
The Postmedieval Latvian Oral Microbiome in the Context of Modern Dental Calculus and Modern Dental Plaque Microbial Profiles

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Archaeological Sample Characteristics, Collection and Burial Site Data
2.2. Modern Dental Calculus and Modern Dental Plaque Sample Collection
2.3. DNA Isolation
2.4. Library Preparation and Shotgun Metagenomics Sequencing
2.5. Sequencing Data Analysis
2.6. Statistical Analysis
3. Results
3.1. Sequencing Data and Ancient DNA Authentication
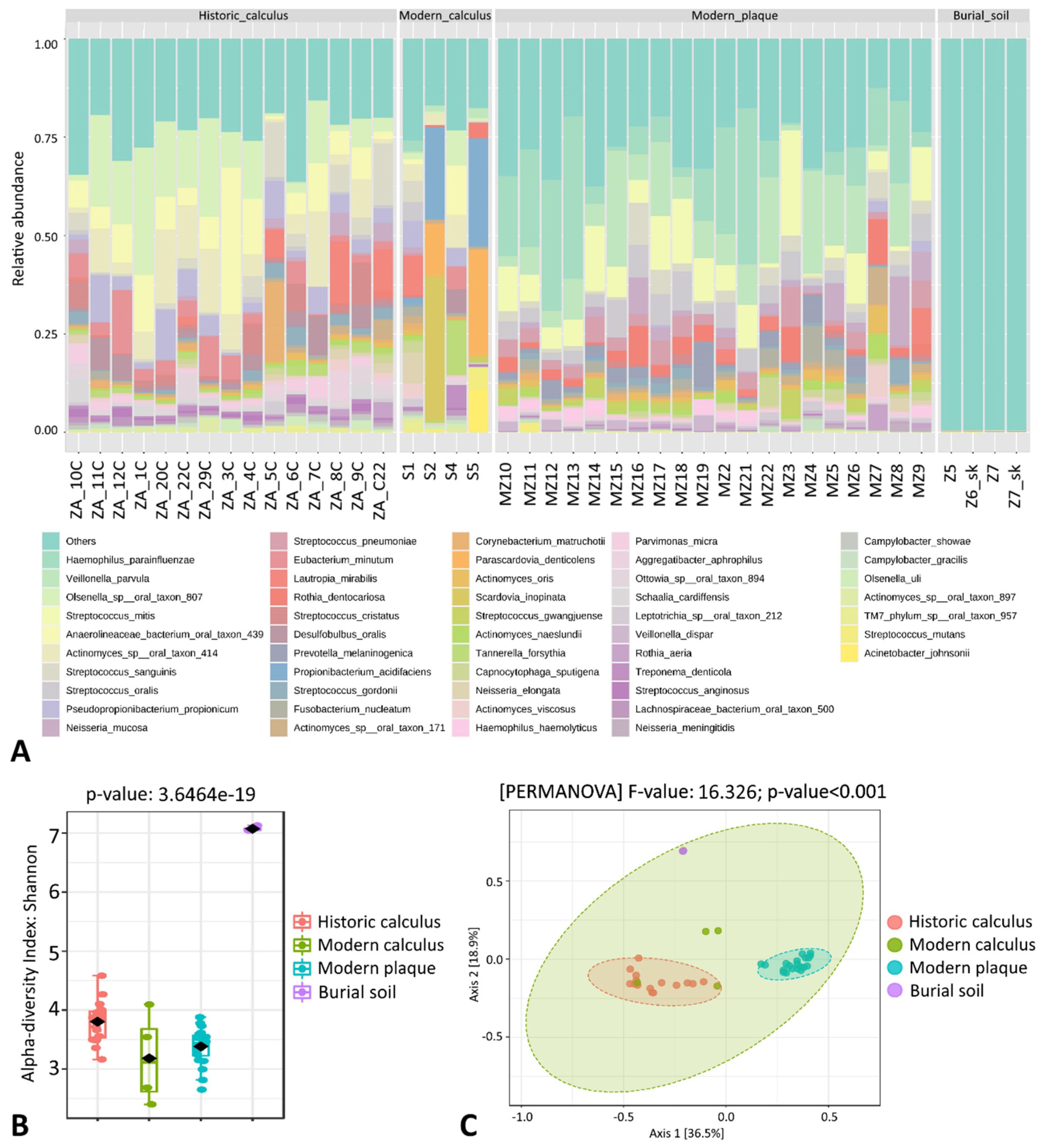
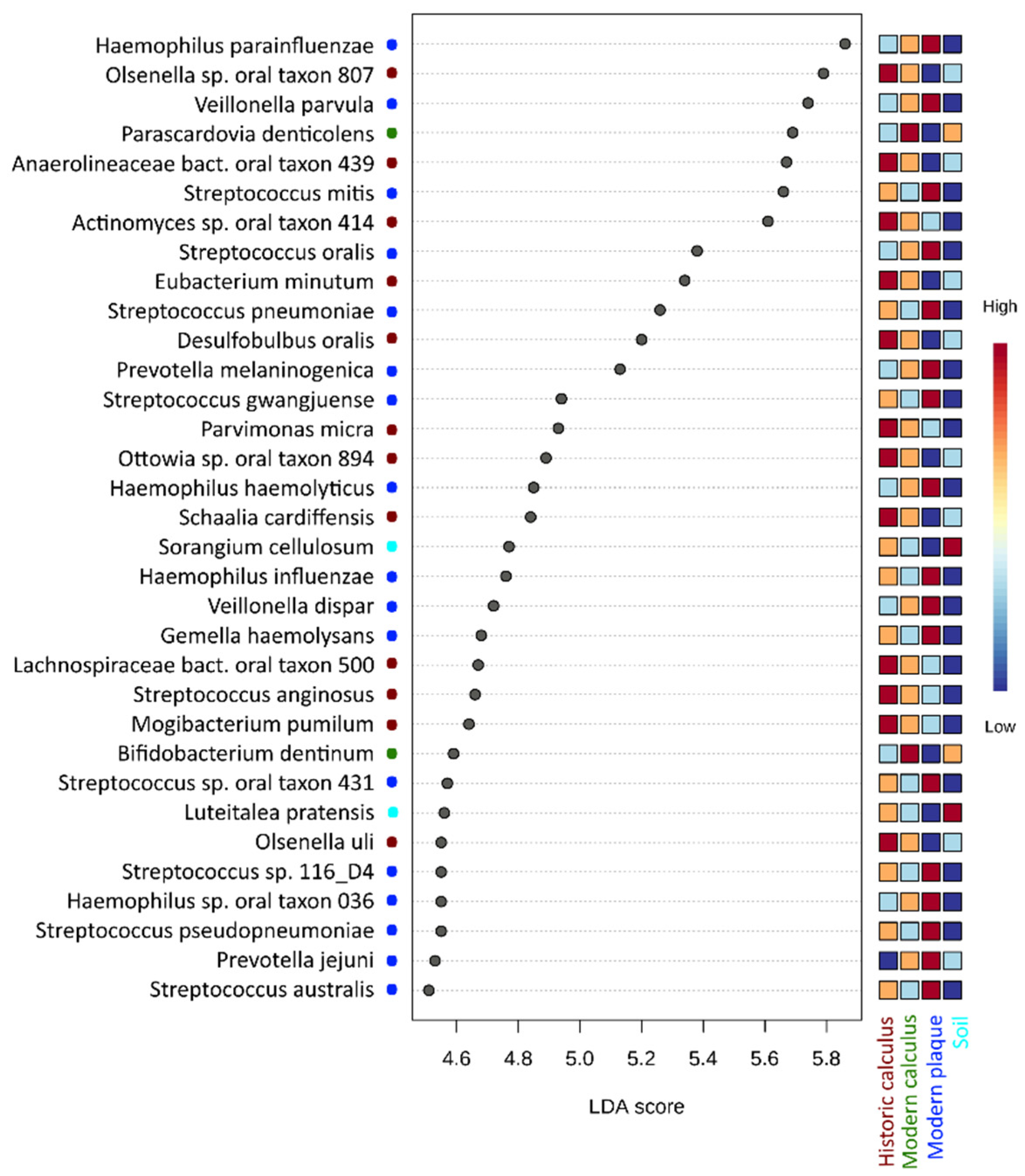
3.2. Taxonomical Analysis of Microbial Profiles at the Species Level
3.3. Taxonomic Analysis of Microbial Profiles of Historic Dental Calculus Samples at the Species Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.R.; Yu, W.-H.; Lakshmanan, A.; Wade, W.G. The Human Oral Microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilian, M.; Chapple, I.L.C.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.L.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome—An update for oral healthcare professionals. Br. Dent. J. 2016, 221, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Verma, D.; Garg, P.K.; Dubey, A.K. Insights into the human oral microbiome. Arch. Microbiol. 2018, 200, 525–540. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.N.; Snesrud, E.; Liu, J.; Ong, A.C.; Kilian, M.; Schork, N.J.; Bretz, W. The Dental Plaque Microbiome in Health and Disease. PLoS ONE 2013, 8, e58487. [Google Scholar] [CrossRef] [Green Version]
- Warinner, C.; Rodrigues, J.F.M.; Vyas, R.; Trachsel, C.; Shved, N.; Grossmann, J.; Radini, A.; Hancock, Y.; Tito, R.Y.; Fiddyment, S.; et al. Pathogens and host immunity in the ancient human oral cavity. Nat. Genet. 2014, 46, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Faller, L.L.; Klitgord, N.; Mazumdar, V.; Ghodsi, M.; Sommer, D.D.; Gibbons, T.R.; Treangen, T.J.; Chang, Y.-C.; Li, S.; et al. Deep Sequencing of the Oral Microbiome Reveals Signatures of Periodontal Disease. PLoS ONE 2012, 7, e37919. [Google Scholar] [CrossRef] [Green Version]
- Ziesemer, K.A.; Mann, A.E.; Sankaranarayanan, K.; Schroeder, H.; Ozga, A.T.; Brandt, B.W.; Zaura, E.; Waters-Rist, A.; Hoogland, M.; Salazar-García, D.C.; et al. Intrinsic challenges in ancient microbiome reconstruction using 16S rRNA gene amplification. Sci. Rep. 2015, 5, 16498. [Google Scholar] [CrossRef]
