plasma membrane. In the cell, the DNA–protein complexes are internalized by the chloroplast. DNA delivery by using signal peptides was successfully performed in tobacco, arabidopsis, rice, and hibiscus, which indicates the applicability of this method for different plant species [50,51,53]. Owing to signal peptides, it was possible to achieve a stable transformation of the plastid genome, while the use of SWCNTs (described above) has so far led to the transient expression of exogenous DNA only.
Although DNA transfer using signal peptides has not yet been optimized for genetic constructs encoding components of genome-editing systems, it is quite a promising technique because it eliminates the need for the preparation of protoplasts, callus formation, and the use of expensive special equipment (gene gun).
4. Practical Application of Chloroplast Genomic Editing Platforms
4.1. Base-Editing Technologies
Although the development of technologies for the genetic editing of the chloroplast genome remains a major challenge at present, active work is in progress in this area and there are some successful results attained already (Table 1).
One of the focus areas is introducing single-nucleotide substitutions via the deamination of the nitrogenous bases of cytosine and adenine by using enzymes cytidine deaminase and adenine deaminase, respectively. Platforms are being developed that include genome- editing tools in combination with transport or signaling molecules and a nitrogenous base deaminase. During 2021–2023, several research groups conducted successful experiments to obtain point mutations in target chloroplast genes [8,24,26,28]. The authors of these works optimized the TALED base editor, previously created for mitochondria, for chloroplasts [24,27,54]. The functionally active TALED editor is a heterodimer, each subunit of which is a hybrid protein consisting of a TALE effector domain, one half of the DddAtox bacterial deaminase, a signal peptide and, if cytidine deaminase is used, an uracil glycosylase inhibitor. As a result of the catalytic activity of deaminases, the conversion of CG pairs into TA or that of AT into GC occurs.
One of the first works on creation and use of such a platform was presented in an article by R. Li and co-authors [55]. The genetically engineered construct for the targeted editing of the rice chloroplast genome included coding sequences of the DNA-binding domain of the TALE effector from Xanthomonas oryzae, chloroplast signal peptide CTP, and cytidine deaminase DddAtox. PsaA, a conservative gene encoding a protein of photosystem I, was chosen as the target gene for converting CG into TA. As a result, the target change in nitrogenous bases was achieved, which led to the appearance of a premature stop codon and, as a consequence, the inactivation of the psaA gene. The mutant plants had an albino phenotype due to decreased chlorophyll production. Sequencing results showed that the editing efficiency reached 64% and the vast majority of the edited lines were homoplasmic.

{kind=link}
Table 1.
Chloroplast genes modified as a result of the application of genome editing technology.
| Target Gene | Genome Editor | Species | Transformation Method | References |
|---|---|---|---|---|
| psaA (chlorophyll a of photosystem I) | ptTALED (plastid-targeted TALE deaminase); CRISPR/Cas9 | rice, lettuce, arabidopsis, chlamydomonas | Agrobacterium, mRNA, biolistic | [25,39,55] |
| psbA (photosynthetic protein, D1) | ptTALED | lettuce, rapeseed, arabidopsis | Agrobacterium, mRNA, polyethylene glycol (PEG)-mediated | [23,25,26] |
| psbB (photosynthetic protein, CP-47) | ptTALED | lettuce, rapeseed | Agrobacterium | [23] |
| rrn16 (16SrRNA) | ptTALED | lettuce, rapeseed, arabidopsis | Agrobacterium, PEG-mediated | [22] |
| rbcL (large, catalytic subunit of RuBisCo) | ptTALED | lettuce, arabidopsis | Agrobacterium | [22] |
| rpoC1 (β subunit of RNA polymerase) | ptTALED | arabidopsis | Agrobacterium | [26] |
Apart from psaA, targeted editing of three more chloroplast genes of lettuce (Lactuca sativa) and arabidopsis was performed—rrn16 encoding 16S ribosomal RNA, psbA encoding a photosystem II protein, and rbcL encoding the large catalytic subunit of RuBisCo, an enzyme that catalyzes CO2 fixation [25]. Mutations in these genes cause resistance to antibiotics (rrn16) and herbicides (psbA), and also lead to an albino phenotype (psaA). The TadA8e adenine deaminase of Escherichia coli was used as a base editor, which catalyzes the deamination of adenine, converting A into G. This system was supplemented with the signal peptide of arabidopsis chloroplasts, RecA1. The work was carried out both for a protoplast culture (lettuce) and with the production of a whole edited plant (arabidopsis). The protoplasts were transfected with messenger RNAs encoding TALED and the signal peptide, while agrobacterium-mediated transformation was used for arabidopsis. The results of deep sequencing showed that, depending on the gene, the frequency varied from 25% to 51% in lettuce protoplasts and reached 99% for arabidopsis in some variants. An important achievement of this work was the production of arabidopsis plants with heritable homoplasmic mutations.
