1. Introduction
Tea (
Camellia sinensis (L.) O. Kuntze, 2
n = 2
x = 30) is one of the most popular non-alcoholic beverages worldwide, and is consumed by approximately 70% of the world’s population for its refreshing taste, attractive aroma, therapeutic uses, and mildly stimulating properties [
1]. It is an economically important tree crop, grown in over 52 countries in Asia, Africa, and South America [
2,
3]. The tea is a woody ever-green perennial plant and recorded to be native to Yunnan and Sichuan provinces in China and the northern part of Myanmar [
4]. In Korea, although tea was introduced from China as early as the seventh century, the development of the tea industry was slow, and production was small [
5].
The importance of using genetic resources in breeding programs to enhance crop genetic potential has been well recognized [
6]. Many germplasm appraisal methods, such as morphology, biochemistry, molecular markers, and sensory evaluation, have been used to evaluate the resources of tea germplasm [
7,
8,
9]. The phenotype can be referred to as a good standard for the evaluation of tea germplasm, because this method is simply based on the morphological traits to analyze the genetic diversity assessment [
10]. Recently, the technology of using molecular markers has been proven to be one of the most effective methods for identifying different tea varieties [
2,
7,
11,
12,
13,
14].
Tea is an out-crossing species, and selected elite genotypes are propagated vegetatively and released as clonal varieties [
13,
15,
16]. Clonal identification is traditionally based on morphological descriptors such as plant shape, stem width, leaf shape, young leaf type, and fruit shape [
15,
17]. However, as in many out-crossing crops, tea is highly heterozygous with most of its morphological, physiological, and biochemical descriptors showing continuous variation and high plasticity [
18,
19]. Korir et al. reported that morphological traits are associated with drawbacks such as the influences of environment on trait expressions, epistatic interactions, and pleiotropic effects among others despite the value of their advantages [
17]. On the contrary, molecular markers are used as they are least affected by environmental factors and indefinite presence. In addition, they offer a possibility to observe the genome directly and thus eliminate the shortcomings inherent in a phenotype observation [
15]. In previous studies, genetic diversity, discrimination and differentiation of tea germplasms have been assessed using different DNA markers such as restriction fragment length polymorphism (RFLP), randomly amplified polymorphic DNA (RAPD), amplified fragment length polymorphism (AFLP), inter-simple sequence repeat (ISSR), and simple sequence repeat (SSR) [
3,
5,
7,
11,
12,
13,
14,
15,
20,
21,
22].
In Korea, the national research institutes collected and conserved excellent tea individuals and investigated their morphological characteristics [
23]. Also, some studies analyzed the genetic diversity of Korean tea germplasm using RFLP and RAPD [
5,
14,
16]. However, the analysis of genetic diversity in Korean tea germplasm is not sufficient as it included a very small population (approximately 20–50 individuals). In the present study, 21 SSR primer pairs were used to analyze 410 tea accessions from Korea, and the aim was: (1) to evaluate the genetic diversity and population structure of Korean tea accessions and (2) to estimate the genetic differentiation and variation source among inferred populations. It is hypothesized that the results of the present study would be helpful to gain a deeper understanding on the genetic diversity, population structure, and differentiation of tea germplasm to guide effective collection, conservation, and application of tea genetic resources in Korea.
4. Discussion
Erosion of plant genetic diversity is a very serious problem caused by modernization and replacement of wild plants or landraces with a few elite varieties [
31,
32]. Therefore, collection and preservation of plant genetic resources are of immense importance for crop breeding to support the demands of a growing human population. Effective management and utilization of plant genetic resources require information about the origin of strains, phenotypic traits, and genetic diversity (identified by molecular techniques) [
33]. In this study, analysis of genetic diversity of 410 tea accessions collected and conserved in the Korea genebank was performed. Genetic diversity provides an assurance of future genetic progress and insurance against unforeseen threats to agricultural production such as disease epidemics or climate changes. Thus, the fate of genetic diversity in these gene pools is of utmost importance if plant breeding will continue to address the pressing needs of society such as increased yield, genetic resistance to diseases and pests, improved nutritional and processing quality of crop products, and reduction in environmental effects [
34].
In the present study, about 85% of tea accessions were collected from Boseong and Suncheon in Jeollanam-do (JN) (
Table 2). According to Eom and Kim, the tea seeds obtained from China were firstly cultivated in Mount Jiri in JN and so a majority of tea plants are included in the Honam region (Jeollanam-do and Jeollabuk-do) [
35]. In addition, tea experiment stations in Boseong experiment station (BES) and Mokpo experiment station (MES, a city close to Suncheon) have collected the tea accessions since the late 1990s [
18]. The two experiment stations have probably collected tea accessions around the area where the institute is located, and thus the largest number of tea accessions was collected in the region. The tea accessions of the two research institutes have been managed as registered tea germplasms of the NAC and appear to cause a regional collectivity imbalance of tea accessions in Korea.
