Growth Trends of Coniferous Species along Elevational Transects in the Central European Alps Indicate Decreasing Sensitivity to Climate Warming

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Sites
2.2. Tree-Ring Analysis
2.3. Climate Data
2.4. Statistical Analysis
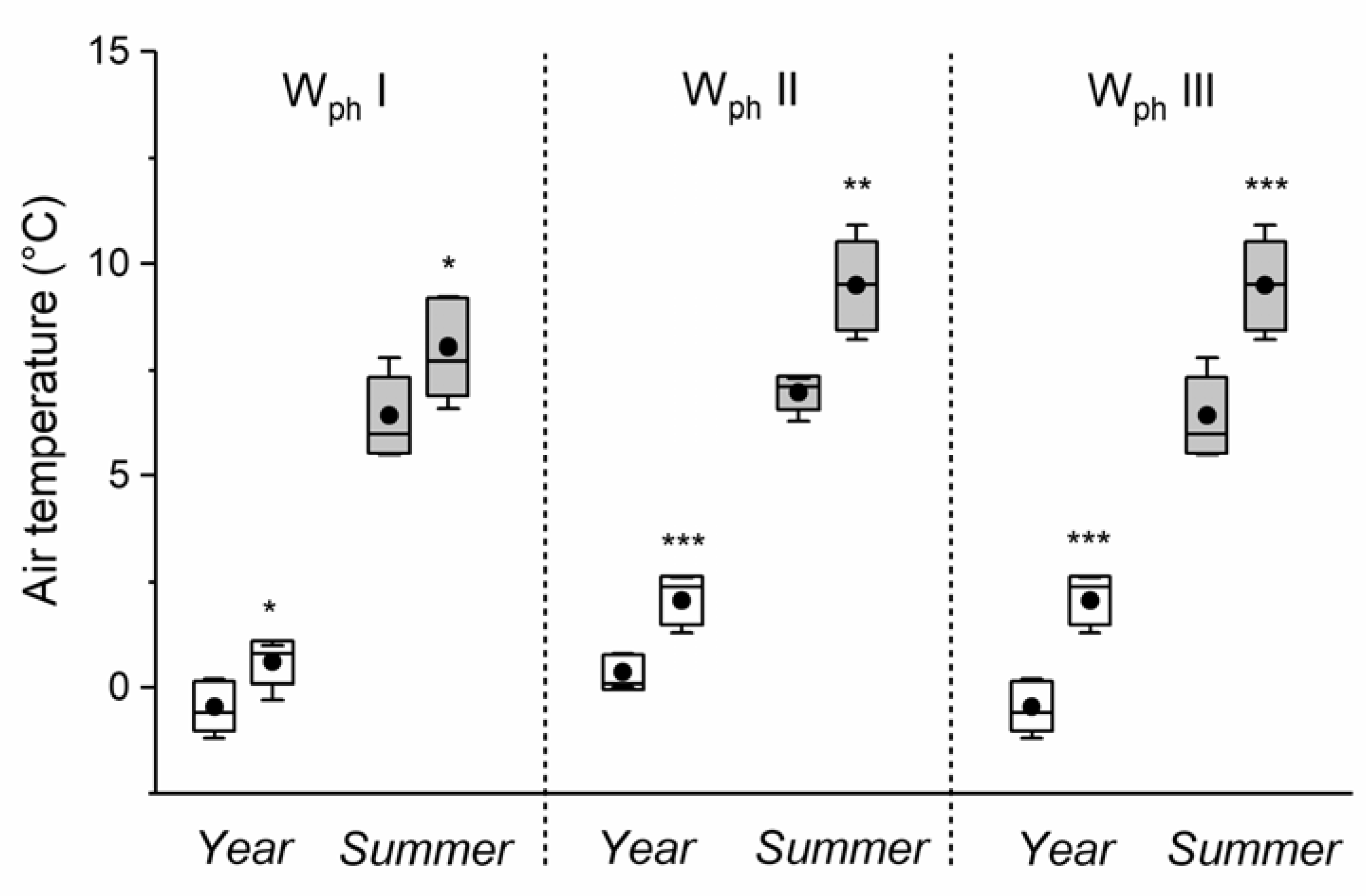
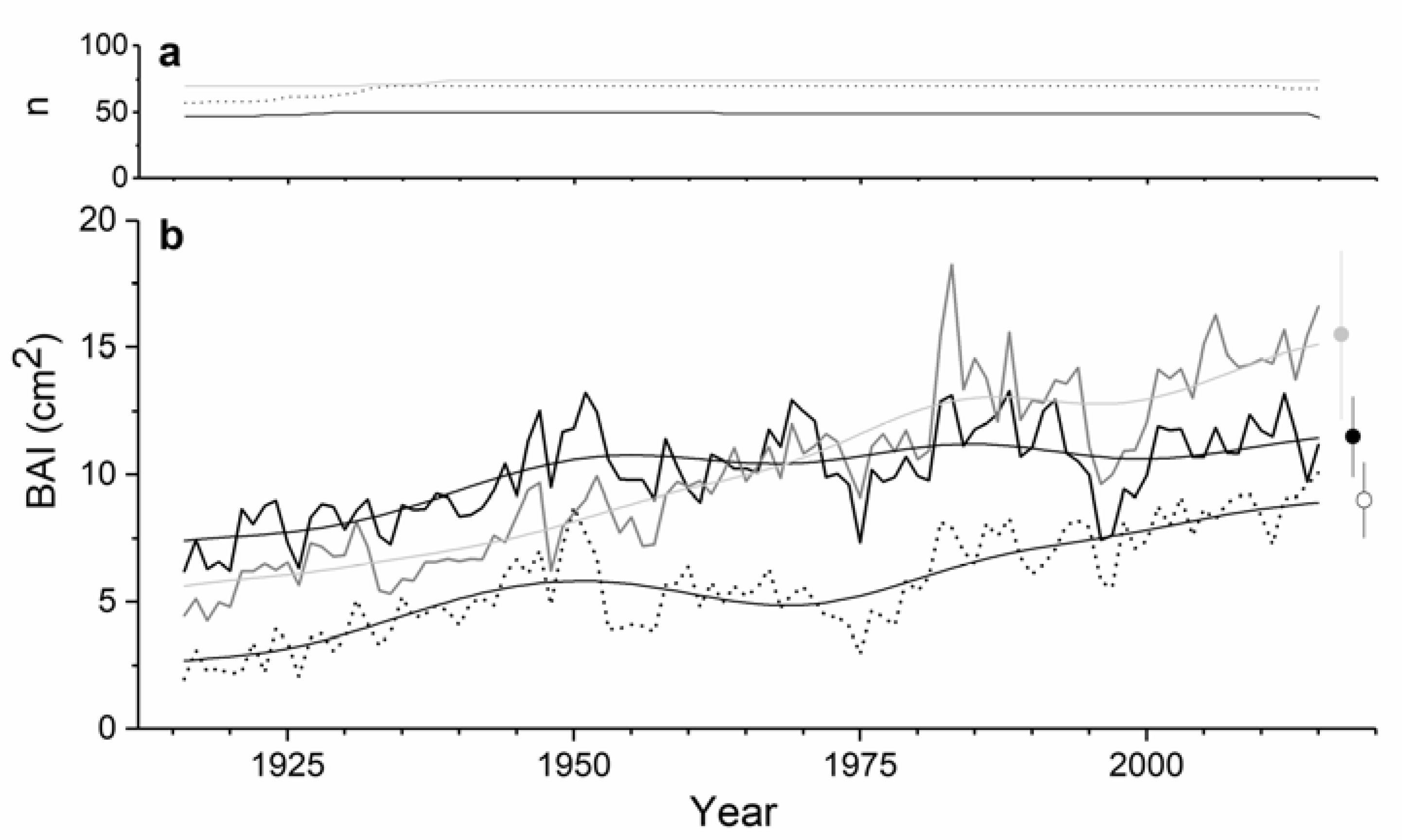
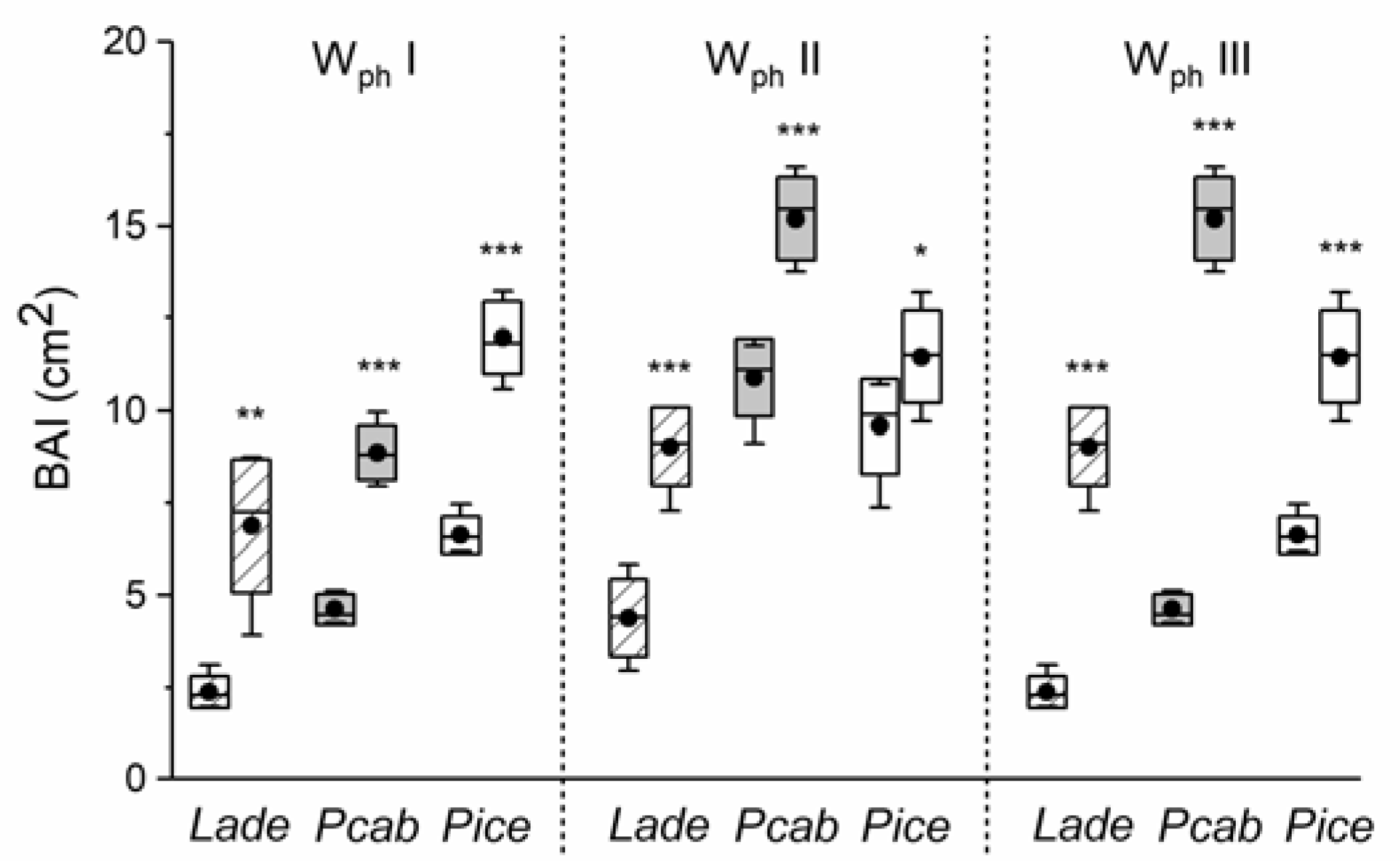
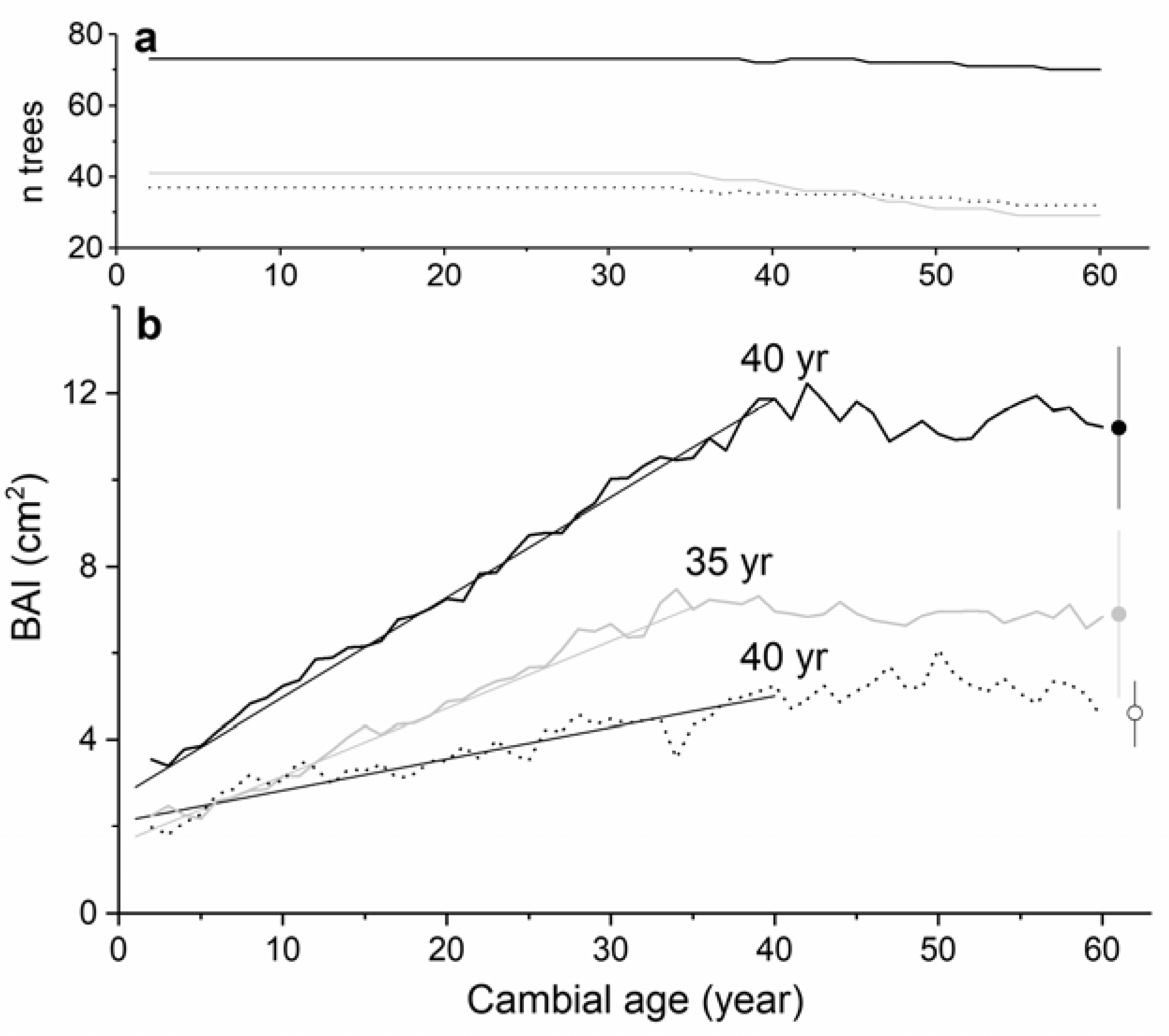
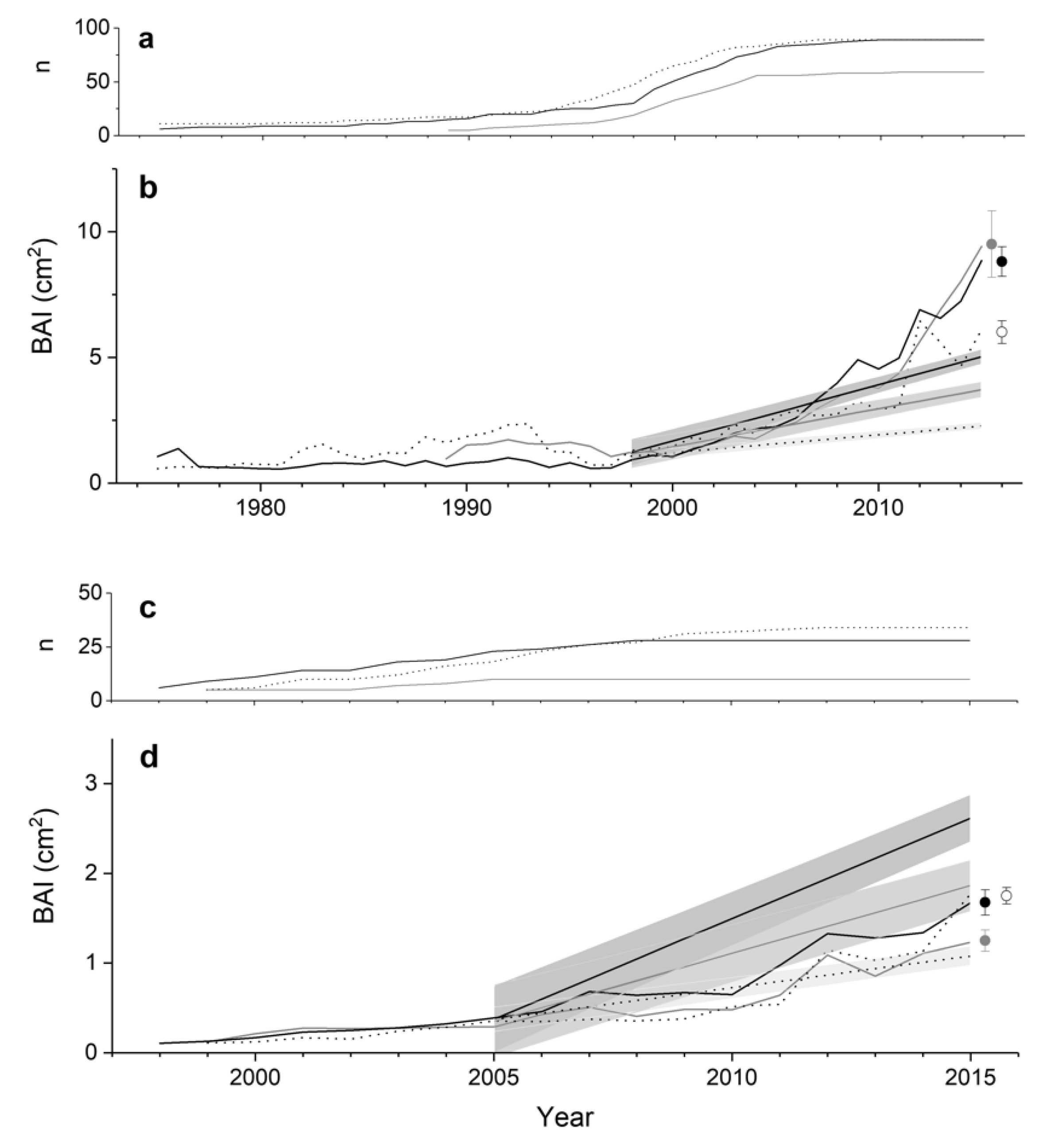
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Körner, C. A re-assessment of high elevation tree line positions and their explanation. Oecologia 1998, 115, 445–459. [Google Scholar] [PubMed] [Green Version]
- Esper, J.; Cook, E.R.; Schweingruber, F.H. Low-frequency signals in long tree-ring chronologies for reconstructing past temperature variability. Science 2002, 295, 2250–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieser, G.; Tausz, M. Trees at Their Upper Limit: Treelife Limitation at the Alpine Timberline. In Plant Ecophysiology; Springer: Berlin/Heidelberg, Germany, 2007; Volume 5. [Google Scholar]
- Körner, C. Alpine Treelines. Funtional Ecology of the Global High Elevation Tree Limits; Springer: Basel, Switzerland, 2012. [Google Scholar]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Gričar, J.; Seo, J.-W.; Rathgeber, C.B.K.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Carrer, M.; Urbinati, C. Age-dependent tree ring growth responses to climate of Larix decidua and Pinus cembra in the Italian Alps. Ecology 2004, 85, 730–740. [Google Scholar] [CrossRef]
- Oberhuber, W. Influence of climate on radial growth of Pinus cembra within the alpine timberline ecotone. Tree Physiol. 2004, 24, 291–301. [Google Scholar] [CrossRef]
- IPCC. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. In Climate Change 2014: Synthesis Report; Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Auer, I.; Böhm, R.; Jurkovic, A.; Lipa, W.; Orlik, A.; Potzmann, R.; Schöner, W.; Ungersböck, M.; Matulla, C.; Briffa, K.; et al. HISTALP—Historical instrumental climatological surface time series of the greater Alpine region 1760–2003. Int. J. Climatol. 2007, 27, 17–46. [Google Scholar] [CrossRef]
