Transactivation of miR-202-5p by Steroidogenic Factor 1 (SF1) Induces Apoptosis in Goat Granulosa Cells by Targeting TGFβR2

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Follicular Separation, Cell Isolation, and Culture
2.2. RNA Isolation and qPCR
2.3. Small RNAs Sequencing of FF-EVs
2.4. Plasmids and siRNAs
2.5. Cell Transfection
2.6. Estradiol ELISA Assay
2.7. Western Blotting (WB)
2.8. Chromatin Immunoprecipitation (ChIP)-PCR Assay
2.9. Dual Luciferase Assays
2.10. Apoptosis Assays
2.11. Cell Proliferation Assays
2.12. Statistical Analysis
3. Results
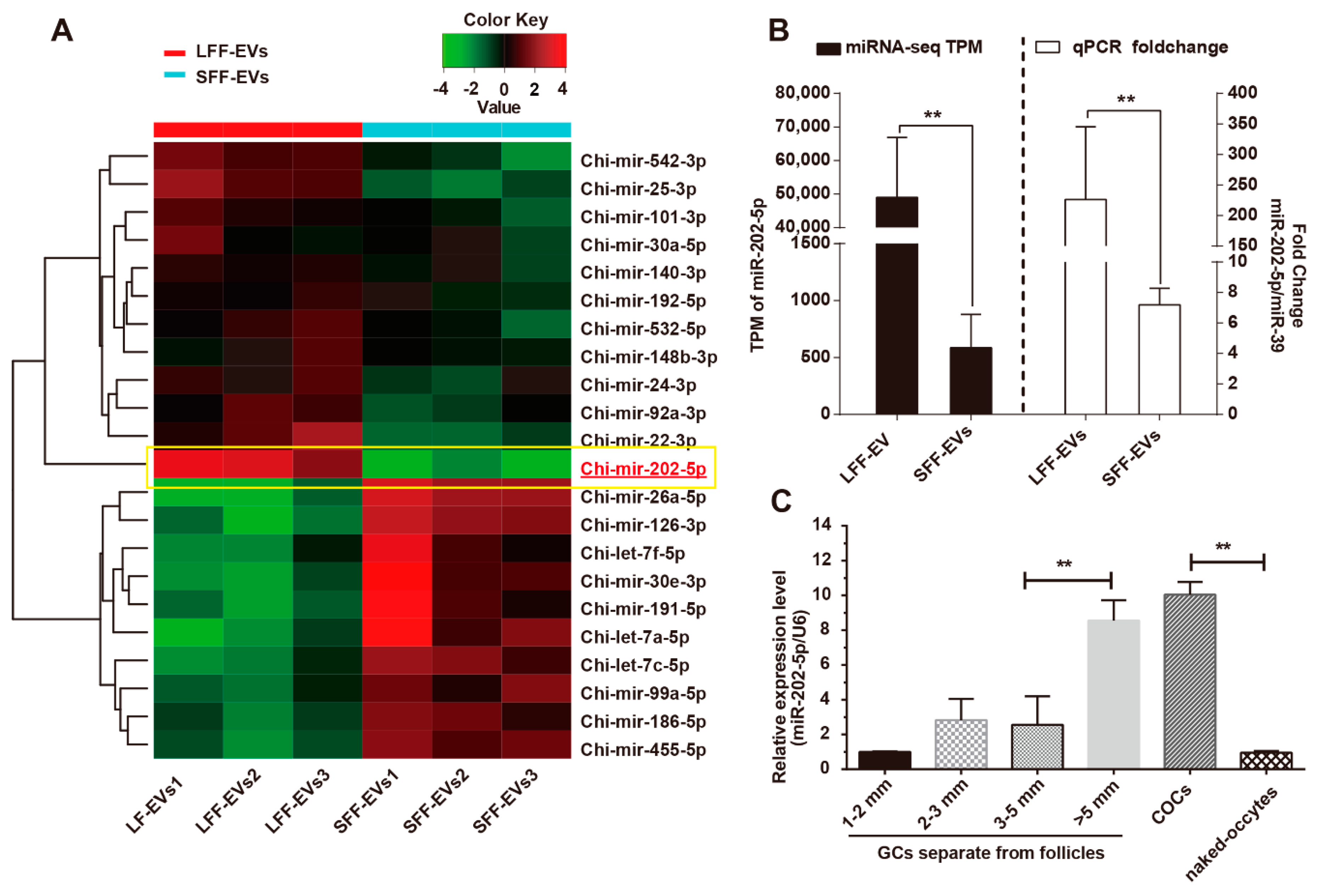
3.1. miR-202-5p Was Highly Expressed in Growing Follicles
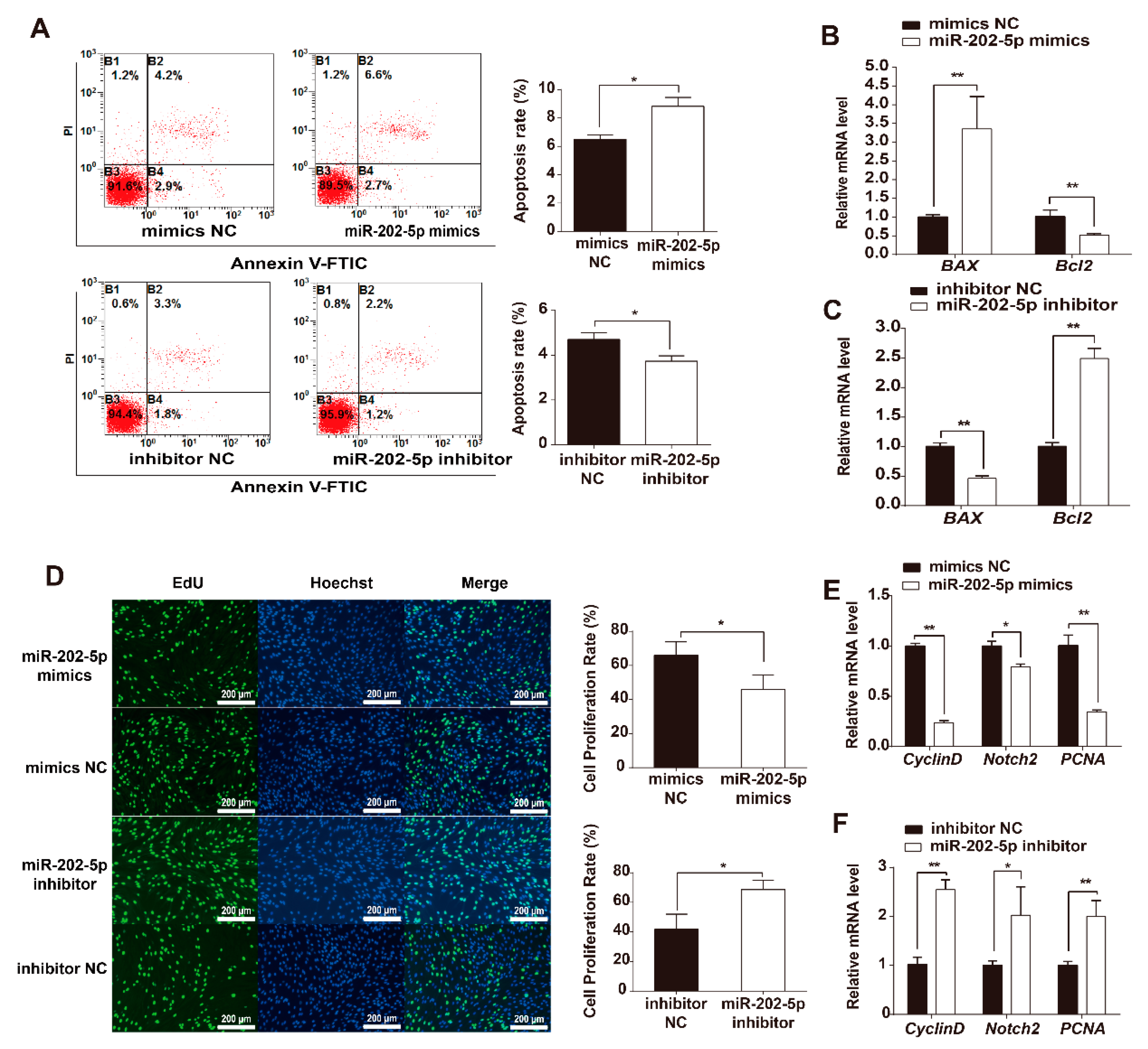
3.2. miR-202-5p Induces GC Apoptosis and Suppresses GC Proliferation In Vitro
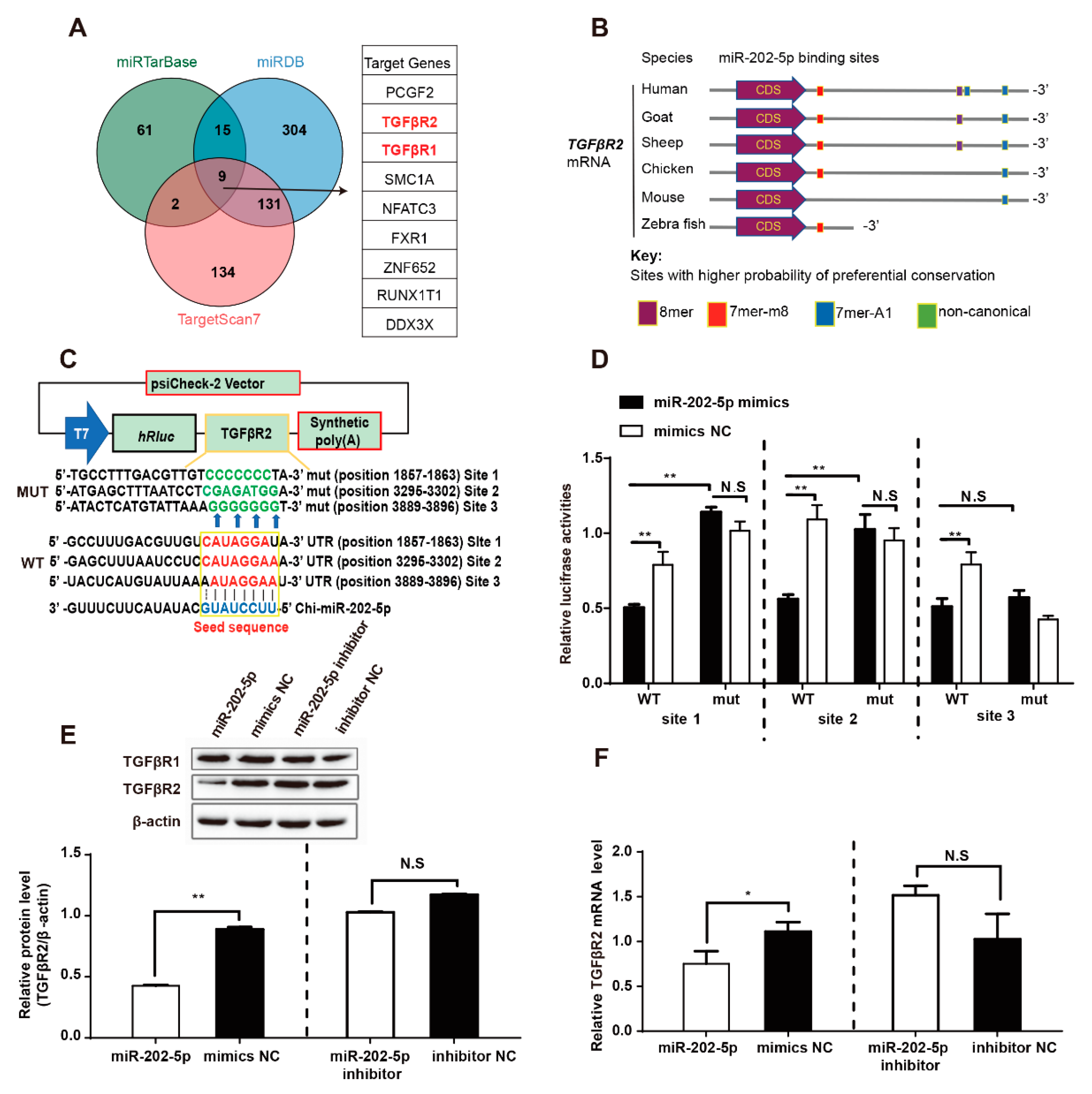
3.3. miR-202-5p Binds and Downregulates TGFβR2 mRNA Expression in GCs
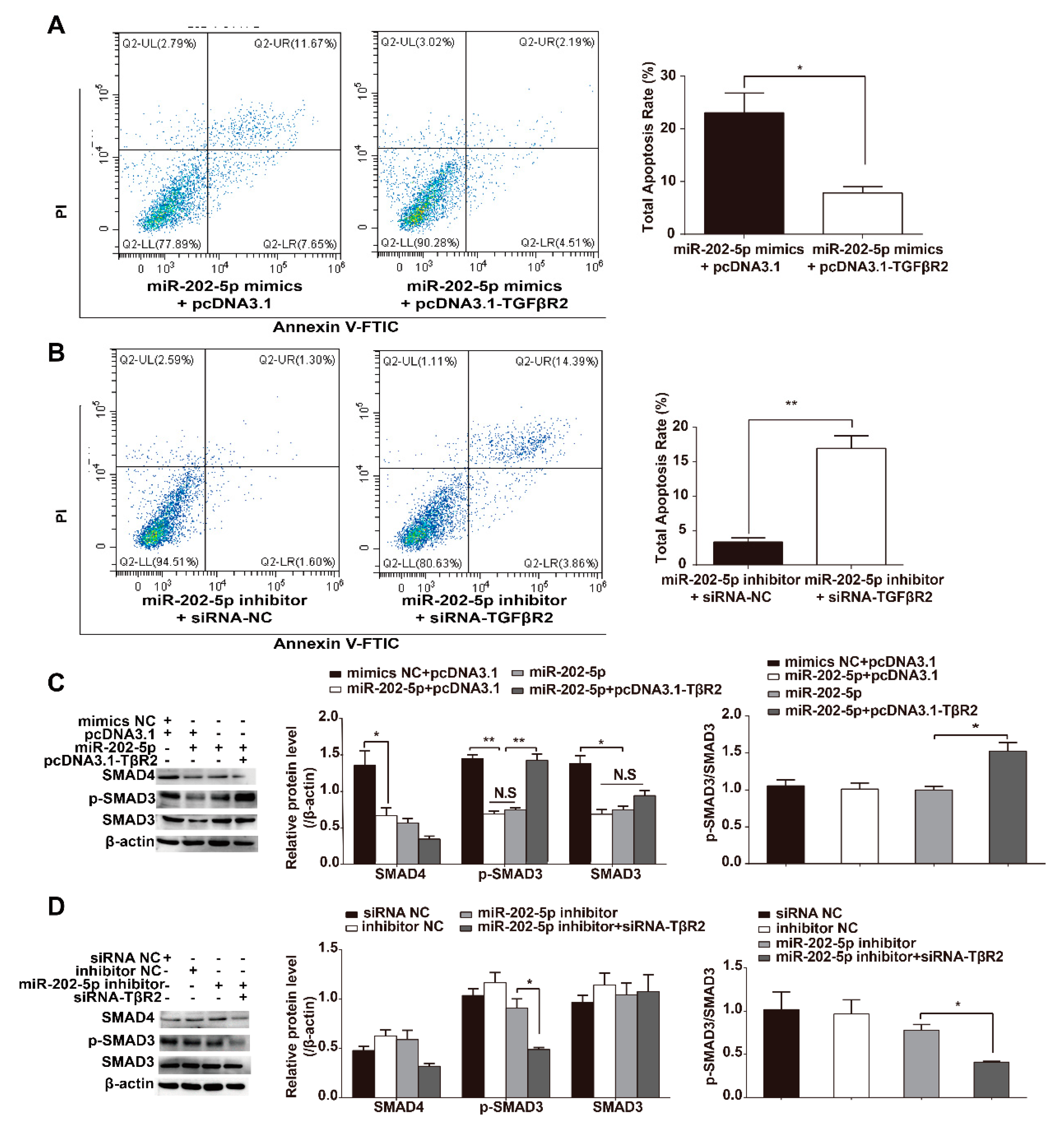
3.4. miR-202-5p Inhibits TGF-β/SMAD Signaling in GCs
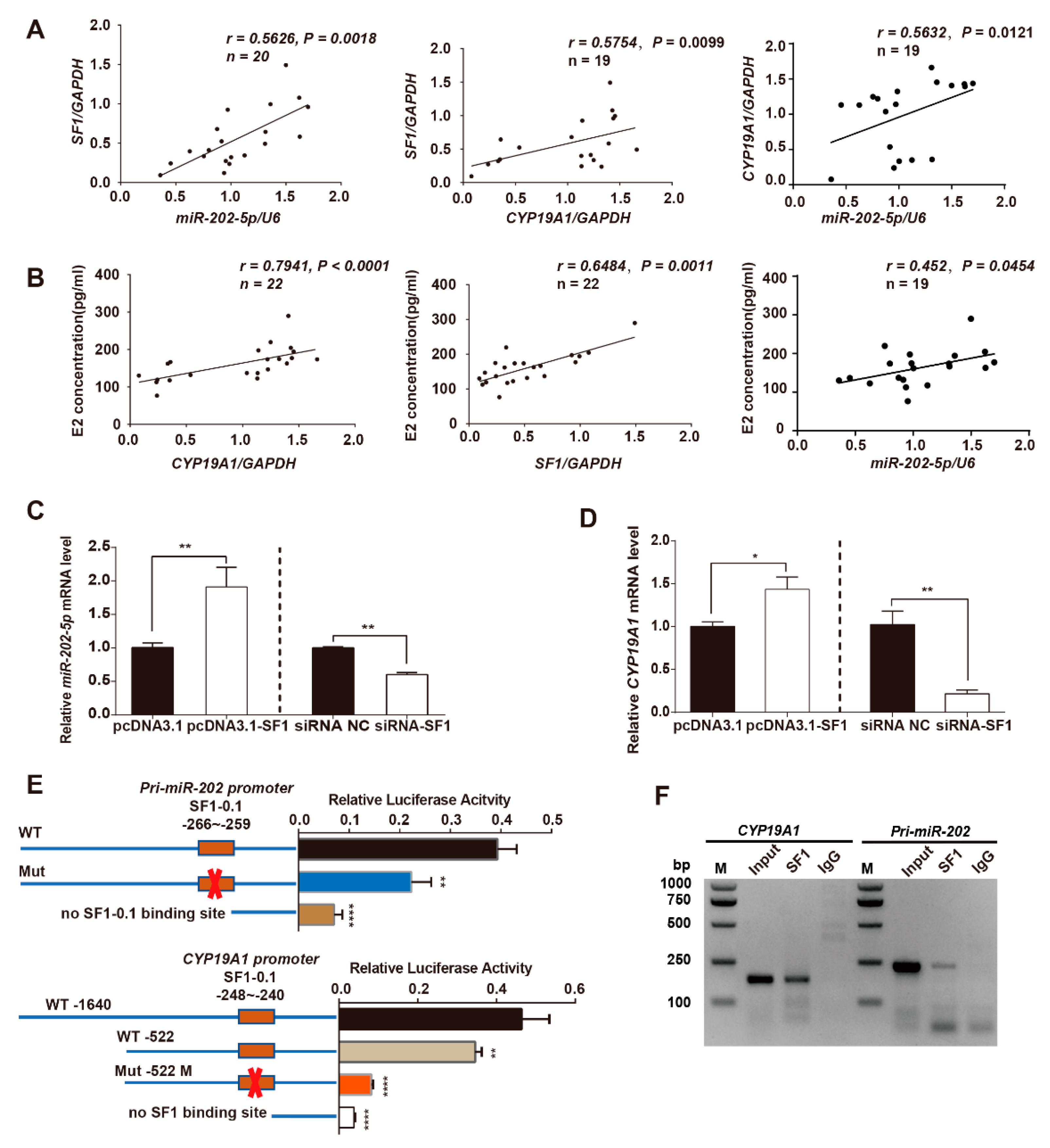
3.5. SF1 Promotes miR-202 and CYP19A1 Expression through Binding to Their Promoter Regions
3.6. miR-202-5p Mediates SF1 Regulation of the Canonical TGF-β/SMAD Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Ethics Statement
Data Availability Statement
References
- Jolly, P.D. Morphological evidence of apoptosis and the prevalence of apoptotic versus mitotic cells in the membrana granulosa of ovarian follicles during spontaneous and induced atresia in ewes. Boil. Reprod. 