
CCL22-Polarized TAMs to M2a Macrophages in Cervical Cancer In Vitro Model

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RT-PCR
2.3. Flowcytometry
2.4. RNA Interference and Plasmid Construction
2.5. Statistics
3. Results
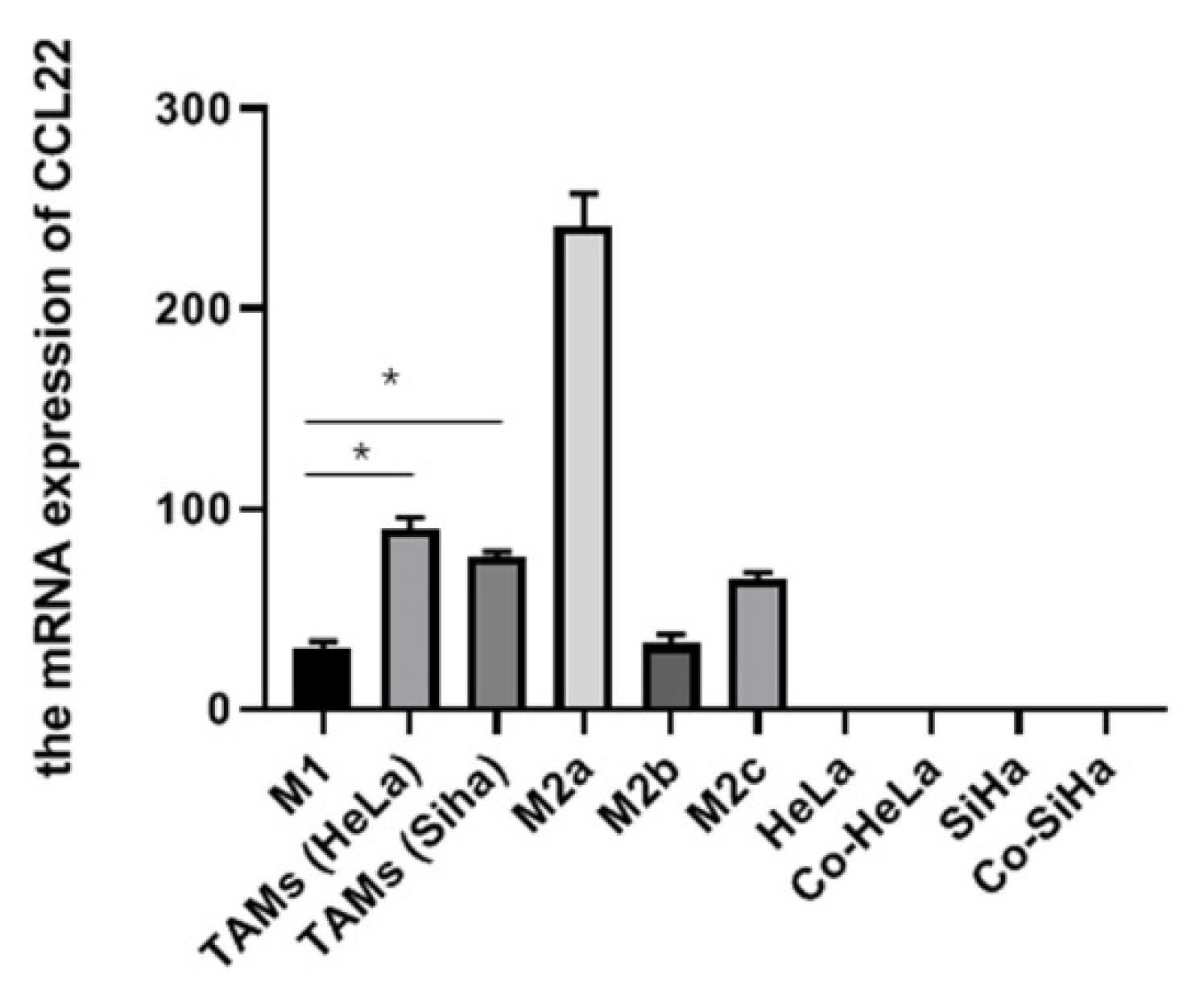
3.1. CCL22 Expression in TAMs of Cervical Cancer In Vitro
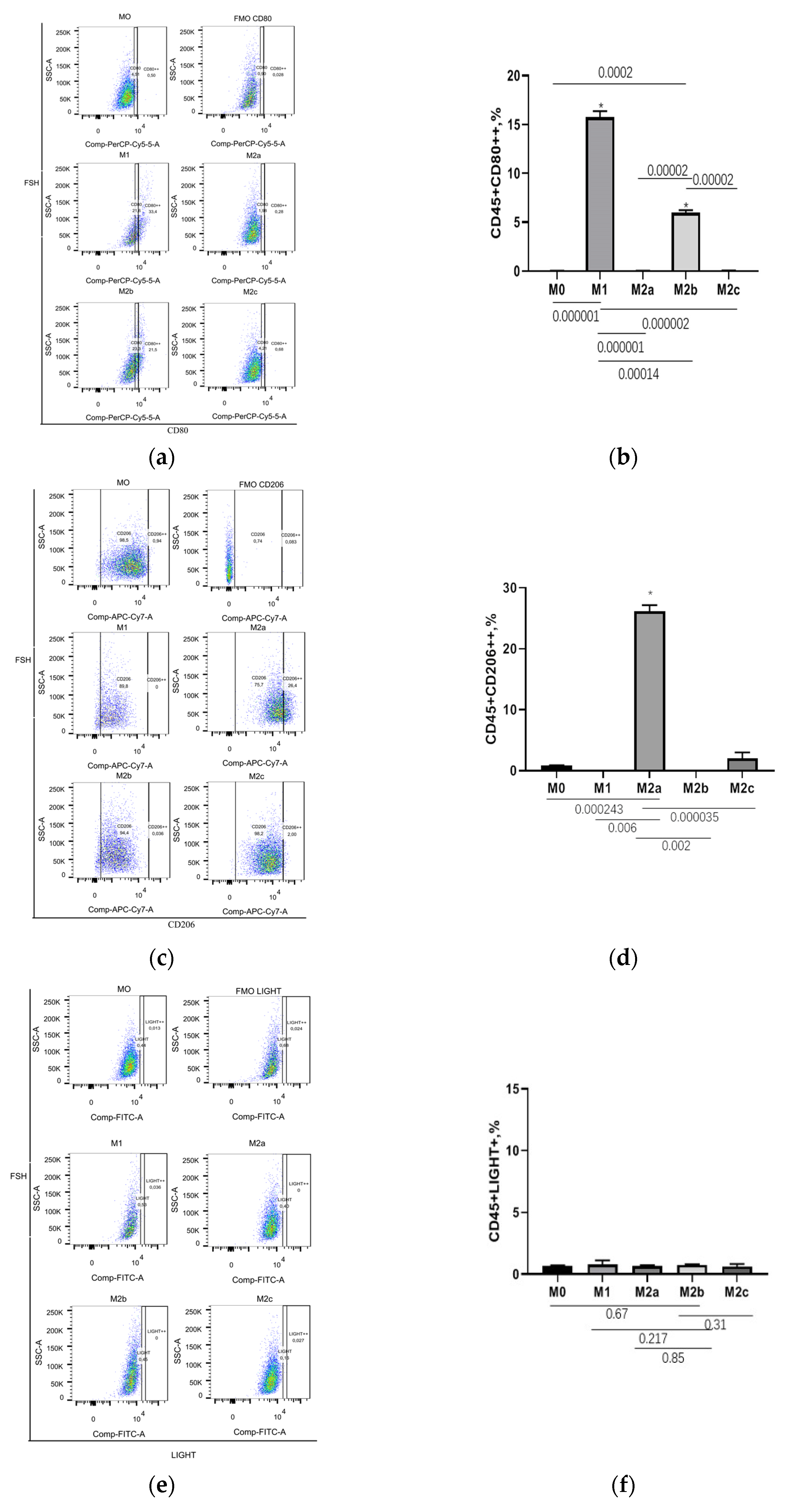
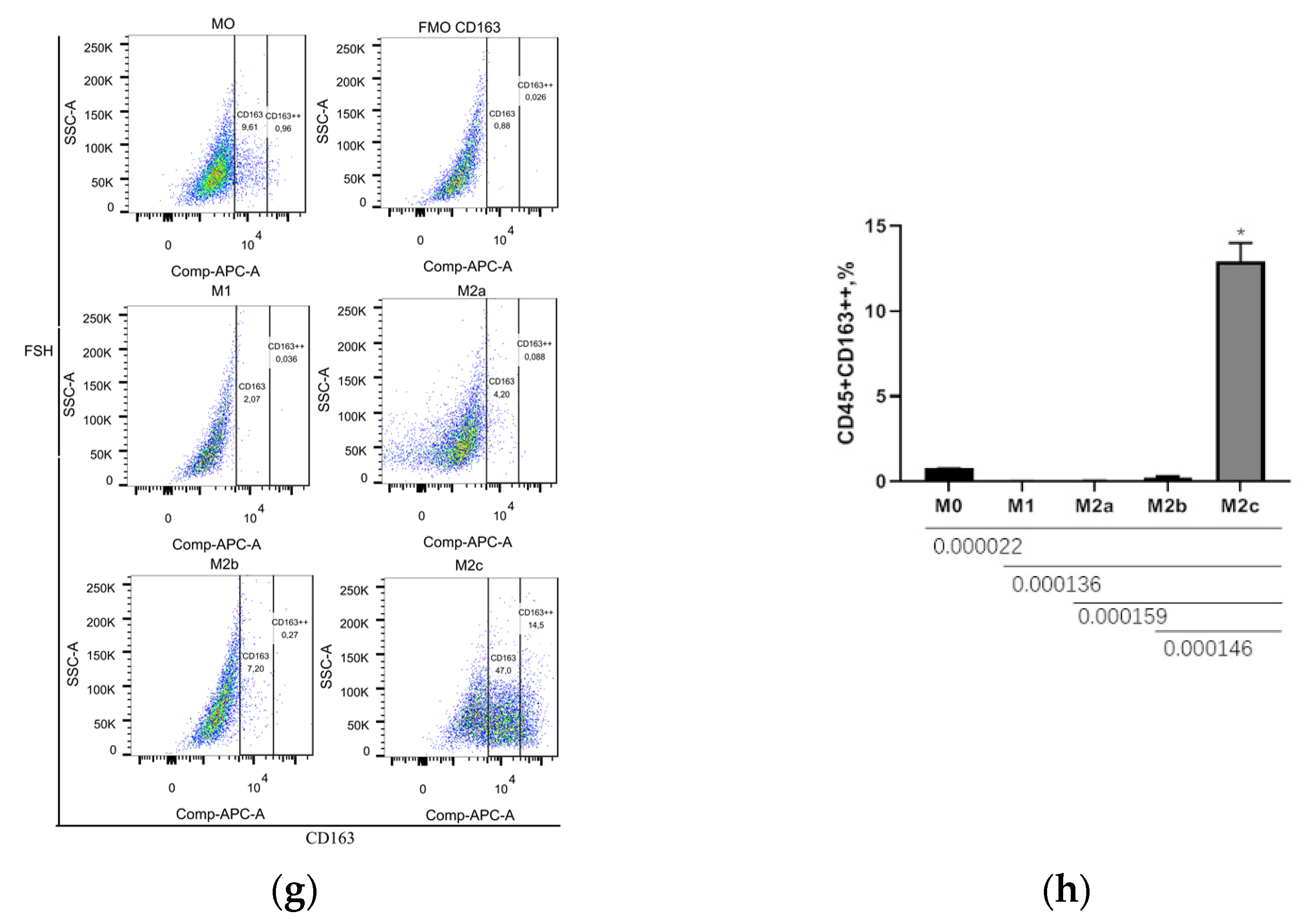
3.2. CD206++ and CD163++ Are the Marker of M2a and M2c Macrophages, Respectively
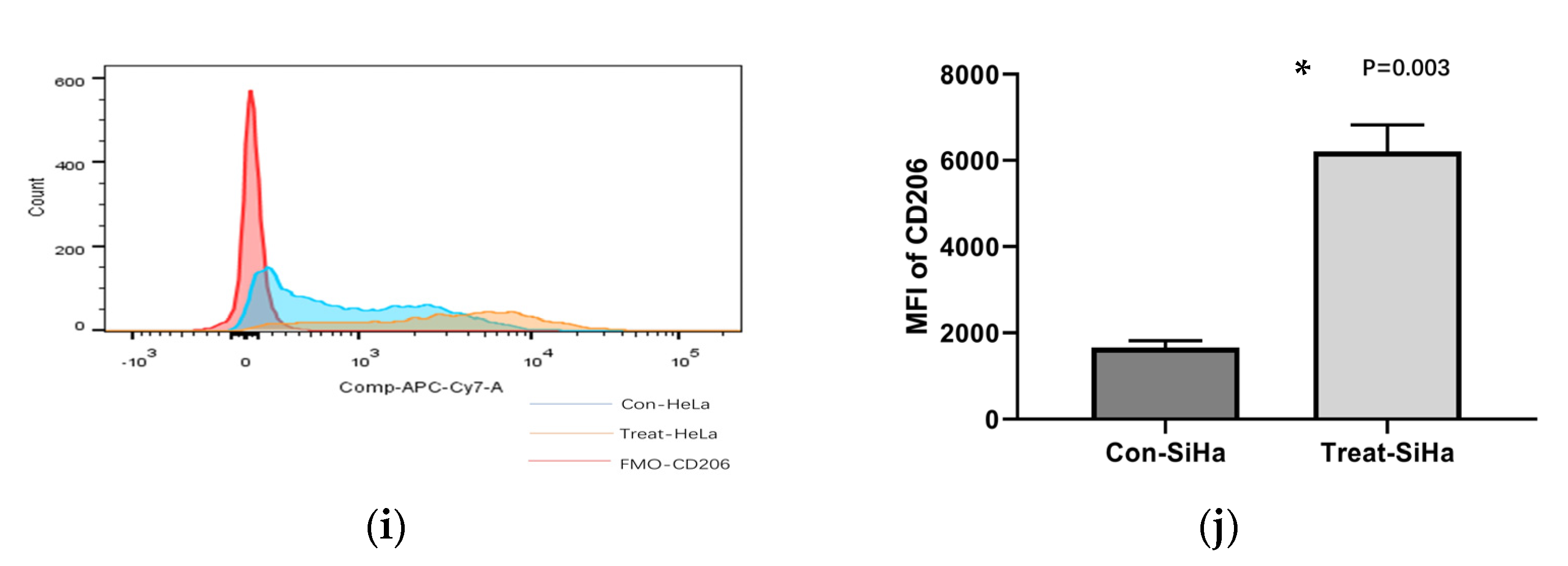
