An Independent Locus Upstream of ASIP Controls Variation in the Shade of the Bay Coat Colour in Horses

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Cohort
2.2. Genotyping
2.3. Study-Specific Genotyping Quality Control (QC)
2.4. Association Analysis
2.5. Gene Annotation
2.6. Genotyping of RALY Indel Polymorphism
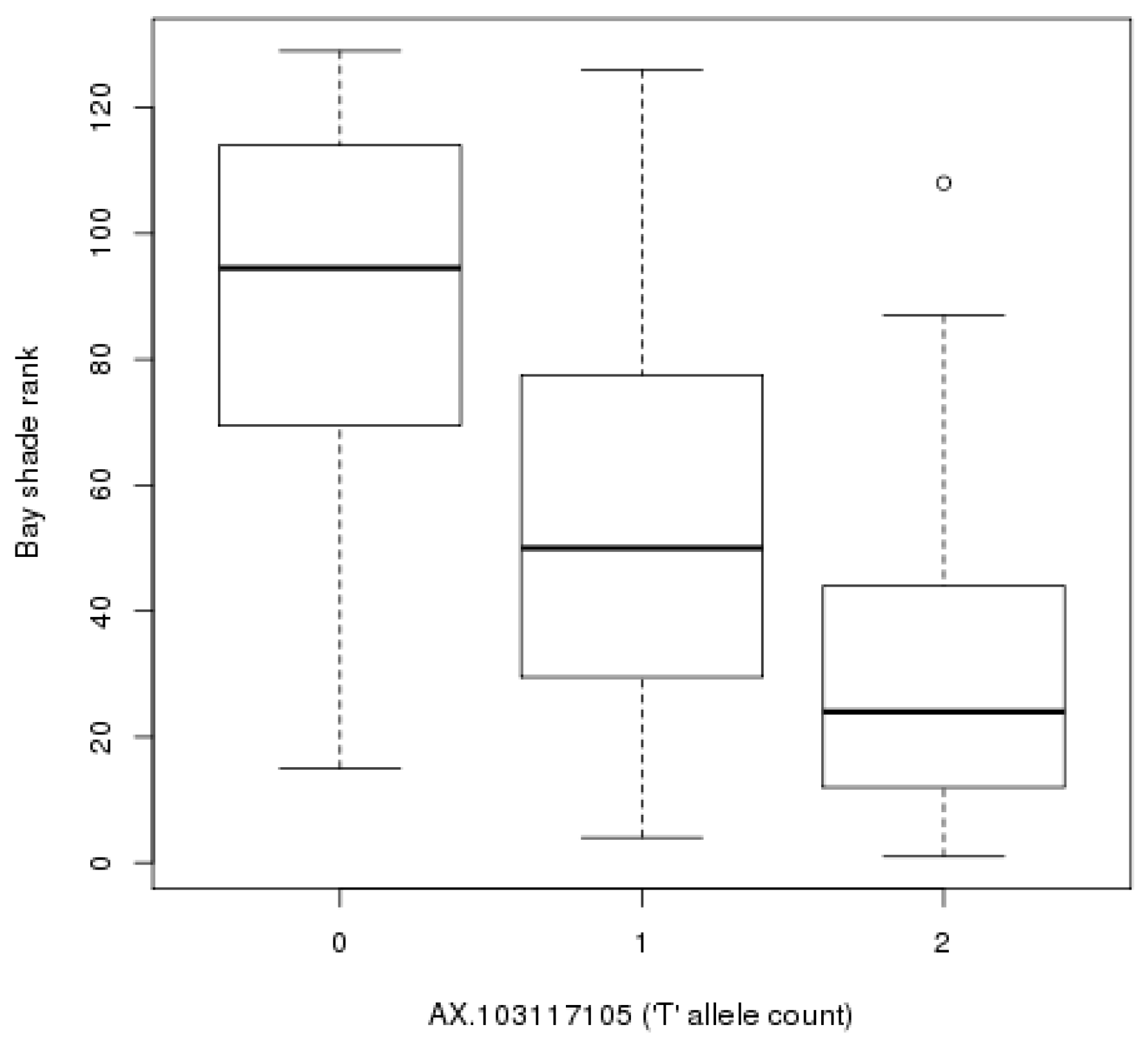
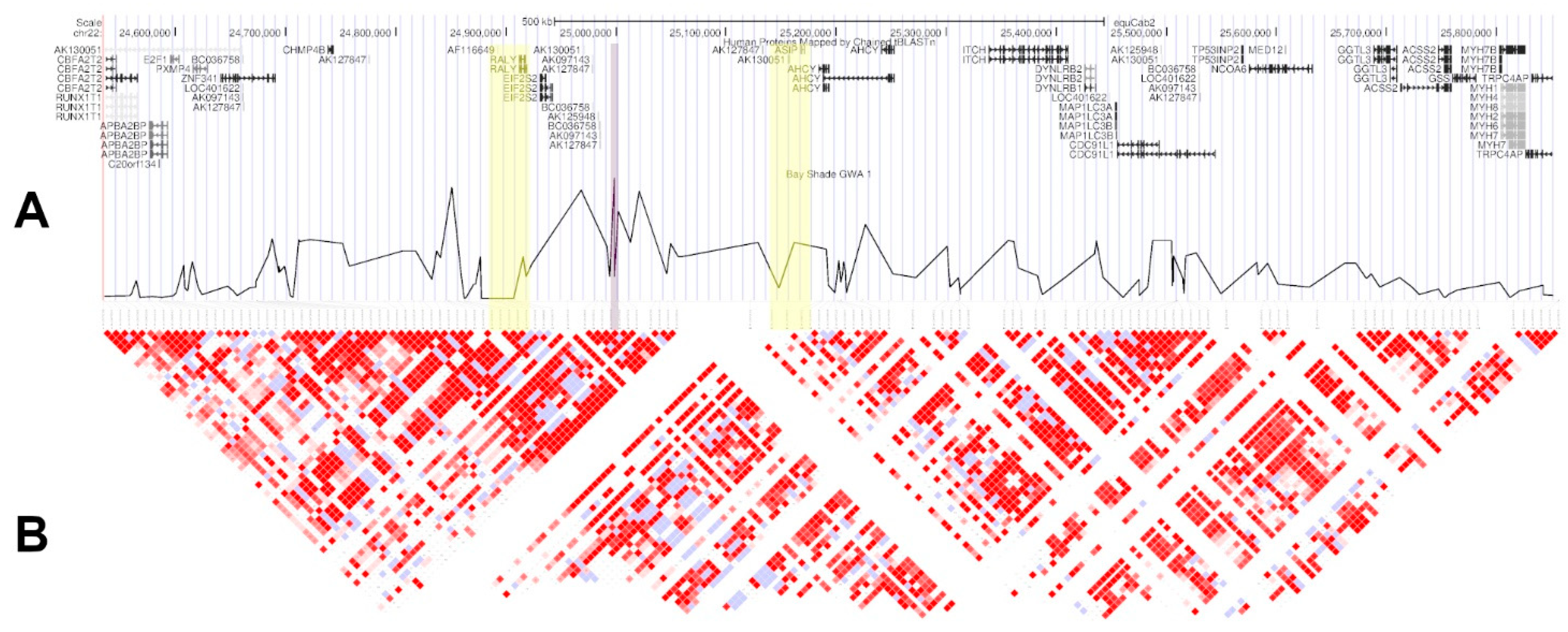
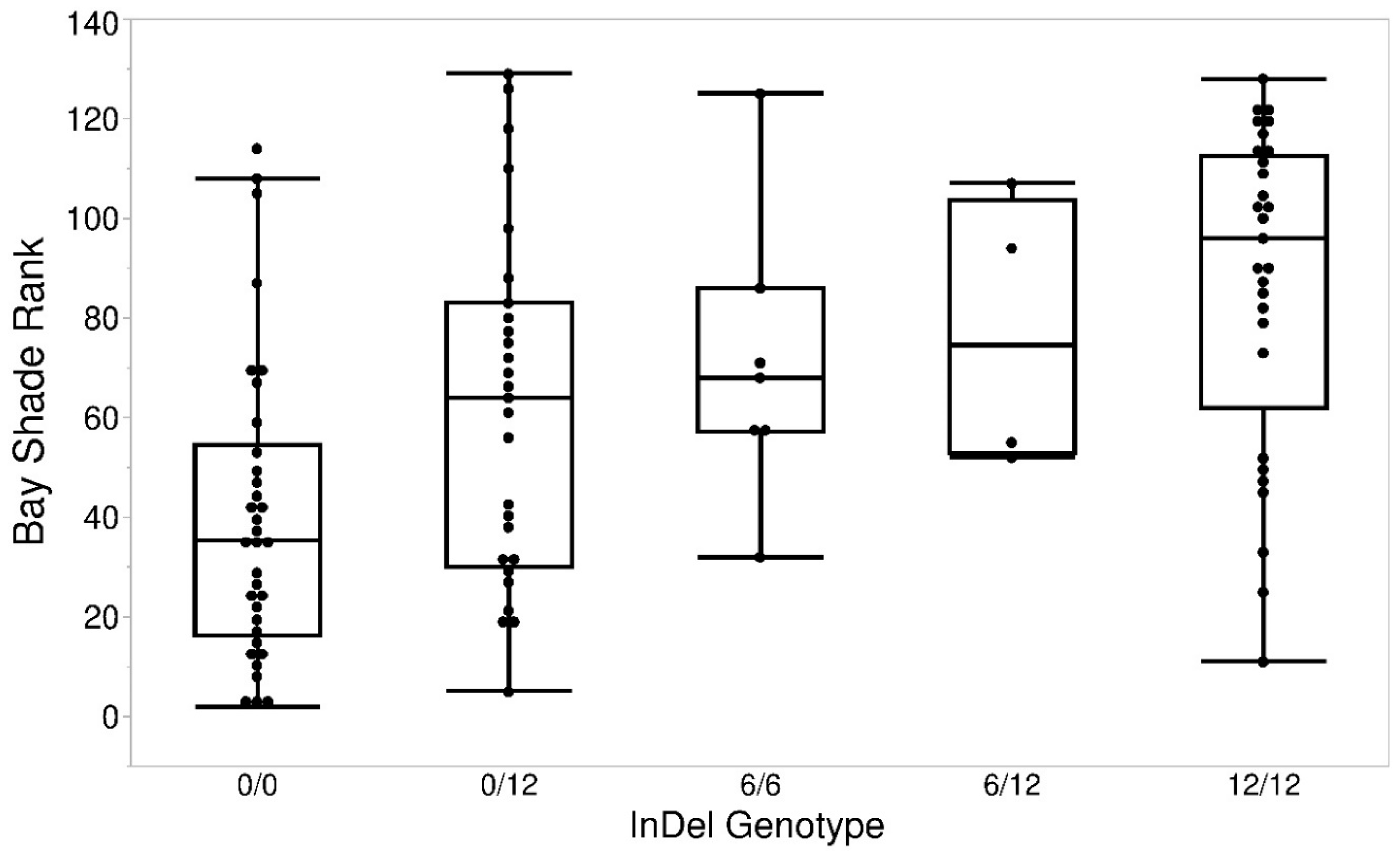
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klungland, H.; Vage, D. Molecular Genetics of Pigmentation in Domestic Animals. Curr. Genom. 2000, 1, 223–242. [Google Scholar] [CrossRef]
- Rieder, S. Molecular tests for coat colours in horses. J. Anim. Breed. Genet. 2009, 126, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Seiji, M.; Shimao, K.; Birbeck, M.S.C.; Fitzpatrick, T.B. Subcellular Localisation of Melanin Biosynthesis. Ann. N. Y. Acad. Sci. 1963, 100, 497–533. