Niche Complementarity and Resistance to Grazing Promote the Invasion Success of Sargassum horneri in North America

1
Department of Ecology, Evolution and Marine Biology, University of California Santa Barbara, Santa Barbara, CA 93106-6150, USA
2
Marine Science Institute, University of California Santa Barbara, Santa Barbara, CA 93106-6150, USA
*
Author to whom correspondence should be addressed.
Diversity 2020, 12(2), 54; https://doi.org/10.3390/d12020054
Submission received: 14 December 2019
/
Revised: 23 January 2020
/
Accepted: 24 January 2020
/
Published: 29 January 2020
(This article belongs to the Special Issue Ecology of Invasive Aquatic Plants)
Abstract
:Invasive species are a growing threat to conservation in marine ecosystems, yet we lack a predictive understanding of ecological factors that influence the invasiveness of exotic marine species. We used surveys and manipulative experiments to investigate how an exotic seaweed, Sargassum horneri, interacts with native macroalgae and herbivores off the coast of California. We asked whether the invasion (i.e., the process by which an exotic species exhibits rapid population growth and spread in the novel environment) of S. horneri is influenced by three mechanisms known to affect the invasion of exotic plants on land: competition, niche complementarity and herbivory. We found that the removal of S. horneri over 3.5 years from experimental plots had little effect on the biomass or taxonomic richness of the native algal community. Differences between removal treatments were apparent only in spring at the end of the experiment when S. horneri biomass was substantially higher than in previous sampling periods. Surveys across a depth range of 0–30 m revealed inverse patterns in the biomass of S. horneri and native subcanopy-forming macroalgae, with S. horneri peaking at intermediate depths (5–20 m) while the aggregated biomass of native species was greatest at shallow (<5 m) and deeper (>20 m) depths. The biomass of S. horneri and native algae also displayed different seasonal trends, and removal of S. horneri from experimental plots indicated the seasonality of native algae was largely unaffected by fluctuations in S. horneri. Results from grazing assays and surveys showed that native herbivores favor native kelp over Sargassum as a food source, suggesting that reduced palatability may help promote the invasion of S. horneri. The complementary life histories of S. horneri and native algae suggest that competition between them is generally weak, and that niche complementarity and resistance to grazing are more important in promoting the invasion success of S. horneri.
1. Introduction
Marine ecosystems are increasingly threatened by invasive species as global trade expands and human-mediated introductions via commercial shipping occur at escalating rates [1,2,3,4,5]. Developing a predictive understanding of factors influencing the success of marine invasive species has clear implications for managing their spread and impacts. Yet relative to terrestrial systems, little is known about the ecological processes that influence marine invasions [6,7]. In terrestrial ecosystems, once an introduced species becomes established, biotic interactions with native species can play a major role in limiting population growth, spread and ecological impacts [8,9,10,11]. These interactions can either promote or inhibit “invasion”, here defined as the process by which an exotic species exhibits rapid population growth and spread in the novel environment [12].
In terrestrial and freshwater plants, biotic interactions such as competition with natives and herbivory can affect invasion success [9,13]. Competition for limited resources among native and invasive species is expected to be most intense when they have similar life histories and resource requirements [14,15,16]; invasion is promoted when exotic plants employ resource acquisition strategies superior to native competitors, reducing their abundance or diversity [17]. Invasion success can also be promoted when exotic species have functional traits or resource requirements that differ from the native biota, which allows them to take advantage of underutilized resources in space and time [16,18,19,20,21]. Such niche complementarity can facilitate invasions by allowing exotics to avoid interacting with natives that have superior competitive abilities. Like native plants, native consumers can promote or hinder invasion depending on their dietary preference. For example, herbivores that prefer exotic plants to natives can inhibit invasion, while those that prefer native plants can facilitate invasion by reducing the strength of competition between exotic and native plants [10,22,23]. Studies aimed at determining the mechanisms affecting the invasiveness of exotic marine macrophytes are needed to derive meaningful generalizations about the role of biotic interactions in influencing the invasibility of a wide range of ecosystems.
The Asian brown alga Sargassum horneri (Turner) C. Agardh, 1820 (Fucales) was first detected in California in 2003 [24] and has since spread throughout southern California and Baja California, Mexico [25]. Several life history characteristics of S. horneri are typical of “weedy” invaders with r-selected traits including broad habitat requirements and high fecundity with >60% of its biomass dedicated to reproductive tissue at its peak fertility [26,27]. It has highly localized propagule dispersal, as well as the ability to disperse long distances via the dislodgement and drifting of buoyant fertile adults [27]. The biomass of S. horneri is strongly seasonal: juveniles prevalent in the summer exhibit rapid growth to several meters in height during the winter, and reproduction and biomass peak in the spring [27]. S. horneri has the potential to compete with native algae by reducing the amount of light reaching algae growing beneath its canopy. Throughout the invaded range, S. horneri has become a dominant macroalga in some areas, but remains rare in others [7,25]. However, it is unclear whether this dominance results from competitive displacement of native species or opportunistic occupation of an underutilized niche.
During its reproductive phase, S. horneri can form dense canopies that shade the bottom, and canopy shading by invasive algae has been shown to cause the decline or exclusion of native seaweeds [28,29,30]. However, it has been hypothesized that the invasion of S. horneri is suppressed in areas dominated by native algae [7], suggesting that niche complementarity rather than competitive superiority accounts for its rapid spread in North America. Detailed information on patterns of distribution of S. horneri and native algae across space (e.g., depths) and through time (e.g., seasons) can provide valuable insight into the relative importance of competition versus niche complementarity in accounting for the invasion success of S. horneri.
The effects of herbivores in structuring temperate marine communities are well documented [31,32,33], but less is known about their potential role in influencing invasions. Exotic seaweeds with traits that deter herbivory (i.e., structural or chemical defenses) can gain an advantage over native competitors in areas with high grazing pressure. Such may be the case for S. horneri as it is in the order Fucales, which is known for having high levels of phenolic compounds that deter grazing [34,35,36]. Thus, preferential consumption of less defended native algae such as laminarian kelps [36,37] could facilitate the spread of S. horneri by weakening competition with other more palatable native algae.
The purpose of this study was to determine the degree to which competition, niche complementarity and herbivory account for patterns of abundance of S. horneri in an area where it has become established. To do this, we documented patterns of co-occurrence between S. horneri and native algae spatially across a depth gradient and temporally over multiple years in experimental plots with S. horneri removed or left intact to evaluate niche complementarity and competition as mechanisms contributing to the invasiveness of S. horneri. If invasion by S. horneri results from its ability to outcompete native algae, then we expected the biomass and taxonomic richness of native algae to increase in areas where we experimentally removed S. horneri. Alternatively, if the invasion success of S. horneri relies on its ability to occupy underutilized resources, then we expected to see little change in the native algal assemblage in response to S. horneri removal. We also performed a field experiment involving the major herbivores to examine their grazing preferences for S. horneri versus other algae. Using a combination of feeding assays and distributional surveys, we tested the hypothesis that herbivores facilitate S. horneri by preferentially consuming native algae.
2. Materials and Methods
2.1. Study System
Field experiments and surveys were conducted on rocky reefs on the leeward side of Santa Catalina Island, located 35 km offshore of Los Angeles, CA, USA. Study reefs consisted of bedrock, boulders and cobble distributed along a moderate slope that transitioned to sand at depths of about 30 m. The reefs were dominated by native macroalgae and the invasive Sargassum horneri. Native macroalgae included the canopy-forming giant kelp Macrocystis pyrifera, subcanopy-forming species of kelp (e.g., Eisenia arborea and Agarum fimbriatum) and fucoid algae (e.g., Sargassum palmeri, Stephanocystis neglecta and Halidrys diocia), and understory-forming foliose and calcified algae. Sessile invertebrates occupied only about 3% of the reef surface. S. horneri has become one of the most common macrophytes on shallow reefs at Santa Catalina Island since its introduction in 2006.
The primary grazers at Santa Catalina Island include sea urchins and herbivorous snails. Centrostephanus coronatus, the most abundant species of urchin, takes refuge in crevices and forages within <1 m from its shelter during the night before returning to the same location before sunrise [38]. This behavior leads to the formation of urchin “halos” where they commonly graze down algae within small home ranges.
2.2. Competition
To test the effects of Sargassum horneri on the abundance and taxonomic richness of native algae, we compared the native algal assemblages in experimental plots from which S. horneri was continually removed (hereafter referred to as S−) with those in unmanipulated control plots with S. horneri left intact (S+) over 3.5 years. We also measured the reduction in the amount of light permeating through its canopy as a potential mechanism of competition. This experiment was conducted at Isthmus Reef (33.4476° N, 118.4898° W) at 6 m depth, within the range where S. horneri is most abundant. Twenty-four 1 m2 plots separated by a distance of at least 2 m were established on areas of reef comprised of >90% rock and with a high density (i.e., at least 30 individuals) of S. horneri. S. horneri was removed from 12 randomly assigned plots (S−) beginning in spring 2014 and every 6 to 12 weeks thereafter until summer 2017. S− plots had a 30 cm wide buffer zone around the perimeter where S. horneri was removed to minimize potential edge effects such as shading by individuals outside of the plot. Removal entailed divers using knives to pry all S. horneri holdfasts off the substrate, minimizing disturbance to the other biota within the plot as much as possible. Since competitive interactions may vary with time and among seasons, we sampled the algal communities in all S+ and S− plots just prior to the initial removal of S. horneri in spring 2014 and quarterly thereafter (i.e., summer, autumn, winter and spring) over three consecutive growing seasons (2014–2015, 2015–2016 and 2016–2017).
Algae were identified to the lowest taxonomic level possible, which in most cases was species (Table S1), and measurements of all understory and subcanopy-forming algae were taken in order to estimate the damp biomass of algae in each plot. The abundance of low-lying understory algae was measured as percent cover using a uniform point contact (UPC) method that involved recording the presence and identity of all algae intersecting 49 points distributed in a grid within each 1 m2 plot. Percent cover was determined as the fraction of points a taxon intersected × 100. Although multiple organisms may intersect a single point if they overlay one another, a taxon was only recorded once at a given point even if it intersected that point multiple times. Using this technique, the percent cover of all taxa combined in a plot can exceed 100%, but the percent cover of any individual species or morphological group cannot. This sampling resolution was sufficient to detect species covering at least 2% of the area in a quadrat. If a species was present in the plot but not recorded at one of the 49 points, then it was assigned a percent cover value of 0.5%. Since percent cover does not necessarily scale with biomass for larger subcanopy-forming algae, we recorded the density and the average size of these taxa. Damp biomass was estimated from density and size data of subcanopy algae and percent cover data of understory algae using taxon-specific relationships obtained from the literature [27,39,40,41] or developed specifically for this project (Table S2).