- Gobiet, A.; Kotlarski, S.; Beniston, M.; Heinrich, G.; Rajczak, J.; Stoffel, M. 21st century climate change in the European Alps–A review. Sci. Total Environ. 2014, 493, 1138–1151. [Google Scholar] [CrossRef]
- Büntgen, U.; Frank, D.; Wilson, R.; Carrer, M.; Urbinati, C. Testing for tree-ring divergence in the European Alps. Glob. Chang. Biol. 2008, 14, 2443–2453. [Google Scholar] [CrossRef]
- St George, S.; Esper, J. Concord and discord among Northern Hemisphere paleotemperature reconstructions from tree rings. Quat. Sci. Rev. 2019, 203, 278–281. [Google Scholar] [CrossRef]
- D’Arrigo, R.; Wilson, R.; Liepert, B.; Cherubini, P. On the ‘divergence problem’ in northern forests: A review of the tree ring evidence and possible causes. Glob. Planet. Chang. 2008, 60, 289–305. [Google Scholar] [CrossRef]
- Oberhuber, W.; Kofler, W.; Pfeifer, K.; Seeber, A.; Gruber, A.; Wieser, G. Long-term changes in tree-ring-climate relationships at Mt. Patscherkofel (Tyrol, Austria) since the mid-1980s. Trees 2008, 22, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cazzolla Gatti, R.; Callaghan, T.; Velichevskaya, A.; Dudko, A.; Fabbio, L.; Battipaglia, G.; Liang, J. Accelerating upward treeline shift in the Altai Mountains under last-century climate change. Sci. Rep. 2019, 9, 7678. [Google Scholar] [CrossRef] [PubMed]
- Wieser, G.; Oberhuber, W.; Gruber, A. Effects of climate change at treeline: Lessons from space-for-time studies, manipulative experiments, and long-term observational records in the Central Austrian Alps. Forests 2019, 10, 508. [Google Scholar] [CrossRef] [Green Version]
- Babst, F.; Bouriaud, O.; Alexander, R.; Trouet, V.; Frank, D. Toward consistent measurements of carbon accumulation: A multi-site assessment of biomass and basal area increment across Europe. Dendrochronologia 2014, 32, 153–161. [Google Scholar] [CrossRef]
