
Homeostatic Regulation of Glucocorticoid Receptor Activity by Hypoxia-Inducible Factor 1: From Physiology to Clinic


1
Department of Biology and Biochemistry, University of Bath, Claverton Down, Bath BA2 7AY, UK
2
The Bateson Centre & The School of Biosciences, Firth Court, University of Sheffield, Western Bank, Sheffield S10 2TN, UK
*
Author to whom correspondence should be addressed.
Cells 2021, 10(12), 3441; https://doi.org/10.3390/cells10123441
Submission received: 12 November 2021
/
Revised: 2 December 2021
/
Accepted: 4 December 2021
/
Published: 7 December 2021
(This article belongs to the Special Issue Immune Modulations by Glucocorticoids: From Molecular Biology to Clinical Research)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Glucocorticoids (GCs) represent a well-known class of lipophilic steroid hormones biosynthesised, with a circadian rhythm, by the adrenal glands in humans and by the inter-renal tissue in teleost fish (e.g., zebrafish). GCs play a key role in the regulation of numerous physiological processes, including inflammation, glucose, lipid, protein metabolism and stress response. This is achieved through binding to their cognate receptor, GR, which functions as a ligand-activated transcription factor. Due to their potent anti-inflammatory and immune-suppressive action, synthetic GCs are broadly used for treating pathological disorders that are very often linked to hypoxia (e.g., rheumatoid arthritis, inflammatory, allergic, infectious, and autoimmune diseases, among others) as well as to prevent graft rejections and against immune system malignancies. However, due to the presence of adverse effects and GC resistance their therapeutic benefits are limited in patients chronically treated with steroids. For this reason, understanding how to fine-tune GR activity is crucial in the search for novel therapeutic strategies aimed at reducing GC-related side effects and effectively restoring homeostasis. Recent research has uncovered novel mechanisms that inhibit GR function, thereby causing glucocorticoid resistance, and has produced some surprising new findings. In this review we analyse these mechanisms and focus on the crosstalk between GR and HIF signalling. Indeed, its comprehension may provide new routes to develop novel therapeutic targets for effectively treating immune and inflammatory response and to simultaneously facilitate the development of innovative GCs with a better benefits-risk ratio.
1. Introduction
The name “glucocorticoid” (GC) is a portmanteau word (glucose + cortex + steroid), which derives from their key role in the regulation of glucose metabolism, their biosynthesis at the level of the adrenal cortex and their steroidal structure. They represent a well-known class of lipophilic steroid hormones synthetized, with a circadian rhythm, by the adrenal glands in humans and by the inter-renal tissue in teleost fish. GC circadian production in mammals is tuned by the hypothalamus-pituitary-adrenal (HPA) axis, which is the equivalent of the hypothalamus-pituitary-inter-renal (HPI) axis in teleost fish. Both are essential for stress adaptation [1,2,3,4]. The axis consists of a highly conserved regulatory system present in all living organisms aimed at maintaining a dynamic equilibrium in the body in response to external and internal stimuli, which is fundamental to assure homeostasis and survival. Cortisol, the end-product of the HPA/I axis, is the main GC both in humans and teleost fish and plays a fundamental role in the maintenance of both resting and stress-related responses [5,6,7,8].
Since their discovery in the 1940s [9], much has been learnt of GC molecular modes of action [10,11,12,13,14,15,16,17,18,19,20,21,22,23]. In particular, the glucocorticoid receptor’s characterization as a DNA-binding protein that regulates transcription initiation [24], the cloning of GR [25,26] and the breakthrough that most of the immunosuppressive actions of GCs occur via interfering with key inflammatory transcriptional regulators such as NF-κB and AP-1 [27,28,29,30], represent the main milestones.
Natural and synthetic glucocorticoids have been widely used for decades as effective anti-inflammatory and immunosuppressive treatments to control pathological disorders, which are very often linked to hypoxia. In particular, they have been broadly used to treat both acute and chronic inflammations, including inflammatory bowel disease, rheumatoid arthritis, multiple sclerosis, eczema and psoriasis, as well as being used in treatment of various leukaemias and in immunosuppressive regimes upon organ transplant [31,32,33,34,35,36]. At any point in time, an estimated ~1% of the total adult UK population receives oral glucocorticoid therapy [37]. However, due to the presence of adverse effects [38] and GC resistance [39,40,41,42], their therapeutic benefits are limited in patients chronically treated with these steroids. Examples of the most common GC-related side effects include osteoporosis, glaucoma, diabetes, skin atrophy, abdominal obesity, dyslipidemia, hypertension in adults and growth retardation in children [16,43,44].
Cortisol exerts its functions through direct binding to the glucocorticoid receptor (GR), but also to the mineralocorticoid receptor (MR), which binds cortisol with even higher affinity [45,46]. As transcription factors, both GR and MR compete for the same ligands, can form heterodimers and homodimers with each other, recognize and bind many of the same hormone response elements on the DNA, and share numerous coregulatory proteins involved in the gene transcription initiation. Importantly, GCs activate MR in most tissues at basal levels, whereas activate GR under stressful conditions or at the diurnal peak [47].
