C:N:P Stoichiometry of Plant, Litter and Soil along an Elevational Gradient in Subtropical Forests of China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
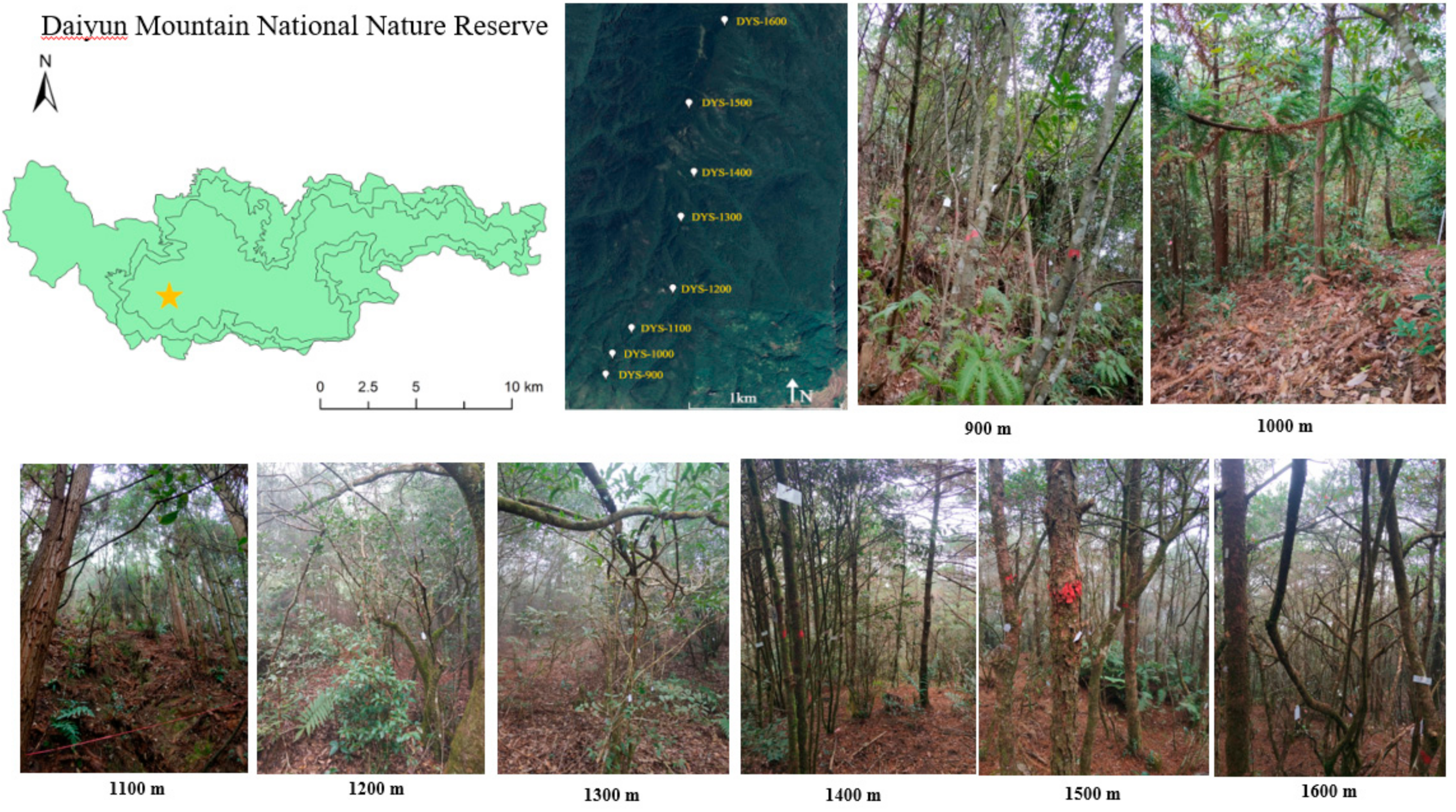
2.1. Study Site
2.2. Sample Plot Setting
2.3. Data Collection
2.4. Data Analysis
3. Results
3.1. Characteristics of C, N and P Contents and Ecological Stoichiometric Ratio of Plants and Litter at Different Elevations
3.2. C:N, C:P and N:P Correlations of Plant, Litter and Soil in Daiyun Mountain
3.3. Structural Equation Model Analysis of Plant, Litter and Main Environmental Factors
4. Discussion
4.1. Stoichiometric Characteristics of Forest Ecosystem in Daiyun Mountain
4.2. Stoichiometric Relationship of Forest Ecosystems in Daiyun Mountain
4.3. Environmental Factors Affecting the Stoichiometric Ratio of Daiyun Mountain Forest Ecosystem
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Index | Evaluation Standard | Final Model Fitting Results |
|---|---|---|
| p-value (Chi-square) | >0.05 | 0.118 |
| GFI | >0.90 | 0.950 |
| CFI | >0.90 | 0.933 |
| IFI | >0.90 | 0.974 |
| SRMR | <0.08 | 0.075 |
| Component | Element | AT | ST |
|---|---|---|---|
| Plant | TC | −0.098 | −0.212 |
| TN | −0.492 * | −0.531 ** | |
| TP | −0.437 * | −0.489 * | |
| C:N | 0.431 * | 0.433 * | |
| C:P | 0.427 * | 0.459 * | |
| N:P | −0.115 | −0.074 | |
| Litter | TC | −0.212 | −0.286 |
| TN | −0.657 ** | −0.782 ** | |
| TP | −0.454 * | −0.511 * | |
| C:N | 0.559 ** | 0.663 ** | |
| C:P | 0.324 | 0.351 | |
| N:P | −0.230 | −0.303 | |
| Soil | TC | 0.006 | 0.105 |
| TN | −0.329 | −0.342 | |
| TP | 0.556 ** | 0.620 ** | |
| C:N | 0.164 | 0.259 | |
| C:P | −0.310 | −0.287 | |
| N:P | −0.424 * | −0.468 * |
References
- Fang, J.Y.; Chen, A.P.; Peng, C.G.; Zhao, S.Q.; Ci, L.J. Changes in forest biomass carbon storage in China between 1949 and 1998. Science 2001, 292, 2320–2322. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.R.; Song, X.; Wang, Q.F.; Liu, Y.F.; Guan, D.X.; Yan, J.H.; Sun, X.M.; Zhang, L.M.; Wen, X.F. Water use efficiency of forest ecosystems in eastern China and its relations to climatic variables. New Phytol. 2008, 177, 927–937. [Google Scholar] [CrossRef]
- Elser, J.J.; Stemer, R.W.; Gorokhova, E.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Harrison, J.F.; Hobbie, S.E.; et al. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Grace, J.; Rayment, M. Respiration in the balance. Nature 2000, 404, 819–820. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, S.E.; Nadelhoffer, K.J.; Hgberg, P. A synthesis: The role of nutrients as constraints on carbon balances in boreal and arctic regions. Plant Soil 2002, 242, 163–170. [Google Scholar] [CrossRef]
