
Formica gagatoides Ruzsky, 1904, and Siberian F. kozlovi Dlussky, 1965 (Hymenoptera: Formicidae); Two or One Species?
 ,
,
Abstract
:1. Introduction

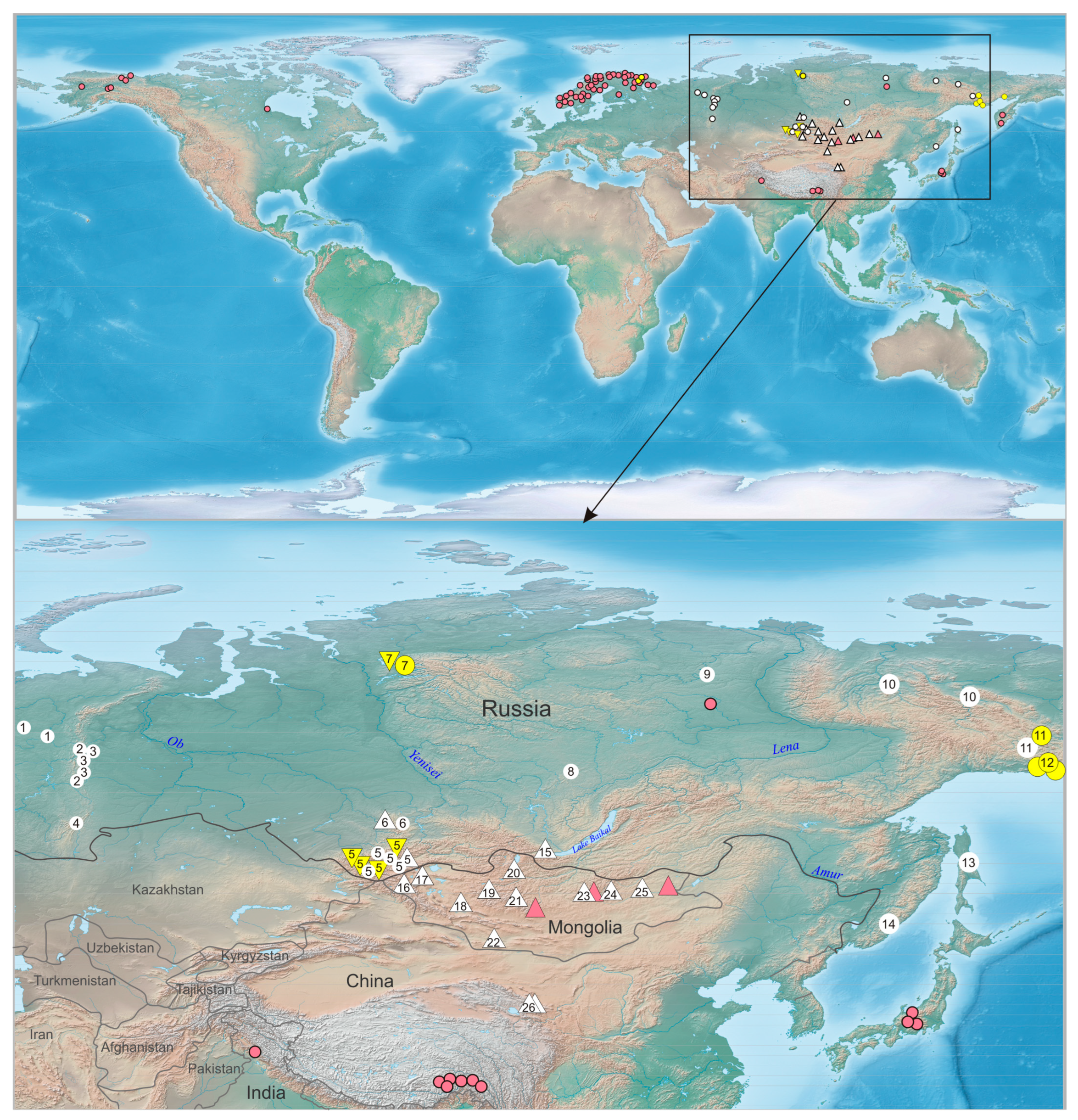
2. Materials and Methods
2.1. Morphological Features
2.2. Molecular Genetics Study