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.; Pruvost, M.; Reissmann, M.; Benecke, N.; Brockmann, G.A.; Castaños, P.; Cieslak, M.; Lippold, S.; Llorente, L.; Malaspinas, A.S.; et al. Coat color variation at the beginning of Horse domestication. Science 2009, 324, 485. [Google Scholar] [CrossRef] [Green Version]
- Fang, M.; Larson, G.; Soares Ribeiro, H.; Li, N.; Andersson, L. Contrasting Mode of Evolution at a Coat Color Locus in Wild and Domestic Pigs. PLoS Genet. 2009, 5, e1000341. [Google Scholar] [CrossRef] [Green Version]
- Tobin, D.J. Human hair pigmentation–Biological aspects. Int. J. Cosmet. Sci. 2008, 30, 233–257. [Google Scholar] [CrossRef]
- Prota, G. Recent advances in the chemistry of melanogenesis in mammals. J. Investig. Dermatol. 1980, 75, 122–127. [Google Scholar] [CrossRef] [Green Version]
- Rieder, S.; Taourit, S.; Mariat, D.; Langlois, B.; Guérin, G. Mutations in the agouti (ASIP), the extension (MCIR), and the brown (TYRP1) loci and their association to coat color phenotypes in horses (Equus caballus). Mamm. Genome 2001, 12, 450–455. [Google Scholar] [CrossRef]
