Fall Ethephon Application Enhances the Freezing Tolerance of Magnolia wufengensis During Overwintering

1
Ministry of Education Key Laboratory for Silviculture and Conservation, College of Forestry, Beijing Forestry University, Beijing 100083, China
2
Fujian Forestry Investigation and Planning Institute, Fuzhou 350003, China
3
Biotechnology Research Center, Three Gorges University, Yichang 443002, China
4
Forestry Bureau of Wufeng County, Wufeng 443400, China
*
Author to whom correspondence should be addressed.
†
Deceased on 18 April 2018.
Forests 2019, 10(10), 868; https://doi.org/10.3390/f10100868
Submission received: 7 August 2019
/
Revised: 24 September 2019
/
Accepted: 27 September 2019
/
Published: 3 October 2019
Abstract
:Magnolia wufengensis L.Y. Ma et L. R. Wang, a rare species which has been introduced and cultivated from southern China to northern China, frequently suffers from freezing injuries. To figure out the influence of ethephon (ETH) application on the cold tolerance during the natural overwintering of M. wufengensis, one-year shoots subjected to ethephon application at four concentrations (0, 700, 1000, and 1500 mg·L−1) were collected to measure the physiological and biochemical changes from September 2017 to 1 April 2018. The fall ETH application increased the freezing tolerance of M. wufengensis, and the optimum concentration for M. wufengensis was 1000 mg·L−1 (T2), which not only improved the shoot freezing tolerance by 1.4 times, but also led to a 25.0% faster cold acclimation rate and 13.7% slower de-acclimation rate. Moreover, 1000 mg·L−1 ETH delayed bud-burst in the spring by 10. 7 d, which was helpful for effectively avoiding cold spells in the spring, and improved the rates of bud survival by 47.1%. The improved freezing tolerance under exogenous ETH application was associated with an increased dehydration and accumulation of proline content. It seems that exogenous ETH application may be used on M. wufengensis grown in northern China to protect against freezing tolerance during the overwintering period.
1. Introduction
Magnolia wufengensis, a new Magnolia species (Magnoliaceae), was discovered in natural secondary forests growing with other native species in Wufeng county, Hubei Province, southern China [1,2]. However, M. wufengensis has a narrow distribution range because of biological characteristics and anthropogenic disturbance, with only 2000 individuals making up the wild community [3]. More seriously, fragmented habitation makes it much more difficult for M. wufengensis to be renewed under natural conditions, leaving it in a critically endangered state [3]. Therefore, it is necessary and urgent to preserve and expand the geographic distribution of M. wufengensis. With years of efforts, on the one hand, we have preserved the wild environment of Wufeng, and have bred Jiaohong No.1, which represents a new of cultivar with striking red flowers (Figure 1) [3]. Increasingly popular for urban greening, M. wufengensis has considerable ornamental and economical value owing to its colorful flowers (pure, dark, or pale red) and varied flower petal numbers (9–25, 32, or 46) [4]. Additionally, M. wufengensis has an extremely economical value because essential oils can be extracted from its flowers, whose main active components include sterols, n-alkanes, and terpenes. In addition, it has been widely used for the afforestation of barren hills in an area of over 45,000,000 m2 in Wufeng county. At the same time, the wood quality of this species is excellent because of its high and straight trunk (its height can reach 25 m), and little branches. Besides, in Chinese furniture markets, it has been frequently used to make furniture, drawing boards, and joinery, which also reflect its excellent wood characteristics (great material, straight texture, and fine structure). Moreover, it can be used as timber-making species. On the other hand, our group have introduced M. wufengensis to many provinces in China. Based on what has and is being done, we found that a high temperature and radiation during the growing seasons were mainly restraining factors in southern China (such as Jiangsu Province and Guangdong Province, data has still not been published yet). Moreover, compared with southern China, large ornamental trees, especially arbors, are very scarce in northern China. It would be of greater significance both in terms of the forestry greening value and economic value than planting in southern China, if M. wufengensis could be introduced successfully. In addition, the capital of China, Beijing, is proposing “Enriching Coloration and Prolonging Green Period of Beijing” to build a more beautiful and greener city. Therefore, M. wufengensis is regarded as a great arbor species and material owing to its amazing ornamental value, and has been introduced to northern China in recent years. However, freezing injuries have been a major factor restraining the sustainability and profitability of M. wufengensis seedling production and have caused huge economic losses in northern China [5,6,7,8].
Freezing injuries adversely affect the growth, productivity, and geographical distribution of horticultural plants [9,10]. The susceptibility of plants to freezing injuries may be due not only to an insufficient freezing tolerance, but also to the timing and rate of cold acclimation and de-acclimation [11]. Cold acclimation, the process by which plants transit from a cold-sensitive to cold-hardy state [12], is essential for the survival of woody plants growing in temperate regions [13]. Cold acclimation usually develops in two stages. In the first stage, the short-day condition induces growth cessation, leaf senescence, and a certain development of freezing tolerance [14,15,16]. The decreasing air temperature observed in the second stage increases the freezing tolerance to the maximum level [17]. Freezing tolerance decreases during the de-acclimation period [18,19,20]. A series of physiological and biochemical responses associated with an increasing freezing tolerance include modifications of the membrane lipid composition, a decreased tissue water content, the accumulation of proline, soluble protein, sugars, and hormone regulation [18,19,20]. In general, the changes which occur during cold acclimation are reversed during de-acclimation.
Ethephon is a kind of plant growth regulator which functions by releasing ethylene when absorbed into plants (pH > 4 [21]). The phytohormone ethylene takes part in many complex physio-chemical processes. In addition, it is closely associated with biotic stresses and abiotic stresses, including pathogen attacks [22], drought stress [23], salt stress [24], and cold stress [25]. The specific role of ethylene’s response to cold stresses and cold acclimation may vary among species. Some studies have reported that ethylene negatively regulated the cold acclimation-induced freezing tolerance of alfalfa seedlings [26], arabidopsis [27], and Bermuda grass [28]; on the contrary, more studies have concluded that increases in ethylene contents cause cold tolerance to be enhanced, including that of wheat [29], tobacco [30], the Musa AAA Cavendish subgroup [31], Secale cereale (rye) [32], tomato [33], and Phaseolus spp. (bean) [34].The mechanism of ethephon’s regulation of cold tolerance is ethylene response factor (ERF) genes, which trigger many cold-response pathways by controlling the expression of downstream genes [30] and related downstream pathways, including plant senescence [34], antioxidant enzymes [35], ice nucleation activity, and the patterns of osmotic proteins [35].
However, to the best of our knowledge, no published reports are available on the influence of exogenous ethephon (ETH) application to Magnolia plants, especially on the cold resistance of M. wufengensis. Therefore, this study begins to focus on this unknown field. Physiological and biochemical responses of field-grown M. wufengensis seedlings to root application ETH treatments during cold acclimation were investigated in this study. The following questions were addressed: (1) Is the role of ethylene in the freezing tolerance of M. wufengensis seedlings positive or negative? (2) What is the optimum ETH application concentration for ultimately improving the freezing tolerance of M. wufengensis? (3) Does ETH affect both acclimation and de-acclimation processes in M. wufengensis?