All but two species of algae recorded in the study plots were native to the region; the non-native Sargassum muticum and Codium fragile occurred in low abundance. Both of these species and S. horneri were excluded from analyses to test specifically for the effects of S. horneri on the native algal assemblages [42]. The surface canopy-forming giant kelp, Macrocystis pyrifera, was present at the beginning of the experiment, but it declined quickly during a warming trend and disappeared by December 2014 for the duration of the study. Consequently, its presence did not factor into our analyses.
The effects of S. horneri removal on the taxonomic richness and aggregate biomass of native algae were evaluated using linear mixed effects models [43]. Taxonomic richness was calculated as the number of unique native algal taxa within each plot, and aggregate biomass was calculated as the summed damp biomass of all native algae within each plot. Since we hypothesized that treatment effects may differ among seasons and develop over time, we included season, treatment (S+ or S−) and days since the start of the experiment (elapsed time) as main effects in the model. To account for variation associated with resampling individual plots, we included plot and the summed damp biomass of native algae within each plot at the start of the experiment prior to the first removal of S. horneri as random effects. Full models with the main effects in question (i.e., season, removal treatment, elapsed time and the interactions of time–removal treatment and season–removal treatment) were compared against null or full models without the effects in question using likelihood ratio tests with chi-square test statistics to select the best fit based on the Akaike Information Criterion (AIC). Model assumptions of normality and homoscedasticity were validated through visual inspection of the residuals, and biomass data were square-root transformed to meet model assumptions. To identify which time periods contributed to the time-by-removal treatment interaction, we used Tukey’s Honest Significant Difference (HSD) post hoc analysis to compare the means of S+ and S− treatments for each sampling period.
Differences in the composition of the algal community between S+ and S− plots were tested using non-metric multi-dimensional scaling (nMDS) and analysis of similarities (ANOSIM). We compared the mean biomass of each taxon in S+ and S− plots in spring and summer 2017, during and after the sampling period when S. horneri removal had a significant effect. We used an unrestricted permutation of raw data (999 permutations) on Bray–Curtis similarity matrices with square-root transformation applied. A similarity percentage (SIMPER) analysis was used to determine the taxa that contributed most to dissimilarity between S+ and S− plots.
To determine the amount of shading caused by the S. horneri canopy we calculated the percent transmission of photosynthetically active radiation (PAR, 400–700 nm) during the spring sampling periods in S− and S+ plots. Light was measured using a handheld spherical quantum sensor (LI-COR Model LI-192) oriented vertically in the center of each plot 30 cm above the bottom. Ten readings of Photosynthetic Photon Flux Density (PPFD in µmol m−2 s−1) were taken in each plot and averaged. Percent transmission was calculated from the average of 10 PPFD readings taken at the surface before and after the dive as:
We assessed how percent transmission of PAR was affected by S. horneri canopy biomass in S+ plots during spring using linear regression. We also tested the hypothesis that the removal of S. horneri increases PAR reaching the bottom compared to unmanipulated plots during spring following the initial removal of S. horneri using a repeated-measures ANOVA with removal treatment as a fixed factor, and plot and year as random factors. We used one-tailed t-tests to determine how the years differed from each other with respect to light transmission because we had an a priori expectation that light would be lower in S+ plots than S– plots. Percent transmission light data were arcsin-transformed prior to analyses to meet the assumptions of ANOVA.
2.3. Complementarity
We examined seasonal patterns of biomass of Sargassum horneri and native algae in the experimental plots described above to test their degree of temporal complementarity. Comparisons of native algae and S. horneri in S+ plots were used to determine whether the seasonality in biomass differed between the two, while comparisons of native algae in S+ and S− plots were used to determine whether seasonal fluctuations in biomass of native algae occurred independent of S. horneri abundance.
We examined the degree of spatial complementarity between S. horneri and native algae by comparing their biomass across the depth range within which most species of brown algae at Santa Catalina Island occur (0–30 m). Scuba divers counted the number of recruit (defined as <5 cm tall) and adult (defined as >5 cm tall) S. horneri and native species of subcanopy-forming macroalgae within 1 m2 quadrats placed every 5 m along transects at four sites that ran perpendicular to shore from the intertidal to 30 m depth or where the reef transitioned to sand, whichever came first. Density data were converted to units of damp biomass using the method described above (see 2.2 Competition). Since these algae grow only on hard bottom substrate, we visually estimated the percent cover of rock within each quadrat and standardized density estimates to m−2 hard bottom. We performed these surveys in April of 2016, the time of year when the biomass of S. horneri reaches its peak [27]. Although smaller native understory species may also compete with S. horneri, limits on bottom time prevented us from sampling them.
Measured depths were adjusted relative to the Mean Lower Low Water (MLLW) and quadrats were binned into depth intervals of 5 m. Between one and three quadrats were sampled within each depth interval at each site, depending on the grade of the reef. The aggregate biomass of native algae within a quadrat was calculated as the sum of the biomass of the juvenile and adult stages of all native species measured. A two-way ANOVA was used to test whether the biomass of S. horneri and the aggregate biomass of native algae varied by depth interval and taxa.
2.4. Herbivory
We performed grazing assays and surveys of benthic algae within and adjacent to urchin halos to assess whether the palatability of S. horneri differed from that of other algae. In September 2016, replicate arrays consisting of Sargassum horneri, its native and introduced congeners S. palmeri and S. muticum and the native kelps Macrocystis pyrifera and Eisenia arborea were deployed at Isthmus Reef for periods of 48 h. Arrays were either exposed to grazing by urchins and snails or placed inside cages nearby that were designed to exclude these grazers. Cages were constructed from 1 cm-gauge plastic mesh and were cylindrical in shape (1 m in height and 0.5 m in diameter) with mesh covering the top. Cages were open at the bottom and a 1 m-wide weighted skirt secured them to the reef and prevented grazers >1 cm from entering. All urchins and snails were removed from the cages at the beginning of each assay.
During each of the four deployments, 15 arrays containing one sample of each of the five target species of algae were placed in urchin halos while another 15 were placed inside cages. Urchin halos were defined as sections of the reef adjacent to a small ledge where >10 urchins were found and grazing activity was apparent from a lack of algae growing within a 30 cm radius. Some herbivorous snails were also present in the halos, including Tegula eiseni, Tegula aureotincta, Megastrea undosa and Norrisia norrisii. Cages were left in the same location for the duration of the experiment, but we selected unique halos for each deployment so that herbivores would be naïve to the arrays. In the day preceding each deployment, we collected and weighed similarly sized blades or thalli of the five target species. Damp weights were quantified prior to deployment and immediately after collection by spin-drying samples for 10 s before weighing them. Three repeat measurements of each sample were taken by re-hydrating the sample and repeating the drying and weighing process. The average of three replicate measurements for each sample was used to optimize our ability to detect small changes in tissue loss.
Herbivore preference was assessed by comparing algal weights measured before and after each deployment in the exposed versus caged arrays. We calculated the percent of biomass lost as:
where G initial and G final represent the mean of the three replicate weights measured for each sample before and after deployment respectively. For each deployment, exposed and caged arrays were randomly paired and the biomass of each species of algae lost due to grazing was calculated as the difference in the change in biomass between paired arrays. One-way ANOVA was used to evaluate whether the biomass lost due to grazing differed by species, and post hoc contrasts were tested for significance with a Tukey HSD test to determine which species were preferentially consumed. Model assumptions of normality and homoscedasticity were validated through visual inspection of the residuals.
To provide a more time-integrated assessment of the feeding preferences of grazers, we tested whether the relative abundance of S. horneri differed from that of native algae in heavily grazed areas during the final deployment. We did this by measuring the percent cover of all subcanopy and understory algae in 1 m2 quadrats placed adjacent to the 15 urchin halos and at 15 nearby reference locations with high algal cover. Percent cover was assessed using the uniform point contact sampling method described above (see 2.2 Competition). We standardized estimates of cover for individual algal taxa to the total cover of subcanopy and understory algae within each quadrat to compare the relative algal composition adjacent to and away from halos. We ignored encrusting algae and unoccupied space in order to focus on the differences between the foliose algal species that are likely to be consumed by the grazers. Algae were identified to the lowest taxonomic level possible, and were analyzed in the following groups: S. horneri, S. palmeri and other native algae (Table S3). We used a two-way ANOVA to test whether the cover of these taxonomic groups differed adjacent to and away from urchin halos, and Tukey HSD post hoc contrasts were used to determine how the taxonomic groups differed from one another. Standardized percent cover data were arcsin-transformed prior to analyses to meet the assumptions of ANOVA.
2.5. Software Used for Statistical Analysis
All univariate statistical models and tests were completed using RStudio (version 1.1.414) for R Statistical Computing Package [44]. Linear mixed models were fit using the lme4 package [45], and post hoc comparisons were performed using the multcomp library [46]. All multivariate analyses were conducted using PRIMER v7.0 [47] and PERMANOVA+ for PRIMER [48].