1997, 56, 837–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, F.; Inoue, N.; Manabe, N.; Ohkura, S. Follicular growth and atresia in mammalian ovaries: Regulation by survival and death of granulosa cells. J. Reprod. Dev. 2012, 58, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terenina, E.; Fabre, S.; Bonnet, A.; Monniaux, D.; Robert-Granie, C.; SanCristobal, M.; Sarry, J.; Vignoles, F.; Gondret, F.; Monget, P.; et al. Differentially expressed genes and gene networks involved in pig ovarian follicular atresia. Physiol. Genom. 2017, 49, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Manabe, N.; Goto, Y.; Matsuda-Minehata, F.; Inoue, N.; Maeda, A.; Sakamaki, K.; Miyano, T. Regulation mechanism of selective atresia in porcine follicles: Regulation of granulosa cell apoptosis during atresia. J. Reprod. Dev. 2004, 50, 493–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, F.M.; Gorospe, W.C. Biochemical Identification of Apoptosis (Programmed Cell Death) in Granulosa Cells: Evidence for a Potential Mechanism Underlying Follicular Atresia*. Endocrinology 1991, 129, 2415–2422. [Google Scholar] [CrossRef]
- Sontakke, S.D.; Mohammed, B.T.; McNeilly, A.S.; Donadeu, F.X. Characterization of microRNAs differentially expressed during bovine follicle development. Reproduction 2014, 148, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Navakanitworakul, R.; Hung, W.-T.; Gunewardena, S.; Davis, J.S.; Chotigeat, W.; Christenson, L.K. Characterization and Small RNA Content of Extracellular Vesicles in Follicular Fluid of Developing Bovine Antral Follicles. Sci. Rep. 2016. [Google Scholar] [CrossRef]