2. Materials and Methods
2.1. Plant Materials and Experimental Treatments
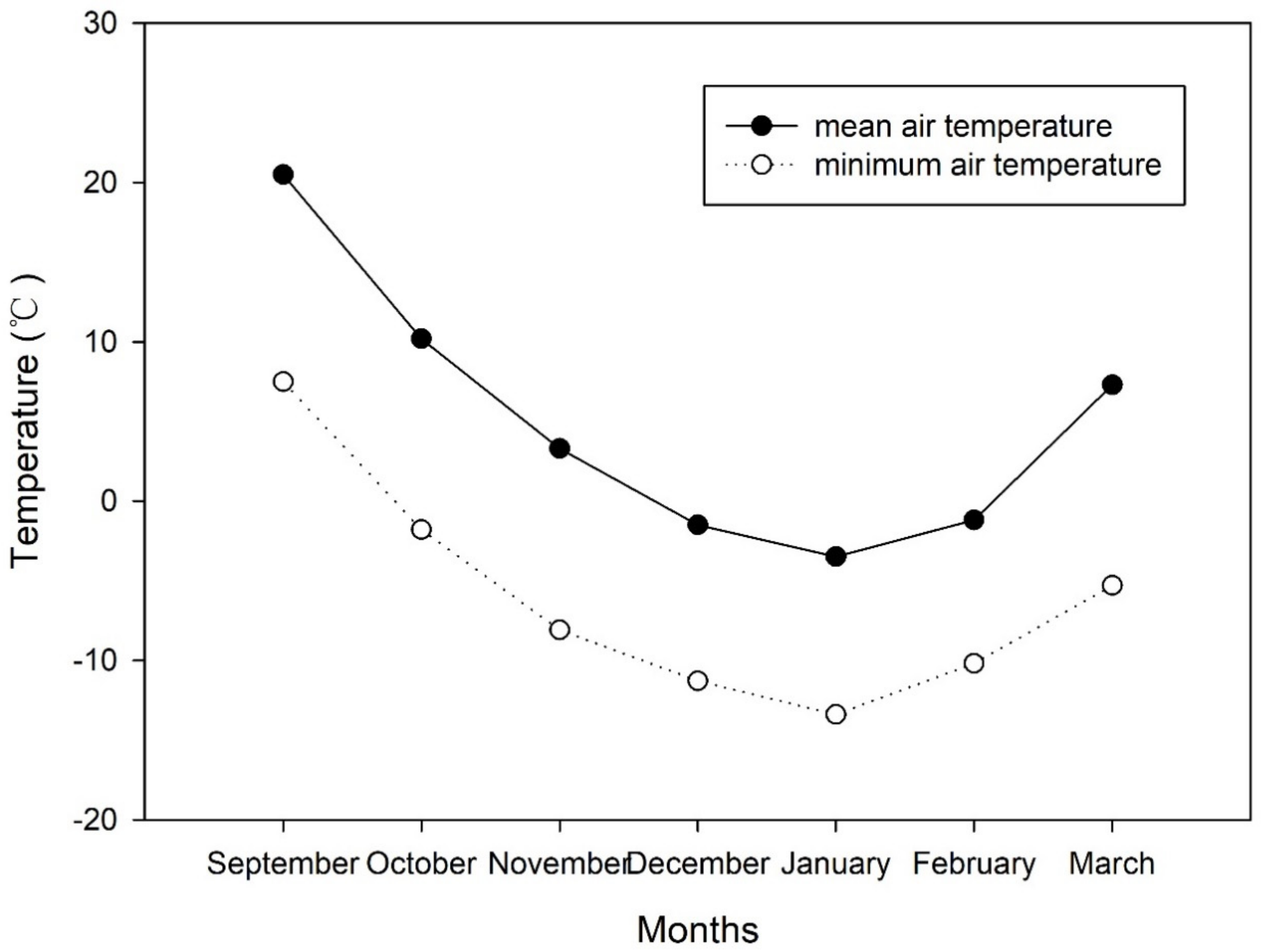
The Dongdadi Experimental Base of Beijing University of Agriculture in Beijing, China (39°48′ N, 116°28′ E), was used in this study. The air temperature data from September 2017 to March 2018 that we recorded are shown in Figure 2. The soil type was cinnamon soil.
In early April 2017, 1-year-old M. wufengensis seedlings from the Rare Plant Institute of Forestry Bureau of Wufeng County in Wufeng, Hubei Province, China (29°56′ N, 110°15′ E), were planted at our experimental site. The plants were grown in five plots at our study site. Each plot (about 10 m × 3 m) contained 20 plants and the planting space was 0.5 m × 0.5 m. From April to 31 August, the seedlings were kept well-irrigated and protected against bacterial pathogens and weed competition. The average height of the seedlings was 524.0 mm, and the average ground diameter of them was 11.4 mm. In our pre-experiment, the optimum time for ETH application was about early September, and ETH concentrations above 1600 mg·L−1 were observed to cause leaf damage (data not shown). Therefore, the seedlings were subjected to four concentrations of ETH, consisting of 0 mg·L−1 (T0), 750 mg·L−1 (T1), 1000 mg·L−1 (T2), and 1500 mg·L−1 (T3), between 16:00 and 18:00 on the 1, 11, and 21 of September. The ETH solution with 40% active ingredients was provided by Sichuan Guoguang Agrochemical Co., Ltd. (Sichuan, China). Each treatment involved three replications. Whole seedlings were sprayed with ETH solutions to runoff with a 5 L handheld sprayer averaging a spray volume of 0.5 L/seedling.
2.2. Determination of Shoot Growth
Shoot length was measured every 3 days from 1 to 30 September. Ten representative seedlings under each treatment were selected and one healthy upper-crown shoot facing the sun on each representative seedling was selected for shoot length measurement by steel tape. The same sampling method was applied to D50B (days to 50% bud-burst), LT50 (50% relative electrolyte leakage), water content, proline content, and days to terminal bud-burst. The relative growth rate (RGR) and days to growth cessation (DGC) of shoot length were measured according to the method of Yang et al. [5,6,7,8]. In brief, the RGR of shoot length was determined by the formula, RGR = (lnv2 − lnv1)/(t2 − t1), where v1 is the value of shoot length on the first measuring day, v2 is the value of shoot length on the last measuring day, t1 is the first measuring day (1 September), and t2 is the last measuring day (30 September). Days to growth cessation (DGC) of shoot length, assessed as the number of days since 1 September to shoot growth cessation, were recorded every three days.
2.3. Determination of Leaf Senescence
For leaf senescence, assessed as the leaf chlorophyll content, ten representative seedlings under each treatment were selected and one healthy upper-crown leaf facing the sun on each seedling was selected for shoot growth measurement, which was determined once per 10 d from 1 September to 31 October using SPAD-502 (Konica Minolta, Japan).
2.4. Determination of Bud Dormancy
Bud dormancy, assessed as days to 50% bud-burst (D50B), was determined monthly from 30 September 2017 to 30 March 2018. The D50B was measured according to the method of Yang et al. [6,8]. In brief, the trays were placed on benches under forcing conditions in a growth chamber (Saifu Corporation, Ningbo, P. R. China) with the following settings: a 24 h photoperiod at 500 µmol m−2s−1 PPFD, 22 °C, and 80% relative humidity. Bud-burst (%) was recorded every day.
2.5. Determination of Shoot Freezing Tolerance
The freezing tolerance of shoots, assessed as the lowest temperature at which 50% injury occurred (LT50), was determined monthly from 30 September 2017 to 30 March 2018. The LT50 was measured according to the method of Yang et al. [5,6,7,8]. In brief, the tubes were incubated in a circulating water bath equipped with a temperature controller (DC1030; Saifu, P.R. China) and cooled at a rate of 2 °C·h−1 until they reached 0 °C. There were seven designed temperatures, including 0, −5, −10, −15, −20, −25, and −30 °C, in our freezing experiment. After being maintained at 0 °C for 1 h, the tubes were cooled at a rate of 5 °C·h−1 until each target temperature was reached, and were maintained for 2 h at each target temperature. LT50 was estimated from an asymmetric sigmoid curve constructed with the Gompertz function, fitted to the percent injury data calculated by the analyzed electrolyte leakage according to the method of Jun et al. [36].
2.6. Determination of Physiological and Biochemical Parameters in the Shoots
The water content and proline content in the shoots were determined monthly from 30 September 2017 to 30 March 2018. The water content was calculated using the formula, water content = (fresh weight − dry weight)/fresh weight. In brief, collected fresh shoots were quickly cut into small pieces and placed in an aluminum box (dried and weighed in advance), and the weight was accurately recorded with an analytical balance. They were dried in an oven at 105 °C for 2 h, and then dried at 80 °C until reaching a constant weight. The proline content was measured according to the method of Bates et al. [37], with a little modification. In brief, freeze-dried shoot material (8 to 20 mg dry weight) was boiled for 10 min in 2.5 mL of 3% aqueous sulfosalicylic acid, and was then cooled and filtered through filter paper. A total of 2 mL of filtrate was reacted with 2 mL of acid ninhydrin and 2 mL of glacial acetic acid in a test tube for 30 min at 100 °C. This was then terminated in an ice bath and 4 mL of toluene was mixed vigorously for 20 sec and centrifuged at 3000 r/min for 5 min. Next, supernatant was collected, and the absorbance was read at 520 nm, using toluene as a blank.
2.7. Determination of Bud-Burst in Spring
Bud-burst was recorded each day for all seedlings from 1 April 2018 onwards. Days to terminal bud-burst was recorded, and final bud survival rates were calculated.
2.8. Statistical Analysis
All data were analyzed using SPSS Statistics 18.0, including a one-way analysis of variance for the main effects of different treatments and a correlation analysis. All tables and figures were produced using Microsoft Word 2016 and Sigmaplot 12.5, respectively.