3.3. CCL22 Could Polarize TAMs toward M2a Macrophages in Cervical Cancer via an Autocrine Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Hirchaud, F.; Hermetet, F.; Ablise, M.; Fauconnet, S.; Vuitton, D.A.; Prétet, J.-L.; Mougin, C. Isoliquiritigenin induces caspase-dependent apoptosis via downregulation of HPV16 E6 expression in cervical cancer Ca Ski Cells. Planta Medica 2013, 79, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Small, W., Jr.; Bacon, M.A.; Bajaj, A.; Chuang, L.T.; Fisher, B.J.; Harkenrider, M.M.; Jhingran, A.; Kitchener, H.C.; Mileshkin, L.R.; Viswanathan, A.N.; et al. Cervical cancer: A global health crisis. Cancer 2017, 123, 2404–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.; Ye, M.; Zhou, J.; Wang, Z.P.; Zhu, X. Recent advances on the molecular mechanism of cervical carcinogenesis based on systems biology technologies. comput. Struct. Biotechnol. J. 2019, 17, 241–250. [Google Scholar] [CrossRef]
- Schmid, M.; Franckena, M.; Kirchheiner, K.; Sturdza, A.; Georg, P.; Dörr, W.; Pötter, R. Distant metastasis in patients with cervical cancer after primary radiotherapy with or without chemotherapy and image guided adaptive brachytherapy. Gynecol. Oncol. 2014, 133, 256–262. [Google Scholar] [CrossRef]
- Opinion Granted Based on Positive Results from Phase 3 KEYNOTE-826 Trial. Available online: https://www.merck.com/news/merck-receives-positive-eu-chmp-opinion-for-keytruda-pembrolizumab-plus-chemotherapy-with-or-without-bevacizumab-as-treatment-for-patients-with-persistent-recurrent-or-metastatic-cervical-c/. (accessed on 25 March 2022).