2.3. Phylogenetic Analysis
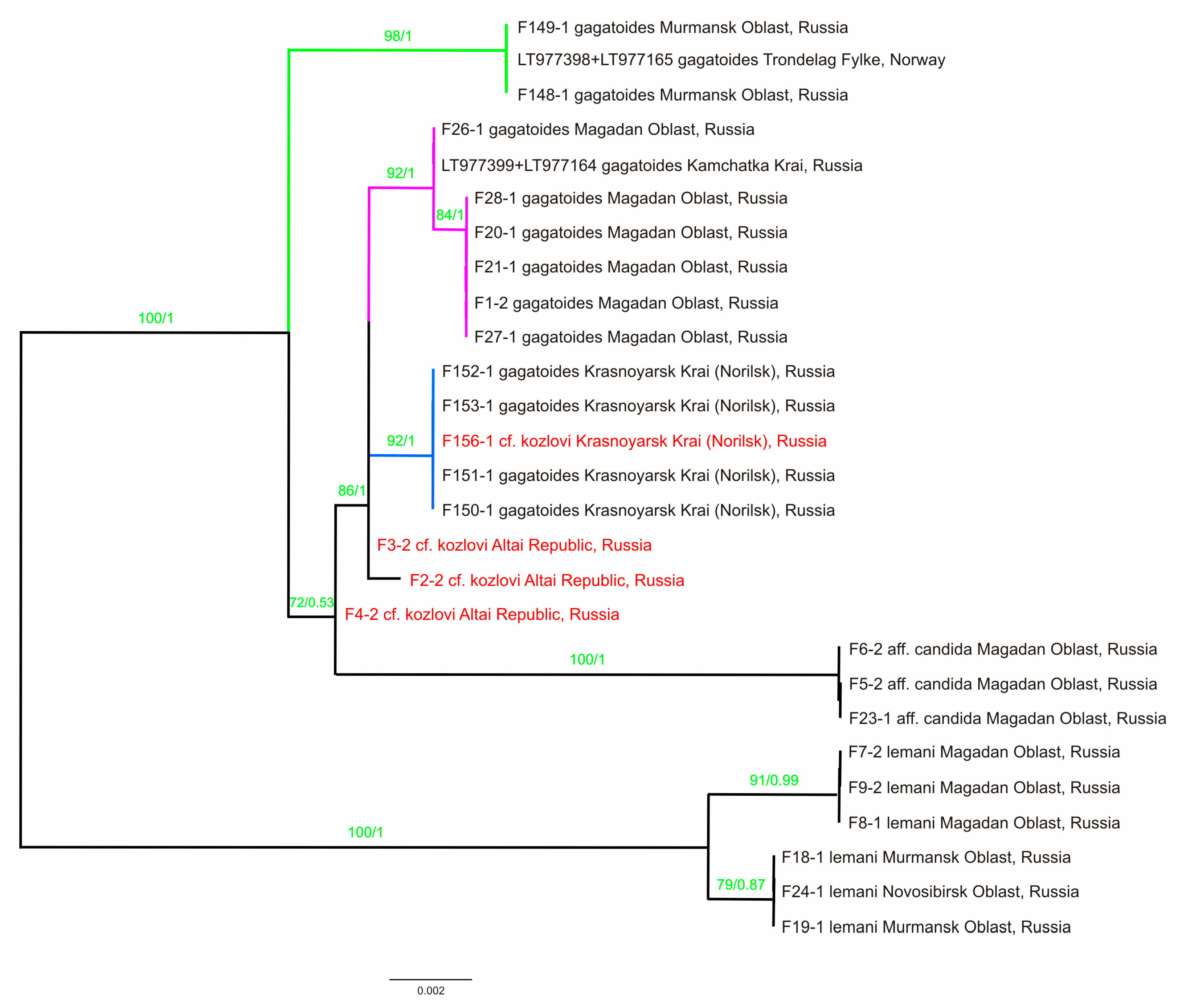
3. Results
3.1. Degree of Morphological Differences between F. cf. kozlovi and F. gagatoides
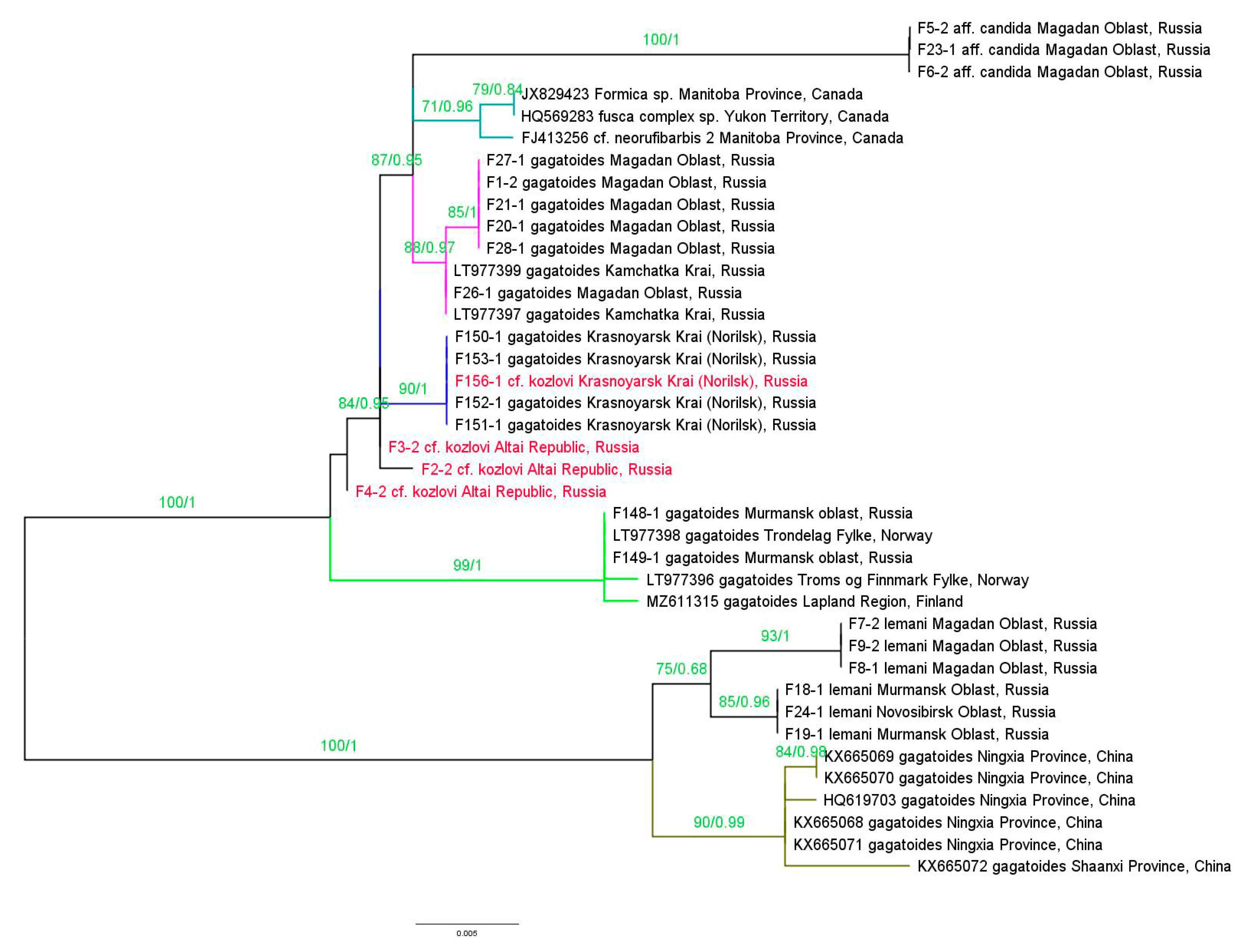
3.2. Molecular Genetic Assay
Diversity of the COI Gene
4. Discussion
5. Conclusions
- For species with a wide range (such as F. gagatoides), it is inappropriate to use their chaetotaxy type as a diagnostic feature, due to its strong variability. In particular, this concerns the number of setae on the mid-leg femora of workers and females, as well as on the pronotum of females.