- Dreger, D. Gene Interactions with Agouti Signaling Protein Produce Complex Pigmentation Phenotypes in the Domestic Dog. Doctoral Dissertation, University of Saskatchewan, Saskatoon, SK, Canada, 2012. [Google Scholar]
- Daverio, M.S.; Rigalt, F.; Romero, S.; Vidal-Rioja, L.; Di Rocco, F. Polymorphisms in MC1R and ASIP genes and their association with coat color phenotypes in llamas (Lama glama). Small Rumin. Res. 2016, 144, 83–89. [Google Scholar] [CrossRef]
- Marklund, L.; Moller, M.J.; Sandberg, K.; Andersson, L. A missense mutation in the gene for melanocyte-stimulating hormone receptor (MC1R) is associated with the chestnut coat color in horses. Mamm. Genome 1996, 7, 895–899. [Google Scholar] [CrossRef]
- Wagner, H.J.; Reissmann, M. New polymorphism detected in the horse MC1R gene. Anim. Genet. 2000, 31, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Patterson Rosa, L.; Walker, N.L.; Mallicote, M.M.; MacKay, R.J.; Brooks, S.A. Genomics of Congenital Idiopathic Anhidrosis in the Stock-Type Horse. Equine Vet. J. 2020. in review. [Google Scholar]
- Cosgrove, E.J.; Sadeghi, R.; Schlamp, F.; Holl, H.M.; Moradi-Shahrbabak, M.; Miraei-Ashtiani, S.R.; Abdalla, S.; Shykind, B.; Troedsson, M.; Stefaniuk-Szmukier, M.; et al. Genome diversity and the origin of the Arabian horse. Sci. Rep. 2020, in press. [Google Scholar]
- Schaefer, R.J.; Schubert, M.; Bailey, E.; Bannasch, D.L.; Barrey, E.; Bar-Gal, G.K.; Brem, G.; Brooks, S.A.; Distl, O.; Fries, R.; et al. Developing a 670k genotyping array to tag ~2M SNPs across 24 horse breeds. BMC Genom. 2017, 18, 565. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Chow, C.; Tellier, L.; Vattikuti, S.; Purcell, S.; Lee, J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- PLINK v1.90.b4.1 2017. Available online: https://www.cog-genomics.org/plink/1.9/ (accessed on 30 March 2017).