- Baxter, I.; Dilkes, B.P. Elemental profiles reflect plant adaptations to the environment. Science 2012, 336, 1661–1663. [Google Scholar] [CrossRef] [Green Version]
- Elser, J. Growth rate-stoichiometry couplings in diverse biota. Ecol. Lett. 2010, 6, 936–943. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Shen, F.; Thompson, J.; Liu, W.; Bardgett, R.D. Stoichiometric traits (N:P) of understory plants contribute to reductions in plant diversity following long-term nitrogen addition in subtropical forest. Ecol. Evol. 2021, 11, 4243–4251. [Google Scholar] [CrossRef]
- Liu, R.; Wang, D. C:N:P stoichiometric characteristics and seasonal dynamics of leaf-root-litter-soil in plantations on the loess plateau. Ecol. Indic. 2021, 127, 107772. [Google Scholar] [CrossRef]
- Zhu, Q.; Liao, K.; Lai, X.M.; Lv, L.G. Scale-dependent effects of environmental factors on soil organic carbon, soil nutrients and stoichiometry under two contrasting land use types. Soil Use Manag. 2021, 37, 243–256. [Google Scholar] [CrossRef]
- Han, W.X.; Fang, J.Y.; Guo, D.L.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef]
- Parton, W.; Silver, W.L.; Burke, I.C.; Grassens, L.; Harmon, M.E.; Currie, W.; King, J.Y.; Adair, E.C.; Brandt, L.; Hart, S.C. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 2007, 315, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Thornton, P.E.; Post, W.M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2013, 22, 737–749. [Google Scholar] [CrossRef]
- Tian, H.; Chen, G.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P ratios in China’ s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Li, Y.; Wu, J.; Liu, S.; Shen, J.; Huang, D.; Su, Y.; Wei, W.; Syers, J.K. Is the C:N:P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China? Glob. Biogeochem. Cycles 2012, 26, 4002. [Google Scholar] [CrossRef]
- Pang, Y.; Tian, J.; Zhao, X.; Chao, Z. The linkages of plant, litter and soil C:N:P stoichiometry and nutrient stock in different secondary mixed forest types in the Qinling Mountains, China. PeerJ 2020, 8, e9274. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 33–47. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, X.; Liang, W.; Dai, G.; Wang, X.; Han, S. Distribution of soil organic carbon fractions along the altitudinal gradient in Changbai Mountain, China. Pedosphere 2011, 21, 615–620. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.F.; Chen, Y.M.; Zhou, Y.; Zheng, H.F.; Xu, Z.F.; Tan, B.; You, C.M.; Zhang, L.; Li, H.; Guo, L.; et al. Litter chemical traits strongly drove the carbon fractions loss during decomposition across an alpine treeline ecotone. Sci. Total Environ. 2021, 753, 142287. [Google Scholar] [CrossRef]