Nakazato and co-authors [26] applied the same technology for editing target nitrogenous bases in three arabidopsis genes. One of them is 16S rRNA, the modification of which confers antibiotic resistance. The other two genes, modifications of which lead to growth retardation, are rpoC1 encoding the beta subunit of DNA-dependent RNA polymerase and psbA encoding a photosystem II protein. A modified version of base editor DdCBE was created, which was named plastid-targeted platinum TALE cytidine deaminase or ptpTALECD [26,28]. For transformation, a single binary vector encoding the ptpTALECD pair was used, which was introduced into the nuclear genome of A. thaliana using the floral dip method. The authors showed that the homoplasmic substitution in the first generation of transformants reached 54%. In addition, it was confirmed that the introduced substitution was stably inherited by the next generation [26,28].
Another group of researchers, using similar editing tools, created an effective platform for generating site-specific point mutations in chloroplasts [23]. With the help of the GoldenGate cloning methodology, a set of plasmids was developed consisting of 424 effector plasmids encoding TALE and 8 expression plasmids encoding a hybrid protein that consists of chloroplast signal peptide CTP and the DddA bacterial cytosine deaminase domain. The combination of effector and expression plasmids resulted in the formation of one of the DdCBE base editor variants. The authors used this approach to induce the base editing of chloroplast genomes in the lettuce and rapeseed callus, and in genes psbA, psbB, and rpoC1, and demonstrated editing frequencies of up to 38%.
In all these works, the TALE-based system was chosen as the genome editing technique, in which the catalytic center was changed from an endonuclease to a deaminase. The CRISPR/Cas9 tool proved to be ineffective for solving these problems, presumably due to the electrochemical potential of the chloroplast double membrane, which creates a barrier to the penetration of guide RNA [13].
4.2. Development of CRISPR/Cas9 System to Increase the Efficiency of Plastid Transformation
One of the most important advantages obtained by modifying the genome of plastids, in particular chloroplasts, is their potential ability to produce very high levels of recombinant proteins, by far exceeding the capabilities of nuclear transformants. However, the efficiency of plastome transformation and the yield of transformant plants still remain at an extremely low level, even when working with model plant species, which seriously hinders the development of this technology [12].
The production of transplastomic plants involves the insertion of genetic constructs encoding target genes and dominant selective markers into the plastid genome. The integration of a new nucleotide sequence into the target site occurs due to the homologous recombination of plastid DNA with an artificially synthesized template. The formation of double-strand breaks is a precondition for the successful integration of a genetic construct. Thus, this process is similar to that of genome editing in the mode of so-called knock-in modifications and requires delivery to chloroplasts not only of the editing tools, but also of the template itself. The following question arises: is there a practical opportunity to increase the efficiency of transformation using CRISPR/Cas9?
To date, two research groups have managed to perform effective CRISPR/Cas9 editing of the chloroplast genome, while achieving the integration of an exogenous DNA sequence into the target site.
Yoo and co-authors developed Edit plasmids that were delivered into the chloroplasts of Chlamydomonas reinhardtii algae via bioballistics [39]. A distinctive feature of these plasmids was their ability to autonomously replicate in chloroplasts and be inherited over generations. The plasmids contained a cassette encoding the Cas9 endonuclease, a cassette for the expression of gRNA, template DNA, and a selectable marker. In this experiment, a DNA fragment carrying the GFP reporter gene flanked by sequences homologous to the Cas9 cleavage sites created by a pair of guide gRNAs was synthesized as the template. To achieve a high level of expression of the created vectors, special attention was given to the experimental selection of a strong promoter. The authors proved that the integration of exogenous donor DNA (a GFP sequence) into target sites of the genome occurred only with the introduction of an editing plasmid containing CRISPR/gRNA. Transformation efficiency reached a frequency of 10% [39].