3. Results
3.1. Competition
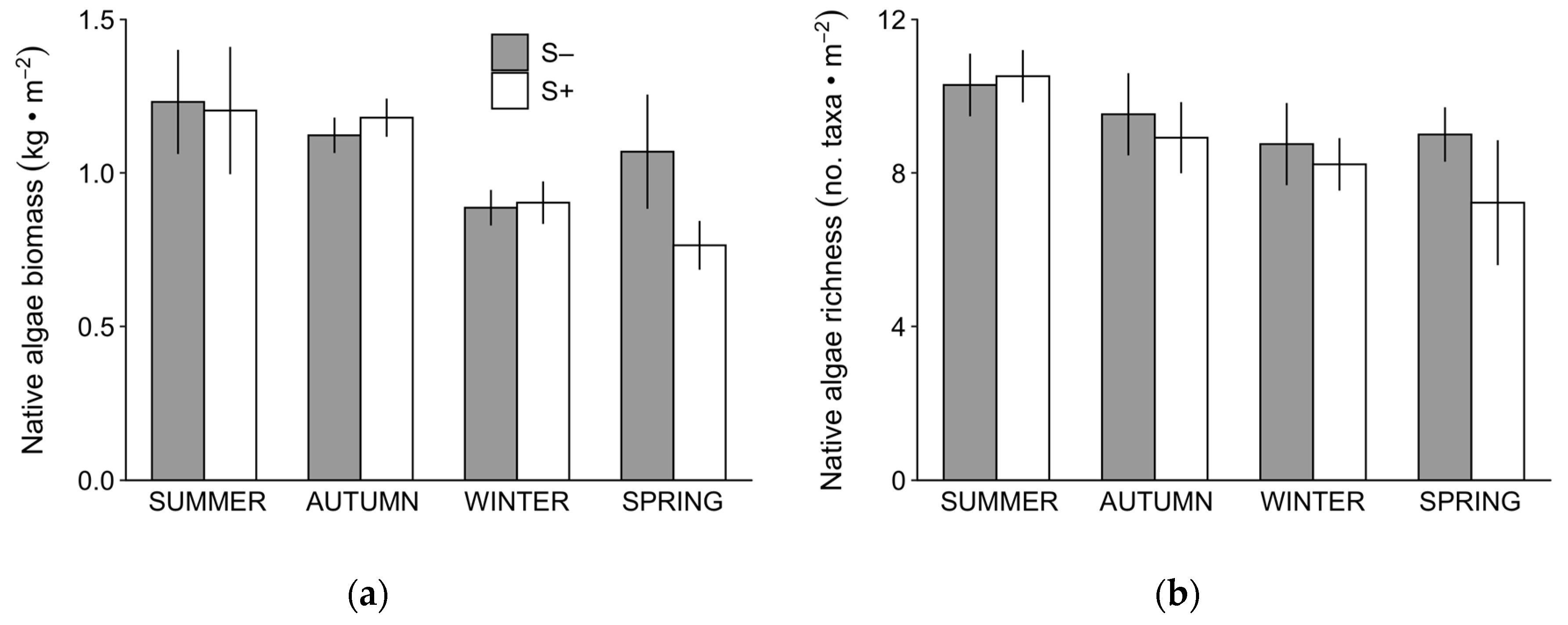
The aggregated biomass and taxonomic richness of native algae varied significantly by season (Table 1). Biomass peaked during summer and autumn, declined by winter and remained low into spring (Figure 1a), while richness also peaked in summer and declined slightly through spring (Figure 1b). The effects of experimentally removing Sargassum horneri on the biomass and species richness of native algae were dependent on season (see season × removal interactions in Table 1).
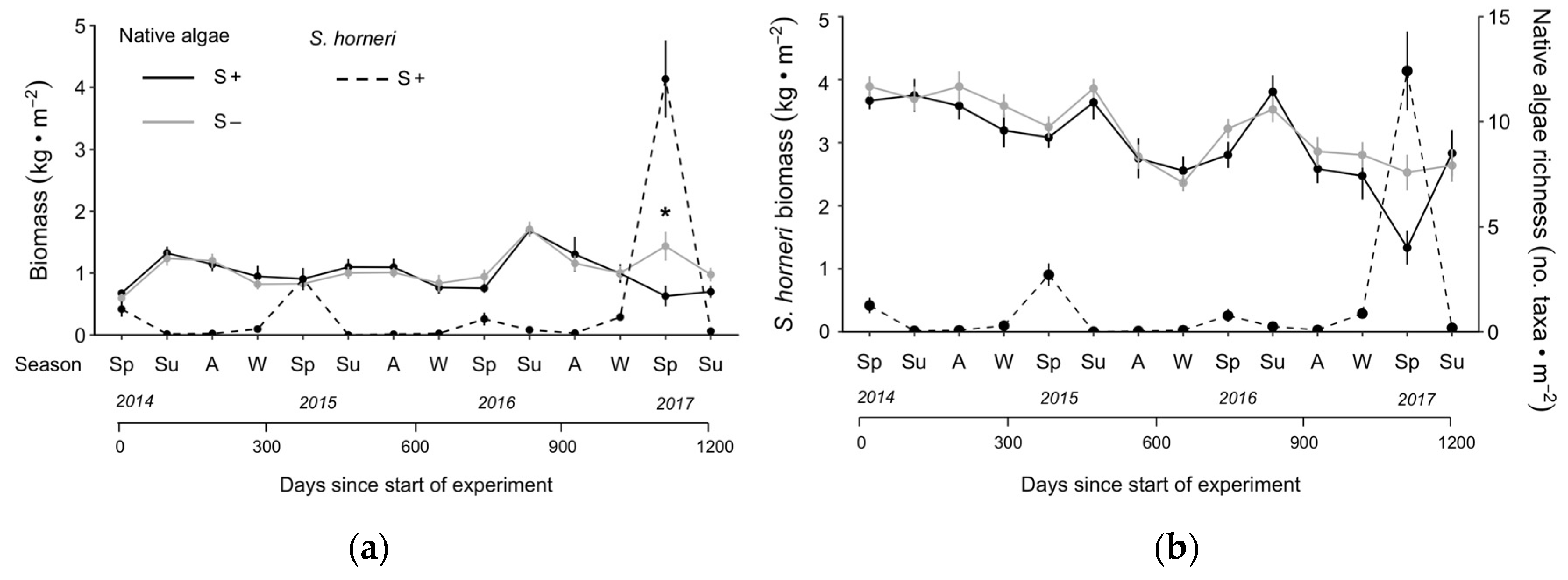
Although there was a significant interaction between season and removal for both biomass and species richness, post hoc tests revealed no particular season as driving the difference (p > 0.05 for all comparisons). Closer examination of the data revealed that the effects of S. horneri removal varied dramatically with days since the start of the experiment (Figure 2) as post hoc testing showed a significant difference in algal biomass between treatments in spring 2017 only, approximately 1100 days since the start of the experiment (Tukey’s HSD, p = 0.002 indicated by * in Figure 2a; all other periods p > 0.05). This difference was driven by a bloom in native algae in S− plots that coincided with a dramatic increase in the biomass of S. horneri in S+ plots (Figure 2a). The biomass of native algae in S− and S+ plots began to converge again by summer 2017 when S. horneri biomass declined. The taxonomic richness of native algae decreased over the course of the study (Figure 2b), independent of the removal of S. horneri (Table 1b).
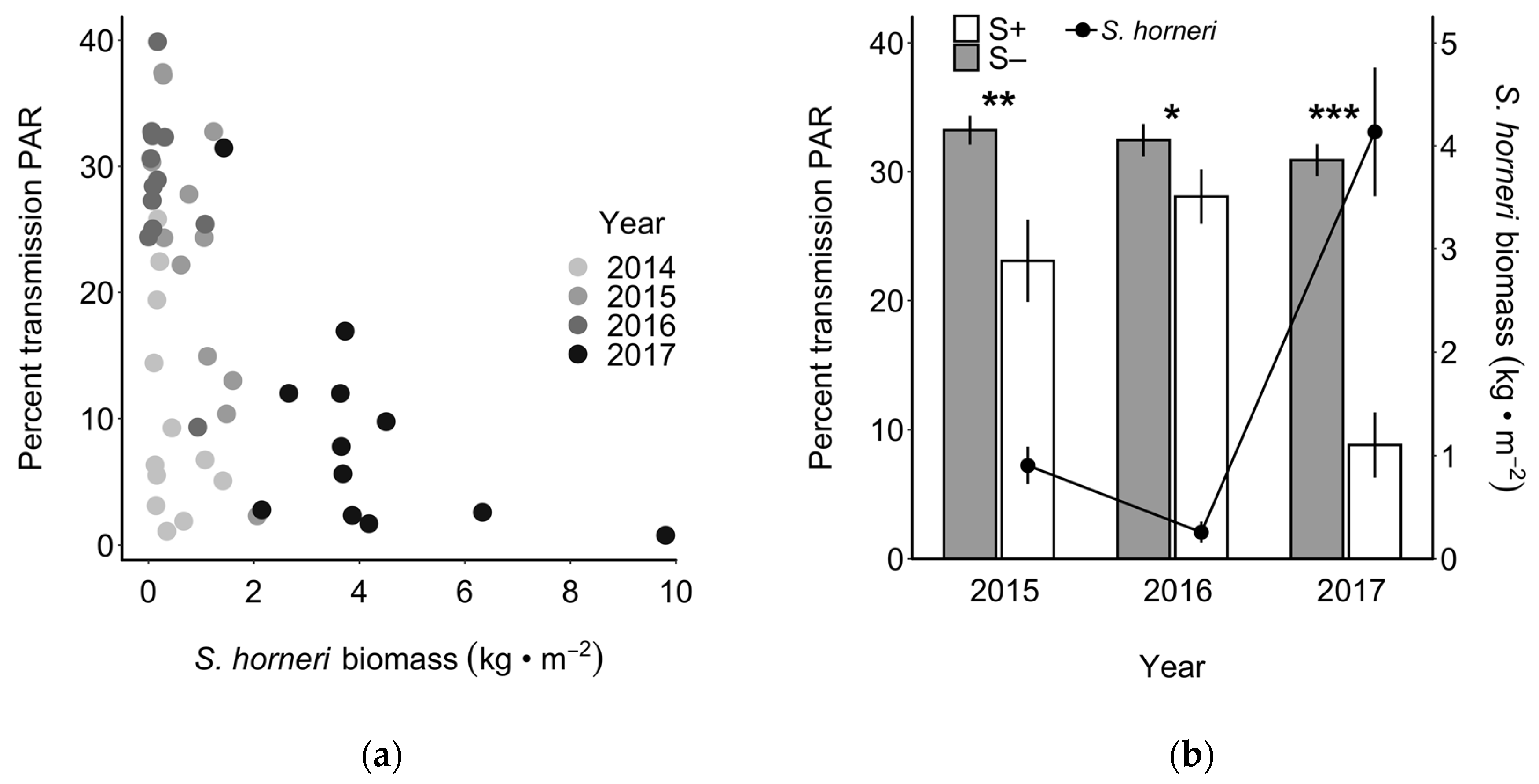
The percent of photosynthetically active radiation (PAR) reaching the bottom in spring was inversely related to the biomass of S. horneri in S+ plots when examined across all four years (Figure 3a; R2 = 0.33, F1,46 = 24.03, p < 0.001). This reduction in light can be attributed to the development of the S. horneri canopy, as evidenced by the significant effect of S. horneri removal on PAR (Figure 3b; ANOVA, F1,22 = 25.2, p < 0.0001). Post hoc tests revealed that S. horneri removal significantly increased PAR in each year (2015: t = 3.00, df = 22, p = 0.003; 2016: t = 1.78, df = 22, p < 0.04; 2017: t = 7.84, df = 22, p < 0.001), especially in 2017 when the biomass of S. horneri in S+ plots was greatest.