- Salzer, M.W.; Larson, E.R.; Bunn, A.G.; Hughes, M.K. Changing climate response in near-treeline bristlecone pine with elevation and aspect. Environ. Res. Lett. 2014, 9, 114007. [Google Scholar] [CrossRef]
- Sullivan, P.F.; Ellison, S.B.Z.; McNown, R.W.; Brownlee, A.H.; Sveinbjoernsson, B. Evidence of soil nutrient availability as the proximate constraint on growth of treeline trees in northwest Alaska. Ecology 2015, 96, 716–727. [Google Scholar] [CrossRef]
- Liang, E.; Leuschner, C.; Dulamsuren, C.; Wagner, B.; Hauck, M. Global warming related tree growth decline and mortality on the north-eastern Tibetan Plateau. Clim. Chang. 2016, 134, 163–176. [Google Scholar] [CrossRef]
- Liu, B.; Wang, Y.; Zhu, H.; Liang, E.; Camarero, J.J. Topography and age mediate the growth responses of Smith fir to climate warming in the southeastern Tibetan Plateau. Int. J. Biometeorol. 2016, 60, 1577–1587. [Google Scholar] [CrossRef]
- Harris, I.C.; Jones, P.D. CRU TS4.01: Climatic Research Unit (CRU) Time-Series (TS) Version 4.01 of High-Resolution Gridded Data of Month-by-Month Variation in Climate (Jan. 1901–Dec. 2016); Centre for Environmental Data Analysis: Norwich, UK, 2017. [Google Scholar] [CrossRef]
- Estrada, F.; Perron, P.; Martínez-López, B. Statistically derived contributions of diverse human influences to twentieth-century temperature changes. Nat. Geosci. 2013, 6, 1050–1055. [Google Scholar] [CrossRef] [Green Version]
- Hegerl, G.C.; Brönnimann, S.; Schurer, A.; Cowan, T. The early 20th century warming: Anomalies, causes, and consequences. WIREs Clim. Chang. 2018, 9, e522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Forests. In Vegetation Ecology of Central Europe; Springer: Berlin/Heidelberg, Germany, 2017; Volume 1. [Google Scholar]