3. Results
3.1. Shoot Growth
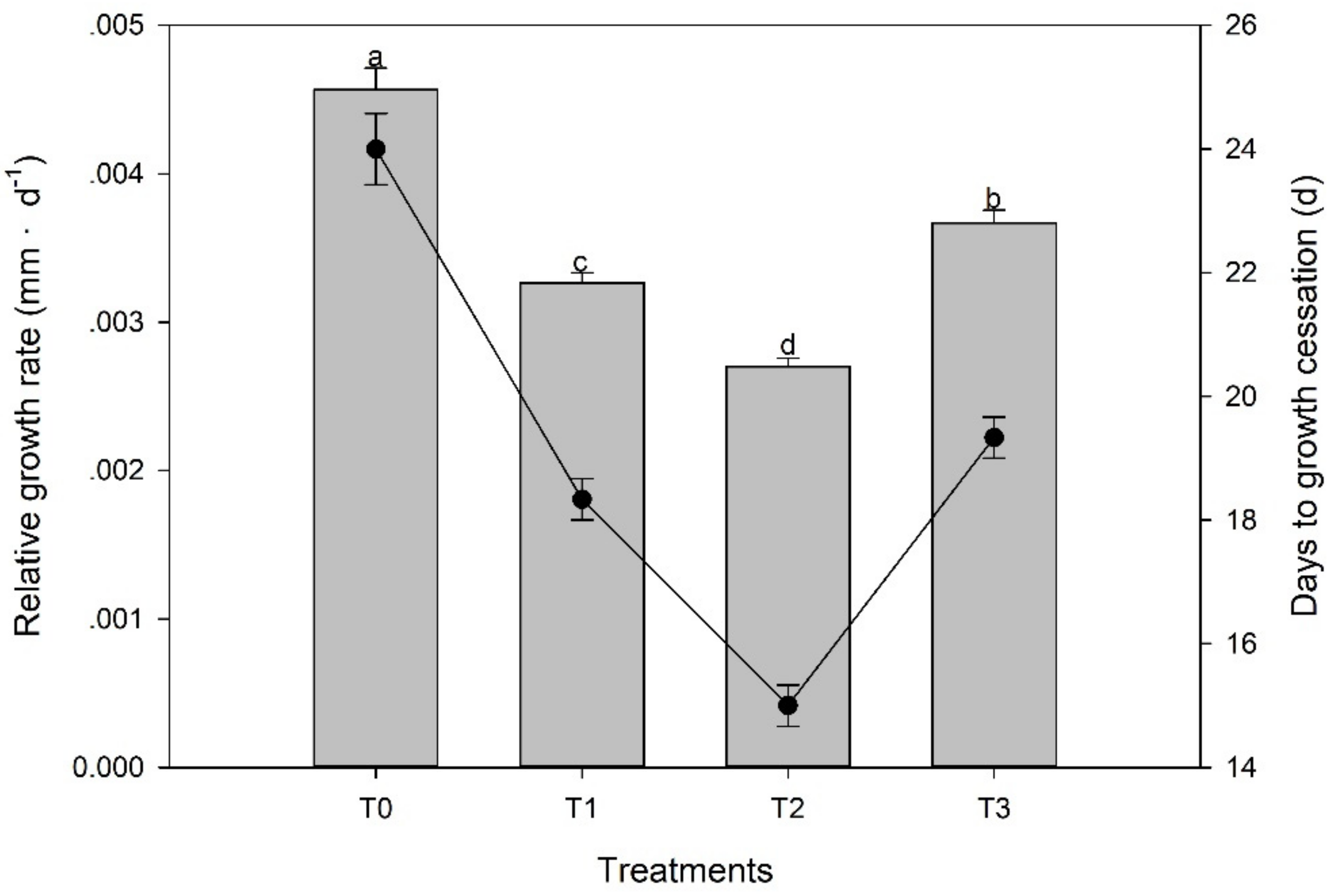
Compared with the seedlings under T0 (Figure 3), exogenous ethephon applications decreased both the RGR and DGC values of M. wufengensis, and the RGR of shoot length decreased by 0.0019 mm·d−1 under T2. The shoots under T2 stopped elongating before 15 September, while it took 9 d longer for T0-treated shoots to stop the growth of seedlings than T2-treated shoots.
3.2. Leaf Senescence
Chlorophyll content-treated ETH decreased after the first application and was lower, while leaf chlorophyll content under T0 remained the same from 1 to 11 September, and then decreased after 11 September (Figure 4). Compared with T0, the range of decline of chlorophyll content was 20.8%, 31.3%, and 14.2% larger under T1, T2, and T3, respectively. The decreased rates of chlorophyll content between 1 September and 31 October were 34.0%, 51.8%, and 23.9% faster under T0, T1, T2, and T3, respectively. Based on the chlorophyll content, the degree and rate of leaf senescence were as follows: T2 > T1 > T3 > T0.
3.3. Effect of ETH on Bud Dormancy
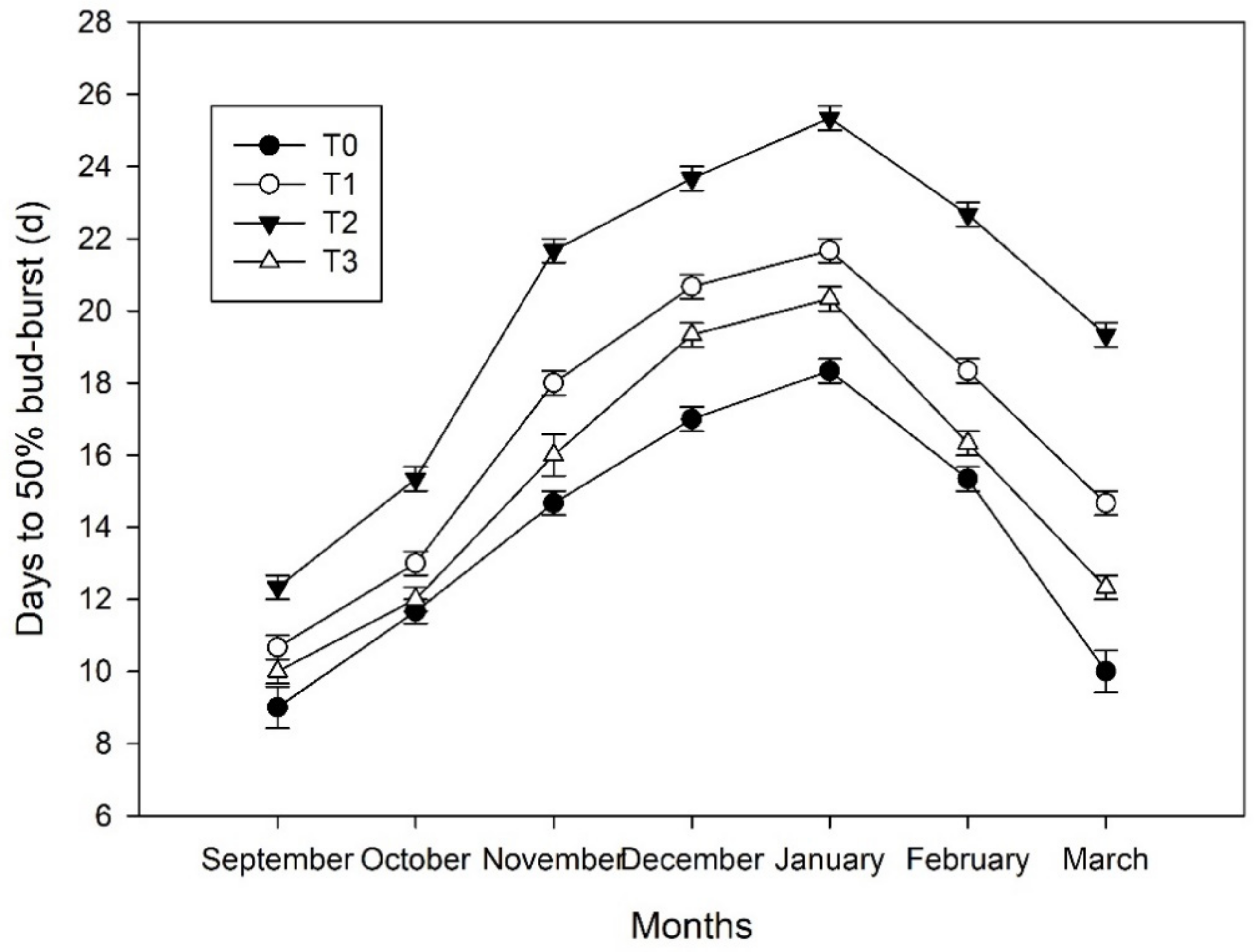
Compared with T0, the D50B values of seedlings treated with ETH were significantly higher during the whole experimental period. The trend of D50B was similar to that of air temperature (Figure 2 and Figure 5). D50B under each treatment continued to increase during cold acclimation and reached the maximum value in January (D50B of 18, 22, 25, and 21 d under T0, T1, T2, and T3, respectively), after which it decreased during de-acclimation (Figure 5). The increased rates of D50B during cold acclimation were 1.9, 2.2, 2.6, and 2.1 d per month under T0, T1, T2, and T3, respectively. Decreased rates of D50B during de-acclimation were 4.2, 3.5, 3.0, and 4.0 d per month under T0, T1, T2, and T3, respectively. Based on the D50B values (seedlings in growth chamber), the seedlings under T2 (ETH 1000 mg·L−1) displayed the significantly deepest bud dormancy levels, followed by T1, T2, and T0.