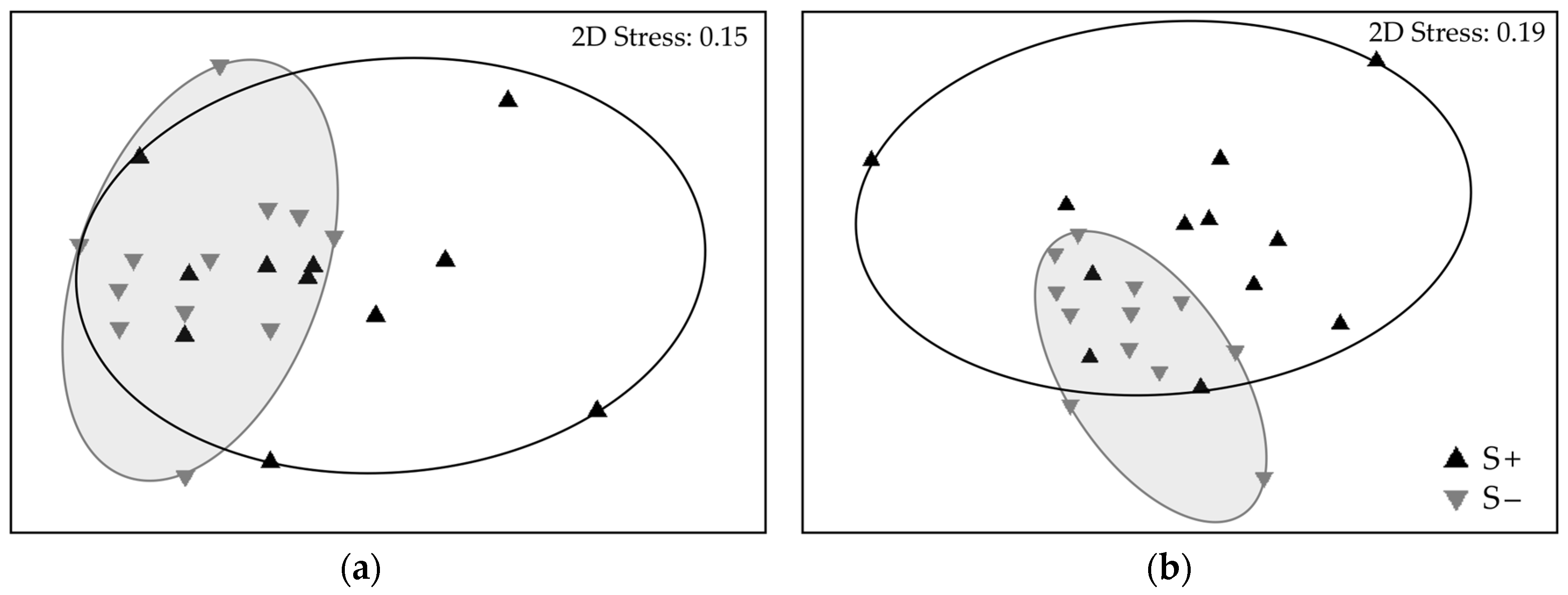
Since S. horneri manipulation had no significant effect on the total biomass of native algae until spring 2017, we restricted our analysis of community structure in S+ and S− plots to data collected during spring and summer 2017. S. horneri removal significantly influenced the native algal assemblages in the spring (Figure 4a; PERMANOVA: Pseudo-F1,21 = 2.90, p = 0.016) and summer (Figure 4b; Pseudo-F1,22 = 2.12, p = 0.041). SIMPER analysis (Table 2) revealed that nearly fifty percent of the dissimilarity between S− and S+ treatments was explained by just two species in spring (Sargassum palmeri and Zonaria farlowii) and three species in summer (Z. farlowii, S. palmeri and Colpomenia sinuosa).
3.2. Complementarity
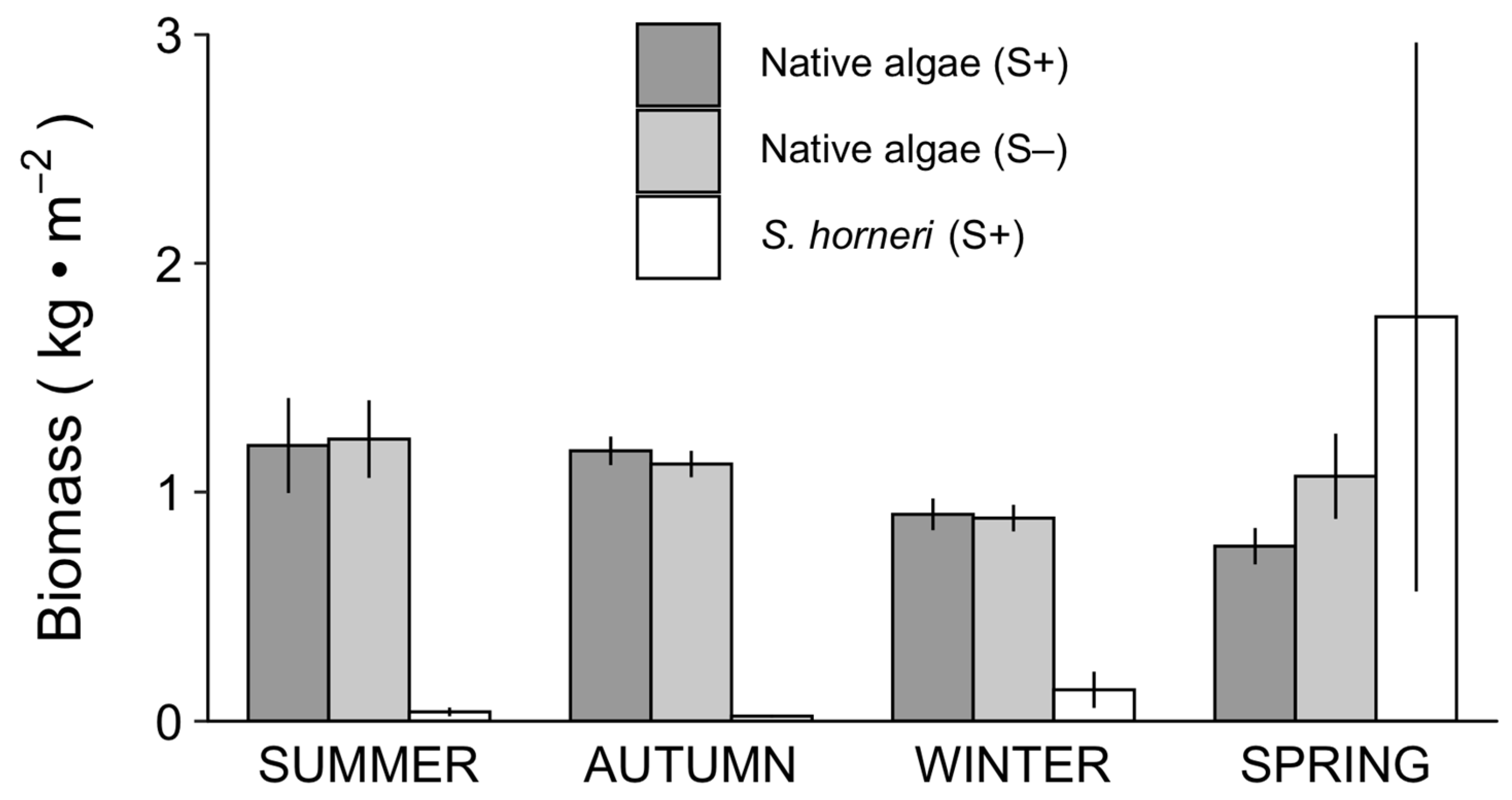
Sargassum horneri displayed a different seasonal pattern in biomass compared to the aggregated biomass of native algae. There was strong seasonality in the biomass of S. horneri in S+ plots, remaining low during summer and autumn, and increasing slightly in winter and dramatically in the spring (Figure 5). By contrast, the aggregated biomass of native algae fluctuated much less throughout the year with highest mean values recorded in summer and biomass declining through winter. In S+ plots, the biomass of native algae continued to decrease into spring, while in S− plots, an increase in the biomass of native algae occurred, which was driven primarily by the native congener S. palmeri in spring 2017.
Results of the depth surveys were consistent with the hypothesis that spatial complementarity with native algae facilitates the invasiveness of S. horneri. Two-way ANOVA revealed that the effect of depth on biomass differed for S. horneri and native algae (F5,1 = 11.78, p < 0.0001 for depth × taxa interaction), and the two were inversely related (Figure 6a). S. horneri was present from the intertidal to the deepest depths sampled, but was most abundant between depths of 5–20 m while the biomass of native algae showed peaks at <5 and >20 m (Figure 6b). The occurrence of specific taxa of native algae varied with depth (Table S4). Biomass of fucoid species (such as Stephanocystis neglecta, Halidrys dioica and Sargassum palmeri) as well as the native kelp Eisenia arborea peaked at shallow depths, while E. arborea also occurred at deeper depths in addition to another native kelp, Agarum fimbriatum.
3.3. Herbivory
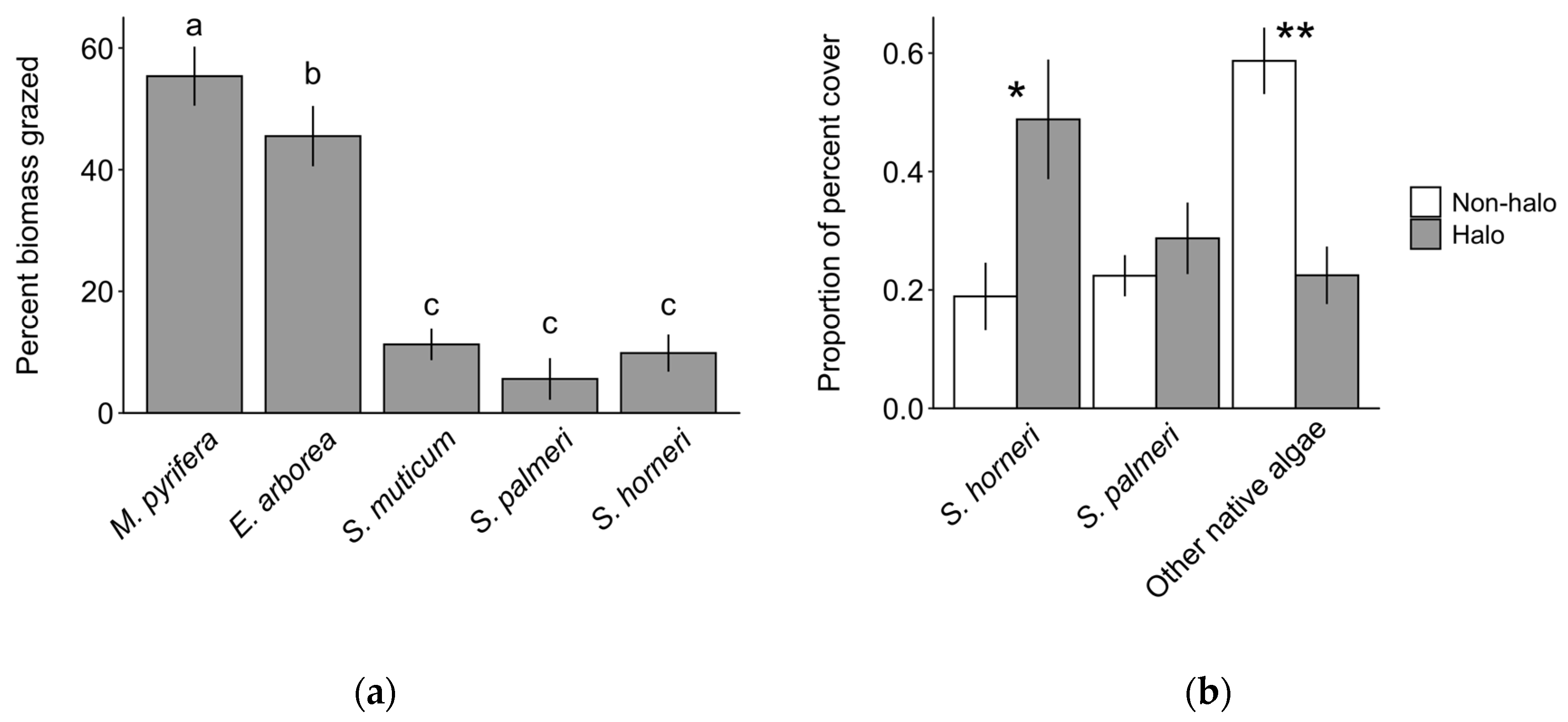
The effects of grazing on the biomass of algae remaining after 48 h assays differed significantly among the five species of algae tested (Figure 7a; ANOVA, F4 = 35.146, p < 0.001). Approximately five times more biomass of Macrocystis pyrifera and four times more biomass of Eisenia arborea was lost due to grazing compared to the three species of Sargassum.