- Velsko, I.M.; Fellows Yates, J.A.; Aron, F.; Hagan, R.W.; Frantz, L.A.F.; Loe, L.; Martinez, J.B.R.; Chaves, E.; Gosden, C.; Larson, G.; et al. Microbial differences between dental plaque and historic dental calculus are related to oral biofilm maturation stage. Microbiome 2019, 7, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogden, A. Advances in the palaeopahtology of teeth and jaws. In Advances in Human Palaeopathology; Pinhasi, R., Mays, S., Eds.; Wiley: New York, NY, USA, 2008; pp. 283–307. [Google Scholar]
- Spirģis, R. Rīgas Sv. Pētera baznīcas kapsētas apbedījumu stratigrāfija (Stratigraphy of the Riga’s St. Peter’s Church cemetery burial). In Senā Rīga 7: Pētījumi Pilsētas Arheoloģijā Un Vēsturē; Latvijas vēstures institūta apgāds: Rīga, Latvia, 2012; pp. 179–225. (In Latvian) [Google Scholar]
- Tilko, S. Apbedīšanas tradīcijas Doma viduslaiku kapsētā (Mortuarypractises in the Dome Medieval Cemetery). In Senā Rīga 1: Pētījumi Pilsētas Arheoloģijā Un Vēsturē; Latvijas vēstures institūta apgāds: Rīga, Latvia, 1998; pp. 273–288. (In Latvian) [Google Scholar]
- Brooks, S.; Suchey, J.M. Skeletal age determination based on the os pubis: A comparison of the Acsádi-Nemeskéri and Suchey-Brooks methods. Hum. Evol. 1990, 5, 227–238. [Google Scholar] [CrossRef]
- Buckberry, J.L.; Chamberlain, A.T. Age estimation from the auricular surface of the ilium: A revised method. Am. J. Phys. Anthr. 2002, 119, 231–239. [Google Scholar] [CrossRef]
- Fulton, T.L.; Shapiro, B. Setting Up an Ancient DNA Laboratory. In Ancient DNA; Shapiro, B., Barlow, A., Heintzman, P.D., Hofreiter, M., Paijmans, J.L.A., Soares, A.E.R., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; Volume 1963, pp. 1–13. ISBN 978-1-4939-9175-4. [Google Scholar]
- Donoghue, H.D. Molecular Palaeopathology of Human Infectious Disease. In Advances in Human Palaeopathology; Pinhasi, R., Mays, S., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2007; pp. 147–176. ISBN 978-0-470-72418-7. [Google Scholar]
- Kazarina, A.; Gerhards, G.; Petersone-Gordina, E.; Kimsis, J.; Pole, I.; Zole, E.; Leonova, V.; Ranka, R. Analysis of the bacterial communities in ancient human bones and burial soil samples: Tracing the impact of environmental bacteria. J. Archaeol. Sci. 2019, 109, 104989. [Google Scholar] [CrossRef]
- Keyser-Tracqui, C.; Ludes, B. Methods for the Study of Ancient DNA. In Forensic DNA Typing Protocols; Humana Press: Totowa, NJ, USA, 2004; Volume 297, pp. 253–264. ISBN 978-1-59259-867-0. [Google Scholar]