- He, X.J.; Hou, E.Q.; Liu, Y.; Wen, D.Z. Altitudinal patterns and controls of plant and soil nutrient concentrations and stoichiometry in subtropical China. Sci. Rep. 2016, 6, 24261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; He, Z.S.; Gu, X.G.; Liu, J.F.; Liu, Y.H. Classification and ordination of the Pinus taiwanensis forest on Daiyun Mountain, Fujian Province, China. Taiwania 2020, 65, 119–128. [Google Scholar]
- Su, S.J.; Liu, J.F.; He, Z.S.; Zheng, S.Q.; Hong, W.; Xu, D.W. Ecological species groups and interspecific association of dominant tree species in Daiyun Mountain National Nature Reserve. J. Mt. Sci. 2015, 12, 637–646. [Google Scholar] [CrossRef]
- Xu, D.W.; Liu, J.F.; Marshall, P.; He, Z.S.; Zheng, S.Q. Leaf litter decomposition dynamics in unmanaged Phyllostachys pubescens stands at high elevations in the Daiyun Mountain National Nature Reserve. J. Mt. Sci. 2017, 14, 2246–2256. [Google Scholar] [CrossRef]
- Jiang, L.; He, Z.S.; Liu, J.F.; Xing, C.; Gu, X.G.; Wei, C.S.; Zhu, J.; Wang, X.L. Elevation Gradient Altered Soil C, N, and P Stoichiometry of Pinus taiwanensis Forest on Daiyun Mountain. Forests 2019, 10, 1089. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.F.; Su, S.J.; He, Z.S.; Jiang, L.; Hong, W. Relationship between Pinus taiwanensis seedling regeneration and the spatial heterogeneity of soil nitrogen in Daiyun Mountain, southeast China. Ecol. Indic. 2020, 115, 106398. [Google Scholar] [CrossRef]
- Condit, R. Tropical Forest Census Plots; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- People’ s Republic of China Forestry. Industry Standard. Forest Soil Analysis Method; The State Forestry Administration of the People’ s Republic of China: Beijing, China, 1999; pp. 71–113. (In Chinese)
- R Core team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Volume 11, p. 7. [Google Scholar]
- Shipley, B. A New inferential test for path models based on directed acyclic graphs. Struct. Equ. Modeling A Multidiscip. J. 2000, 7, 206–218. [Google Scholar] [CrossRef]
- Rosseel, Y. lavaan: An R package for structural equation modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Redfield, A.C. The biological control of chemical factors in the environment. Am. Sci. 1958, 46, 205–221. [Google Scholar]
- Feng, L.; Hu, J.; Xie, Y.; Cong, H.; Chen, X.; Deng, Z. Foliar stoichiometry of carbon, nitrogen, and phosphorus in wetland sedge Carex brevicuspis along a small-scale elevation gradient. Ecol. Indic. 2017, 92, 322–329. [Google Scholar]
- Schinner, F. Soil microbial activities and litter decomposition related to altitude. Plant Soil 1982, 65, 87–94. [Google Scholar] [CrossRef]
- Ren, S.J.; Yu, G.R.; Jiang, C.M.; Fang, H.J.; Sun, X.M. Stoichiometric characteristics of leaf carbon, nitrogen, and phosphorus of 102 dominant species in forest ecosystems along the North-South Transect of East China. Chin. J. Appl. Ecol. 2012, 23, 581–586. (In Chinese) [Google Scholar]