Surveys revealed that the taxonomic composition of algae varied between areas adjacent to and away from urchin halos (Figure 7b; Table S3). There was a significant interaction between taxonomic group and proximity on the relative percent cover (ANOVA, F2,1 = 12.97, p < 0.0001). Post hoc tests revealed that the cover of S. horneri was approximately two times greater near the halos (p = 0.01). By contrast, the proximity to halos had no effect on the cover of S. palmeri (p = 0.98), while that of other native algae taxa near halos was about one third of the level away from halos (p = 0.001).
4. Discussion
The ability of invasive plants to outcompete native flora for limited resources has been well documented [13,49,50] and is the primary mechanism that has been attributed to the successful invasion of Sargassum muticum in the coastal waters off Washington state, USA [30]. Its congener, S. horneri, has a similar potential to displace native algae as a result of shading caused by the high canopy biomass it achieves during the spring [27]. However, we found little evidence that competitive superiority explains the high invasiveness of S. horneri in California as its sustained removal had a minimal effect on the biomass and composition of native algae over a 3.5-year period. Taxonomic richness of the native flora declined over the course of this study but was unresponsive to S. horneri removal. The total biomass of native algae was also unaffected by S. horneri manipulation until 2017, when it increased sharply in plots where S. horneri had been removed. The increase was driven primarily by a perennial congener, S. palmeri. This bloom of S. palmeri coincided with a large increase in the ambient biomass of S. horneri in spring 2017, which dramatically reduced the amount of light reaching the bottom in non-removal plots. Studies of aquatic plants and animals, marsh grasses and marine macroalgae have shown that impacts scale with the abundance of an invader (e.g., [51,52,53,54]). In this study, S. horneri had no detectable effects until it reached extremely high abundance, at which point only modest impacts to the native algal community occurred, driven primarily by a single closely related species.
The strength of competition between introduced and native species can vary spatially and temporally, depending on fluctuations in biomass driven by species’ life histories or environmental factors [55]. The seasonal phenology of the macroalgal community suggested that S. horneri’s peak biomass was generally complementary to that of most of the native macroalgae, whose biomass tended to be highest in summer. This pattern was consistent regardless of the presence of S. horneri (i.e., in removal and non-removal plots) except during spring 2017 when S. horneri was extremely abundant, suggesting it was not a consequence of S. horneri, but rather a natural cycle. This conclusion is substantiated by similar estimates of seasonal biomass of native algae at Santa Catalina Island and elsewhere in southern California prior to invasion by S. horneri [39,56]. Since the giant kelp, M. pyrifera, was absent from our survey and experimental sites throughout nearly the entire course of this study, it did not factor into our analyses. However, like the other native algae we observed, the biomass of M. pyrifera in southern California often peaks in the summer and autumn and drops during winter and spring due to wave-induced disturbance to the canopy [57]. Hence, the success of S. horneri may be attributed in part to the decreased abundance of native algae during its period of peak growth and reproduction.
The depth distribution of S. horneri relative to that of native subcanopy algae could reflect the strength of their competitive interactions or physiological preferences for different parts of the environment. We found that S. horneri displayed spatial complementarity with other subcanopy algae as it was most abundant at intermediate depths (5–20 m), while native algae were most abundant at shallower (<5 m) and deeper (>20 m) depths. That the depth distributions of native subcanopy algae observed in our surveys were similar to those reported by others at Santa Catalina Island prior to the arrival of S. horneri [58,59,60,61] suggests that their lower abundance at intermediate depths was not due to competition with S. horneri.
The reasons for the peak in S. horneri abundance at intermediate depths in our study are unknown. However, the distribution of S. horneri in other regions indicates great versatility in light requirements, and opportunistic growth in situations where competition is minimal. For example, in its native range in Japan, S. horneri grows from the intertidal to 20 m [62] but is most common on shallow reefs from the low intertidal to 4 m [63]. In Baja California, Mexico, near the southern extent of its invaded range, S. horneri has been reported to occur from the intertidal [64,65] to at least 8 m depth [66]. Perhaps robust subcanopy-forming macroalgal communities at Santa Catalina Island deter S. horneri at very deep (>20 m) and very shallow (<5 m) depths, while increased space and light available at intermediate depths allow S. horneri to thrive with minimal competition. Such appears to be the case for the annual Asian kelp, Undaria pinnatifida, whose invasion success in the United Kingdom has been attributed in part to its broad depth range as well as its niche dissimilarities with native algae as the abundances of U. pinnatifida and native algae were inversely correlated along a depth gradient [67].
Our findings revealed that S. horneri has the greatest biomass at depths where, and times when, the abundance of native macroalgae is lowest. The consistent phenology of S. horneri in its native and invaded range [27] and of most native algae in the presence or absence of S. horneri suggest that niche complementarity between them occurs throughout the year. Recent work by Sullaway and Edwards [68] at nearby sites at Santa Catalina Island supports this idea, showing that S. horneri increased rather than decreased levels of community production and respiration in this system. They concluded that S. horneri takes advantage of environmental conditions that disturb native algae and thrives as a consequence of disturbance, rather than causing an ecosystem shift due to its ability to outcompete the native flora [69]. Consistent with this idea is the observation by Caselle et al. [7] that S. horneri abundance at nearby Anacapa Island was significantly lower in older, well-established marine protected areas (MPAs) where the abundance of native algae was high relative to newly established MPAs. These authors argued that the differences in S. horneri abundance between new and old MPAs reflect stronger competition between native algae and S. horneri in the older MPAs where native algae flourish. Thus, niche complementarity may allow S. horneri to achieve high abundance only in places where competition from native algae is not strong.
Herbivores can influence the invasion success of freshwater and marine macrophytes directly through consumption of the invader, or they can mediate interspecific competition through preferential consumption of native species [23,36,37,70]. These preferences may arise from morphological differences or chemical defenses. For example, algae in the order Fucales (which includes the genus Sargassum) typically have high levels of phenolic compounds that are known to deter grazing [37]. Our results are consistent with this hypothesis, demonstrating that grazers consumed the native kelps M. pyrifera and E. arborea while avoiding S. horneri and its congeners S. palmeri and S. muticum. Our results also support the hypothesis posed by Caselle et al. [7] that urchins avoid S. horneri and preferentially consume native algae in areas where they co-occur, thereby reducing the potential for competition between them.
The composition of the benthic algal community reflected the grazer preferences we observed. Centrostephanus centrotus, the most abundant species of sea urchin in our study, is known to display strong feeding preferences, decreasing the abundance of favored species dramatically before switching to less-preferred species [38]. We found that native foliose algae were reduced and S. horneri was more dominant adjacent to urchin halos compared to nearby reference areas. Interestingly, we found no biomass response to grazing by its perennial congener S. palmeri, which is native to southern California. Thus while grazers avoided both species of Sargassum in favor of native foliose algae, only S. horneri responded to a lack of herbivory with increased abundance. It may be that S. horneri is able to colonize space created on the reef more readily than S. palmeri due to its annual life history and high fecundity. Traits related to rapid growth and high fecundity, as well as deterrence to herbivory, are often associated with invasive plants [71]. However, defenses often come at a fitness cost [72] and shorter lived, r-selected plants are not typically heavily defended [73]. Yet S. horneri is a species with r-selected traits that allow it to rapidly colonize available space, and it is also a member of an order of algae that typically displays high levels of chemical defense. These traits undoubtedly contribute to the ability of S. horneri to proliferate in places where interactions with native species are weak.
5. Conclusions
We found that the high propensity of S. horneri to invade southern California reefs results largely from its ability to occupy resources underutilized by native species in space and time and to resist grazing relative to native algae. Its annual life history, high fecundity and capacity for widespread dispersal further enhance its ability to colonize novel habitats. The complementary phenology of S. horneri and native algae suggest competition between them is generally weak, which is consistent with the results of our 3.5-year manipulative experiment. Our findings indicate the greatest potential for competitive interactions between S. horneri and native algae is at intermediate depths during spring when S. horneri peaks in biomass. Future work testing the effects of S. horneri on native algae should focus on this depth range and season. Collectively, our results highlight the importance of considering exotic marine species in the context of the invasibility of native assemblages when assessing their invasiveness and developing management strategies for controlling their spread.
Supplementary Materials
The following are available online at https://www.mdpi.com/1424-2818/12/2/54/s1, Table S1: List of native subcanopy and understory algal taxa recorded in the Sargassum horneri removal experiment, Table S2: Coefficients (a = intercept, b = slope), r2, and p values or SE for formulas to convert size-specific density, or percent cover to damp biomass (g), Table S3: Proportional percent cover of algae adjacent to and away from sea urchin halos, Table S4: Depth distribution of Sargassum horneri and native subcanopy algae. The data presented in this manuscript are available online for download from the Long Term Ecological Research Network Data Portal, doi:10.6073/pasta/2c2237bb3cee86e7c6d9488e8ce2795d [74].