3.4. Freezing Tolerance in the Shoots
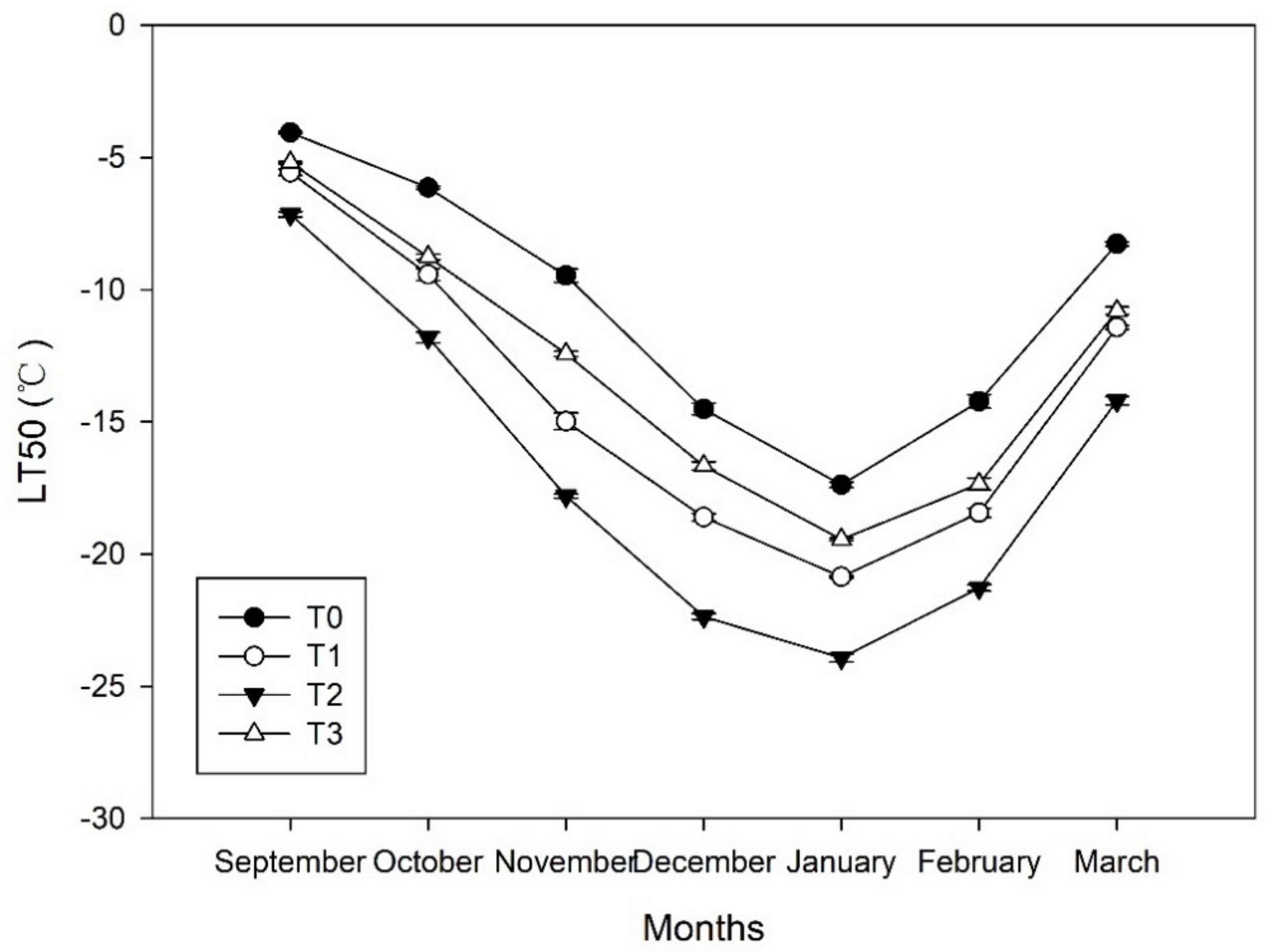
The LT50 values of the shoots treated with ETH were significantly lower than those of the shoots under T0 during overwintering. Specifically, LT50 values under each treatment changed coincidently with air temperature (Figure 2), which continued to decrease during cold acclimation, and reached the minimum value in January (LT50 of −17.4, −20.9, −23.9, and −19.5 °C under T0, T1, T2, and T3, respectively), before then increasing (Figure 6). The decreased rates of LT50 during cold acclimation were 2.7, 3.0, 3.4, and 2.9 °C per month under T0, T1, T2, and T3, respectively. Based on the LT50 values, the freezing tolerance of the seedlings could be ordered as T2 > T1 > T3 > T0 during overwintering.
3.5. Effect of ETH on Biochemical Parameters in Shoots
During the entire overwintering period, seedlings treated with ETH exhibited significantly lower water values than those treated with T0. The tendency of the water content in the shoots under each treatment to change was highly similar to that of LT50, and the water content reached the minimum value in January (the values of 51.2%, 43.9%, 37.9%, and 46.9% under T0, T1, T2, and T3, respectively), before then increasing (the values of 65.5%, 58.7%, 53.7%, and 60.0% under T0, T1, T2, and T3, respectively) (Figure 7). The water content significantly declined under T2 (ETH 1000 mg·L−1) from late September 2017 to late January 2018. The decreased rates of water content during cold acclimation were 6.1%, 5.3%, 7.1%, and 5.4% per month under T0, T1, T2, and T3, respectively. The water contents could be ordered as T0 > T3 > T1 > T2.
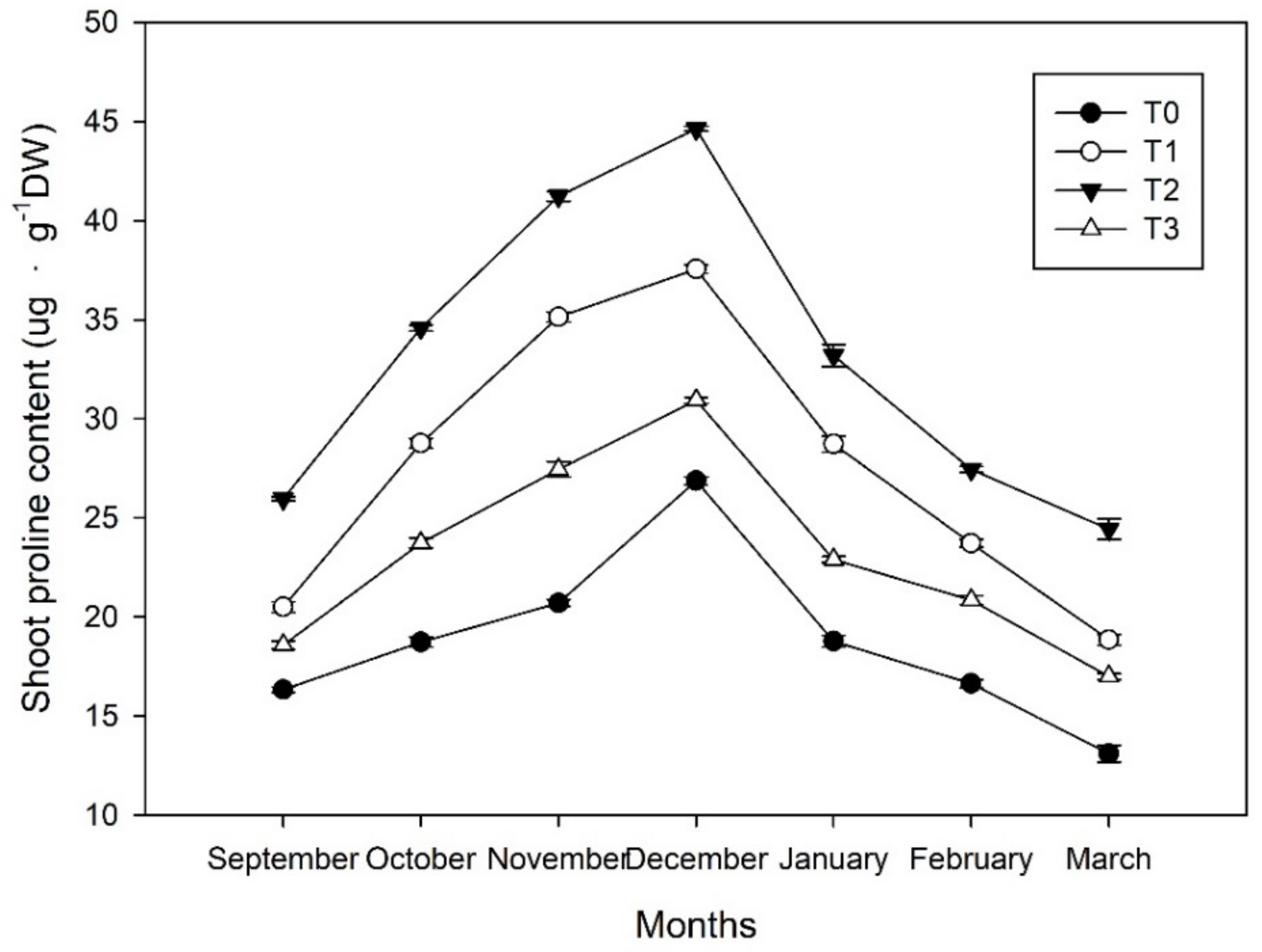
The proline content in the shoots under each treatment increased and reached its maximum level in December, before then decreasing (Figure 8). The shoots’ maximum proline content levels (26.9, 37.6, 44.6, and 30.9 ug·g−1 dry weight (DW) under T0, T1, T2, and T3, respectively). In March 2018, the values were 13.1, 18.8, 24.4, and 17.0 ug·g−1 DW under T0, T1, T2, and T3, respectively. All ethephon treatments could accelerate proline accumulation, while T2 was much more effective than the others. Proline could be ordered as T2 > T1 >T3 > T0.