- Maciag, P.C.; Radulovic, S.; Rothman, J. The first clinical use of a live-attenuated Listeria monocytogenes vaccine: A phase I safety study of Lm-LLO-E7 in patients with advanced carcinoma of the cervix. Vaccine 2009, 27, 3975–3983. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Locati, M. New vistas on macrophage differentiation and activation. Eur. J. Immunol. 2007, 37, 14–16. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Locati, M. Macrophage polarization comes of age. Immunity 2005, 23, 344–346. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Xu, J.; Warren, C.M.; Duan, D.; Li, X.; Wu, L.; Iruela-Arispe, M.L. Endothelial cells provide an instructive niche for the differentiation and functional polarization of M2-like macrophages. Blood 2012, 120, 3152–3162. [Google Scholar] [CrossRef] [Green Version]
- Georgoudaki, A.-M.; Prokopec, K.E.; Boura, V.F.; Hellqvist, E.; Sohn, S.; Östling, J.; Dahan, R.; Harris, R.A.; Rantalainen, M.; Klevebring, D.; et al. Reprogramming tumor-associated macrophages by antibody targeting inhibits cancer progression and metastasis. Cell Rep. 2016, 15, 2000–2011. [Google Scholar] [CrossRef] [Green Version]
- Mosser, D.M. The many faces of macrophage activation. J. Leukoc. Biol. 2003, 73, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.; Keshav, S.; Harris, N.; Gordon, S. Interleukin 4 potently enhances murine macrophage mannose receptor activity: A marker of alternative immunologic macrophage activation. J. Exp. Med. 1992, 176, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.F.; Mosser, D.M. A novel phenotype for an activated macro-phage: The type 2 activated macrophage. J. Leukoc. Biol. 2002, 72, 101–106. [Google Scholar] [PubMed]
- Hong, S.; Qian, J.; Yang, J.; Li, H.; Kwak, L.W.; Yi, Q. Roles of idiotype-specific T cells in myeloma cell growth and survival: Th1 and CTL Cells are tumoricidal while Th2 cells promote tumor growth. Cancer Res. 2008, 68, 8456–8464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Fujimoto, S. A tumor-specific Th2 clone initiating tumor rejection via primed CD8+ cytotoxic T-lymphocyte activation in mice. Cancer Res. 1996, 56, 5005–5011. [Google Scholar]
- Mattes, J.; Hulett, M.; Xie, W.; Hogan, S.; Rothenberg, M.E.; Foster, P.; Parish, C. Immunotherapy of cytotoxic T cell-resistant tumors by T helper 2 cells: An eotaxin and STAT6-dependent process. J. Exp. Med. 2003, 197, 387–393. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Little, A.C.; Pathanjeli, P.; Wu, Z.; Bao, L.; Goo, L.E.; Yates, J.; Oliver, C.R.; Soellner, M.B.; Merajver, S.D. IL-4/IL-13 Stimulated macrophages enhance breast cancer invasion via Rho-GTPase regulation of synergistic VEGF/CCL-18 signaling. Front. Oncol. 2019, 9, 456. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.-M.; Xiao, X.; Lao, X.-M.; Wei, Y.; Liu, R.-X.; Zeng, Q.-H.; Wang, J.-C.; Ouyang, F.-Z.; Chen, D.-P.; Chan, K.-W.; et al. Polarization of tissue-resident TFH-Like cells in human hepatoma bridges innate monocyte inflammation and M2b macrophage polarization. Cancer Discov. 2016, 6, 1182–1195. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Koh, J.; Ko, J.S.; Kim, H.Y.; Lee, H.; Chung, D.H. Ubiquitin E3 ligase pellino-1 inhibits IL-10-mediated M2c polarization of macrophages, thereby suppressing tumor growth. Immune Netw. 2019, 19, e32. [Google Scholar] [CrossRef]
- Pathria, P.; Louis, T.L.; Varner, J.A. Targeting tumor-associated macrophages in cancer. Trends Immunol. 2019, 40, 310–327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhu, X.-D.; Sun, H.-C.; Xiong, Y.-Q.; Zhuang, P.-Y.; Xu, H.-X.; Kong, L.-Q.; Wang, L.; Wu, W.-Z.; Tang, Z.-Y. Depletion of tumor-associated macrophages enhances the effect of sorafenib in metastatic liver cancer models by antimetastatic and antiangiogenic effects. Clin. Cancer Res. 2010, 16, 3420–3430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchini, G.; Gianni, L. The immune system and response to HER2-targeted treatment in breast cancer. Lancet Oncol. 2014, 15, e58–e68. [Google Scholar] [CrossRef]