Author Contributions
Conceptualization, D.C.R., L.M.M. and S.J.H.; methodology, D.C.R., L.M.M. and S.J.H.; software, L.M.M.; validation, L.M.M.; formal analysis, L.M.M.; investigation, D.C.R. and L.M.M. resources, D.C.R.; data curation, L.M.M.; writing—original draft preparation, L.M.M.; writing—review and editing, D.C.R., L.M.M. and S.J.H.; visualization, L.M.M.; supervision, D.C.R. and S.J.H.; project administration, L.M.M.; funding acquisition, D.C.R., L.M.M. and S.J.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by California Sea Grant, grant number NA14OAR4170075; National Oceanic and Atmospheric Administrations’ Office of National Marine Sanctuaries Dr. Nancy Foster Scholarship, grant number NA13NOS4290006; and the U.S. National Science Foundation’s (NSF) Long Term Ecological Research Program, grant number OCE1232779. The APC was funded by NSF.
Acknowledgments
We thank the many volunteers and staff from the University of California at Santa Barbara and University of Southern California’s Wrigley Marine Science Center who assisted with fieldwork, especially P. Salinas-Ruiz. We also thank C. D’Antonio for helpful insights during the development of this manuscript, J. Peters for statistical advice, and two anonymous reviewers for their suggestions.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Cohen, A.N.; Carlton, J.T. Accelerating invasion rate in a highly invaded estuary. Science 1998, 279, 555–558. [Google Scholar] [CrossRef] [Green Version]
- Stachowicz, J.J.; Terwin, J.R.; Whitlatch, R.B.; Osman, R.W. Linking climate change and biological invasions: Ocean warming facilitates nonindigenous species invasions. Proc. Natl. Acad. Sci. USA 2002, 99, 15497–15500. [Google Scholar] [CrossRef] [Green Version]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- Sorte, C.J.; Williams, S.L.; Zerebecki, R.A. Ocean warming increases threat of invasive species in a marine fouling community. Ecology 2010, 91, 2198–2204. [Google Scholar] [CrossRef]
- Seebens, H.; Gastner, M.T.; Blasius, B. The risk of marine bioinvasion caused by global shipping. Ecol. Lett. 2013, 16, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Papacostas, K.J.; Rielly-Carroll, E.W.; Georgian, S.E.; Long, D.J.; Princiotta, S.D.; Quattrini, A.M.; Reuter, K.E.; Freestone, A.L. Biological mechanisms of marine invasions. Mar. Ecol. Prog. Ser. 2017, 565, 251–268. [Google Scholar] [CrossRef] [Green Version]
- Caselle, J.C.; Davis, K.; Marks, L.M. Marine management affects the invasion success of a non-native species in a temperate reef system in California, USA. Ecol. Lett. 2018, 21, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Methuen: London, UK, 1958; 181p. [Google Scholar]
- Levine, J.M.; Adler, P.B.; Yelenik, S.G. A meta-analysis of biotic resistance to exotic plant invasions. Ecol. Lett. 2004, 7, 975–989. [Google Scholar] [CrossRef] [Green Version]
- Maron, J.L.; Vilà, M. When do herbivores affect plant invasion? Evidence for the natural enemies and biotic resistance hypotheses. Oikos 2001, 95, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.D.; Hay, M.E. Biotic resistance to plant invasions? Native herbivores prefer non-native plants. Ecol. Lett. 2005, 8, 959–967. [Google Scholar] [CrossRef]
- Valéry, L.; Fritz, H.; Lefeuvre, J.C.; Simberloff, D. In search of a real definition of the biological invasion phenomenon itself. Biol. Invasions 2008, 10, 1345–1351. [Google Scholar] [CrossRef]
- Fleming, J.P.; Dibble, E.D. Ecological mechanisms of invasion success in aquatic macrophytes. Hydrobiologia 2015, 746, 23–37. [Google Scholar] [CrossRef]
- MacArthur, R.; Levins, R. The limiting similarity, convergence, and divergence of coexisting species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Spencer, D.F.; Rejmanek, M. Propagule type influences competition between two submersed aquatic macrophytes. Oecologia 1989, 81, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Petruzzella, A.; Manschot, J.; van Leeuwen, C.H.A.; Grutters, B.M.C.; Bakker, E.S. Mechanisms of invasion resistance of aquatic plant communities. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Madsen, J.D. Predicting invasion success of Eurasian watermilfoil. J. Aquat. Plant Manag. 1998, 36, 28–32. [Google Scholar] [CrossRef]
- Crawley, M.J.; Harvey, P.H.; Purvis, A. Comparative ecology of the native and alien floras of the British Isles. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1996, 351, 1251–1259. [Google Scholar] [CrossRef]
- Lloret, F.; Medail, F.; Brundu, G.; Camarda, I.; Moragues, E.; Rita, J.; Lambdon, P.; Hulme, P. Species attributes and invasion success by alien plants on Mediterranean islands. J. Ecol. 2005, 93, 512–520. [Google Scholar] [CrossRef]
- Mack, R.N. Phylogenetic constraint, absent life forms, and preadapted alien plants: A prescription for biological invasions. Int. J. Plant Sci. 2003, 164, 185–196. [Google Scholar] [CrossRef]
- Muthukrishnan, R.; Hansel-Welch, N.; Larkin, D.J.; Nilsson, C. Environmental filtering and competitive exclusion drive biodiversity-invasibility relationships in shallow lake plant communities. J. Ecol. 2018, 106, 2058–2070. [Google Scholar] [CrossRef]
- Grosholz, E. Avoidance by grazers facilitates spread of an invasive hybrid plant. Ecol. Lett. 2010, 13, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Pulzatto, M.M.; Lolis, L.A.; Louback-Franco, N.; Mormul, R.P. Herbivory on freshwater macrophytes from the perspective of biological invasions: A systematic review. Aquat. Ecol. 2018, 52, 297–309. [Google Scholar] [CrossRef]
- Miller, K.A.; Engle, J.M.; Uwai, S.; Kawai, H. First report of the Asian seaweed Sargassum filicinum Harvey (Fucales) in California, USA. Biol. Invasions 2007, 9, 609–613. [Google Scholar] [CrossRef] [Green Version]
- Marks, L.M.; Salinas-Ruiz, P.; Reed, D.C.; Holbrook, S.J.; Culver, C.S.; Engle, J.M.; Kushner, D.J.; Caselle, J.E.; Freiwald, J.; Williams, J.P.; et al. Range expansion of a non-native, invasive macroalga Sargassum horneri (Turner) C. Agardh, 1820 in the eastern Pacific. BioInvasions Rec. 2015, 4, 243–248. [Google Scholar] [CrossRef]
- Baker, H.G. Characteristics and modes of origin of weeds. In The Genetics of Colonizing Species; Baker, H.G., Stebbins, G.L., Eds.; Academic Press: New York, NY, USA, 1965; Volume X, pp. 147–168. [Google Scholar]
- Marks, L.M.; Reed, D.C.; Holbrook, S.J. Life history traits of the invasive seaweed Sargassum horneri at Santa Catalina Island, California. Aquat. Invasions 2018, 13, 339–350. [Google Scholar] [CrossRef]
- DeWreede, R.E. Sargassum muticum (Fucales, Phaeophyta): Regrowth and interaction with Rhodomelalarix (Ceramiales, Rhodophyta). Phycologia 1983, 22, 153–160. [Google Scholar] [CrossRef]
- Casas, G.; Scrosati, R.; Piriz, L. The invasive kelp Undaria pinnatifida (Phaeophyceae, Laminariales) reduces native seaweed diversity in Nuevo Gulf (Patagonia, Argentina). Biol. Invasions 2004, 6, 411–416. [Google Scholar] [CrossRef]
- Britton-Simmons, K.H. Direct and indirect effects of the introduced alga Sargassum muticum on benthic, subtidal communities of Washington State, USA. Mar. Ecol. Prog. Ser. 2004, 27, 61–78. [Google Scholar] [CrossRef] [Green Version]
- Mann, K.H. Destruction of kelp beds by sea urchins: A cyclical phenomenon or irreversible degradation? Helgolander Wissenshaftlichen Meeresuntersuchungen 1977, 30, 455–732. [Google Scholar] [CrossRef] [Green Version]
- Shears, N.T.; Babcock, R.C. Marine reserves demonstrate top-down control of community structure on temperate reefs. Oecologia 2002, 132, 131–142. [Google Scholar] [CrossRef]
- Lafferty, K.D. Fishing for lobsters indirectly increases epidemics in sea urchins. Ecol. Appl. 2004, 14, 1566–1573. [Google Scholar] [CrossRef]
- Geiselman, J.A.; McConnell, O.J. Polyphenols in brown algae Fucus vesiculosus and Ascophyllum nodusum: Chemical defenses against the marine herbivorous snail, Littorina littorea. J. Chem. Ecol. 1981, 8, 1115–1133. [Google Scholar] [CrossRef] [PubMed]
- Alstyne, K.L. Herbivore grazing increases polyphenolic defenses in the intertidal brown algae Fucus distichus. Ecology 1988, 69, 655–663. [Google Scholar] [CrossRef]
- Estes, J.A.; Steinberg, P.D. Predation, herbivory, and kelp evolution. Paleobiology 1988, 4, 19–36. [Google Scholar] [CrossRef]
- Van Alstyne, K.L.; McCarthy, J.J., III; Hustead, C.L.; Duggins, D.O. Geographic variation in polyphenolic levels of Northeastern Pacific kelps and rockweeds. Mar. Biol. 1999, 133, 371–379. [Google Scholar] [CrossRef]
- Vance, R.R.; Schmitt, R.J. The effect of the predator-avoidance behavior of the sea urchin, Centrostephanus coronatus, on the breadth of its diet. Oecologia 1979, 44, 21–25. [Google Scholar] [CrossRef]
- Harrer, S.L.; Reed, D.C.; Miller, R.J.; Holbrook, S.J. Patterns and controls of the dynamics of net primary production by understory macroalgal assemblages in giant kelp forests. J. Phycol. 2013, 49, 248–257. [Google Scholar] [CrossRef]
- Nelson, J.; Reed, D.; Harrer, S.; Miller, R. SBC LTER: Reef: Coefficients for estimating biomass from body size or percent cover for kelp forest species. Environ. Data Initiat. 2020, in press. [Google Scholar]
- Santa Barbara Coastal Long Term Ecological Research; SBC LTER (University of California Santa Barbara): Santa Barbara, CA, USA, 2019; Unpublished data.