- Wang, J.Y.; Wang, S.Q.; Li, R.L.; Yan, J.H.; Sha, L.Q.; Han, S.J. C:N:P stoichiometric characteristics of four forest types’ dominant tree species in China. Chin. J. Plant Ecol. 2011, 35, 587–595. (In Chinese) [Google Scholar]
- Hu, Q.; Sheng, M.; Bai, Y.; Xiao, H. Response of C, N, and P stoichiometry characteristics of Broussonetia papyrifera to altitude gradients and soil nutrients in the karst rocky ecosystem, SW China. Plant Soil 2020, 14, 1–14. [Google Scholar] [CrossRef]
- Xiang, X.Y.; Chen, S.K.; Pan, P.; Ouyang, X.Z.; Ning, J.K.; Li, Q. Stoichiometric traits of C, N and P of leaf-litter-soil system of Pinus massoniana forest. J. For. Environ. 2019, 39, 11–17. (In Chinese) [Google Scholar]
- Wang, P.A.; Gong, Y.Q.; Wang, Q.W.; Ren, M.X.; Bo, F.J.; Zhang, Y.X.; Guo, J.P. Carbon, nitrogen and phosphorus stoichiometric characteristics of needle leaf- leaf litter- soil from Larix principis-rupprechtii plantation with different ages. J. Northwest For. Univ. 2020, 35, 1–9. (In Chinese) [Google Scholar]
- Bai, X.J.; Zeng, Q.C.; An, S.S.; Wang, B.R. Ecological stoichiometry characteristics of leaf-litter-soil in different plantations on the Loess Plateau, China. Chin. J. Appl. Ecol. 2016, 27, 3823–3830. [Google Scholar]
- Vitousek, P. Nutrient cycling and nutrient use efficiency. Am. Nat. 1982, 119, 553–572. [Google Scholar] [CrossRef]
- Tessier, J.T.; Raynal, D.J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. J. Appl. Ecol. 2003, 40, 523–534. [Google Scholar] [CrossRef]
- Guo, Y.; Yan, Z.; Gheyret, G.; Zhou, G.; Xie, Z.; Tang, Z. The community-level scaling relationship between leaf nitrogen and phosphorus changes with plant growth, climate and nutrient limitation. J. Ecol. 2020, 108, 1276–1286. [Google Scholar] [CrossRef]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2010, 164, 243–266. [Google Scholar] [CrossRef]
- Koerselman, W.; Meuleman, A.F.M. The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Tong, R.; Zhou, B.Z.; Jiang, L.L.; Ge, X.G.; Cao, Y.H.; Shi, J.X. Leaf litter carbon, nitrogen and phosphorus stoichiometry of Chinese fir (Cunninghamia lanceolata) across China. Glob. Ecol. Conserv. 2021, 27, e1542. [Google Scholar] [CrossRef]
- Aerts, R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship. Oikos 1997, 79, 439–449. [Google Scholar] [CrossRef]
- Guo, C.; Liu, S.B.; Xiang, Y.Z.; Tang, X.L.; Liu, H.T.; Yao, B.; Luo, X.Q. Impact of living mulch on soil C:N:P stoichiometry in orchards across China: A meta-analysis examining climatic, edaphic, and biotic dependency. Pedosphere 2020, 30, 26–34. [Google Scholar]