- Godiska, R.; Chantry, D.; Raport, C.J.; Sozzani, S.; Allavena, P.; Leviten, D.; Mantovani, A.; Gray, P.W. Human macrophage–derived chemokine (MDC), a novel chemoattractant for monocytes, monocyte-derived dendritic cells, and natural killer cells. J. Exp. Med. 1997, 185, 1595–1604. [Google Scholar] [CrossRef] [Green Version]
- Chang, M.; McNinch, J.; Elias, C., 3rd; Manthey, C.L.; Grosshans, D.; Meng, T.; Boone, T.; Andrew, D.P. Molecular Cloning and functional characterization of a novel CC chemokine, stimulated T cell chemotactic protein (STCP-1) that specifically acts on activated T lymphocytes. J. Biol. Chem. 1997, 272, 25229–25237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaniel, C.; Pardali, E.; Sallusto, F.; Speletas, M.; Ruedl, C.; Shimizu, T.; Seidl, T.; Andersson, J.; Melchers, F.; Rolink, A.G.; et al. Activated murine B lymphocytes and dendritic cells produce a novel CC chemokine which acts selectively on activated T Cells. J. Exp. Med. 1998, 188, 451–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Gray, P.A.; Van Damme, J. Macrophage-derived chemokine (MDC). J. Leukoc. Biol. 2000, 68, 400–404. [Google Scholar]
- Curiel, T.J.; Coukos, G.; Zou, L.; Alvarez, X.; Cheng, P.; Mottram, P.; Evdemon-Hogan, M.; Conejo-Garcia, J.R.; Zhang, L.; Burow, M.; et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat. Med. 2004, 10, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Yang, C.; Wang, S.; Shi, D.; Zhang, C.; Lin, X.; Xiong, B. M2 macrophages confer resistance to 5-fluorouracil in colorectal cancer through the activation of CCL22/PI3K/AKT signaling. OncoTargets Ther. 2019, 12, 3051–3063. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Li, Y.; Wei, X.; Zhang, Q.; Jia, H.; Quan, S.; Cao, D.; Wang, L.; Yang, T.; Zhao, J.; et al. Negative immune factors might predominate local tumor immune status and promote carcinogenesis in cervical carcinoma. Virol. J. 2017, 14, 5. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Schmoeckel, E.; Kost, B.P.; Kuhn, C.; Vattai, A.; Vilsmaier, T.; Mahner, S.; Mayr, D.; Jeschke, U.; Heidegger, H.H. Higher CCL22+ Cell Infiltration is associated with poor prognosis in cervical cancer patients. Cancers 2019, 11, 2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jaguin, M.; Houlbert, N.; Fardel, O.; Lecureur, V. Polarization profiles of human M-CSF-generated macrophages and comparison of M1-markers in classically activated macrophages from GM-CSF and M-CSF origin. Cell. Immunol. 2013, 281, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.X.; Zhang, S.X.; Wu, H.J.; Rong, X.L.; Guo, J. M2b macrophage polarization and its roles in diseases. J. Leukoc. Biol. 2019, 106, 345–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeyda, M.; Farmer, D.; Todoric, J.; Aszmann, O.; Speiser, M.; Györi, G.; Zlabinger, G.; Stulnig, T. Human adipose tissue macrophages are of an anti-inflammatory phenotype but capable of excessive pro-inflammatory mediator production. Int. J. Obes. 2007, 31, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colin, S.; Chinetti-Gbaguidi, G.; Staels, B. Macrophage phenotypes in atherosclerosis. Immunol. Rev. 2014, 262, 153–166. [Google Scholar] [CrossRef]
- De Vos van Steenwijk, P.J.; Ramwadhdoebe, T.; Goedemans, R.; Doorduijn, E.; van Ham, J.; Gorter, A.; van Hall, T.; Kuijjer, M.; van Poelgeest, M.; van der Burg, S.; et al. Tumor-infiltrating CD14-positive myeloid cells and CD8-positive T-cells prolong survival in patients with cervical carcinoma. Int. J. Cancer 2013, 133, 2884–2894. [Google Scholar] [CrossRef]
- Pedraza-Brindis, E.J.; Sánchez-Reyes, K.; Hernández-Flores, G.; Bravo-Cuellar, A.; Jave-Suárez, L.F.; Aguilar-Lemarroy, A.; Gómez-Lomelí, P.; López-López, B.A.; Ortiz-Lazareno, P.C. Culture supernatants of cervical cancer cells induce an M2 phenotypic profile in THP-1 macrophages. Cell. Immunol. 2016, 310, 42–52. [Google Scholar] [CrossRef]