- Thomsen, M.S.; Wernberg, T.; South, P.M.; Schiel, D.R. To include or not to include (the invader in community analyses)? that is the question. Biol. Invasions 2016, 18, 1515–1521. [Google Scholar] [CrossRef]
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models, 2nd ed.; Cambridge University Press: London, UK, 2006; p. 648. [Google Scholar]
- RStudio Team (2019). RStudio: Integrated Development for R. RStudio, Inc.: Boston, MA, USA. Available online: http://www.rstudio.com/ (accessed on 14 December 2019).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial, 1st ed.; PRIMER-E: Plymouth, UK, 2015; p. 300. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2018; p. 214. [Google Scholar]
- Levine, J.M.; Vilà, M.; D’Antonio, C.M.; Dukes, J.S.; Grigulis, K.; Lavorel, S. Mechanisms underlying the impacts of exotic plant invasions. Proc. R. Soc. Lond. B 2003, 270, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Vilà, M.; Weiner, J. Are invasive plant species better competitors than native plant species? – evidence from pair-wise experiments. Oikos 2004, 105, 229–238. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Wernberg, T.; Tuya, F.; Silliman, B.R. Evidence for impacts of nonindigenous macroalgae: A meta-analysis of experimental field studies. J. Phycol. 2009, 45, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Theuekauf, S.J.; Pukett, B.J.; Theuekauf, K.W.; Theuerkauf, E.J.; Eggleston, D.B. Density-dependent role of an invasive marsh grass, Phragmites australis, on ecosystem service provision. PLoS ONE 2017, 12, e0173007. [Google Scholar] [CrossRef] [Green Version]
- White, L.F.; Shurin, J.B. Density dependent effects of an exotic marine macroalga on native community diversity. J. Exp. Mar. Biol. Ecol. 2011, 405, 111–119. [Google Scholar] [CrossRef]
- Olson, E.R.; Doherty, J.M. Macrophyte diversity-abundance relationship with respect to invasive and native dominants. Aquat. Bot. 2014, 119, 111–119. [Google Scholar] [CrossRef]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Benes, K.M.; Carpenter, R.C. Kelp canopy facilitates understory algal assemblage via competitive release during early stages of secondary succession. Ecology 2015, 96, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Reed, D.C.; Rassweiler, A.; Arkema, K. Density derived estimates of standing crop and net primary production in the giant kelp Macrocystis pyrifera. Mar. Biol. 2009, 156, 2077–2083. [Google Scholar] [CrossRef] [Green Version]
- Kastendiek, J. Competitor-mediated coexistence: Interactions among three species of benthic macroalgae. J. Exp. Mar. Biol. Ecol. 1982, 62, 201–210. [Google Scholar] [CrossRef]
- Anderson, T.W. Role of macroalgal structure in the distribution and abundance of a temperate reef fish. Mar. Ecol. Prog. Ser. 1994, 113, 279–290. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Behrens, M.D.; Davis, G.E.; Haaker, P.L.; Kushner, D.J.; Richards, D.V.; Taniguchi, I.K.; Tegner, M.J. Habitat of endangered white abalone, Haliotis sorenseni. Biol. Conservation 2004, 116, 191–194. [Google Scholar] [CrossRef]
- Morrow, K.; Carpenter, R. Shallow kelp canopies mediate macroalgal composition: effects on the distribution and abundance of Corynactis californica (Corallimorpharia). Mar. Ecol. Prog. Ser. 2008, 361, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T. Japanese species of Sargassum subgenus Bactrophycus (Phaeophyta, Fucales). J. Fac. of Sci., Hokkaido Univ. Ser. V (Botany) 1983, 13, 99–246. [Google Scholar]
- Tkaba, M.; Mizokami, A. Seasonal fluctuation of Sargassum communities and vertical distribution of Sargassaceae at Kuroshima Island in the western Akinada. Bull. Hiroshima. Fish. Exp. Stn. 1982, 12, 33–44. [Google Scholar]
- Aguilar-Rosas, L.E.; Aguilar-Rosas, R.; Kawai, H.; Uwai, S.; Valenzuela-Espinoza, E. New record of Sargassum filicinum Harvey (Fucales, Phaeophyceae) in the Pacific coast of Mexico. Algae 2007, 22, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Trejo, G.I.; Ibarra-Obando, S.E.; Aguilar-Rosas, L.E.; Poumian-Tapia, M.; Solana-Arellano, E. Presence of Sargassum horneri at Todos Santos Bay, Baja California, Mexico: its effects on the local macroalgae community. Am. J. Plant Sci. 2015, 6, 2693–2707. [Google Scholar] [CrossRef] [Green Version]
- Riosmena-Rodríguez, R.; Boo, G.H.; López-Vivas, J.M.; Hernández-Velasco, A.; Sáenz-Arroyo, A.; Boo, S.M. The invasive seaweed Sargassum filicinum (Fucales, Phaeophyceae) is on the move along the Mexican Pacific coastline. Bot. Mar. 2012, 55, 547–551. [Google Scholar] [CrossRef]
- Epstein, G.; Hawkins, S.; Smale, D. Identifying niche and fitness dissimilarities in invaded marine macroalgal canopies within the context of contemporary coexistence theory. Sci. Rep. 2019, 9, 8816. [Google Scholar] [CrossRef] [Green Version]
- Sullaway, G.; Edwards, M. Impacts of the non-native alga, Sargassum horneri, on benthic community production in a California Kelp Forest. Mar. Ecol. Prog. Ser. 2020, in press. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Turkington, R. Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 2005, 86, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Bulleri, F.; Tamburello, L.; Beneditti-Cecchi, L. Loss of consumers alters the effects of resident assemblages on the local spread of an introduced macroalga. Oikos 2009, 118, 269–279. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: where do we stand. In Biol. Invasions; Nentwig, W., Ed.; Springer-Verlag: Berlin, Germany, 2007; pp. 97–125. [Google Scholar] [CrossRef]
- Koricheva, J. Meta-analysis of sources of variation in fitness costs of plant antiherbivore defenses. Ecology 2002, 83, 176–190. [Google Scholar] [CrossRef]
- Strauss, S.; Rudgers, J.A.; Lau, J.A.; Irwin, R.E. Direct and ecological costs of resistance to herbivory. Trends Ecol. Evol. 2002, 17, 278–285. [Google Scholar] [CrossRef]
- Marks, L.; Reed, D.; Holbrook, S. SBC LTER: REEF: Data to support "Niche complementarity and resistance to grazing promote the invasion success of Sargassum horneri in North America". Environ. Data Initiat 2020. [Google Scholar] [CrossRef]
Figure 1.
Mean (± SE) biomass (a) and taxonomic richness (b) of all native algae measured in Sargassum horneri-removal (S−; grey bars) and non-removal (S+; white bars) plots. N = 4 years for summer, and 3 years for autumn, winter and spring.
Figure 1.
Mean (± SE) biomass (a) and taxonomic richness (b) of all native algae measured in Sargassum horneri-removal (S−; grey bars) and non-removal (S+; white bars) plots. N = 4 years for summer, and 3 years for autumn, winter and spring.

Figure 2.
Mean (± SE) biomass (a) and taxonomic richness (b) of native algae in Sargassum horneri-removal (S−; grey solid line) and non-removal (S+; black solid line) plots, overlaid by biomass of S. horneri in non-removal plots (S+; dashed line). Asterisk indicates sampling period where multiple comparisons tests (Tukey’s HSD, p < 0.05) indicated a significant difference between treatments. N = 12 plots per sampling period. First data points in each series are from the pre-removal census.
Figure 2.
Mean (± SE) biomass (a) and taxonomic richness (b) of native algae in Sargassum horneri-removal (S−; grey solid line) and non-removal (S+; black solid line) plots, overlaid by biomass of S. horneri in non-removal plots (S+; dashed line). Asterisk indicates sampling period where multiple comparisons tests (Tukey’s HSD, p < 0.05) indicated a significant difference between treatments. N = 12 plots per sampling period. First data points in each series are from the pre-removal census.

Figure 3.
Percent transmission (i.e., percent of surface light reaching the bottom) of photosynthetically active radiation (PAR, 400–700 nm) related to Sargassum horneri biomass during spring. (a) Percent transmission of PAR related to S. horneri biomass in non-removal (S+) plots only, with each sampling year indicated by a different shade. (b) Effect of S. horneri removal on PAR. The left y-axis shows percent transmission of PAR (mean ± SE) in S. horneri-removal (S–; grey bars) and non-removal (S+; white bars) plots, and the right y-axis shows damp biomass of S. horneri (± SE) in non-removal (S+) plots when light measurements were taken. Asterisks indicate sample dates where t-tests indicated significant differences between treatments (*, **, ***: p < 0.05, 0.01, and 0.001, respectively).
Figure 3.
Percent transmission (i.e., percent of surface light reaching the bottom) of photosynthetically active radiation (PAR, 400–700 nm) related to Sargassum horneri biomass during spring. (a) Percent transmission of PAR related to S. horneri biomass in non-removal (S+) plots only, with each sampling year indicated by a different shade. (b) Effect of S. horneri removal on PAR. The left y-axis shows percent transmission of PAR (mean ± SE) in S. horneri-removal (S–; grey bars) and non-removal (S+; white bars) plots, and the right y-axis shows damp biomass of S. horneri (± SE) in non-removal (S+) plots when light measurements were taken. Asterisks indicate sample dates where t-tests indicated significant differences between treatments (*, **, ***: p < 0.05, 0.01, and 0.001, respectively).

Figure 4.
Non-metric multidimensional scaling (nMDS) plots showing benthic algal assemblage structure in plots where Sargassum horneri was removed (S−; grey) and in non-removal plots (S+; black) sampled in 2017. Data are presented by season as (a) spring and (b) summer. N = 24 plots. Analysis used damp biomass with a square root transform and Bray–Curtis similarity index. Two-dimensional (2D) stress values indicate the degree of mismatch between the predicted values from the regression of the similarity matrix and the distances between samples.
Figure 4.
Non-metric multidimensional scaling (nMDS) plots showing benthic algal assemblage structure in plots where Sargassum horneri was removed (S−; grey) and in non-removal plots (S+; black) sampled in 2017. Data are presented by season as (a) spring and (b) summer. N = 24 plots. Analysis used damp biomass with a square root transform and Bray–Curtis similarity index. Two-dimensional (2D) stress values indicate the degree of mismatch between the predicted values from the regression of the similarity matrix and the distances between samples.

Figure 5.
Seasonal mean (± SE) damp biomass of native algae (all species combined) and Sargassum horneri in S. horneri-removal (S−) and non-removal (S+) plots. N = 4 years for summer, and 3 years for autumn, winter and spring.
Figure 5.
Seasonal mean (± SE) damp biomass of native algae (all species combined) and Sargassum horneri in S. horneri-removal (S−) and non-removal (S+) plots. N = 4 years for summer, and 3 years for autumn, winter and spring.

Figure 6.