In this study, the mean Nei’s gene diversity (GD, 0.792) across 21 SSR markers was higher than other studies; 0.652 in 280 tea accessions using 23 SSR markers [
7], 0.640 in 450 tea accessions using 96 EST-SSR marker [
22], 0.543 in 185 Chinese tea cultivars using 48 SSR markers [
13], and 0.680 in 64 Sri Lankan tea cultivars using 33 EST- or genomic-SSR markers [
3]. The gene diversity of a locus, also known expected heterozygosity, is a fundamental measure of genetic variation in a population, and describes the proportion of heterozygosis expected under Hardy–Weinberg equilibrium [
36]. As tea is an open pollinated plant, the tea plant shows highly heterogeneous and consequently broad genetic variation [
13]. The obtained results also showed high gene diversity in a manner similar to the previously reported data. However, Yao et al., mentioned that the comparison of the degree of genetic diversity between different studies is difficult as the analysis may be affected by various factors like sampling schemes, number of SSR markers, sizes of SSR repeats, and location of SSR in the genome [
22].
Contrary to the result of higher gene diversity, 410 tea accessions in this study were characterized by an extreme dearth of genetic diversity as revealed by an overall Simpson’s Dominance (
λ) of 0.998. Furthermore, the AMOVA revealed there was no significant difference among populations, suggesting low genetic diversity across the entire collected region. In addition, the standardized index of association (rbarD, 0.0583, (
p < 0.001)) supported the hypothesis of clonal population structure based on the linkage disequilibrium tests, where the null hypothesis of random mating was rejected for all populations. Under clonal propagation, heterozygosis and allelic diversity at each locus are expected to increase [
37,
38]. While high levels of clonality tend to increase genetic variation within the population, an opposite effect is expected on genetic differentiation among populations and on genotypic diversity, both decreasing with the rate of clonal reproduction [
37,
39]. Indeed, the 410 tea accessions in this study were landraces and are likely to have been collected from private farms. As breeding a reliable cultivar for a private farm is nearly impossible, almost all the tea gardens consist of seedling tea plants from the local and wild origin with great morphological variations [
23]. BES and MES also collected and investigated the morphological characteristics of tea germplasm and many variations were observed in the number of stems, stem length, leaf area, and leaf color which were within the limits of the investigation of the morphological characteristics [
40]. Due to the lack of sufficient studies on the genetic diversity of Korean tea germplasm, a few researchers argued over the importance of genetic collection and preservation of tea accessions [
14,
21].
Previous studies performed the analysis of genetic diversity of different tea accessions using molecular markers like RFLP, RAPD, and SSR [
3,
5,
21,
22]. In addition, the STRUCTURE software was used to analyze the population structure of tea germplasm [
2,
3,
13,
22]. To analyze the genetic diversity and population structure of Korean tea accessions, 21 SSR markers and DAPC analysis were used in this study. The DAPC method provides an interesting alternative to STRUCTURE software as it does not require that populations should be in Hard-Weinberg equilibrium and can handle large sets of data without using parallel processing software [
41]. DAPC analysis divided the population into well-defined clusters associated with provenance, ploidy, taxonomy and breeding program of the genotypes and related to their genetic structure [
42]. According to Rosyara et al., STRUCTURE, EIGENSTRAT, and DAPC exhibit the ability to control population structure in association with mapping studies [
43]. EIGENSTRAT and DAPC were slightly better than STRUCTURE but DAPC led to a better separation among populations. In this study, DAPC (four clusters) analysis provided a more detailed clustering within tea accessions than STRUCTURE (two populations) (
Figure S1). Campoy et al., reported that their results of population structure in sweet cherry using STRUCTURE and DAPC showed good consistency between the two methods and DAPC analysis provided a more detailed clustering among the populations compared to STRUCTURE analysis [
41].