- Chen, C.; Zhang, S.J.; Li, L.D.; Liu, Z.D.; Chen, J.L.; Gu, X.; Wang, L.F.; Fang, X. Carbon, nitrogen and phosphorus stoichiometry in leaf, litter and soil at different vegetation restoration stages in the mid-subtropical region of China. Chin. J. Plant Ecol. 2019, 43, 658–671. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, G.Q.; Zhang, P.; Peng, S.Z.; Chen, Y.M.; Cao, Y. The coupling of leaf, litter, and soil nutrients in warm temperate forests in northwestern China. Sci. Rep. 2017, 7, 11754. [Google Scholar] [CrossRef]
- Bo, F.J.; Zhang, Y.X.; Chen, H.Y.H.; Wang, P.G.; Ren, X.M.; Guo, J.P. The C:N:P stoichiometry of planted and natural Larix principis-rupprechtii stands along altitudinal gradients on the Loess Plateau, China. Forests 2020, 11, 363. [Google Scholar] [CrossRef] [Green Version]
- Johannes, R.; Philip, C.B.; Erland, B. The microbial PLFA composition as affected by pH in an arable soil. Soil Biol. Biochem. 2009, 42, 516–520. [Google Scholar]
- Bååth, E.; Anderson, T.H. Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol. Biochem. 2003, 35, 955–963. [Google Scholar] [CrossRef]
- Thomas, S.; Katharina, M.K.; Emanuel, S.; Katja, S.G.; Ellersdorfer, G.; Bernd, R.; Andreas, R.; Sophie, Z.B.; Kathrin, R. Who is who in litter decomposition? Metaproteomics reveals major microbial players and their biogeochemical functions. Multidiscip. J. Microb. Ecol. 2012, 6, 1749–1762. [Google Scholar]
- Qin, Y.Y.; Feng, Q.; Adamowski, J.F.; Zhu, M.; Zhang, X.F. Community level response of leaf stoichiometry to slope aspect in a montane environment: A case study from the Central Qilian Mountains, China. Glob. Ecol. Conserv. 2021, 28, e01703. [Google Scholar] [CrossRef]
- Ai, Z.M.; He, L.R.; Xin, Q.; Yang, T.; Liu, G.B.; Xue, S. Slope aspect affects the non-structural carbohydrates and C:N:P stoichiometry of Artemisia sacrorum on the Loess Plateau in China. Catena 2017, 152, 9–17. [Google Scholar] [CrossRef]
- Waigwa, A.N.; Mwangi, B.N.; Wahiti, G.R.; Omengo, F.; Zhou, Y.D.; Wang, Q.F. Variation of morphological and leaf stoichiometric traits of two endemic species along the elevation gradient of Mount Kenya, East Africa. J. Plant Ecol. 2020, 13, 785–792. [Google Scholar] [CrossRef]
- Chen, L.L.; Qiang, D.; Yuan, Z.Y.; Mu, X.M.; Kallenbach, R.L. Age-related C:N:P stoichiometry in two plantation forests in the Loess Plateau of China. Ecol. Eng. 2018, 120, 14–22. [Google Scholar] [CrossRef]



| Items | Components | 900 m | 1000 m | 1100 m | 1200 m | 1300 m | 1400 m | 1500 m | 1600 m |
|---|---|---|---|---|---|---|---|---|---|
| TC (g/kg) | Plant | 458.49 ± 0.33 dB | 480.88 ± 2.61 aA | 496.89 ± 3.74 abA | 471.00 ± 1.93 cA | 478.95 ± 2.61 abA | 470.38 ± 0.54 cA | 494.36 ± 1.75 bcB | 470.42 ± 0.11 cA |
| Litter | 433.71 ± 0.78 eC | 473.44 ± 1.27 bB | 475.44 ± 1.41 bA | 409.30 ± 2.51 fB | 460.29 ± 0.99 dB | 467.33 ± 0.57 cB | 480.06 ± 1.37 aA | 460.31 ± 0.69 dB | |