Another successful targeted delivery of editing tools to chloroplasts via bioballistics was performed for the transformation of the tobacco plastome. The authors used Cas9 and guide RNA to increase the number of double-strand breaks in the target site, activate the DNA damage repair mechanism and promote the integration of exogenous donor DNA into the target region of the plastome. Genes encoding editing tools were delivered into chloroplasts either in the co-transformation mode or as part of an expression cassette along with target genes and a selectable marker in a single vector. The results showed that the efficiency of chloroplast transformation with this supplement increased six- to ten-fold [40].
5. Conclusions
Editing plastid genomes today is a new direction in plant bioengineering that holds good prospects. Despite the fact that the peculiarities of the plastid genome organization require the specific adaptation of genome editing methods, these methods allow for solving a very wide range of problems, including the modification of genes responsible for photosynthesis and increasing plastome transformation efficiency. One of the key factors for effective plastome editing or modification is using suitable methods for delivering editing tools and a template for homologous recombination. Some of these methods are already in active use, while others are still under development. However, plastid genome editing systems are actively developing and new methods for delivering programmable nucleases are constantly being invented. Accordingly, approaches to conducting research in this area need to be adapted with respect to new technologies that will make it possible to find a simple and effective solution for editing the genomes of plant organelles.
Author Contributions
Conceptualization, analysis of literary data, writing, N.V.D. and Y.V.S.; funding acquisition, Y.V.S. All authors have read and agreed to the published version of the manuscript.
Funding
The work was supported by grant RSF № 23-24-00545.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rashid, B.; Tariq, M.; Khalid, A.; Shams, F.; Ali, Q.; Ashraf, F.; Ghaffar, I.; Khan, M.I.; Rehman, R.; Husnain, T. Crop improvement: New approaches and modern techniques. Plant Gene Trait. 2017, 8, 18–30. [Google Scholar] [CrossRef]
- Fiaz, S.; Ahmar, S.; Saeed, S.; Riaz, A.; Mora-Poblete, F.; Jung, K.H. Evolution and application of genome editing techniques for achieving food and nutritional security. Int. J. Mol. Sci. 2021, 22, 5585. [Google Scholar] [CrossRef]
- Fichtner, F.; Urrea Castellanos, R.; Ülker, B. Precision genetic modifications: A new era in molecular biology and crop improvement. Planta 2014, 239, 921–939. [Google Scholar] [CrossRef]
- Govindan, G.; Ramalingam, S. Programmable Site-Specific Nucleases for targeted genome engineering in higher Eukaryotes. J. Cell Physiol. 2016, 231, 2380–2392. [Google Scholar] [CrossRef] [PubMed]
- Das, D.; Singha, D.L.; Paswan, R.R.; Chowdhury, N.; Sharma, M.; Reddy, P.S.; Chikkaputtaiah, C. Recent advancements in CRISPR/Cas technology for accelerated crop improvement. Planta 2022, 255, 109. [Google Scholar] [CrossRef]
- Liu, H.; Chen, W.; Li, Y.; Sun, L.; Chai, Y.; Chen, H.; Nie, H.; Huang, C. CRISPR/Cas9 technology and its utility for crop improvement. Int. J. Mol. Sci. 2022, 23, 10442. [Google Scholar] [CrossRef]
- Basu, U.; Riaz Ahmed, S.; Bhat, B.A.; Anwar, Z.; Ali, A.; Ijaz, A.; Gulzar, A.; Bibi, A.; Tyagi, A.; Nebapure, S.M.; et al. A CRISPR way for accelerating cereal crop improvement: Progress and challenges. Front. Genet. 2023, 13, 866976. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chang, L.; Zhang, J. Advancing organelle genome transformation and editing for crop improvement. Plant Commun. 2021, 4, 100141. [Google Scholar] [CrossRef]