3.6. Effect of ETH on Bud-Burst in Spring
As seen in Figure 11, compared to T0, all ethephon applications significantly delayed (p < 0.05) bud-burst 3.7, 10.7, and 6.4 d under T1, T2, and T3, respectively (Figure 9). In addition, three concentrations of ETH significantly (p < 0.05) increased the bud survival rates by 18.6%, 32.0%, and 15.7% under T1, T2, and T3, respectively (Figure 9). To sum up, days to bud-burst and the bud survival rate could be ordered as T2 > T1 >T3 > T0.
4. Discussion
Vegetative growth in late fall has been shown to negatively correlate with winter-hardiness [38]. Our experimental results showed that the exogenous ethephon inhibited shoot growth of M. wufengensis (Figure 3). The effects of ETH on growth are linked to the interactions between ETH and other plant hormones [28]. Additionally, ETH treatments of 1000 mg·L−1 advanced the growth cessation of M. wufengensis seedlings by 9 d (Figure 3). This suggests that the seedlings under this ETH concentration underwent cold acclimation earlier, because growth cessation is an early physiological step in the cold acclimation process [39].
Leaf senescence is another early process and sign of cold acclimation [40], which remobilizes nutrient substances from leaves to overwintering tissues like shoots. The degradation of chlorophyll occurs in mesophyll cells during leaf senescence [41]. In our experiment, ethephon application significantly accelerated the degradation of chlorophyll of M. wufengensis seedlings (Figure 4), and a similar function of inducing leaf senescence by ethephon has been verified in previous studies [28,42,43]. In addition, the leaf senescence process induced by ethephon may depend on the initial application date, frequency, concentrations, and intervals [42]. Senescence is a combination function of ethylene and cytokinin ratios [28]. In addition, in Arabidopsis thaliana, ethylene accelerates leaf senescence by directly activating chlorophyll catabolic genes which shed light on the primary molecular mechanism between ethylene and leaf senescence [43]. In this study, faster leaf senescence under T2 (ETH 1000 mg·L−1) may indicate stronger freezing tolerance at an earlier point in time. This is because faster senescence tends to imply better and more adequate cold acclimation [40]. In addition, cold-hardy species usually exhibit leaf senescence earlier than cold-sensitive species [44].
According to the inhibitory sources, bud dormancy in woody species can be divided into three categories: para-dormancy (inhibition from distal organs), endodormancy (inhibition from internal bud signals), and eco-dormancy (inhibition from unfavorable environmental conditions) [45]. Buds initially go through the endodormancy phase and then the eco-dormancy phase during cold acclimation after the chilling requirement has been satisfied. It can be concluded that the ETH application of 1000 mg·L−1 could induce the endodormancy of M. wufengensis (Figure 5), because single cuttings (not para-dormant) were used in favorable growing environments (not eco-dormant) [46]. Although there was a lack of direct data, many researchers have reported that ethephon could alter the chilling/heat requirement of buds, affect bud development, prolong the period of bud dormancy, and delay bloom effectively [47,48,49]. Deeper dormancy during overwintering was related to more cold-hardiness [8]. This result may mean that seedlings sprayed with 1000 mg·L−1 ETH (T2) are more tolerant than the others.
Similar to other woody perennial plants, the freezing tolerance of M. wufengensis was strengthened during cold acclimation, and peaked in January [5,6,7,8,50]. Based on LT50 values, ETH application could enhance the freezing tolerance of seedlings during overwintering, of which 1000 mg·L−1 ETH (T2) was the most effective concentration (Figure 6). Exogenous ethylene application has previously been reported to enhance the freezing tolerance in many plants [29,30,31,32,33,34,51]. In addition, relatively more and severer freezing damage in one-year shoots of the seedlings without ETH application was observed in midwinter January (visual observation), indicating the effectiveness of ETH for increasing the freezing tolerance of M. wufengensis.
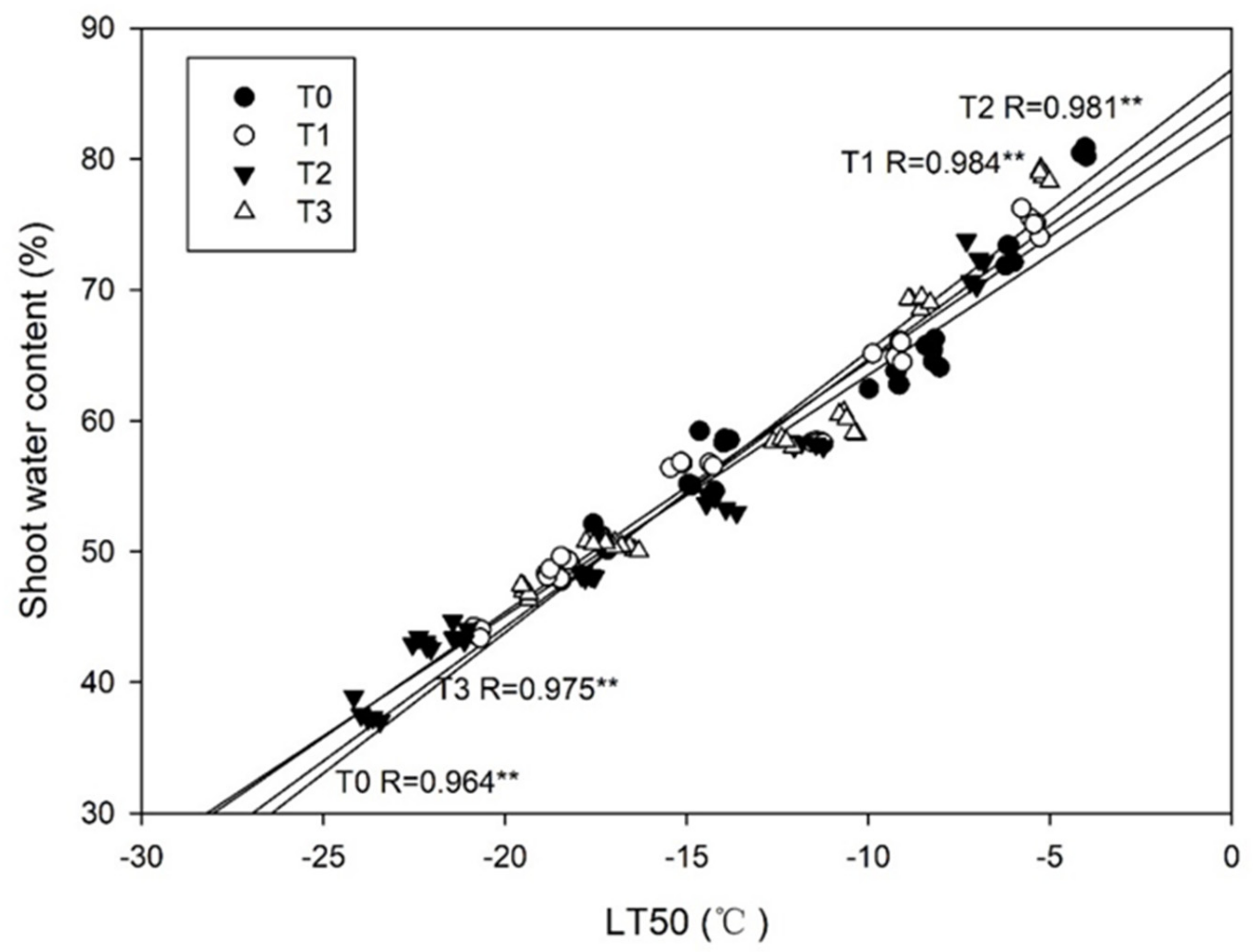
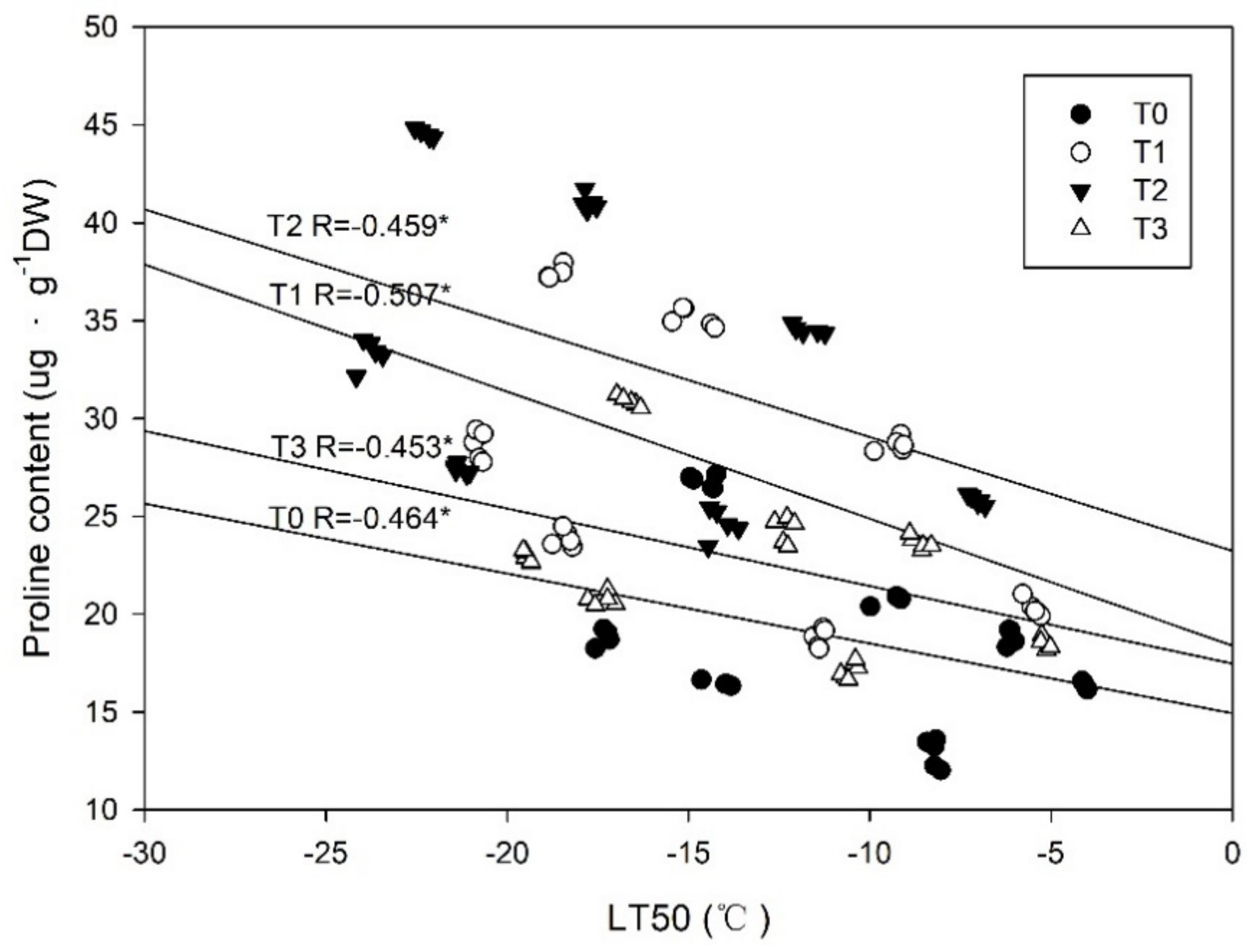
An increased freezing tolerance is commonly connected to a reduced water content [52], which was well-observed in our study. A significant relationship between water content and freezing tolerance in M. wufengensis seedlings was detected (Figure 10). Plant cells are usually dehydrated under the extracellular freezing process, and a dehydration-induced freezing tolerance has been well-illustrated [10]. Therefore, the observed differences in water content among treatments may explain, at least partly, the superior freezing tolerance under an ETH application of 1000 mg·L−1. It has been demonstrated in four bermudagrass cultivars that water stress in late autumn can help plants acquire early cold acclimation and dormancy, which may also be the consequence of the induced abscisic acid (ABA) synthesis caused by water stress [53]. In this paper, freezing tolerance was significantly correlated with proline content in M. wufengensis seedlings, which was consistent with [35,51] (Figure 11). Proline plays multiple roles in plant stress tolerance, as a mediator of osmotic adjustment, a stabilizer of proteins and membranes, an inducer of osmotic stress-related genes, and a scavenger of reactive oxygen species (ROS) [54,55,56,57]. Hence, the higher proline content under the ETH application of 1000 mg·L−1 also explained the increased freezing tolerance.