- Wigginton, J.E.; Cutler, D.J.; Abecasis, G.R. A note on exact tests of Hardy-Weinberg equilibrium. Am. J. Hum. Genet. 2005, 76, 887–893. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A Tool for Genome-wide Complex Trait Analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Abdellaoui, A.; Hottenga, J.J.; De Knijff, P.; Nivard, M.G.; Xiao, X.; Scheet, P.; Brooks, A.; Ehli, E.A.; Hu, Y.; Davies, G.E.; et al. Population structure, migration, and diversifying selection in the Netherlands. Eur. J. Hum. Genet. 2013, 21, 1277–1285. [Google Scholar] [CrossRef] [Green Version]
- Shang, S.; Yu, Y.; Zhao, Y.; Dang, W.; Zhang, J.; Qin, X.; Irwin, D.M.; Wang, Q.; Liu, F.; Wang, Z.; et al. Synergy between MC1R and ASIP for coat color in horses (Equus caballus). J. Anim. Sci. 2019, 97, 1578–1585. [Google Scholar] [CrossRef]
- MacKowski, M.; Wodas, L.; Brooks, S.A.; Cieslak, J. TBX3 and ASIP genotypes reveal discrepancies in officially recorded coat colors of Hucul horses. Animal 2019, 13, 1811–1816. [Google Scholar] [CrossRef]
- Druml, T.; Grilz-Seger, G.; Horna, M.; Brem, G. Discriminant Analysis of Colour Measurements Reveals Allele Dosage Effect of ASIP/MC1R in Bay Horses. Czech J. Anim. Sci. 2018, 63, 347–355. [Google Scholar]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; (version 3.4.1 (“Single Candle”)); Available online: https://www.R-project.org/ (accessed on 30 June 2017).
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbino, D.R.; Achuthan, P.; Akanni, W.; Amode, M.R.; Barrell, D.; Bhai, J.; Billis, K.; Cummins, C.; Gall, A.; Girón, C.G.; et al. Ensembl 2018. Nucleic Acids Res. 2018, 46, D754–D761. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The Human Genome Browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Kalbfleisch, T.S.; Rice, E.S.; DePriest, M.S.; Walenz, B.P.; Hestand, M.S.; Vermeesch, J.R.; O′Connell, B.L.; Fiddes, I.T.; Vershinina, A.O.; Saremi, N.F.; et al. Improved reference genome for the domestic horse increases assembly contiguity and composition. Commun. Biol. 2018, 1, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wade, C.M.; Giulotto, E.; Sigurdsson, S.; Zoli, M.; Gnerre, S.; Imsland, F.; Lear, T.L.; Adelson, D.L.; Bailey, E.; Bellone, R.R.; et al. Genome sequence, comparative analysis, and population genetics of the domestic horse. Science 2009, 326, 865–867. [Google Scholar] [CrossRef] [Green Version]
- Pruitt, K.D.; Brown, G.R.; Hiatt, S.M.; Thibaud-Nissen, F.; Astashyn, A.; Ermolaeva, O.; Farrell, C.M.; Hart, J.; Landrum, M.J.; McGarvey, K.M.; et al. RefSeq: An update on mammalian reference sequences. Nucleic Acids Res. 2014, 42, D756–D763. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Mucaki, E.J.; Shirley, B.C.; Rogan, P.K. Prediction of Mutant mRNA Splice Isoforms by Information Theory-Based Exon Definition. Hum. Mutat. 2013, 34, 557–565. [Google Scholar] [CrossRef]
- Dreger, D.L.; Parker, H.G.; Ostrander, E.A.; Schmutz, S.M. Identification of a mutation that is associated with the saddle tan and black-and-tan phenotypes in Basset Hounds and Pembroke Welsh Corgis. J. Hered. 2013, 104, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Bultman, S.J.; Michaud, E.J.; Woychik, R.P. Molecular characterization of the mouse agouti locus. Cell 1992, 71, 1195–1204. [Google Scholar] [CrossRef]