- An, Y.; Wang, Y.; Wang, X.; Xiao, J. Development of chloroplast transformation and gene expression regulation technology in land plants. Front. Plant Sci. 2022, 13, 1037038. [Google Scholar] [CrossRef]
- Yu, Y.; Yu, P.C.; Chang, W.J.; Yu, K.; Lin, C.S. Plastid transformation: How does it work? Can it be applied to crops? What can it offer? Int. J. Mol. Sci. 2020, 21, 4854. [Google Scholar] [CrossRef]
- Bock, R. Engineering plastid genomes: Methods, tools, and applications in basic research and biotechnology. Annu. Rev. Plant Biol. 2015, 66, 211–241. [Google Scholar] [CrossRef] [PubMed]
- Rozov, S.M.; Sidorchuk, Y.V.; Deineko, E.V. Transplastomic plants: Problems of production and their solution. Russ. J. Plant Physiol. 2022, 69, 20. [Google Scholar] [CrossRef]
- Son, S.; Park, S.R. Challenges facing CRISPR/Cas9-based genome editing in plants. Front. Plant Sci. 2022, 13, 902413. [Google Scholar] [CrossRef] [PubMed]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 23, 134. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Zhu, H. Modified gene editing systems: Diverse bioengineering tools and crop improvement. Front. Plant Sci. 2022, 13, 847169. [Google Scholar] [CrossRef]
- Bracher, A.; Whitney, S.M.; Hartl, F.U.; Hayer-Hartl, M. Biogenesis and metabolic maintenance of rubisco. Annu. Rev. Plant Biol. 2017, 68, 29–60. [Google Scholar] [CrossRef]
- Khalil, A.M. The genome editing revolution: Review. J. Genet. Eng. Biotechnol. 2020, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Tariq, A.; Mushtaq, M.; Yaqoob, H.; Bhat, B.A.; Zargar, S.M.; Raza, A.; Ali, S.; Charagh, S.; Mubarik, M.S.; Zaman, Q.U.; et al. Putting CRISPR-Cas system in action: A golden window for efficient and precise genome editing for crop improvement. GM Crops Food 2023, 14, 1–27. [Google Scholar] [CrossRef]
- Sakuma, T.; Woltjen, K. Nuclease-mediated genome editing: At the front-line of functional genomics technology. Dev. Growth Differ. 2014, 56, 2–13. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Nain, V. TALENs-an indispensable tool in the era of CRISPR: A mini review. J. Genet. Eng. Biotechnol. 2021, 21, 125. [Google Scholar] [CrossRef]
- Cermak, T.; Doyle, E.L.; Christian, M.; Wang, L.; Zhang, Y.; Schmidt, C.; Baller, J.A.; Somia, N.V.; Bogdanove, A.J.; Voytas, D.F. Efcient design and assembly of custom TALEN and other TAL efector-based constructs for DNA targeting. Nucleic Acids Res. 2011, 39, e82. [Google Scholar] [CrossRef]
- Ahmad, N.; Nielsen, B.L.; Mansoor, S. Editing the plastid genome of recalcitrant plant species. Trends Genet. 2021, 37, 955–957. [Google Scholar] [CrossRef]
- Kang, B.C.; Bae, S.J.; Lee, S.; Lee, J.S.; Kim, A.; Lee, H.; Baek, G.; Seo, H.; Kim, J.; Kim, J.S. Chloroplast and mitochondrial DNA editing in plants. Nat. Plants 2021, 7, 899–905. [Google Scholar] [CrossRef]
- Mok, B.Y.; de Moraes, M.H.; Zeng, J.; Bosch, D.E.; Kotrys, A.V.; Raguram, A.; Hsu, F.; Radey, M.C.; Peterson, S.B.; Mootha, V.K.; et al. A bacterial cytidine deaminase toxin enables CRISPR-free mitochondrial base editing. Nature 2020, 583, 631–637. [Google Scholar] [CrossRef]
- Mok, Y.G.; Hong, S.; Bae, S.J.; Cho, S.I.; Kim, J.S. Targeted A-to-G base editing of chloroplast DNA in plants. Nat. Plants 2022, 8, 1378–1384. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, I.; Okuno, M.; Yamamoto, H.; Tamura, Y.; Itoh, T.; Shikanai, T.; Takanashi, H.; Tsutsumi, N.; Arimura, S.I. Targeted base editing in the plastid genome of Arabidopsis thaliana. Nat. Plants 2021, 7, 906–913. [Google Scholar] [CrossRef]