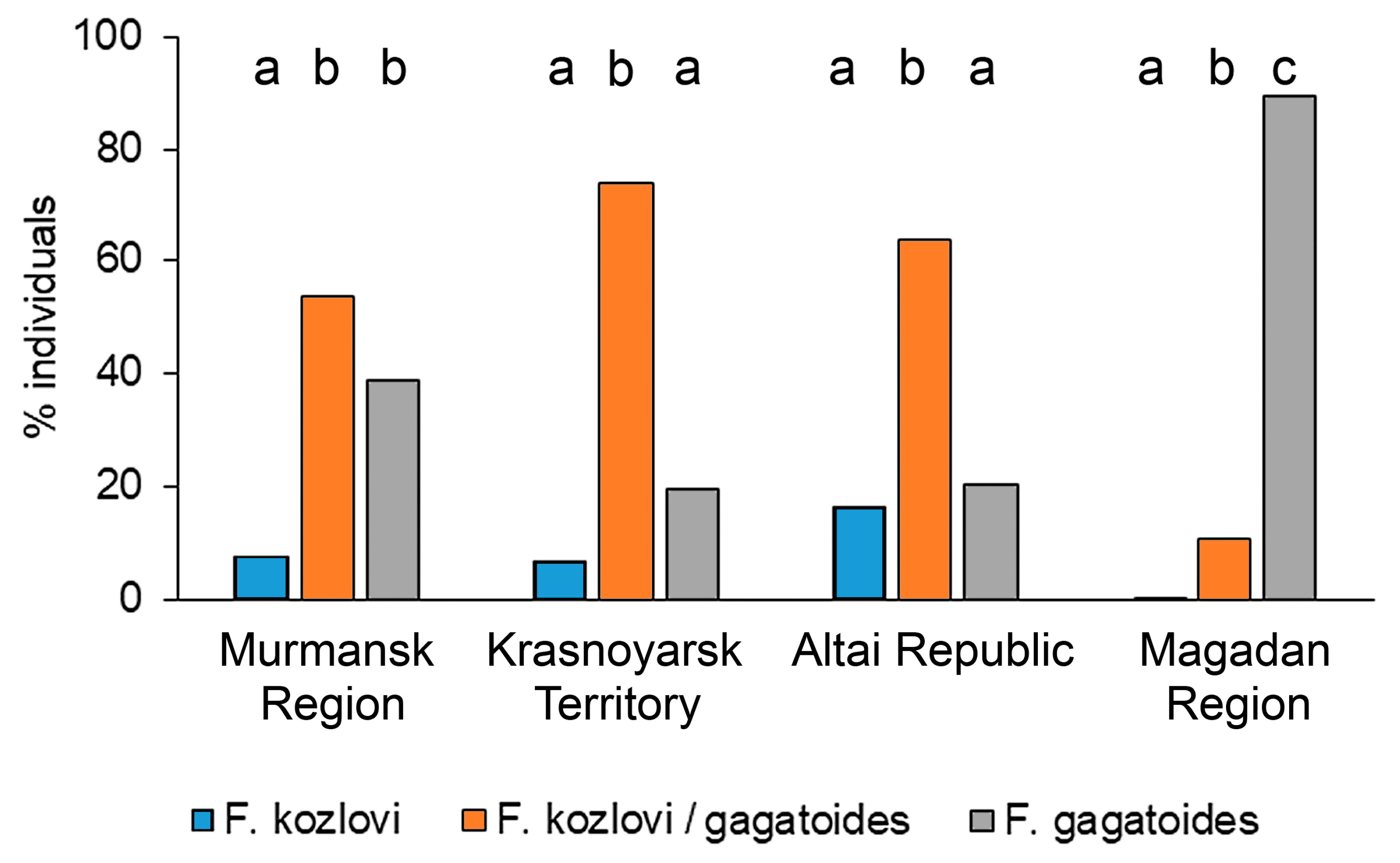
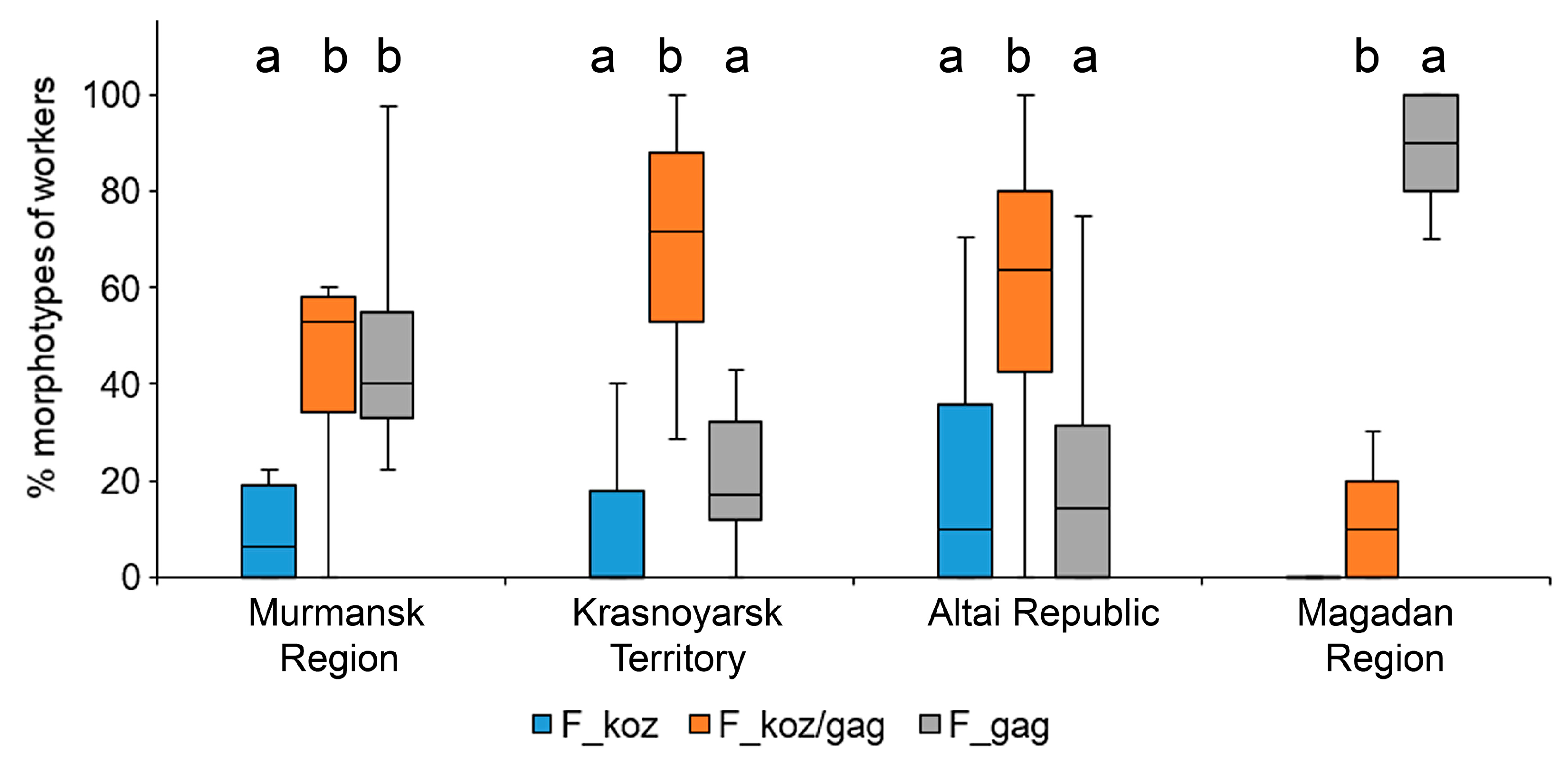
- In representatives of the F. gagatoides/F. cf. kozlovi complex in Siberia, a high diversity of variants was noted, according to the main morphological diagnostic characters for the workers and females—the number of setae on the mid-leg femora and pronotum, as well as the pubescence of the first gaster tergites. In Siberia, the diversity of the morphotypes among the workers was the highest (Krasnoyarsk Territory—eight, Altai—seven), with a predominance of intermediate forms (Krasnoyarsk Territory—73.6%, Altai—63.4%). In females, five morphotypes were noted, with a predominance of “mixed” forms (about 60%). As for “pure” morphotypes in females, rather high proportions of F. kozlovi (30.8%) and F. gagatoides (28.6%) were noted in Altai and the Krasnoyarsk Territory, respectively.
- The results of the molecular genetic analysis of the representatives of the Siberian F. cf. kozlovi and Asian F. gagatoides, involving specimens with both “pure” and “mixed” morphotypes, revealed the conspecifity of the studied specimens and significantly expanded the knowledge on the distribution of F. gagatoides.
- The data obtained raise the issue of the F. kozlovi species status, which requires an additional investigation into F. kozlovi materials from the areas of the description of its type specimens (Mongolia and China). In addition, the data from the molecular analysis also raise the issue of the concpecifity of the Asian and European branches of F. gagatoides.