Spatial co-occurrence of Sargassum horneri and native algae. Data are damp biomass of S. horneri and aggregated damp biomass of all native algae measured within 1 m2 quadrats sampled along transects running perpendicular to shore. (a) Points represent individual quadrats, and are shaded by depth bin. N = 64 quadrats sampled across 4 sites. (b) Bars represent mean (± SE) biomass of S. horneri (white) and the native algae (grey) by 5 m depth bins. The mean and SE of individual species is provided in Table S4. N = 4 sites per depth bin except 25–30 m where N = 3 sites.
Figure 6.
Spatial co-occurrence of Sargassum horneri and native algae. Data are damp biomass of S. horneri and aggregated damp biomass of all native algae measured within 1 m2 quadrats sampled along transects running perpendicular to shore. (a) Points represent individual quadrats, and are shaded by depth bin. N = 64 quadrats sampled across 4 sites. (b) Bars represent mean (± SE) biomass of S. horneri (white) and the native algae (grey) by 5 m depth bins. The mean and SE of individual species is provided in Table S4. N = 4 sites per depth bin except 25–30 m where N = 3 sites.

Figure 7.
Evidence for consumer avoidance of Sargassum horneri. (a) The difference in percent change in biomass (mean ± SE) in randomly paired samples of algae deployed in urchin halos and away from halos inside mesh cages over 48-h periods. Lower-case letters differentiate statistically significant differences between species (Tukey’s HSD, p < 0.05). N = 57 paired arrays from four deployments. (b) Composition of algae adjacent to urchin halos (grey bars) and in nearby reference areas (white bars). Data are the mean proportion of the total percent cover of algae measured in 1 m2 quadrats. N = 15 quadrats sampled per treatment. Asterisks indicate a significant difference between treatments (*, **: p < 0.05, 0.01, respectively).
Figure 7.
Evidence for consumer avoidance of Sargassum horneri. (a) The difference in percent change in biomass (mean ± SE) in randomly paired samples of algae deployed in urchin halos and away from halos inside mesh cages over 48-h periods. Lower-case letters differentiate statistically significant differences between species (Tukey’s HSD, p < 0.05). N = 57 paired arrays from four deployments. (b) Composition of algae adjacent to urchin halos (grey bars) and in nearby reference areas (white bars). Data are the mean proportion of the total percent cover of algae measured in 1 m2 quadrats. N = 15 quadrats sampled per treatment. Asterisks indicate a significant difference between treatments (*, **: p < 0.05, 0.01, respectively).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results from likelihood ratio tests and model selection for determining the influence of experimental removal of Sargassum horneri on the (a) biomass and (b) taxonomic richness of native algae. Independent variables included were: Days since the start of the experiment (Days), S. horneri removal treatment (Removal), and season of the sampling period (Season). Individual variables were tested against the null model and interactions were tested against additive models with the same parameters. Models were ranked according to Akaike Information Criterion (AIC) selection with lower AIC values indicating a better fit of the data. Significance was based on chi-square test statistics. Statistically significant p-values are in bold text.
Table 1.
Results from likelihood ratio tests and model selection for determining the influence of experimental removal of Sargassum horneri on the (a) biomass and (b) taxonomic richness of native algae. Independent variables included were: Days since the start of the experiment (Days), S. horneri removal treatment (Removal), and season of the sampling period (Season). Individual variables were tested against the null model and interactions were tested against additive models with the same parameters. Models were ranked according to Akaike Information Criterion (AIC) selection with lower AIC values indicating a better fit of the data. Significance was based on chi-square test statistics. Statistically significant p-values are in bold text.
| Variables | Model | df | AIC | χ2 | Chi df | p (<χ 2) |
|---|---|---|---|---|---|---|
| a. Biomass of native algae1 | ||||||
| Individual parameters | Null | 4 | 2140.4 | |||
| Days | 5 | 2139.9 | 2.52 | 1 | 0.112 | |
| Removal | 5 | 2141.9 | 0.51 | 1 | 0.473 | |
| Season | 7 | 2106.3 | 40.22 | 3 | < 0.001 | |
| Interactions | Days + Removal | 6 | 2141.4 | |||
| Days × Removal | 7 | 2132.2 | 11.15 | 1 | < 0.001 | |
| Season + Removal | 8 | 2107.7 | ||||
| Season × Removal | 11 | 2104.7 | 9.02 | 3 | 0.029 | |
| b. Richness of native algae | ||||||
| Individual parameters | Null | 4 | 1489.7 | |||
| Days | 5 | 1403.8 | 87.83 | 1 | < 0.001 | |
| Removal | 5 | 1491.6 | 0.04 | 1 | 0.842 | |
| Season | 7 | 1447.1 | 48.54 | 3 | < 0.001 | |
| Interactions | Days + Removal | 6 | 1405.8 | |||
| Days × Removal | 7 | 1407.1 | 0.72 | 1 | 0.397 | |
| Season + Removal | 8 | 1449.1 | ||||
| Season × Removal | 11 | 1446.6 | 8.54 | 3 | 0.036 |
1 data square-root transformed.
Table 2.
Composition of the native algae present in spring and summer 2017 in unmanipulated (S+) plots and those where Sargassum horneri was removed (S−). Data are damp biomass (mean ± SE g·m−2) and the percent contribution of individual taxa to the top 70% of the dissimilarity between S+ and S− treatments in SIMPER analysis.
Table 2.
Composition of the native algae present in spring and summer 2017 in unmanipulated (S+) plots and those where Sargassum horneri was removed (S−). Data are damp biomass (mean ± SE g·m−2) and the percent contribution of individual taxa to the top 70% of the dissimilarity between S+ and S− treatments in SIMPER analysis.
| Taxonomic | Taxon | Spring | Summer | ||||
|---|---|---|---|---|---|---|---|
| Group | S+ | S− | % | S+ | S− | % | |
| Subcanopy | Stephanocystis neglecta | 25.9 ± 15.3 | 27.2 ± 11.3 | 7.6 | 27.5 ± 10.3 | 19.0 ± 8.1 | 5.9 |
| algae | Sargassum palmeri | 415.1 ± 154.1 | 911.3 ± 232.5 | 29.0 | 172.3 ± 45.8 | 262.6 ± 67.6 | 14.3 |
| Understory algae | Articulated coralline spp. | 0 ± 0 | 1.6 ± 1.6 | . | 0.4 ± 0.4 | 1.6 ± 1.6 | . |
| Asparagopsis taxiformis | 0 ± 0 | 0 ± 0 | . | 0.3 ± 0.3 | 1.1 ± 1.1 | . | |
| Brown blade spp. | 0 ± 0 | 6.0 ± 4.3 | . | 0 ± 0 | 0 ± 0 | . | |
| Cladophora graminea | 0 ± 0 | 0 ± 0 | . | 0.1 ± 0.1 | 0 ± 0 | . | |
| Colpomenia sinuosa | 0 ± 0 | 10.6 ± 8.4 | . | 148.3 ± 72.3 | 12.7 ± 5.4 | 12.3 | |
| Chondria californica | 0 ± 0 | 0.3 ± 0.3 | 1.1 ± 0.5 | 0.5 ± 0.4 | |||
| Corallina chilensis | 20.2 ± 10.6 | 30.5 ± 13.0 | 7.2 | 13.6 ± 7.4 | 12.3 ± 7.0 | . | |
| Dictyopteris undulata | 3.9 ± 2.2 | 23.5 ± 6.1 | 6.6 | 14.8 ± 6.7 | 45.9 ± 12.4 | 7.2 | |
| Dictyota spp. | 1.1 ± 0.7 | 20.4 ± 7.7 | . | 24.0 ± 11.2 | 24.0 ± 11.6 | . | |
| Filamentous brown spp. | 0 ± 0 | 0 ± 0 | . | 0.1 ± 0.1 | 0 ± 0 | . | |
| Filamentous green spp. | 0 ± 0 | 0.2 ± 0.2 | . | 0.2 ± 0.2 | 0.5 ± 0.3 | . | |
| Filamentous red spp. | 1.1 ± 1.1 | 1.2 ± 1.0 | . | 0 ± 0 | 2.4 ± 1.9 | . | |
| Green foliose spp. | 0 ± 0 | 0.5 ± 0.5 | . | 0 ± 0 | 0 ± 0 | . | |
| Halicystis ovalis | 0 ± 0 | 0 ± 0 | . | 0.2 ± 0.2 | 0.7 ± 0.4 | . | |
| Haliptylon gracile | 20.6 ± 11.5 | 20.4 ± 7.9 | . | 37.8 ± 21.3 | 28.7 ± 13.4 | 6.6 | |
| Hydroclathrus clathratus | 0 ± 0 | 0 ± 0 | . | 8.5 ± 4.2 | 5.3 ± 4.3 | . | |
| Laurencia pacifica | 0.6 ± 0.6 | 3.6 ± 1.1 | . | 7.7 ± 3.0 | 11.3 ± 5.2 | . | |
| Lithothrix aspergillum | 19.8 ± 10.0 | 21.0 ± 11.5 | 6.5 | 33.7 ± 20.9 | 25.1 ± 15.0 | 6.3 | |
| Plocamium cartilagineum | 2.7 ± 2.3 | 2.2 ± 1.5 | . | 17.3 ± 10.5 | 2.4 ± 1.4 | . | |
| Pterocladia capillacea | 4.6 ± 4.6 | 5.3 ± 4.3 | . | 8.5 ± 4.2 | 6.4 ± 4.3 | . | |
| Rhodymenia californica | 0 ± 0 | 1.9 ± 1.3 | . | 1 ± 0.5 | 0.6 ± 0.4 | . | |
| Scytosiphon lomentaria | 0 ± 0 | 0 ± 0 | . | 1 ± 0.7 | 0 ± 0 | . | |
| Zonaria farlowii | 171.9 ± 34.9 | 347.6 ± 72.2 | 16.2 | 182.7 ± 47.1 | 513.2 ± 84.7 | 17.7 | |
| Cumulative % contribution to dissimilarity | - | - | 73.1 | - | - | 70.3 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marks, L.M.; Reed, D.C.; Holbrook, S.J. Niche Complementarity and Resistance to Grazing Promote the Invasion Success of Sargassum horneri in North America. Diversity 2020, 12, 54. https://doi.org/10.3390/d12020054
AMA Style
Marks LM, Reed DC, Holbrook SJ. Niche Complementarity and Resistance to Grazing Promote the Invasion Success of Sargassum horneri in North America. Diversity. 2020; 12(2):54. https://doi.org/10.3390/d12020054
Chicago/Turabian StyleMarks, Lindsay M., Daniel C. Reed, and Sally J. Holbrook. 2020. "Niche Complementarity and Resistance to Grazing Promote the Invasion Success of Sargassum horneri in North America" Diversity 12, no. 2: 54. https://doi.org/10.3390/d12020054
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.