- Maliga, P. Engineering the plastid and mitochondrial genomes of flowering plants. Nat. Plants 2022, 8, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, I.; Okuno, M.; Itoh, T.; Tsutsumi, N.; Arimura, S.I. Characterization and development of a plastid genome base editor, ptpTALECD. Plant J. 2023, 115, 1151–1162. [Google Scholar] [CrossRef]
- Hille, F.; Richter, H.; Wong, S.P.; Bratovic, M.; Ressel, S.; Charpentier, E. The Biology of CRISPR-Cas: Backward and Forward. Cell 2018, 172, 1239–1259. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Jiang, F.; Taylor, D.W.; Sternberg, S.H.; Kaya, E.; Ma, E.; Anders, C.; Hauer, M.; Zhou, K.; Lin, S.; et al. Structures of Cas9 endonucleases reveal RNA-mediated conformational activation. Science 2014, 343, 1247997. [Google Scholar] [CrossRef]
- Kim, H.; Kim, J.S. A guide to genome engineering with programmable nucleases. Nat. Rev. Genet. 2014, 15, 321–334. [Google Scholar] [CrossRef]
- Razzaq, A.; Saleem, F.; Kanwal, M.; Mustafa, G.; Yousaf, S.; Imran Arshad, H.M.; Hameed, M.K.; Khan, M.S.; Joyia, F.A. Modern trends in plant genome editing: An inclusive review of the CRISPR/Cas9 toolbox. Int. J. Mol. Sci. 2019, 20, 4045. [Google Scholar] [CrossRef]
- Jasin, M.; Haber, J.E. The democratization of gene editing: Insights from site-specific cleavage and double-strand break repair. DNA Repair 2016, 44, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Luan, X.; Liu, Y.; Wang, L.; Wang, J.; Yang, S.; Liu, S.; Zhang, J.; Liu, H.; Yao, D. Strategies and methods for improving the efficiency of CRISPR/Cas9 gene editing in plant molecular breeding. Plants 2023, 12, 1478. [Google Scholar] [CrossRef] [PubMed]
- Ozyigit, I.I.; Yucebilgili Kurtoglu, K. Particle bombardment technology and its applications in plants. Mol. Biol. Rep. 2020, 47, 9831–9847. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.U.; Ling, A.P.K. Gene introduction approaches in chloroplast transformation and its applications. J. Genet. Eng. Biotechnol. 2021, 19, 148. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Nannas, N.J.; Fu, F.F.; Shi, J.; Aspinwall, B.; Parrott, W.A.; Dawe, R.K. Genome-scale sequence disruption following biolistic transformation in rice and maize. Plant Cell 2019, 31, 368–383. [Google Scholar] [CrossRef] [PubMed]
- Yoo, B.C.; Yadav, N.S.; Orozco, E.M., Jr.; Sakai, H. Cas9/gRNA-mediated genome editing of yeast mitochondria and Chlamydomonas chloroplasts. PeerJ 2020, 8, e8362. [Google Scholar] [CrossRef]
- Tang, N.; Xia, Y.; Zhan, Y.; Dan, J.; Yu, M.; Bu, X.; Cao, M. Improvement of chloroplast transformation using CRISPR/Cas9. J. Biobased Mater. Bioenergy 2020, 14, 401. [Google Scholar] [CrossRef]
- Glass, Z.; Lee, M.; Li, Y.; Xu, Q. Engineering the delivery system for CRISPR-based genome editing. Trends Biotechnol. 2017, 36, 173–185. [Google Scholar] [CrossRef]
- Tian, L.; Chou, H.L.; Fukuda, M.; Kumamaru, T.; Okita, T.W. mRNA Localization in Plant Cells. Plant Physiol. 2020, 182, 97–109. [Google Scholar] [CrossRef]
- Gómez, G.; Pallás, V. Noncoding RNA mediated traffic of foreign mRNA into chloroplasts reveals a novel signaling mechanism in plants. PLoS ONE 2010, 19, e12269. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.F.; Huang, Y.P.; Chen, L.H.; Hsu, Y.H.; Tsai, C.H. Chloroplast phosphoglycerate kinase is involved in the targeting of Bamboo mosaic virus to chloroplasts in Nicotiana benthamiana plants. Plant Physiol. 2013, 3, 1598–1608. [Google Scholar] [CrossRef] [PubMed]
- Newkirk, G.M.; de Allende, P.; Jinkerson, R.E.; Giraldo, J.P. Nanotechnology approaches for chloroplast biotechnology advancements. Front. Plant Sci. 2021, 12, 691295. [Google Scholar] [CrossRef] [PubMed]