- Generally, the data obtained significantly expand the knowledge of the distribution, morphological variability, and ecotypes of F. gagatoides.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhigulskaya, Z.A. Ants (Hymenoptera, Formicidae) of the Kurai Basin of Southeastern Altai. Tomsk State Univ. J. 2011, 350, 189–195. [Google Scholar]
- Antonov, I.A.; Pleshanov, A.S. Ecological-geographical features of myrmecofauna of Baikal region. Bull. Buryat St. Univ. Biol. Geograph. 2011, 4, 104–108. [Google Scholar]
- Bayartogtokh, B.; Aibek, U.; Pfeiffer, M.; Yamane, S. Diversity and biogeography of ants in Mongolia (Hymenoptera: Formicidae). Asian Myrmecol. 2014, 6, 63–82. [Google Scholar]
- Cronin, A.L.; Donnerhack, O.; Seidel, F.; Yamanaka, T. Fine-scale variation in natural nitrogen isotope ratios of ants (Hymenoptera: Formicidae). Entomol. Exp. Appl. 2015, 157, 354–359. [Google Scholar] [CrossRef]
- Dlussky, G.M. Ants of the genus Formica of Mongolia and Northeast Tibet. Ann. Zool. 1965, 23, 15–43. [Google Scholar]
- Dlussky, G.M. Ants of the Genus Formica (Hymenoptera: Formicidae g. Formica); Nauka: Moscow, Russia, 1967; p. 236. [Google Scholar]
- Sonobe, R.; Dlussky, G.M. On two ant species of the genus Formica (Hymenoptera, Formicidae) from Japan. Kontyû 1977, 45, 23–25. [Google Scholar]
- Dmitrienko, V.K.; Petrenko, E.S. Ants of Taiga Biocenoses of Siberia; Nauka: Novosibirsk, Russia, 1976; p. 220. [Google Scholar]
- Kupyanskaya, A.N.; Shabalin, S.A. Ants (Hymenoptera, Formicidae) of conifer-broad-leaved mixed forests of South Sikhote-Alin. Euroasian Ent. J. 2012, 11, 249–260. [Google Scholar]
- Berman, D.I.; Alfimov, A.V.; Zhigulskaya, Z.A.; Leirikh, A.N. Overwintering and Cold-Hardiness of Ants in the Northeast of Asia; Pensoft Publishers: Sofia/Moscow, Russia, 2010; p. 294. [Google Scholar]
- Francoeur, A. Ants (Hymenoptera: Formicidae) of the Yukon. In Insects of the Yukon; Danks, H.V., Downes, J.A., Eds.; Biological Survey of Canada (Terrestrial Arthropods): Ottawa, ON, Canada, 1997; pp. 901–910. [Google Scholar]
- Antmaps. Available online: https://antmaps.org/?mode=species&species=Formica.gagatoides (accessed on 11 March 2023).
- Schär, S.; Talavera, G.; Espadaler, X.; Rana, J.D.; Andersen, A.A.; Cover, S.P.; Vila, R. Do Holarctic ant species exist? Trans-Beringian dispersal and homoplasy in the Formicidae. J. Biogeogr. 2018, 45, 1917–1928. [Google Scholar] [CrossRef]
- Xu, Z.; Chu, J.; Zhang, C.; Yu, N. Ant Species and Distribution Pattern in Gongbo Nature Reserve in Southeastern Tibet. Sichuan J. Zool. 2011, 30, 118–123. [Google Scholar]
- Liu, C.; Dudley, K.L.; Xu, Z.; Economo, E.P. Mountain metacommunities: Climate and spatial connectivity shape ant diversity in a complex landscape. Ecography 2018, 41, 101–112. [Google Scholar] [CrossRef]
- Bharti, H.; Sharma, Y.P.; Bharti, M.; Pfeiffer, M. Ant species richness, endemicity and functional groups, along an elevational gradient in the Himalayas. Asian Myrmecol. 2013, 5, 79–101. [Google Scholar]
- Dlussky, G.M.; Zryanin, V.A. Keys for identifying species of the genus Formica. In Monitoring of the Formica Ants; Zakharov, A.A., Ed.; KMK Scientific Press: Moscow, Russia, 2013; pp. 52–60. [Google Scholar]
- Seifert, B. The “Black Bog Ant” Formica picea Nylander, 1846—A species different from Formica candida Smith, 1878 (Hymenoptera: Formicidae). Myrmecol. Nachr. 2004, 6, 29–38. [Google Scholar]