- Key, F.M.; Posth, C.; Krause, J.; Herbig, A.; Bos, K.I. Mining Metagenomic Data Sets for Ancient DNA: Recommended Protocols for Authentication. Trends Genet. 2017, 33, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Dabney, J.; Meyer, M.; Paabo, S. Ancient DNA Damage. Cold Spring Harb. Perspect. Biol. 2013, 5, a012567. [Google Scholar] [CrossRef] [PubMed]
- Paabo, S. Ancient DNA: Extraction, characterization, molecular cloning, and enzymatic amplification. Proc. Natl. Acad. Sci. USA 1989, 86, 1939–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goecks, J.; Nekrutenko, A.; Taylor, J.; The Galaxy Team. Galaxy: A comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 2010, 11, R86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosakovsky Pond, S.; Wadhawan, S.; Chiaromonte, F.; Ananda, G.; Chung, W.-Y.; Taylor, J.; Nekrutenko, A.; The Galaxy Team. Windshield splatter analysis with the Galaxy metagenomic pipeline. Genome Res. 2009, 19, 2144–2153. [Google Scholar] [CrossRef] [Green Version]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Breitwieser, F.P.; Thielen, P.; Salzberg, S.L. Bracken: Estimating species abundance in metagenomics data. PeerJ Comput. Sci. 2017, 3, e104. [Google Scholar] [CrossRef]
- Pavian: Interactive Analysis of Metagenomics Data for Microbiomics and Pathogen Identification|bioRxiv. Available online: https://www.biorxiv.org/content/10.1101/084715v1 (accessed on 11 June 2020).
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Knights, D.; Kuczynski, J.; Charlson, E.S.; Zaneveld, J.; Mozer, M.C.; Collman, R.G.; Bushman, F.D.; Knight, R.T.; Kelley, S.T. Bayesian community-wide culture-independent microbial source tracking. Nat. Methods 2011, 8, 761–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbig, A.; Maixner, F.; Bos, K.I.; Zink, A.; Krause, J.; Huson, D.H. MALT: Fast alignment and analysis of metagenomic DNA sequence data applied to the Tyrolean Iceman. bioRxiv 2017, 050559. [Google Scholar] [CrossRef] [Green Version]
- Jónsson, H.; Ginolhac, A.; Schubert, M.; Johnson, P.L.F.; Orlando, L. mapDamage2.0: Fast approximate Bayesian estimates of ancient DNA damage parameters. Bioinformatics 2013, 29, 1682–1684. [Google Scholar] [CrossRef] [PubMed]
- Peltzer, A.; Jäger, G.; Herbig, A.; Seitz, A.; Kniep, C.; Krause, J.; Nieselt, K. EAGER: Efficient ancient genome reconstruction. Genome Biol. 2016, 17, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance: Non-parametric manova for ecology. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Weyrich, L.S.; Farrer, A.G.; Eisenhofer, R.; Arriola, L.A.; Young, J.; Selway, C.A.; Handsley-Davis, M.; Adler, C.J.; Breen, J.; Cooper, A. Laboratory contamination over time during low-biomass sample analysis. Mol. Ecol. Resour. 