- Liu, J.; Yao, W.; Yao, Y.; Du, X.; Zhou, J.; Ma, B.; Liu, H.; Li, Q.; Pan, Z. MiR-92a inhibits porcine ovarian granulosa cell apoptosis by targetingSmad7gene. Febs Lett. 2014, 588, 4497–4503. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Li, X.; Yao, Y.; Li, Q.; Pan, Z.; Li, Q. miR-1275 controls granulosa cell apoptosis and estradiol synthesis by impairing LRH-1/CYP19A1 axis. Biochim. Et Biophys. Acta (Bba) - Bioenerg. 2018, 1861, 246–257. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, J.; Pan, Z.; Du, X.; Li, X.; Ma, B.; Yao, W.; Li, Q.; Liu, H. The let-7g microRNA promotes follicular granulosa cell apoptosis by targeting transforming growth factor-β type 1 receptor. Mol. Cell. Endocrinol. 2015, 409, 103–112. [Google Scholar] [CrossRef]
- Wainwright, E.N.; Jorgensen, J.S.; Kim, Y.; Truong, V.; Bagheri-Fam, S.; Davidson, T.; Svingen, T.; Fernandez-Valverde, S.L.; McClelland, K.S.; Taft, R.J.; et al. Sox9 regulates microrna mir-202-5p/3p expression during mouse testis differentiation1. Biol. Reprod 2013, 89, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Cai, T.; Zheng, C.; Lin, X.; Wang, G.; Liao, S.; Wang, X.; Gan, H.; Zhang, D.; Hu, X. Microrna-202 maintains spermatogonial stem cells by inhibiting cell cycle regulators and rna binding proteins. Nucleic Acids Res. 2017, 45, 4142–4157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabaja, A.A.; Mielnik, A.; Robinson, B.D.; Wosnitzer, M.S.; Schlegel, P.N.; Paduch, D.A. Possible germ cell-sertoli cell interactions are critical for establishing appropriate expression levels for the sertoli cell-specific microrna, mir-202-5p, in human testis. Basic Clin. Androl. 2015, 25, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalak, P.; Malone, J.H. Testis-derived microRNA profiles of African clawed frogs (Xenopus) and their sterile hybrids. Genom. 