- Tollmann, A. Geologie von Österreich Band 1. In Die Zentralalpen; Deuticke: Wien, Austria, 1977. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree ring dating and measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- LeBlanc, D.C. Relationships between breast-height and whole-stem growth indices for red spruce on Whiteface mountain, New York. Can. J. For. Res. 1990, 20, 1399–1407. [Google Scholar] [CrossRef]
- Biondi, F.; Qeadan, F. A theory-driven approach to tree-ring standardization: Defining the biological trend from expected basal area increment. Tree Ring Res. 2008, 64, 81–96. [Google Scholar] [CrossRef] [Green Version]
- Peters, R.L.; Groenendijk, P.; Vlam, M.; Zuidema, P.A. Detecting long-term growth trends using tree rings: A critical evaluation of methods. Glob. Chang. Biol. 2015, 21, 2040–2054. [Google Scholar] [CrossRef]
- Wieser, G.; Matyssek, R.; Luzian, R.; Zwerger, P.; Pindur, P.; Oberhuber, W.; Gruber, A. Effects of atmospheric and climate change at the timberline of the Central European Alps. Ann. For. Sci. 2009, 66, 402. [Google Scholar] [CrossRef] [Green Version]
- Erb, K.H.; Kastner, T.; Luyssaert, S.; Houghton, R.A.; Kuemmerle, T.; Olofsson, P.; Haberl, H. Bias in the attribution of forest carbon sinks. Nat. Clim. Chang. 2013, 3, 854–856. [Google Scholar] [CrossRef] [Green Version]
- Büntgen, U.; Frank, D.C.; Nievergelt, D.; Esper, J. Summer temperature variations in the European Alps, AD 755–2004. J. Clim. 2006, 19, 5606–5623. [Google Scholar] [CrossRef]
- Carrer, M.; Nola, P.; Eduard, J.L.; Motta, R.; Urbinati, C. Regional variability of climate-growth relationships in Pinus cembra high elevation forests in the Alps. J. Ecol. 2007, 95, 1072–1083. [Google Scholar] [CrossRef]
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Wieser, G.; Oberhuber, W.; Gruber, A.; Leo, M.; Matyssek, R.; Grams, T.E.E. Stable water use efficiency under climate change of three sympatric conifer species at the alpine treeline. Front. Plant Sci. 2016, 7, 799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saulnier, M.; Edouard, J.L.; Corona, C.; Guibal, F. Climate/growth