2019, 19, 982–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol. 2014, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Socransky, S.; Haffajee, A.; Cugini, M.; Smith, C.; Kent, R.L. Microbial complexes in subgingival plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Rodriguez, T.M.; Fornaciari; Luciani, S.; Marota, I.; Vercellotti, G.; Toranzos, G.A.; Giuffra, V.; Cano, R.J. Commensal and Pathogenic Members of the Dental Calculus Microbiome of Badia Pozzeveri Individuals from the 11th to 19th Centuries. Genes 2019, 10, 299. [Google Scholar] [CrossRef] [Green Version]
- Nord, C.-E.; Sjöberg, L.; Wadström, T. Pseudomonas Aeruginosa in Oral Infections. Acta Odontol. Scand. 1972, 30, 371–381. [Google Scholar] [CrossRef]
- Souto, R.; Silva-Boghossian, C.M.; Colombo, A.P.V. Prevalence of Pseudomonas aeruginosa and Acinetobacter spp. in subgingival biofilm and saliva of subjects with chronic periodontal infection. Braz. J. Microbiol. 2014, 45, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Blum, W.E.H.; Zechmeister-Boltenstern, S.; Keiblinger, K.M. Does Soil Contribute to the Human Gut Microbiome? Microorganisms 2019, 7, 287. [Google Scholar] [CrossRef] [Green Version]
- Petersone-Gordina, E.; Roberts, C.; Millard, A.R.; Montgomery, J.; Gerhards, G. Dental disease and dietary isotopes of individuals from St Gertrude Church cemetery, Riga, Latvia. PLoS ONE 2018, 13, e0191757. [Google Scholar] [CrossRef] [Green Version]
- Eisenhofer, R.; Kanzawa-Kiriyama, H.; Shinoda, K.-I.; Weyrich, L.S. Investigating the demographic history of Japan using ancient oral microbiota. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190578. [Google Scholar] [CrossRef] [PubMed]
- Cross, K.L.; Chirania, P.; Xiong, W.; Beall, C.J.; Elkins, J.G.; Giannone, R.J.; Griffen, A.L.; Guss, A.M.; Hettich, R.L.; Joshi, S.S.; et al. Insights into the Evolution of Host Association through the Isolation and Characterization of a Novel Human Periodontal Pathobiont, Desulfobulbus oralis. mBio 2018, 9, e02061-17. [Google Scholar] [CrossRef] [Green Version]
- Raitapuro-Murray, T.; Molleson, T.I.; Hughes, F.J. The prevalence of periodontal disease in a Romano-British population c. 200–400 AD. Br. Dent. J. 2014, 217, 459–466. [Google Scholar] [CrossRef]