2008, 91, 158–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armisen, J.; Gilchrist, M.J.; Wilczynska, A.; Standart, N.; Miska, E.A. Abundant and dynamically expressed mirnas, pirnas, and other small rnas in the vertebrate xenopus tropicalis. Genome Res. 2009, 19, 1766–1775. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, W.; Jin, Y.; Jia, P.; Jia, K.; Yi, M. Mir-202-5p is a novel germ plasm-specific microrna in zebrafish. Sci. Rep. 2017. [Google Scholar] [CrossRef]
- Gay, S.; Bugeon, J.; Bouchareb, A.; Henry, L.; Delahaye, C.; Legeai, F.; Montfort, J.; Le Cam, A.; Siegel, A.; Bobe, J.; et al. MiR-202 controls female fecundity by regulating medaka oogenesis. PloS Genet. 2018. [Google Scholar] [CrossRef]
- Donadeu, F.X.; Sontakke, S.D.; Ioannidis, J. And MicroRNA indicators of follicular steroidogenesis. Reprod. Fertil. Dev. 2016. [Google Scholar] [CrossRef]
- Donadeu, F.; Schauer, S. Differential miRNA expression between equine ovulatory and anovulatory follicles. Domest. Anim. Endocrinol. 2013, 45, 122–125. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Liang, Z.; Jiahao, C.; Zhizhong, L.; Xianxin, L.; Xueda, H.; Yi, H.; Xiaokun, Z.; Chaozhao, L.; Yong, W.; Liang, S. Integrated profiling of micrornas and mrnas: Micrornas located on xq27.3 associate with clear cell renal cell carcinoma. PLoS ONE 2010. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E.A.J.; Siiteri, P.K. The involvement of human placental microsomal cytochrome P-450 in aromatization. J. Boil. Chem. 1974, 249, 5373–5378. [Google Scholar]