In general, early-bloom buds are more susceptible to spring frost damage than later-bloom buds. Exogenous ETH application could decrease the rate of de-acclimation, delaying the period of bud dormancy and bloom [48,49,58]. In this study, ETH prolonged bud-burst and improved the bud survival rate to different extents, which would protect buds against spring frost.
In sum, in recent years, ethylene (or ethephon) has attracted more and more attention with the development of molecular biotechnology. Previous studies have mostly focused on how ethylene or ethylene influences plants under an artificial low temperature of an artificial environment [27,30,34,35] (Table 1). They have also included early studies on the freeze resistance of ethylene to plants, while ignoring the physiological effects of ethylene on plants during the natural overwintering period [26,28,29,31,32,34] (Table 1). This study was the first study about ethephon on Magnoliaceae plants during the entire natural wintering period, which included the effects of ethephon application in late autumn on the growth cessation, leaf senescence, freezing resistance, and the survival rate of winter buds after spring, so it has a very strong practical significance and guiding role. According to the results, in addition, this study not only concluded that ethephon application at the end of autumn could enhance the freezing resistance in winter, but also explained its intrinsic physiological mechanism, that is, by reducing the water content and promoting the accumulation of proline (small molecular substances), so it preliminarily explored the intrinsic mechanism of ethephon improving the wintering performance of M. wufengensis.
5. Conclusions
Freezing injuries have been a restricting factor limiting whether Magnolia wufengensis can be planted in certain areas. A high freezing tolerance could effectively protect plants against freezing tolerance during the natural overwintering period, which includes mid-winter extremely low temperatures and cold spells in the spring. The freezing injuries tend to happen to delicate stems and young shoots, especially tips with growing points. The branches of young trees suffering from freezing injuries stop growing, and renewed growth in the next year will affect the normal growth of trees, reduce the biomass, and may also decrease the wood quality, which will affect the harvest time and quality of wood cultivated as timber forest. Therefore, a high freezing tolerance can improve the survival rate of afforestation, shorten the harvest time of wood, and improve the quality of wood in the future. ETH treatment could have a positive role in enhancing the freezing tolerance, growth, and afforestation of M. wufengensis. Firstly, late-autumn ETH treatments could enhance the freezing tolerance and increase the survival rate, which are conducive to tree growth. In general, injured and poorly growing plants tend to be more vulnerable to infection with other adverse conditions, including various viruses and fungi biotic stresses, as well as abiotic stresses. Therefore, the high freezing tolerance of plants obtained by ETH treatment could decrease the risk of injury or sickness and help to maintain tree health. Secondly, ETH treatment could accelerate shoot growth cessation and leaf senescence, and fasten cold acclimation progress, which includes periderm formation. It may have some impact on the wood chain. Thirdly, ETH treatment could have a long and positive role in freezing tolerance which can be maintained throughout the natural overwintering period, representing a convenient, effective, and practical method to apply on a large scale.
In conclusion, ETH foliar application has a positive role in enhancing the freezing tolerance of M. wufengensis, and the optimum concentration in this study was 1000 mg·L−1, which initiated a cascade of steps for advancing the cold acclimation process of M. wufengensis. An improved freezing tolerance under ETH application was associated with a water content decrease and the accumulation of proline. We suggest that foliar ETH application may be used on M. wufengensis grown in northern China to improve its overwintering performance where there are short growing seasons and early fall frost events.
Author Contributions
L.M., conceived and designed the experiments, and supervised the work; F.C., coordinated and supervised the work; Z.S., coordinated and supervised the work, partly supported the final experiment; Y.Y., designed the experiments and supervised the work; C.C., partly conducted the experiments, ran data analyses, and made the graphs and table; X.D., designed the experiments, conducted the experiments, ran data analyses, and wrote the paper.