- Cropley, J.E.; Suter, C.M.; Beckman, K.B.; Martin, D.I.K. Germ-Line Epigenetic Modification of the Murine A vy allele by Nutritional Supplementation. Proc. Natl. Acad. Sci. USA 2006, 103, 17308–17312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaud, E.J.; Bultman, S.J.; Klebig, M.L.; Van Vugt, M.J.; Stubbs, L.J.; Russell, L.B.; Woychik, R.P. A Molecular Model for the Genetic and Phenotypic Characteristics of the Mouse Lethal Yellow (AY) Mutation. Proc. Natl. Acad. Sci. USA 1994, 91, 2562–2566. [Google Scholar] [CrossRef] [Green Version]
- Hysi, P.G.; Valdes, A.M.; Liu, F.; Furlotte, N.A.; Evans, D.M.; Bataille, V.; Visconti, A.; Hemani, G.; McMahon, G.; Ring, S.M.; et al. Genome-wide association meta-analysis of individuals of European ancestry identifies new loci explaining a substantial fraction of hair color variation and heritability. Nat. Genet. 2018, 50, 652–656. [Google Scholar] [CrossRef] [Green Version]
- Dupin, E.; Sommer, L. Neural crest progenitors and stem cells: From early development to adulthood. Dev. Biol. 2012, 366, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Lightbody, T. Foal with Overo lethal white syndrome born to a registered quarter horse mare. Can. Vet. J. 2002, 43, 715–717. [Google Scholar]
- Bellone, R.R.; Holl, H.; Setaluri, V.; Devi, S.; Maddodi, N.; Archer, S.; Sandmeyer, L.; Ludwig, A.; Foerster, D.; Pruvost, M.; et al. Evidence for a Retroviral Insertion in TRPM1 as the Cause of Congenital Stationary Night Blindness and Leopard Complex Spotting in the Horse. PLoS ONE 2013, 8, e78280. [Google Scholar] [CrossRef] [Green Version]
- Andersson, L.S.; Wilbe, M.; Viluma, A.; Cothran, G.; Ekesten, B.; Ewart, S.; Lindgren, G. Equine Multiple Congenital Ocular Anomalies and Silver Coat Colour Result from the Pleiotropic Effects of Mutant PMEL. PLoS ONE 2013, 8, e75639. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, L.N.; Staiger, E.A.; Albright, J.D.; Brooks, S.A. The MC1R and ASIP coat color loci may impact behavior in the horse. J. Hered. 2016, 107, 214–219. [Google Scholar] [CrossRef]
- Klebig, M.L.; Wilkinson, J.E.; Geisler, J.G.; Woychik, R.P. Ectopic expression of the agouti gene in transgenic mice causes obesity, features of type II diabetes, and yellow fur. Proc. Natl. Acad. Sci. USA 1995, 92, 4728–4732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curik, I.; Druml, T.; Seltenhammer, M.; Sundström, E.; Pielberg, G.R.; Andersson, L.; Sölkner, J. Complex Inheritance of Melanoma and Pigmentation of Coat and Skin in Grey Horses. PLoS Genet. 2013, 9, e1003248. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed Group | Sample Size | β (se) | p-Value |
|---|---|---|---|
| All combined | 126 | −30.0 (3.6) | 2.18 × 10−13 |
| Arabians | 61 | −24.1 (5.3) | 2.47 × 10−5 |
| Persian Horses | 23 | −33.7 (8.8) | 9.95 × 10−4 |
| Quarter Horses | 39 | −41.8 (6.0) | 3.00 × 10−8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corbin, L.J.; Pope, J.; Sanson, J.; Antczak, D.F.; Miller, D.; Sadeghi, R.; Brooks, S.A. An Independent Locus Upstream of ASIP Controls Variation in the Shade of the Bay Coat Colour in Horses. Genes 2020, 11, 606. https://doi.org/10.3390/genes11060606
Corbin LJ, Pope J, Sanson J, Antczak DF, Miller D, Sadeghi R, Brooks SA. An Independent Locus Upstream of ASIP Controls Variation in the Shade of the Bay Coat Colour in Horses. Genes. 2020; 11(6):606. https://doi.org/10.3390/genes11060606
Chicago/Turabian StyleCorbin, Laura J., Jessica Pope, Jacqueline Sanson, Douglas F. Antczak, Donald Miller, Raheleh Sadeghi, and Samantha A. Brooks. 2020. "An Independent Locus Upstream of ASIP Controls Variation in the Shade of the Bay Coat Colour in Horses" Genes 11, no. 6: 606. https://doi.org/10.3390/genes11060606