relationships in a Pinus cembra high-elevation network in the Southern French Alps. Ann. For. Sci. 2011, 68, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Babst, F.; Poulter, B.; Trouet, V.; Tan, K.; Neuwirth, B.; Wilson, R.J.S.; Carrer, M.; Grabner, M.; Tegel, W.; Levanič, T.; et al. Site- and species-specific responses of forest growth to climate across the European continent. Glob. Ecol. Biogeogr. 2013, 22, 706–717. [Google Scholar] [CrossRef]
- Primicia, I.; Camarero, J.J.; Janda, P.; Čada, V.; Morrissey, R.C.; Trotsiuk, V.; Bače, R.; Teodosiu, M.; Svoboda, M. Age, competition, disturbance and elevation effects on tree and stand growth response of primary Picea abies forest to climate. For. Ecol. Manag. 2015, 354, 77–86. [Google Scholar] [CrossRef]
- Szeicz, J.M.; MacDonald, G.M. Age dependent tree-ring growth response of subarctic white spruce to climate. Can. J. For. Res. 1994, 24, 120–132. [Google Scholar] [CrossRef]
- Girardin, M.P.; Guo, X.J.; Bernier, P.Y.; Raulier, F.; Gauthier, S. Changes in growth of pristine boreal North American forests from 1950 to 2005 driven by landscape demographics and species traits. Biogeosciences 2012, 9, 2523–2536. [Google Scholar] [CrossRef] [Green Version]
- Esper, J.; Niederer, R.; Bebi, P.; Frank, D. Climate signal age effects-Evidence from young and old trees in the Swiss Engadin. For. Ecol. Manag. 2008, 255, 3783–3789. [Google Scholar] [CrossRef]
- Cherubini, P.; Dobbertin, M.; Innes, J.L. Potential sampling bias in long-term forest growth trends reconstructed from tree rings: A case study from the Italian Alps. For. Ecol. Manag. 1998, 109, 103–118. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Gloor, E.; Zuidema, P.A. Detecting evidence for CO2 fertilization from tree ring studies: The potential role of sampling biases. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]