- Guigon, C.J.; Mazaud, S.; Forest, M.G.; Brailly-Tabard, S.; Coudouel, N.; Magre, S. Unaltered Development of the Initial Follicular Waves and Normal Pubertal Onset in Female Rats after Neonatal Deletion of the Follicular Reserve. Endocrinology 2003, 144, 3651–3662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Agno, J.E.; Edson, M.A.; Nagaraja, A.K.; Nagashima, T.; Matzuk, M.M. Transforming Growth Factor β Receptor Type 1 Is Essential for Female Reproductive Tract Integrity and Function. PLoS Genet. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.; Pan, Z.; Li, Q.; Liu, H.; Li, Q. SMAD4 feedback regulates the canonical TGF-β signaling pathway to control granulosa cell apoptosis. Cell Death Dis. 2018. [Google Scholar] [CrossRef]
- Du, X.; Zhang, L.; Li, X.; Pan, Z.; Liu, H.; Li, Q. TGF-β signaling controls FSHR signaling-reduced ovarian granulosa cell apoptosis through the SMAD4/miR-143 axis. Cell Death Dis. 2016. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Wang, S.; Luo, A.; Ding, T.; Lai, Z.; Shen, W.; Ma, X.; Cao, C.; Shi, L.; Jiang, J.; et al. Expression Patterns and Regulatory Functions of MicroRNAs During the Initiation of Primordial Follicle Development in the Neonatal Mouse Ovary1. Boil. Reprod. 2013. [Google Scholar] [CrossRef]
- Yao, G.; Yin, M.; Lian, J.; Tian, H.; Liu, L.; Li, X.; Sun, F. MicroRNA-224 Is Involved in Transforming Growth Factor-β-Mediated Mouse Granulosa Cell Proliferation and Granulosa Cell Function by Targeting Smad4. Mol. Endocrinol. 2010, 24, 540–551. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Du, X.; Zhou, J.; Pan, Z.; Liu, H.; Li, Q. Microrna-26b functions as a proapoptotic factor in porcine follicular granulosa cells by targeting sma-and mad-related protein 4. Biol. Reprod. 2014. [Google Scholar] [CrossRef]