- Gilev, A.V. Polar ant Formica gagatoides Ruzsky, 1904. In Red Book of the Sverdlovsk Region: Animals, Plants, Mushrooms; Korytin, N.S., Ed.; OOO Mir: Ekaterinburg, Russia, 2018; p. 155. [Google Scholar]
- Blinova, S.V. The structure of myrmecocomplexes of the Kuznetsk Alatau. Bull. Higher Educat. Institut. N. Caucas. Reg. Natur. Sci. 2009, 4, 82–85. [Google Scholar]
- Kupyanskaya, A.N. Ants (Hymenoptera, Formicidae) of Sakhalin and Moneron Islands. In Flora and fauna of Sakhalin Island (Materials of the Sakhalin Island International Project), Part 2; Storozhenko, S.Y., Ed.; Dalnauka: Vladivostok, Russia, 2005; pp. 168–188. [Google Scholar]
- Zhigulskaya, Z.A.; Shekhovtsov, S.V.; Poluboyarova, T.V.; Berman, D.I. Formica picea and F. candida (Hymenoptera: Formicidae): Synonyms or Two Species? Diversity 2022, 14, 613. [Google Scholar] [CrossRef]
- Vaulin, O.V.; Karagodin, D.A.; Zakharov, I.K.; Baricheva, E.M. Dynamics of Malaria Mosquito Species Composition in Siberian Populations Detected by Restriction Analysis. Russ. J. Genet. 2018, 54, 838–847. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, S. Phylogenetic Relationships Based on DNA Barcoding Among 16 Species of the Ant Genus Formica (Hymenoptera: Formicidae) from China. J. Insect Sci. 2017, 17, 117. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Ji, Y.; Zhang, D.; He, L. Evolutionary conservation and versatility of a new set of primers for amplifying the ribosomal internal transcribed spacer regions in insects and other invertebrates. Mol. Ecol. Notes 2003, 3, 581–585. [Google Scholar] [CrossRef]
- Vaulin, O.V.; Novikov, Y.M. Phylogenetic relationships between Palaearctic species of the Anopheles maculipennis complex (Diptera: Culicidae) revealed by different approaches and markers. The problem of consensus. Vavilov J. Genet. Breed. 2016, 20, 695–703. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Hebert, P.D.; Ratnasingham, S.; de Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B 2003, 270, S96–S99. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [PubMed]
- Borowiec, M.L.; Cover, S.P.; Rabeling, C. The evolution of social parasitism in Formica ants revealed by a global phylogeny. Proc. Natl. Acad. Sci. USA 2021, 118, e2026029118. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Nei, M.; Rooney, A.P. Concerted and birth-and-death evolution of multigene families. Annu. Rev. Genet. 2005, 39, 121–152. [Google Scholar] [CrossRef]
- Schmidt, S.; Driver, F.; De Barro, P. The phylogenetic characteristics of three different 28S rRNA gene regions in Encarsia (Insecta, Hymenoptera, Aphelinidae). Org. Divers. Evol. 2006, 6, 127–139. [Google Scholar] [CrossRef]
- Sonnenberg, R.; Nolte, A.W.; Tautz, D. An evaluation of LSU rDNA D1–D2 sequences for their use in species identification. Front. Zool. 2007, 4, 6. [Google Scholar] [CrossRef]
- Fournier, D.; Aron, S. Hybridization and invasiveness in social insects—The good, the bad and the hybrid. Curr. Opin. Insect Sci. 2021, 46, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Stahlhut, J.K.; Fernandez-Triana, J.; Adamowicz, S.J.; Buck, M.; Goulet, H.; Hebert, P.D.N.; Huber, J.T.; Merilo, M.T.; Sheffield, C.S.; Woodcock, T.; et al. DNA barcoding reveals diversity of Hymenoptera and the dominance of parasitoids in a sub-arctic environment. BMC Ecol. 2013, 13, 2. [Google Scholar] [CrossRef]