- Proctor, D.M.; Fukuyama, J.A.; Loomer, P.M.; Armitage, G.C.; Lee, S.A.; Davis, N.M.; Ryder, M.I.; Holmes, S.P.; Relman, D.A. A spatial gradient of bacterial diversity in the human oral cavity shaped by salivary flow. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Farrer, A.G.; Bekvalac, J.; Redfern, R.; Gully, N.; Dobney, K.; Cooper, A.; Weyrich, L. Biological and cultural drivers of oral microbiota in Medieval and Post-Medieval London, UK. bioRxiv 2018, 343889. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample ID | City | Cemetery | Century, AD | Year of Excavation | Age (Years) | Sex | Lifestyle | Possible Diseases | Tooth Type |
|---|---|---|---|---|---|---|---|---|---|
| ZA_1C | Riga | Dom Square | 16–17th | 1986 | 30–40 | Male | Town/city, middle classes | Inflammation of the clavicle | Molar |
| ZA_3C | Riga | Dom Square | 16–17th | 1986 | 45–50 | Female | Town/city, middle classes | Caries | Incisor |
| ZA_4C | Riga | St. Peter’s Church | 16–17th | 2004 | 30–35 | Male | Town/city | - | Incisor |
| ZA_5C | Riga | St. Peter’s Church | 16–17th | 2004 | 25–30 | Male | Town/city | Fractured rib and left arm’s fracture | Incisor |
| ZA_6C | Riga | St. Peter’s Church | 16–17th | 2004 | 20–25 | Male | Town/city | Non-specific inflammation of the lower leg bone surface | Premolar |
| ZA_7C | Riga | St. Gertrude’s Church | 16–17th | 2006 | 14–15 | Unknown | Countryside, commuter town | - | Incisor |
| ZA_8C | Riga | St. Gertrude’s Church | 17th | 2006 | 45–50 | Male | Countryside, commuter town | Deforming arthrosis (joints) | Incisor |
| ZA_9C | Riga | St. Gertrude’s Church | 16–17th | 2006 | 25–30 | Male | Countryside, commuter town | Vertebral fracture with local inflammation | Molar |
| ZA_10C | Riga | St. Gertrude’s Church | 16–17th | 2006 | 55–60 | Male | Countryside, commuter town | Caries, tooth root abscess, arthritis in joints | Molar |
| ZA_11C | Cesis | St. John’s Church | 17th | 2015 | 35–40 | Male | Town/city, aristocracy | Multiple tooth decay, toe osteomyelitis | Incisor |
| ZA_12C | Cesis | St. John’s Church | 17th | 2015 | 35–40 | Female | Town/city, aristocracy | Non-specific inflammatory process on the surface of the leg bones | Premolar |
| ZA_20C | Kuldiga | Church of the Holy Trinity, Roman Catholic Church | 16–17th | 2015 | 45–50 | Female | Town/city, lower classes | - | Molar |
| ZA_22C | Kuldiga | Church of the Holy Trinity, Roman Catholic Church | 16–17th | 2015 | 30–35 | Female | Town/city, lower classes | - | Incisor |
| ZA_29C | Jelgava | St. Trinity’s Church | 16–17th | 2009 | 35–40 | Female | Town/city, aristocracy | Caries, arthritis in joints, non-specific inflammation | Premolar |
| ZA_C22 | Riga | St. Gertrude’s Church | 17th | 2006 | 40–50 | Unknown | Countryside, commuter town | Deforming arthrosis, pelvic joint | Incisor |