- Moustakas, A.; Heldin, C.-H. The regulation of TGF signal transduction. Development 2009, 136, 3699–3714. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, Y.; Xu, X.; Li, J.; Yuan, F.; Bo, S.; Qiao, J.; Xia, G.; Su, Y.; Zhang, M. Transforming growth factor-β is involved in maintaining oocyte meiotic arrest by promoting natriuretic peptide type C expression in mouse granulosa cells. Cell Death Dis. 2019. [Google Scholar] [CrossRef]
- Li, Y.; Jin, Y.; Liu, Y.; Shen, C.; Dong, J.; Xu, J. SMAD3 regulates the diverse functions of rat granulosa cells relating to the FSHR/PKA signaling pathway. Reproduction 2013, 146, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Schilling, B.; Yeh, J. Expression of transforming growth factor (TGF)-β1, TGF-β2, and TGF-β3 and of type I and II TGF-β receptors during the development of the human fetal ovary. Fertil. Steril. 1999, 72, 147–153. [Google Scholar] [CrossRef]
- Roy, S.K.; Kole, A.R. Ovarian transforming growth factor-beta (TGF-beta) receptors: In-vitro effects of follicle stimulating hormone, epidermal growth factor and TGF-beta on receptor expression in human preantral follicles. Mol. Hum. Reprod. 1998, 4, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Mody, H.R.; Hung, S.W.; Pathak, R.K.; Griffin, J.; Cruz-Monserrate, Z.; Govindarajan, R. miR-202 Diminishes TGFβ Receptors and Attenuates TGFβ1-Induced EMT in Pancreatic Cancer. Mol. Cancer Res. 2017, 15, 1029–1039. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Fan, B.; Zhao, Y.; Fang, J. MicroRNA-202 inhibits cell proliferation, migration and invasion of glioma by directly targeting metadherin. Oncol. Rep. 2017, 38, 1670–1678. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Cai, J.; Xie, W.; Luo, H.; Yang, F. miR-202 suppresses prostate cancer growth and metastasis by targeting PIK3CA. Exp. Med. 2018, 16, 1499–1504. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, C.; Michael, D.; Mahendroo, M.; Simpson, E. Exon-specific northern analysis and rapid amplification of cDNA ends (RACE) reveal that the proximal promoter II (PII) is responsible for aromatase cytochrome P450 (CYP19) expression in human ovary. Mol. Cell. Endocrinol. 1993, 97, 1–6. [Google Scholar] [CrossRef]
- Rice, D.A.; Mouw, A.R.; Bogerd, A.M.; Parker, K.L. A shared promoter element regulates the expression of three steroidogenic enzymes. Mol. Endocrinol. 1991, 5, 1552–1561. [Google Scholar] [CrossRef] [Green Version]
- Morohashi, K.; Honda, S.; Inomata, Y.; Handa, H.; Omura, T. A common trans-acting factor, Ad4-binding protein, to the promoters of steroidogenic P-450s. J. Boil. Chem. 1992, 267, 17913–17919. [Google Scholar]
- Parker, K.L.; Schimmer, B.P. Steroidogenic factor 1: A key determinant of endocrine development and function. Endocr. Rev. 1997, 18, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.L. Steroidogenic Factor 1: An Essential Mediator of Endocrine Development. Recent Prog. Horm. Res. 2002, 57, 19–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schimmer, B.P.; White, P.C. Minireview: Steroidogenic factor 1: Its roles in differentiation, development, and disease. Mol. Endocrinol. 2010, 24, 1322–1337. [Google Scholar] [CrossRef] [Green Version]
- Pelusi, C.; Ikeda, Y.; Zubair, M.; Parker, K.L. Impaired follicle development and infertility in female mice lacking steroidogenic factor 1 in ovarian granulosa cells. Boil. Reprod. 2008, 79, 1074–1083. [Google Scholar] [CrossRef]