- Nehrbass-Ahles, C.; Babst, F.; Klesse, S.; Nötzli, M.; Bouriaud, O.; Neukom, R.; Dobbertin, M.; Frank, D. The influence of sampling design on tree-ring-based quantification of forest growth. Glob. Chang. Biol. 2014, 20, 2867–2885. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Brienen, R.J.W.; Gloor, E.; Phillips, O.L.; Prior, L.D. Detecting trends in tree growth: Not so simple. Trends Plant Sci. 2013, 18, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Duchesne, L.; Houle, D.; Ouimet, R.; Caldwell, L.; Gloor, M.; Brienen, R. Large apparent growth increase in boreal forests inferred from tree-rings are an artefact of sampling design. Sci. Rep. 2019, 9, 6832. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.; Yang, J. Effects of microsite on growth of Pinus cembra in the subalpine zone of the Austrian Alps. Ann. For. Sci. 2004, 61, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Sendall, K.M.; Stefanski, A.; Wei, X.; Rich, R.L.; Montgomery, R.A. Boreal and temperate trees show strong acclimation of respiration to warming. Nature 2016, 531, 633–636. [Google Scholar] [CrossRef]
- Girardin, M.P.; Bouriaud, O.; Hogg, E.H.; Kurz, W.; Zimmermann, N.E.; Metsaranta, J.M.; Jong Rde Frank, D.C.; Esper, J.; Büntgen, U.; Guo, X.J.; et al. No growth stimulation of Canda’s boreal forest under half-century of combined warming and CO2 fertilization. Proc. Natl. Acad. Sci. USA 2016, 113, E8406–E8414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, P.; Wang, X.; Sun, H.; Fan, Y.; Wu, Y.; Lin, X.; Chang, J. Forest type and height are important in shaping the altitudinal change of radial growth response to climate change. Sci. Rep. 2019, 9, 1336. [Google Scholar] [CrossRef] [Green Version]
- Hoch, G.; Körner, C. The carbon charging of pines at the climatic treeline: A global comparison. Oecologia 2003, 135, 10–21. [Google Scholar] [CrossRef]
- Hoch, G.; Körner, C. Global patterns of mobile carbon stores in trees at the high-elevation treeline. Glob. Ecol. Biogeogr. 2012, 21, 861–871. [Google Scholar] [CrossRef]