- Santana, I.; Hu, P.; Jeon, S.J.; Castillo, C.; Tu, H.; Giraldo, J.P. Peptide-mediated targeting of nanoparticles with chemical cargoes to chloroplasts in Arabidopsis plants. Bio-Protocol 2021, 20, e4060. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.Y.; Lew, T.T.S.; Sweeney, C.J.; Koman, V.B.; Wong, M.H.; Bohmert-Tatarev, K.; Snell, K.D.; Seo, J.S.; Chua, N.H.; Strano, M.S. Chloroplast-selective gene delivery and expression in planta using chitosan-complexed single-walled carbon nanotube carriers. Nat. Nanotechnol. 2019, 14, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Odahara, M.; Miyamoto, T.; Numata, K. Fusion peptide-based biomacromolecule delivery system for plant cells. ACS Biomater. Sci. Eng. 2021, 7, 2246–2254. [Google Scholar] [CrossRef]
- Liu, B.R.; Chen, C.W.; Huang, Y.W.; Lee, H.J. Cell-penetrating peptides for use in development of transgenic plants. Molecules 2023, 28, 3367. [Google Scholar] [CrossRef]
- Yoshizumi, T.; Oikawa, K.; Chuah, J.A.; Kodama, Y.; Numata, K. Selective gene delivery for integrating exogenous DNA into plastid and mitochondrial genomes using peptide-DNA complexes. Biomacromolecules 2018, 19, 1582–1591. [Google Scholar] [CrossRef]
- Thagun, C.; Chuah, J.A.; Numata, K. Targeted gene delivery into various plastids mediated by clustered cell-penetrating and chloroplast-targeting eptides. Adv. Sci. 2019, 6, 1902064. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, K.; Tateishi, A.; Odahara, M.; Kodama, Y.; Numata, K. Imaging of the entry pathway of a cell-penetrating peptide-DNA complex from the extracellular space to chloroplast nucleoids across multiple membranes in Arabidopsis leaves. Front. Plant Sci. 2021, 12, 759871. [Google Scholar] [CrossRef] [PubMed]
- Odahara, M.; Horii, Y.; Itami, J.; Watanabe, K.; Numata, K. Functional peptide-mediated plastid transformation in tobacco, rice, and kenaf. Front. Plant Sci. 2022, 13, 989310. [Google Scholar] [CrossRef]
- Lee, H.; Lee, S.; Baek, G.; Kim, A.; Kang, B.-C.; Seo, H.; Kim, J.-S. Mitochondrial DNA editing in mice with DddA-TALE fusion deaminases. Nat. Commun. 2021, 12, 1190. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Char, S.N.; Liu, B.; Liu, H.; Li, X.; Yang, B. High-efficiency plastome base editing in rice with TAL cytosine deaminase. Mol. Plant. 2021, 6, 1412–1414. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram of editing tools’ delivery to chloroplasts for the case of nuclear transformation. CH—chloroplast; CTP—chloroplast transit peptide; ZFN—zinc-finger nuclease; TALEN—transcription activator-like effector nucleases; FokI—catalytic subunit of the bacterial endonuclease; Cas9—CRISPR associated protein.
Figure 1.
Schematic diagram of editing tools’ delivery to chloroplasts for the case of nuclear transformation. CH—chloroplast; CTP—chloroplast transit peptide; ZFN—zinc-finger nuclease; TALEN—transcription activator-like effector nucleases; FokI—catalytic subunit of the bacterial endonuclease; Cas9—CRISPR associated protein.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dorogova, N.V.; Sidorchuk, Y.V. The Search of a Molecular “Swiss Knife” for Chloroplast Genomic Editing. Horticulturae 2023, 9, 1338. https://doi.org/10.3390/horticulturae9121338
AMA Style
Dorogova NV, Sidorchuk YV. The Search of a Molecular “Swiss Knife” for Chloroplast Genomic Editing. Horticulturae. 2023; 9(12):1338. https://doi.org/10.3390/horticulturae9121338
Chicago/Turabian StyleDorogova, Natalya V., and Yuriy V. Sidorchuk. 2023. "The Search of a Molecular “Swiss Knife” for Chloroplast Genomic Editing" Horticulturae 9, no. 12: 1338. https://doi.org/10.3390/horticulturae9121338
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.