- Smith, M.A.; Fernandez-Triana, J.; Roughley, R.; Hebert, P.D. DNA barcode accumulation curves for understudied taxa and areas. Mol. Ecol. Resour. 2009, 9, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Roslin, T.; Somervuo, P.; Pentinsaari, M.; Hebert, P.D.N.; Agda, J.; Ahlroth, P.; Anttonen, P.; Aspi, J.; Blagoev, G.; Blanco, S.; et al. A molecular-based identification resource for the arthropods of Finland. Mol. Ecol. Res. 2022, 22, 803–822. [Google Scholar] [CrossRef]
- Chesnokova, S.V.; Omelchenko, L.V. The Spatial Typological Organization of the Ant Population of the North-Eastern Altay. Sib. J. Ecol. 2004, 11, 481–492. [Google Scholar]
- Chesnokova, S.V.; Omelchenko, L.V. Ants of Central Altai: Spatial-typological structure and classification of communities. Entomol. Rev. 2011, 91, 253–263. [Google Scholar] [CrossRef]
- Pfeiffer, M.; Schultz, R.; Radchenko, A.; Yamane, S.; Woyciechowski, M.; Aibek, U.; Seifert, B. A critical checklist of the ants of Mongolia (Hymenoptera: Formicidae). Bonn. Zool. Beitr. 2007, 55, 1–8. [Google Scholar]
- Aibek, U.; Yamane, S. Comparison of ant communities (Hymenoptera: Formicidae) in different habitat types in North-Central Mongolia with special reference to their response to livestock grazing. Cauc. Entomol. Bull. 2015, 11, 111–120. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amplified Region | Forward Primer | Reverse Primer | Annealing Temperature | Reference |
|---|---|---|---|---|
| COI | GGTCA-ACAAA-TCATA- AAGAT-ATTGG | TAAAC-TTCAG-GGTGA-CCAAA-AAATC-A | 53 °C | [25] |
| ITS1 | TCAC-ACCGC-CCGTC- GCTAC-TA | ATGTG-CGTTC-RAAAT-GTCGA-TGTTC-A | 58 °C | [26] |
| D2 28S | 5’-GGAGT-CGTGT-TGCTT- GATAG-TGCAG-3’ | 5’-TTCTT-GGTCC-GTGTT-TCAAG-ACGGG-3’ | 60 °C | [27] |
| Region | Morphotypes of Workers, % | Number of Individuals | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1-1 | 1-2 | 1-3 | 1-4 | 2-1 | 2-2 | 2-3 | 2-4 | ||
| Murmansk Region | 2.08 | 42.71 | 23.96 | 2.08 | 5.21 | 14.58 | 9.38 | 0.00 | 96 |
| Krasnoyarsk Territory (Norilsk) | 3.45 | 16.09 | 11.49 | 1.15 | 17.24 | 43.68 | 4.60 | 2.30 | 87 |
| Altai Republic | 0.00 | 20.29 | 10.87 | 1.81 | 0.36 | 50.36 | 13.04 | 3.26 | 276 |
| Magadan Region | 54.96 | 34.35 | 2.29 | 0.00 | 6.11 | 2.29 | 0.00 | 0.00 | 131 |
| Region | Proportion of Females with Different Morphotypes, % | Number of Individuals | ||||
|---|---|---|---|---|---|---|
| F1-1 | F1-2 | F2-1 | F2-2 | F2-3 | ||
| Murmansk Region | 0 | 0 | 0 | 25.0 | 75.0 | 4 |
| Krasnoyarsk Territory (Norilsk) | 28.6 | 0 | 57.1 | 0 | 14.3 | 7 |
| Altai Republic | 7.7 | 7.7 | 7.7 | 46.2 | 30.8 | 13 |
| Magadan Region | 50 | 50 | 0 | 0 | 0 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chesnokova, S.V.; Vaulin, O.V.; Zhigulskaya, Z.A.; Novgorodova, T.A. Formica gagatoides Ruzsky, 1904, and Siberian F. kozlovi Dlussky, 1965 (Hymenoptera: Formicidae); Two or One Species? Diversity 2023, 15, 686. https://doi.org/10.3390/d15050686
Chesnokova SV, Vaulin OV, Zhigulskaya ZA, Novgorodova TA. Formica gagatoides Ruzsky, 1904, and Siberian F. kozlovi Dlussky, 1965 (Hymenoptera: Formicidae); Two or One Species? Diversity. 2023; 15(5):686. https://doi.org/10.3390/d15050686
Chicago/Turabian StyleChesnokova, Svetlana V., Oleg V. Vaulin, Zoya A. Zhigulskaya, and Tatiana A. Novgorodova. 2023. "Formica gagatoides Ruzsky, 1904, and Siberian F. kozlovi Dlussky, 1965 (Hymenoptera: Formicidae); Two or One Species?" Diversity 15, no. 5: 686. https://doi.org/10.3390/d15050686