- Gruber, A.; Pirkebner, D.; Oberhuber, W.; Wieser, G. Spatial and seasonal variations in mobile carbohydrates in Pinus cembra in the timberline ecotone of the Central Austrian Alps. Eur. J. For. Res. 2011, 130, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Handa, I.T.; Körner, C.; Hättenschwiler, S. A test of the treeline carbon limitation hypothesis by in situ CO2 enrichment and defoliation. Ecology 2005, 86, 1288–1300. [Google Scholar] [CrossRef]
- Dawes, M.A.; Hättenschwiler, S.; Bebi, P.; FHagedorn ITHanda CKörner, C. Rixen Species-specific tree growth responses to 9 years of CO2 enrichment at the alpine treeline. J. Ecol. 2011, 99, 383–394. [Google Scholar]
- Tranquillini, W. Physiological Ecology of the Alpine Timberline. In Tree Existence in High Altitudes with Special Reference to the European Alps; Ecological Studies 31; Springer: Berlin/Heidelberg, Germany, 1979. [Google Scholar]
- Hadley, J.L.; Smith, W.K. Wind effects on needles of timberline conifers: Seasonal influence of mortality. Ecology 1986, 67, 12–19. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevation (m asl) | ΔE | |||||||
|---|---|---|---|---|---|---|---|---|
| Site | Lat | Lon | Subalpine | Treeline | Krummholz | (m) | Aspect | Slope (°) |
| Patscherkofel (PTK) | 47.206 | 11.452 | 1960–2010 | 2080–2130 | 2140–2180 | 220 | WSW–W | 10–30 |
| Viggarspitze (VIG) | 47.213 | 11.480 | 2030–2050 | 2120–2060 | 2220–2270 | 240 | SSW | 10–40 |
| Morgenköpfl (MOR) | 47.179 | 11.469 | 1880–2020 | 2060–2120 | 2130–2220 | 340 | SW–NW | 10–30 |
| Kaserstattalm (KAS) | 47.115 | 11.290 | 1640–1800 | 1950–1990 | - | 350 | SE–SW | 20–30 |
| Elferlift (ELF) | 47.096 | 11.305 | 1630–1810 | - | - | 180 | NW–NE | 20–30 |
| Lazauntal (SCH1) | 46.756 | 10.779 | 2040–2290 | - | - | 250 | E | 30–45 |
| Finailtal (SCH2) | 46.743 | 10.827 | 2120–2250 | - | - | 130 | W | 30–50 |
| Subalpine | Treeline | Krummholz-Limit | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Pice | Lade | Pcab | Pice | Lade | Pcab | Pice | Lade | Pcab | |