| Sample Type | Sex | Teeth Health * | Age (Years) ** | |
|---|---|---|---|---|
| MZ2 | Supragingival dental plaque | Male | healthy | 23 |
| MZ3 | Supragingival dental plaque | Male | treated | 23 |
| MZ4 | Supragingival dental plaque | Male | treated | 28 |
| MZ5 | Supragingival dental plaque | Female | treated | 23 |
| MZ6 | Supragingival dental plaque | Male | treated | 24 |
| MZ7 | Supragingival dental plaque | Female | treated | 24 |
| MZ8 | Supragingival dental plaque | Male | treated | 24 |
| MZ9 | Supragingival dental plaque | Male | treated | 22 |
| MZ10 | Supragingival dental plaque | Female | healthy | 23 |
| MZ11 | Supragingival dental plaque | Female | treated | 22 |
| MZ12 | Supragingival dental plaque | Female | treated | 22 |
| MZ13 | Supragingival dental plaque | Female | healthy | 22 |
| MZ14 | Supragingival dental plaque | Female | treated | 23 |
| MZ15 | Supragingival dental plaque | Female | treated | 23 |
| MZ16 | Supragingival dental plaque | Female | treated | 22 |
| MZ17 | Supragingival dental plaque | Female | treated | 22 |
| MZ18 | Supragingival dental plaque | Female | treated | 19 |
| MZ19 | Supragingival dental plaque | Male | treated | 22 |
| MZ21 | Supragingival dental plaque | Female | healthy | 23 |
| MZ22 | Supragingival dental plaque | Female | healthy | 20 |
| S1 | Supragingival dental calculus | Male | treated | N/A |
| S2 | Supragingival dental calculus | Male | treated | N/A |
| S4 | Supragingival dental calculus | Female | treated | N/A |
| S5 | Supragingival dental calculus | Male | treated | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazarina, A.; Petersone-Gordina, E.; Kimsis, J.; Kuzmicka, J.; Zayakin, P.; Griškjans, Ž.; Gerhards, G.; Ranka, R. The Postmedieval Latvian Oral Microbiome in the Context of Modern Dental Calculus and Modern Dental Plaque Microbial Profiles. Genes 2021, 12, 309. https://doi.org/10.3390/genes12020309
Kazarina A, Petersone-Gordina E, Kimsis J, Kuzmicka J, Zayakin P, Griškjans Ž, Gerhards G, Ranka R. The Postmedieval Latvian Oral Microbiome in the Context of Modern Dental Calculus and Modern Dental Plaque Microbial Profiles. Genes. 2021; 12(2):309. https://doi.org/10.3390/genes12020309
Chicago/Turabian StyleKazarina, Alisa, Elina Petersone-Gordina, Janis Kimsis, Jevgenija Kuzmicka, Pawel Zayakin, Žans Griškjans, Guntis Gerhards, and Renate Ranka. 2021. "The Postmedieval Latvian Oral Microbiome in the Context of Modern Dental Calculus and Modern Dental Plaque Microbial Profiles" Genes 12, no. 2: 309. https://doi.org/10.3390/genes12020309