Funding
This research was financially supported by the Special Fund for Forest Scientific Research in the Public Welfare (Grant No. 201504704).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ma, L.Y.; Wang, L.R.; He, S.C.; Liu, X.; Wang, X.Q. A new species of Magnolia (Magnoliaceae) from Hubei, China. Bull. Bot. Res. 2006, 26, 4–7. [Google Scholar]
- Ma, L.Y.; Wang, L.R.; He, S.C.; Liu, X.; Wang, X.Q. A new variety of Magnolia (Magnoliaceae) from Hubei, China. Bull. Bot. Res. 2006, 26, 517–519. [Google Scholar]
- Sang, Z.Y.; Ma, L.Y.; Chen, F.J.; Zhang, P.; Zhu, Y.C. Protection status and utilization countermeasure of germplasm resources of the Magnolia wufengensis in Wufeng County. Hubei Agric. Sci. 2011, 50, 1564–1567. [Google Scholar]
- Xiao, A.H.; Ma, L.Y.; Sang, Z.Y.; Chen, F.J. Magnolia wufengensis ‘Jiaohong No. 1′: A Magnolia Cultivar with Striking Red Flowers. HortScience 2018, 53, 573–574. [Google Scholar] [CrossRef]
- Yang, Y.; Jia, Z.K.; Chen, F.J.; Sang, Z.Y.; Ma, L.Y. Comparative analysis of natural cold acclimation and de-acclimation of two Magnolia species with different winter hardiness. Acta Physiol. Plant 2015, 37, 129. [Google Scholar] [CrossRef]
- Yang, Y.; Jia, Z.K.; Chen, F.J.; Sang, Z.Y.; Duan, J.; Ma, L.Y. Natural cold acclimatisation and de-acclimatisation of Magnolia wufengensis in response to alternative methods of application of abscisic acid. J. Hortic. Sci. Biotechnol. 2015, 90, 704–710. [Google Scholar] [CrossRef]
- Yang, Y.; Jia, Z.K.; Chen, F.J.; Sang, Z.Y.; Ma, L.Y. Physiological and biochemical processes of Magnolia wufengensis in response to foliar abscisic acid application during natural cold acclimation. HortScience 2015, 50, 387–394. [Google Scholar] [CrossRef]
- Yang, Y.; Jia, Z.K.; Chen, F.J.; Sang, Z.Y.; Duan, J.; Ma, L.Y. Effect of exogenous abscisic acid on cold acclimation in two Magnolia species. Biol. Plant. 2016, 60, 555–562. [Google Scholar] [CrossRef]
- Weiser, C.J. Cold resistance and injury in woody plants. Science 1970, 169, 1269–1278. [Google Scholar] [CrossRef]
- Pearce, R.S. Plant freezing and damage. Ann. Bot. 2001, 87, 417–424. [Google Scholar] [CrossRef]
- Suojala, T.; Lindén, L. Frost hardiness of Philadelphus and Hydrangea clones during ecodormancy. Acta Agric. Sca. B Soil Plant Sci. 1997, 47, 58–63. [Google Scholar]
- Zabadal, T.J.; Dami, I.E.; Goffinet, M.C.; Martinson, T.E.; Chien, M.L. Winter Injury to Grapevines and Methods of Protection; Michigan State University Extension Bulletin: East Lasing, MI, USA, 2007; p. 2930. [Google Scholar]
- Teets, T.M.; Hummel, R.L.; Guy, C.L. Cold-acclimation of Hibiscus rosasinensis L. and Hybiscus syriacus L. in natural and controlled environments. Plant Cell Environ. 1989, 12, 495–502. [Google Scholar] [CrossRef]
- Davis, J.D.; Evert, R.F. Seasonal cycle of phloem development in woody vines. Bot. Gaz. 1970, 131, 128–138. [Google Scholar] [CrossRef]
- Wolpert, J.A.; Howell, G.S. Cold acclimation of concord grapevines III. Relationship between cold hardiness, tissue water content, and shoot maturation. Vitis 1986, 25, 151–159. [Google Scholar]
- Palva, E.T.; Thtihariju, S.; Tamminen, I.; Puhakdinen, T.; Laitinen, R. Biological mechanisms of low temperature stress response: Cold acclimation and development of freezing tolerance in plants. Jpn. Intl. Res. Cent. Agric. Sci. 2002, 23, 9–15. [Google Scholar]
- Artlip, T.; Wisniewski, M.E. Tissue specific expression of a dehydrin gene in one-year-old ‘Rio Oso Gem’ peach trees. J. Am. Soc. Hortic. Sci. 1997, 122, 784–787. [Google Scholar] [CrossRef]
- Guy, C.L. Cold acclimation and freezing stress tolerance: Role of protein metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 187–223. [Google Scholar] [CrossRef]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef]
- Welling, A.; Palva, E.T. Molecular control of cold acclimation in trees. Physiol. Plant 2008, 127, 167–181. [Google Scholar] [CrossRef]
- Diesburg, K. A new growth regulator for golf course turf-grass. Golf Course Manag. 1999, 67, 49–51. [Google Scholar]
- Chen, H.M.; Xue, L.; Chintamanani, S.; Germain, H.; Lin, H.Q.; Cui, H.T.; Cai, R.; Zuo, J.R.; Tang, X.Y.; Li, X.; et al. Ethylene insensitive3-like1 repress salicylic acid induction deficient2 expression to negatively regulate plant innate immunity in Arabidopsis. Plant Cell 2009, 21, 2527–2540. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.Y.; Zhang, J.F.; Zhang, H.W.; Zhang, Z.J.; Quan, R.D.; Zhou, S.R.; Huang, R.F. Transcriptional Activation of OsDERF1 in OsERF3 and OsAP2-39 Negatively Modulates Ethylene Synthesis and Drought Tolerance in Rice. PLoS ONE 2011, 6, e25216. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.H.; Liu, J.; He, X.J.; Mu, R.L.; Zhou, H.L.; Chen, S.Y.; Zhang, J.S. Modulation of ethylene responses affects plant salt-stress responses. Plant Physiol. 2007, 143, 707–719. [Google Scholar] [CrossRef] [PubMed]
- Eremina, M.; Rozhon, W.; Poppenberger, B. Hormonal control of cold stress responses in plants. Cell. Mol. Life Sci. 2016, 73, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Liu, W.; Xia, X.; Wang, T.; Zhang, W.H. Cold acclimation-induced freezing tolerance of Medicago truncatula seedlings is negatively regulated by ethylene. Physiol. Plant. 2014, 152, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Tian, S.; Hou, L.; Huang, X.; Zhang, X.; Guo, H.; Yang, S. Ethylene signaling negatively regulates freezing tolerance by repressing expression of CBF and type-A ARR genes in Arabidopsis. Plant Cell 2012, 24, 2578–2595. [Google Scholar] [CrossRef] [PubMed]
- Munshaw, G.C.; Ervin, E.H.; Beasley, J.S.; Shang, C.; Zhang, X.; Parrish, D.J. Effects of late-season ethephon applications on cold tolerance parameters of four bermudagrass cultivars. Crop Sci. 2010, 50, 1022–1029. [Google Scholar] [CrossRef]
- Kosová, K.; Prášil, I.T.; Vítámvás, P.; Dobrev, P.; Motyka, V.; Floková, K.; Novák, O.; Turečková, V.; Rolčik, J.; Pešek, B.; et al. Complex phytohormone responses during the cold acclimation of two wheat cultivars differing in cold tolerance, winter Samanta and spring Sandra. J. Plant Physiol. 2012, 169, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.J.; Huang, R.F. Enhanced tolerance to freezing in tobacco and tomato overexpressing transcription factor TERF2/LeERF2 is modulated by ethylene biosynthesis. Plant Mol. Biol. 2010, 73, 241–249. [Google Scholar] [CrossRef]
- Wei, D.; Li, Y.R.; Di, N.N.; Bi, Z.Q.; Bu, Z.Y. Effect of ethephon on cold resistance of young plants of Banana (Musa AAA Cavendish subgroup). Chin. J. Trop. Crop. 2009, 30, 1447–1451. (In Chinese) [Google Scholar]
- Yu, X.M.; Griffith, M.; Wiseman, S.B. Ethylene induces antifreeze activity in winter rye leaves. Plant Physiol. 2001, 126, 1232–1240. [Google Scholar] [CrossRef] [PubMed]
- Ciardi, J.A.; Deikman, J.; Orzolek, M.D. Increased ethylene synthesis enhances chilling tolerance in tomato. Physiol. Plant. 1997, 101, 333–340. [Google Scholar] [CrossRef]
- Guye, M.G.; Vigh, L.; Wilson, L.M. Chilling-induced ethylene production in relation to chill-sensitivity in Phaseolus spp. J. Exp. Bot. 1987, 38, 680–690. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, H.; Pan, X.; Chen, X.L.; Zhang, Z.J.; Lu, X.Y.; Huang, R.F. Overexpression of ethylene response factor TERF2 confers cold tolerance in rice seedlings. Transgenic Res. 2011, 20, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Jun, H.L.; Duk, J.Y.; Su, J.K.; Doil, C.; Hee, J.L. Intraspecies differences in cold hardiness, carbohydrate content and β-amylase gene expression of Vaccinium corymbosum during cold acclimation and deacclimation. Tree Physiol. 2012, 32, 1533–1540. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, L.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dhont, C. Nitrogen reserves, spring regrowth and winter survival of field-grown alfalfa (Medicago sativa) defoliated in the autumn. Ann. Bot. (Lond.) 2006, 97, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.J.; Pellett, N.E.; Klein, R.M. Phytochrome control of growth cessation and initiation of cold acclimation in selected woody plants. Plant Physiol. 1972, 50, 262–265. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Pallardy, S.G. Acclimation and adaptive responses of woody plants to environmental stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Mao, L.Z.; Lu, H.F.; Wang, Q.; Cai, M.M. Comparative photosynthesis characteristics of Calycanthus chinensis and Chimonanthus praecox. Photosynthetica 2007, 45, 601–605. [Google Scholar] [CrossRef]
- Jing, H.C.; Schippers, J.H.; Hille, J.M.; Dijkwel, P.P. Ethylene-induced leaf senescence depends on age-related changes and OLD genes in Arabidopsis. J. Exp. Bot. 2005, 56, 2915–2923. [Google Scholar] [CrossRef] [PubMed]
- Qiu, K.; Li, Z.P.; Yang, Z.; Chen, J.Y.; Wu, S.X.; Zhu, X.Y.; Gao, S.; Gao, J.; Ren, G.D.; Kuai, B.K.; et al. Ein3 and ore1 accelerate degreening during ethylene-mediated leaf senescence by directly activating chlorophyll catabolic genes in arabidopsis. PLoS Genet. 2015, 11, e1005399. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.Y.; Zhang, Y.L.; Shao, H.; Lu, J. Differential physio-biochemical responses to cold stress of cold-tolerant and non-tolerant grapes (Vitis L.) from China. J. Agron. Crop Sci. 2010, 196, 212–219. [Google Scholar] [CrossRef]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endodormancy, paradormancy, and ecodormancy physiological terminology and classification for dormancy research. Hortic. Sci. 1987, 22, 371–377. [Google Scholar]
- Zhang, Y.; Mechlin, T.; Dami, I. Foliar application of abscisic acid induces dormancy responses in greenhouse-grown grapevines. Hortic. Sci. 2011, 46, 1271–1277. [Google Scholar] [CrossRef]
- Durner, E.F.; Gianfagna, T.J. Fall ethephon application increases peach flower bud resistance to low-temperature stress. J. Am. Soc. Hortic. Sci. 1987, 113, 404–406. [Google Scholar]
- Durner, E.F.; Gianfagna, T.J. Ethephon prolongs dormancy and enhances supercooling in peach flower buds. J. Am. Soc. Hortic. Sci. 1991, 116, 500–506. [Google Scholar] [CrossRef]
- Gianfagna, T.J. The effect of LAB 173711 and ethephon on time of flowering and cold hardiness of peach flower buds. J. Plant Growth Regul. 1991, 10, 191–195. [Google Scholar] [CrossRef]
- Lim, C.C.; Arora, R. Comparing Gompertz and Richards functions to estimate freezing injury in Rhododendron using electrolyte leakage. J. Am. Soc. Hortic. Sci. 1998, 123, 246–252. [Google Scholar] [CrossRef]
- Wang, W.J.; Wang, Z.P.; Ping, J.C.; Zhang, Y.H. Effect of Ethephon on Several Cold Hardiness Indexes of Carbernet sauvignon. Sino-Overseas Grapevine Wine 2005, 5, 13–14. [Google Scholar]
- Guy, C.L. Freezing tolerance of plants: Current understanding and selected emerging concepts. Can. J. Bot. 2003, 81, 1216–1223. [Google Scholar] [CrossRef]
- Keller, M. The Science of Grapevines: Anatomy and Physiology, 1st ed.; Academic Press: Burlington, MA, USA, 2010. [Google Scholar]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, G.C.; Murata, N. The unusually strong stabilising effects of glycine betaine on the structure and function of the oxygen-evolving photosystem II complex. Photosynth. Res. 1995, 44, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Matysik, J.; Alia, A.; Bhalu, B.; Mohanty, P. Molecular mechanisms of quenching of reactive oxygen species by proline understress in plants. Curr. Sci. 2002, 82, 525–532. [Google Scholar]
- Crisosto, C.H.; Lombard, P.B.; Fuchigami, L.H. Effect of ethephon and hand defoliation on dormant bud ethylene levels, bloom, delay and yield components of “Red heaven” peach. Acta Hortic. 1987, 10, 201–211. [Google Scholar]
Figure 1.