| PTK | 124 ± 43/8 | 133 ± 45/5 | 146 ± 38/10 | 37 ± 15/15 | 32 ± 19/14 | 26 ± 18/13 | 15 ± 5/10 | 12 ± 3/10 | 23 ± 17/10 |
| VIG | 138 ± 39/8 | 128 ± 28/6 | 85 ± 11/5 | 22 ± 7/10 | 29 ± 17/11 | - | 14 ± 4/9 | 10 ± 3/12 | - |
| MOR | 118 ± 6/10 | 103 ± 12/10 | - | 25 ± 7/11 | 31 ± 20/13 | - | 17 ± 3/9 | 17 ± 5/12 | - |
| KAS | - | 133 ± 23/10 | 142 ± 23/24 | 14 ± 3/53 | 18 ± 4/51 | 16 ± 4/46 | - | - | - |
| ELF | - | 175 ± 47/25 | 150 ± 43/35 | - | - | - | - | - | - |
| SCH1 | 270 ± 54/24 | - | - | - | - | - | - | - | - |
| SCH2 | - | 128 ± 35/14 | - | - | - | - | - | - | - |
| mean ± SD | 199 ± 42 | 133 ± 32 | 131 ± 29 | 25 ± 8 | 27 ± 15 | 21 ± 11 | 15 ± 4 | 13 ± 4 | 23 ± 17 |
| n total | 50 | 70 | 74 | 89 | 89 | 59 | 28 | 34 | 10 |
| Species | WphI | WphII | WphIII |
|---|---|---|---|
| Pinus cembra | 5.35 ± 0.84 a | 1.89 ± 1.78 b | 4.84 ± 1.33 a |
| Larix decidua | 4.49 ± 1.91 a | 4.65 ± 0.54 a | 6.66 ± 1.05 b |
| Picea abies | 4.24 ± 0.82 a | 4.28 ± 1.23 a | 10.57 ± 1.22 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oberhuber, W.; Bendler, U.; Gamper, V.; Geier, J.; Hölzl, A.; Kofler, W.; Krismer, H.; Waldboth, B.; Wieser, G. Growth Trends of Coniferous Species along Elevational Transects in the Central European Alps Indicate Decreasing Sensitivity to Climate Warming. Forests 2020, 11, 132. https://doi.org/10.3390/f11020132
Oberhuber W, Bendler U, Gamper V, Geier J, Hölzl A, Kofler W, Krismer H, Waldboth B, Wieser G. Growth Trends of Coniferous Species along Elevational Transects in the Central European Alps Indicate Decreasing Sensitivity to Climate Warming. Forests. 2020; 11(2):132. https://doi.org/10.3390/f11020132
Chicago/Turabian StyleOberhuber, Walter, Ursula Bendler, Vanessa Gamper, Jacob Geier, Anna Hölzl, Werner Kofler, Hanna Krismer, Barbara Waldboth, and Gerhard Wieser. 2020. "Growth Trends of Coniferous Species along Elevational Transects in the Central European Alps Indicate Decreasing Sensitivity to Climate Warming" Forests 11, no. 2: 132. https://doi.org/10.3390/f11020132