- Li, L.; Yu, S.; Zang, C. Low Necroptosis Process predicts poor treatment outcome of human papillomavirus positive cervical cancers by decreasing tumor-associated macrophages M1 polarization. Gynecol. Obstet. Investig. 2018, 83, 259–267. [Google Scholar] [CrossRef]
- Schildberger, A.; Rossmanith, E.; Eichhorn, T.; Strassl, K.; Weber, V. Monocytes, peripheral blood mononuclear cells, and THP-1 cells exhibit different cytokine expression patterns following stimulation with lipopolysaccharide. Mediat. Inflamm. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bruckmeier, M.; Kuehnl, A.; Culmes, M.; Pelisek, J.; Eckstein, H.-H. Impact of oxLDL and LPS on C-type Natriuretic Peptide System is Different between THP-1 Cells and Human Peripheral Blood Monocytic Cells. Cell. Physiol. Biochem. 2012, 30, 199–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hijiya, N.; Miyake, K.; Akashi, S.; Matsuura, K.; Higuchi, Y.; Yamamoto, S. Possible involvement of toll-like receptor 4 in endothelial cell activation of larger vessels in response to lipopolysaccharide. Pathobiology 2002, 70, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z. The use of THP-1 cells as a model for mimicking the function and regulation of monocytes and macrophages in the vasculature. Atherosclerosis 2012, 221, 2–11. [Google Scholar] [CrossRef]
- Aldo, P.B.; Craveiro, V.; Guller, S.; Mor, G. Effect of culture conditions on the phenotype of THP-1 monocyte cell Line. Am. J. Reprod. Immunol. 2013, 70, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Tarique, A.; Logan, J.; Thomas, E.; Holt, P.; Sly, P.D.; Fantino, E. Phenotypic, functional, and plasticity features of classical and alternatively activated human macrophages. Am. J. Respir. Cell Mol. Biol. 2015, 53, 676–688. [Google Scholar] [CrossRef] [PubMed]
- Saghaeian-Jazi, M.; Mohammadi, S.; Sedighi, S. Culture and differentiation of monocyte derived macrophages using human serum: An optimized method. Zahedan J. Res. Med Sci. 2016, 18, e7362. [Google Scholar] [CrossRef] [Green Version]
- Zizzo, G.; Cohen, P.L. The PPAR-γ antagonist GW9662 elicits differentiation of M2c-like cells and upregulation of the MerTK/Gas6 axis: A key role for PPAR-γ in human macrophage polarization. J. Inflamm. 2015, 12, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Spaccamela, V.M.; Valencia, R.G.; Pastukhov, O.; Duppenthaler, A.; Dettmer, M.S.; Erb, J.; Steiner, U.C.; Hillinger, S.; Speckmann, C.; Ehl, S.; et al. High levels of IL-18 and IFN-γ in Chronically inflamed tissue in chronic granulomatous disease. Front. Immunol. 2019, 10, 2236. [Google Scholar] [CrossRef]
- Iqbal, S.; Kumar, A. Characterization of In vitro Generated Human Polarized Macrophages. J. Clin. Cell. Immunol. 2015, 6, 10–4172. [Google Scholar] [CrossRef] [Green Version]
- Gotovina, J.; Bianchini, R.; Fazekas-Singer, J.; Herrmann, I.; Pellizzari, G.; Haidl, I.D.; Hufnagl, K.; Karagiannis, S.N.; Marshall, J.S.; Jensen-Jarolim, E. Epinephrine drives human M2a allergic macrophages to a regulatory phenotype reducing mast cell degranulation in vitro. Allergy 2020, 75, 2939–2942. [Google Scholar] [CrossRef]
- Raggi, F.; Pelassa, S.; Pierobon, D.; Penco, F.; Gattorno, M.; Novelli, F.; Eva, A.; Varesio, L.; Giovarelli, M.; Bosco, M.C. Regulation of human macrophage M1–M2 polarization balance by hypoxia and the triggering receptor expressed on myeloid Cells-1. Front. Immunol. 2017, 8, 1097. [Google Scholar] [CrossRef] [PubMed]