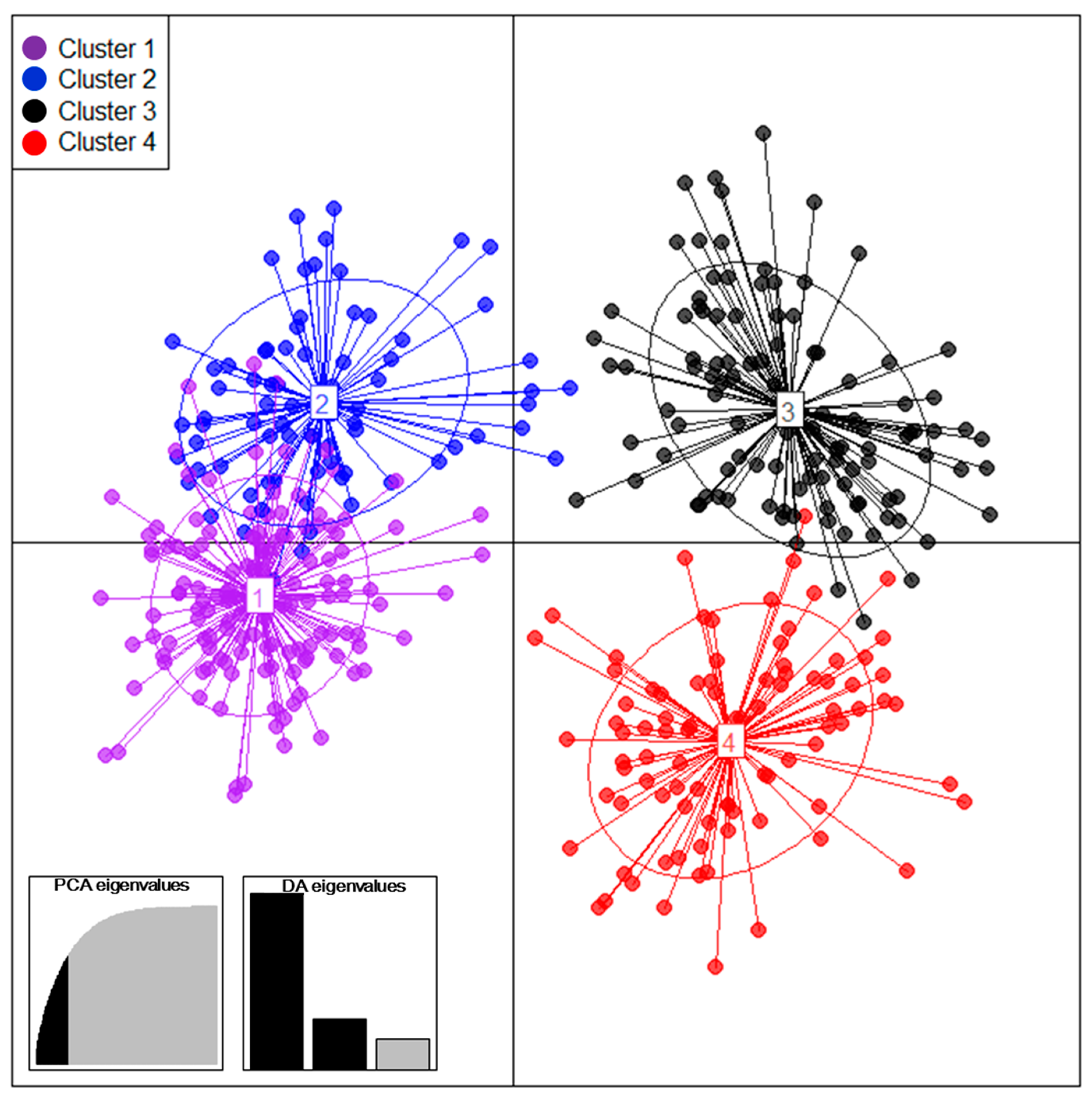
As per the result of DAPC, 410 tea accessions were divided into four clusters (
Figure 1). Among them, cluster 1 and 2 showed a higher frequency of rare alleles and genetic diversity, and there was high gene flow (Nm = 45.120) between two clusters. Yao et al., reported that a majority of rare SSR alleles and higher diversity were observed in the tea accessions from Yunnan and its neighboring provinces, considered as an original center of the tea plant in China [
22]. They also reported that the allele number, genetic diversity, and PIC value of tea germplasm significantly decreased with the distance away from the origin center of the tea plant. Although a particular collection area cannot be designated as an origin, tea accessions contained in cluster 1 and 2 are thought to be the origin of the Korean tea germplasm due to their ratio of rare alleles and higher genetic diversity.
Kaundun et al., reported that tea accessions collected from Korea showed higher genetic diversity than those from Taiwan and Japan [
21]. On the other hand, Jeong and Park mentioned that the genetic variation in Korean tea population is smaller compared to Chinese or Japanese wild tea populations [
18]. The results of the present study confirmed that 410 tea accessions collected and conserved in Korea genebank exhibit the narrow genetic variations. Park et al., suggested that the low genetic diversity of Korean tea was established from a limited gene stock from China [
14]. The short history and relatedly homogeneous environment in which they were introduced in the southwestern part of the country did not favor population differentiation. In addition, loss of diversity was exacerbated by the mass destruction of tea plantations in the fourteenth century due to political and religious reasons [
44]. Consequently, tea being a highly outcrossing species, variability is mostly expected within rather than between the populations as predicted by Hamrick [
45].

 ,
, 


{kind=link}