- Lai, W.; Yeh, Y.; Fang, W.; Wu, L.; Harada, N.; Wang, P.; Ke, F.; Lee, W.; Hwang, J. Calcineurin and crtc2 mediate fsh and tgfβ1 upregulation of cyp19a1 and nr5a in ovary granulosa cells. J. Mol. Endocrinol. 2014, 53, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Derebecka-Holysz, N.; Lehmann, T.P.; Holysz, M.; Trzeciak, W.H. SMAD3 inhibits SF-1-dependent activation of the CYP17 promoter in H295R cells. Mol. Cell. Biochem. 2007, 307, 65–71. [Google Scholar] [CrossRef]
- Ning, L.; Yinglei, X.; Yimeng, Y.; Guidong, Y.; Hui, T.; Guishuan, W.; Jie, L.; Yong, W.; Fei, S. Steroidogenic factor-1 is required for tgf-beta3-mediated 17beta-estradiol synthesis in mouse ovarian granulosa cells. Endocrinology 2011, 152, 3213–3225. [Google Scholar]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Q.; Jin, M.; Wang, Y.; Liu, J.; Kalds, P.; Wang, Y.; Yang, Y.; Wang, X.; Chen, Y. Transactivation of miR-202-5p by Steroidogenic Factor 1 (SF1) Induces Apoptosis in Goat Granulosa Cells by Targeting TGFβR2. Cells 2020, 9, 445. https://doi.org/10.3390/cells9020445
Ding Q, Jin M, Wang Y, Liu J, Kalds P, Wang Y, Yang Y, Wang X, Chen Y. Transactivation of miR-202-5p by Steroidogenic Factor 1 (SF1) Induces Apoptosis in Goat Granulosa Cells by Targeting TGFβR2. Cells. 2020; 9(2):445. https://doi.org/10.3390/cells9020445
Chicago/Turabian StyleDing, Qiang, Miaohan Jin, Yaoyue Wang, Jiao Liu, Peter Kalds, Ying Wang, Yuxin Yang, Xiaolong Wang, and Yulin Chen. 2020. "Transactivation of miR-202-5p by Steroidogenic Factor 1 (SF1) Induces Apoptosis in Goat Granulosa Cells by Targeting TGFβR2" Cells 9, no. 2: 445. https://doi.org/10.3390/cells9020445