- Faget, J.; Biota, C.; Bachelot, T.; Gobert, M.; Treilleux, I.; Goutagny, N.; Durand, I.; Léon-Goddard, S.; Blay, J.Y.; Caux, C.; et al. Early Detection of Tumor Cells by Innate Immune Cells Leads to Treg Recruitment through CCL22 Production by Tumor Cells. Cancer Res. 2011, 71, 6143–6152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobert, M.; Treilleux, I.; Bendriss-Vermare, N.; Bachelot, T.; Goddard-Leon, S.; Arfi, V.; Biota, C.; Doffin, A.C.; Durand, I.; Olive, D.; et al. Regulatory T cells recruited through CCL22/CCR4 are selectively activated in lymphoid infiltrates surrounding primary breast tumors and lead to an adverse clinical outcome. Cancer Res. 2009, 69, 2000–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anz, D.; Rapp, M.; Eiber, S.; Koelzer, V.H.; Thaler, R.; Haubner, S.; Knott, M.; Nagel, S.; Golic, M.; Wiedemann, G.M.; et al. Suppression of intratumoral CCL22 by Type I interferon inhibits migration of regulatory T cells and blocks cancer progression. Cancer Res. 2015, 75, 4483–4493. [Google Scholar] [CrossRef] [Green Version]
- Wiedemann, G.M.; Knott, M.M.; Vetter, V.K.; Rapp, M.; Haubner, S.; Fesseler, J.; Kühnemuth, B.; Layritz, P.; Thaler, R.; Kruger, S.; et al. Cancer cell-derived IL-1alpha induces CCL22 and the recruitment of regulatory T cells. Oncoimmunology 2016, 5, e1175794. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Bracci, P.M.; McCoy, L.S.; Hsuang, G.; Wiemels, J.L.; Rice, T.; Zheng, S.; Kelsey, K.T.; Wrensch, M.R.; Wiencke, J.K. Serum macrophage-derived chemokine/CCL22 levels are associated with glioma risk, CD4 T cell lymphopenia and survival time. Int. J. Cancer 2015, 137, 826–836. [Google Scholar] [CrossRef]
- Kimura, S.; Nanbu, U.; Noguchi, H.; Harada, Y.; Kumamoto, K.; Sasaguri, Y.; Nakayama, T. Macrophage CCL22 expression in the tumor microenvironment and implications for survival in patients with squamous cell carcinoma of the tongue. J. Oral Pathol. Med. 2019, 48, 677–685. [Google Scholar] [CrossRef]
- Muthuswamy, R.; Urban, J.; Lee, J.-J.; Reinhart, T.A.; Bartlett, D.; Kalinski, P. Ability of mature dendritic cells to interact with regulatory T cells is imprinted during maturation. Cancer Res. 2008, 68, 5972–5978. [Google Scholar] [CrossRef] [Green Version]
- Jetten, N.; Verbruggen, S.; Gijbels, M.J.; Post, M.J.; De Winther, M.P.J.; Donners, M.M.P.C. Anti-inflammatory M2, but not pro-inflammatory M1 macrophages promote angiogenesis in vivo. Angiogenesis 2014, 17, 109–118. [Google Scholar] [CrossRef]
- Shan, K.; Feng, N.; Cui, J.; Wang, S.; Qu, H.; Fu, G.; Li, J.; Chen, H.; Wang, X.; Wang, R.; et al. Resolvin D1 and D2 inhibit tumour growth and inflammation via modulating macrophage polarization. J. Cell. Mol. Med. 2020, 24, 8045–8056. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Populations | M0 | M1 | M2a | M2b | M2c |
|---|---|---|---|---|---|
| CD45+CD80++ | 0.03 ± 0.01 | 15.76 ± 0.61 | 0.04 ± 0.01 | 5.99 ± 0.26 | 0.06 ± 0.01 |
| CD45+LIGHT+ | 0.68 ± 0.04 | 0.99 ± 0.23 | 0.67 ± 0.06 | 0.76 ± 0.04 | 0.62 ± 0.21 |
| CD45+CD206++ | 0.85 ± 0.08 | 0.00 ± 0.00 | 26.13 ± 1.03 | 0.03 ± 0.01 | 2.00 ± 1.00 |
| CD45+CD163++ | 0.77 ± 0.06 | 0.04 ± 0.01 | 0.53 ± 0.04 | 0.25 ± 0.05 | 12.63 ± 1.52 |
| MFI | FMO | con-HeLa | si-HeLa | con-Siha | si-Siha |
|---|---|---|---|---|---|
| CD206 | 164 ± 23 | 6741 ± 213 | 3921 ± 796 | 3862 ± 116 | 2885 ± 491 |
| MFI | FMO | C-HeLa | T-HeLa | C-Siha | T-Siha |
|---|---|---|---|---|---|
| CD206 | 164 ± 23 | 1736 ± 97 | 5846 ± 196 | 1658 ± 159 | 6208 ± 617 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Sudan, K.; Schmoeckel, E.; Kost, B.P.; Kuhn, C.; Vattai, A.; Vilsmaier, T.; Mahner, S.; Jeschke, U.; Heidegger, H.H. CCL22-Polarized TAMs to M2a Macrophages in Cervical Cancer In Vitro Model. Cells 2022, 11, 2027. https://doi.org/10.3390/cells11132027
Wang Q, Sudan K, Schmoeckel E, Kost BP, Kuhn C, Vattai A, Vilsmaier T, Mahner S, Jeschke U, Heidegger HH. CCL22-Polarized TAMs to M2a Macrophages in Cervical Cancer In Vitro Model. Cells. 2022; 11(13):2027. https://doi.org/10.3390/cells11132027
Chicago/Turabian StyleWang, Qun, Kritika Sudan, Elisa Schmoeckel, Bernd Peter Kost, Christina Kuhn, Aurelia Vattai, Theresa Vilsmaier, Sven Mahner, Udo Jeschke, and Helene Hildegard Heidegger. 2022. "CCL22-Polarized TAMs to M2a Macrophages in Cervical Cancer In Vitro Model" Cells 11, no. 13: 2027. https://doi.org/10.3390/cells11132027