| TN (g/kg) | Plant | 14.99 ± 0.96 bA | 12.55 ± 0.28 cdA | 12.22 ± 0.53 bcdA | 15.10 ± 1.14 bcA | 12.67 ± 0.49 bcdA | 16.07 ± 2.51 dA | 14.83 ± 0.93 cdA | 18.19 ± 1.01 aA |
| Litter | 8.9 ± 0.18 dC | 8.33 ± 0.46 dC | 10.90 ± 0.48 cC | 13.50 ± 0.14 bB | 10.37 ± 0.48bC | 11.43 ± 0.41 aB | 11.64 ± 1.86 bC | 14.73 ± 0.19 bC | |
| TP (g/kg) | Plant | 1.24 ± 0.14 cdA | 1.12 ± 0.06 cdA | 1.18 ± 0.07 cdA | 1.33 ± 0.09 bcA | 1.50 ± 0.08 bA | 1.40 ± 0.33 aA | 1.38 ± 0.10 dA | 1.44 ± 0.10 cdA |
| Litter | 0.43 ± 0.07 cdB | 0.5 ± 0.04 abB | 0.38 ± 0.02 cB | 0.53 ± 0.01 abB | 0.55 ± 0.06 aB | 0.50 ± 0.06 abB | 0.52 ± 0.05 abB | 0.59 ± 0.04 aB | |
| C:N | Plant | 30.71 ±1.89 abB | 38.35 ± 0.95 abB | 39.09 ± 1.93 abA | 31.38 ± 2.37 abA | 37.86 ± 1.28 abA | 30.05 ± 5.01 aA | 33.49 ± 2.26 aA | 25.94 ± 1.43 bB |
| Litter | 48.77 ± 0.92 bC | 57.05 ± 3.44 aC | 43.70 ± 1.46 cB | 30.32 ± 0.19 eA | 44.40 ± 1.14 cdeC | 40.96 ±0.67 eC | 41.32 ± 4.56 cdB | 31.26 ± 0.42 deC | |
| C:P | Plant | 372.54 ± 38.42 bcB | 430.56 ± 23.75 abB | 405.08± 26.47 abB | 355.16 ± 27.58 cdB | 321.00 ± 16.34 dB | 351.62 ± 74.76 dB | 361.19 ± 28.14 aB | 327.66 ± 22.38 abcA |
| Litter | 1028.07 ± 163.17 abB | 959.83 ± 80.83 abB | 1250.04 ±76.39 aB | 776.90 ± 15.9 bB | 843.25 ± 91.46 bA | 955.02 ± 102.88 abB | 938.12 ± 101.61 abB | 789.95 ± 53.06 bA | |
| N:P | Plant | 12.10 ± 0.53 abB | 11.24 ±0.78 abB | 10.36 ± 0.36 abC | 11.34 ± 0.69 bcC | 8.48 ± 0.26 cB | 11.63 ± 1.27 cB | 10.79 ± 0.47 abB | 12.66 ± 0.92 aB |
| Litter | 21.11 ± 3.56 cdB | 16.81 ± 0.71 dA | 28.73 ± 2.57 abB | 25.63 ± 0.50 abcB | 18.99 ± 1.88 abcA | 23.43 ± 3.53 aA | 22.73 ± 3.29 abcA | 25.28 ± 1.83 bcdA |
| Factors | Plant | Litter | Soil |
|---|---|---|---|
| ELE | −0.481 * | −0.429 * | 0.3319 * |
| pH | 0.266 | 0.2153 | −0.4296 * |
| Slope | 0.301 * | 0.3297 * | 0.0738 |
| SWC | −0.41 | −0.3561 | 0.3159 |
| AT | 0.356 ** | 0.2456 | 0.3118 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, B.; Chen, L.; Jiang, L.; Zhu, J.; Chen, J.; Huang, Q.; Liu, J.; Xu, D.; He, Z. C:N:P Stoichiometry of Plant, Litter and Soil along an Elevational Gradient in Subtropical Forests of China. Forests 2022, 13, 372. https://doi.org/10.3390/f13030372
Chen B, Chen L, Jiang L, Zhu J, Chen J, Huang Q, Liu J, Xu D, He Z. C:N:P Stoichiometry of Plant, Litter and Soil along an Elevational Gradient in Subtropical Forests of China. Forests. 2022; 13(3):372. https://doi.org/10.3390/f13030372
Chicago/Turabian StyleChen, Bo, Lyuyi Chen, Lan Jiang, Jing Zhu, Jiajia Chen, Qingrong Huang, Jinfu Liu, Daowei Xu, and Zhongsheng He. 2022. "C:N:P Stoichiometry of Plant, Litter and Soil along an Elevational Gradient in Subtropical Forests of China" Forests 13, no. 3: 372. https://doi.org/10.3390/f13030372