A Magnolia wufengensis flower in Wufeng County, Hubei Province, P.R. China.

Figure 2.
Dynamics of air temperature at the experimental site (39°48′ N, 116°28′ E) from September 2017 to March 2018.
Figure 2.
Dynamics of air temperature at the experimental site (39°48′ N, 116°28′ E) from September 2017 to March 2018.

Figure 3.
Relative growth rate (RGR) in the shoots of Magnolia wufengensis under ethephon (ETH) treatments is shown in the bar chart. Days to growth cessation in the shoots of Magnolia wufengensis under ETH treatments is shown in the line graph. T0, T1, T2, and T3 indicate the ETH application of 0, 700, 1000, and 1500 mg·L−1, respectively. Different letters indicate significant differences (p < 0.05) among treatments. Data are presented as the means ± SDs (n = 10). The same is shown below.
Figure 3.
Relative growth rate (RGR) in the shoots of Magnolia wufengensis under ethephon (ETH) treatments is shown in the bar chart. Days to growth cessation in the shoots of Magnolia wufengensis under ETH treatments is shown in the line graph. T0, T1, T2, and T3 indicate the ETH application of 0, 700, 1000, and 1500 mg·L−1, respectively. Different letters indicate significant differences (p < 0.05) among treatments. Data are presented as the means ± SDs (n = 10). The same is shown below.

Figure 4.
Dynamics of leaf chlorophyll (ug·cm−2) in Magnolia wufengensis under ethephon (ETH) treatments.
Figure 4.
Dynamics of leaf chlorophyll (ug·cm−2) in Magnolia wufengensis under ethephon (ETH) treatments.

Figure 5.
Dynamics of the bud dormancy level in Magnolia wufengensis under ethephon (ETH) treatments.
Figure 5.
Dynamics of the bud dormancy level in Magnolia wufengensis under ethephon (ETH) treatments.

Figure 6.
Dynamics of the freezing tolerance of shoots in Magnolia wufengensis under ethephon (ETH) treatments.
Figure 6.
Dynamics of the freezing tolerance of shoots in Magnolia wufengensis under ethephon (ETH) treatments.

Figure 7.
Dynamics of the water content of shoots in Magnolia wufengensis under ethephon (ETH) treatments.
Figure 7.
Dynamics of the water content of shoots in Magnolia wufengensis under ethephon (ETH) treatments.

Figure 8.
Dynamics of the proline content of shoots in Magnolia wufengensis under ethephon (ETH) treatments.
Figure 8.
Dynamics of the proline content of shoots in Magnolia wufengensis under ethephon (ETH) treatments.

Figure 9.
Bud survival rate (%) of Magnolia wufengensis under ethephon (ETH) treatments is shown in the bar chart. Days to terminal bud-burst of Magnolia wufengensis under ETH treatments is shown in the line graph.
Figure 9.
Bud survival rate (%) of Magnolia wufengensis under ethephon (ETH) treatments is shown in the bar chart. Days to terminal bud-burst of Magnolia wufengensis under ETH treatments is shown in the line graph.

Figure 10.
Correlation coefficients between the freezing tolerance (estimated as LT50) and water content in the shoots of Magnolia wufengensis under ETH treatments. Note: T0, T1, T2, and T3 indicate the application of 0, 700, 1000, and 1500 mg·L−1 ETH, respectively. The regression line is T2, T1, T3, and T0 from the top to bottom. ** indicate significance at p < 0.01, respectively.
Figure 10.
Correlation coefficients between the freezing tolerance (estimated as LT50) and water content in the shoots of Magnolia wufengensis under ETH treatments. Note: T0, T1, T2, and T3 indicate the application of 0, 700, 1000, and 1500 mg·L−1 ETH, respectively. The regression line is T2, T1, T3, and T0 from the top to bottom. ** indicate significance at p < 0.01, respectively.

Figure 11.
Correlation coefficients between the freezing tolerance (estimated as LT50) and proline content in the shoots of Magnolia wufengensis under ETH treatments. Note: T0, T1, T2, and T3 indicate the application of 0, 700, 1000, and 1500 mg·L−1 ETH, respectively. The regression line is T2, T1, T3, and T0 from the top to bottom. * indicate significance at p < 0.05, respectively.
Figure 11.
Correlation coefficients between the freezing tolerance (estimated as LT50) and proline content in the shoots of Magnolia wufengensis under ETH treatments. Note: T0, T1, T2, and T3 indicate the application of 0, 700, 1000, and 1500 mg·L−1 ETH, respectively. The regression line is T2, T1, T3, and T0 from the top to bottom. * indicate significance at p < 0.05, respectively.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summaries of previous studies on the effect of ethephon or ethylene on the chilling/freezing tolerance of plants.
Table 1.
Summaries of previous studies on the effect of ethephon or ethylene on the chilling/freezing tolerance of plants.
| Role in Freezing Tolerance | Species | By which Bio Synthetic Pathways or Molecular Mechanism |
|---|---|---|
| Positive | Wheat [29] | Ethylene precursor amino cyclopropane carboxylic acid was quickly upregulated |
| Tobacco [30] | TERF2/LeERF2 is involved in enhancing freezing tolerance | |
| Secale cereale (rye) [32] | Ethylene induces antifreeze activity | |
| Tomato [33] | maintaining chlorophyll a fluorescence levels (Fv/Fm) | |
| Phaseolus spp. (bean) [34] | The more chill-tolerant cultivars produced greater amounts of chilling-induced ethylene | |
| Rice [35] | Overexpression of ethylene response factor TERF2 confers cold tolerance in rice | |
| Medicago truncatula [26] | Ethylene level was negatively correlated with freezing tolerance as inhibition of ethylene biosynthesis by inhibitors of ethylene biosynthesis enhanced freezing tolerance | |
| Negative | Arabidopsis [27] | Repressing expression of CBF and Type-A ARR genes |
| Bermudagrass [28] | negatively affected proline concentrations of some cultivars |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Duan, X.; Cai, C.; Yang, Y.; Chen, F.; Sang, Z.; Ma, L. Fall Ethephon Application Enhances the Freezing Tolerance of Magnolia wufengensis During Overwintering. Forests 2019, 10, 868. https://doi.org/10.3390/f10100868
AMA Style
Duan X, Cai C, Yang Y, Chen F, Sang Z, Ma L. Fall Ethephon Application Enhances the Freezing Tolerance of Magnolia wufengensis During Overwintering. Forests. 2019; 10(10):868. https://doi.org/10.3390/f10100868
Chicago/Turabian StyleDuan, Xiaojing, Chuheng Cai, Yang Yang, Faju Chen, Ziyang Sang, and Luyi Ma. 2019. "Fall Ethephon Application Enhances the Freezing Tolerance of Magnolia wufengensis During Overwintering" Forests 10, no. 10: 868. https://doi.org/10.3390/f10100868
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.