Once bound together, they form an active complex which can function in the nucleus to modulate the transcription of effector proteins, as well as in the cytoplasm to hamper targeted transcription factors activity. Historically, these functions have been coined genomic and nongenomic modes of action, respectively [48,49,50,51]. Importantly, GCs and their kindred intracellular receptors, represent critical checkpoints in the endocrine control of vertebrate energy homeostasis. Indeed, if HPA axis activity is not accurately regulated, GC imbalance may result in different pathological conditions such as hypertension, severe cardiovascular, immunological and metabolic complications (e.g., Addison’s disease (GC deficiency) and Cushing’s syndrome (GC excess)) [52,53,54]. In addition, alterations or flaws in the HPA axis response are tightly associated with a broad range of inflammatory and autoimmune diseases, both in humans and in animal models. The latter include Crohn’s disease, rheumatoid arthritis, colitis, inflammatory bowel disease, multiple sclerosis (whose animal equivalent is autoimmune encephalomyelitis), dermatitis, and asthma. Inflammatory conditions include fibromyalgia, chronic fatigue syndrome, depression, and post-traumatic stress disorder (PTSD) [55,56,57,58,59,60,61]. Moreover, even if the biological effects induced by GCs are usually adaptive, their abnormal activity may contribute to a series of acute metabolic diseases which include insulin resistance, obesity, and type 2 diabetes [62,63]. Thus, furthering the research on how GCs precisely work and interact with other pathways may provide better tools to treat these diseases and simultaneously allow the development of selective GR agonists and specific drug-targeting strategies.
Similarly to GCs that are involved in numerous homeostatic maintenance activities (e.g., metabolism of protein, carbohydrate and lipid, etc.) [64], the HIF signalling pathway exerts a pivotal role in ensuring homeostasis, the preservation of which is essential for the correct functioning of the cell. In this regard, the ability to perceive and quickly respond to changes related to environmental oxygen availability is controlled by the hypoxia-inducible factor transcription factors (HIF) family. Hypoxia is a common pathophysiological occurrence, with a profound impact both on human and animal physiology, in which oxygen availability to cells, tissues or to an organ is reduced below a certain threshold (O2 levels < 2%) [65,66]. HIF transcription factors are key homeostatic regulators which coordinate a metabolic shift from aerobic to anaerobic metabolism to assure cell survival, both in mammals and in zebrafish [67,68,69,70,71].
The HIF pathway is finely regulated by the PHD3-VHL-E3 ubiquitin ligase complex the aim of which is to maintain low basal HIF levels that can rapidly increase to promptly respond when oxygen levels decrease. This avoids any activation of the HIF pathway under normoxic conditions. As a transcription factor, HIF drives the hypoxic response via binding to specific hypoxia-response elements (HREs). These are involved in decreasing oxygen consumption and increasing oxygen and nutrient delivery [72,73,74]. Interestingly, HIF signalling can tune its own activation via negative feedback by inducing the expression of the oxygen sensors proteins (PHDs), in particular prolyl hydroxylase 3 (PHD3) in zebrafish and PHD2 in humans and mice [75,76].
However, although the HIF response is aimed at restoring tissue oxygenation and perfusion, it may sometimes be maladaptive and may contribute to the onset of different pathological conditions (e.g., inflammation, stroke, tissue ischemia and growth of solid tumours) [65]. Thus, both glucocorticoids and hypoxia-induced transcriptional responses have been shown to exert crucial roles in tissue homeostasis and in the regulation of cellular responses to stress and inflammation [77,78,79,80,81,82].
Recent studies have also strengthened the knowledge on the important crosstalk between these two major signalling pathways. To this purpose, the aim of the present review is to discuss the evidence accumulated to date about such crosstalk. We also describe novel mechanisms by which GR and HIF influence each other, both in vitro and in vivo, and how these could be exploited to develop novel therapeutic targets required to overcome GC-related and HIF-related diseases. To this end, the use of the zebrafish (Danio rerio) as an effective in vivo model organism has become more and more important to study how both hypoxic and glucocorticoid signalling function in vivo. Indeed, zebrafish share all the components of the human HIF and GC pathway, and the zebrafish has been demonstrated to be a very informative and genetically tractable organism for studying both hypoxia and the stress response both in physiological and pathophysiological conditions [76,83,84,85,86,87]. Finally, the ease use of medium to high throughput drug screening [88] and genetic tractability via the CRISPR/Cas9 based mutagenesis method [89,90,91] make the zebrafish particularly suitable not only for genetic dissection of pathways themselves, but also crosstalk between pathways studied at the level of whole organism.
2. Glucocorticoids
2.1. Biosynthesis, Secretion and Availability
GCs are essential steroid hormones biosynthesized and secreted by the adrenal cortex/inter-renal gland both in a circadian manner and in response to stress. The latter is generally defined as a status of real or perceived threat to homeostasis. Assuring homeostasis in the presence of stressors requires the activation of an intricate series of coordinated biological responses performed by the nervous, endocrine and immune systems [62,92]. The key anatomical structures that regulate the stress response are located both in the central nervous system and in peripheral tissues. The primary effectors of the stress response are localized in the paraventricular nucleus (PVN) of the hypothalamus, in the anterior lobe of the pituitary gland and at the level of the adrenal gland. These three main structures are generally referred to as the hypothalamic-pituitary-adrenal (HPA) axis in humans, and as the hypothalamic-pituitary-inter-renal axis (HPI) in zebrafish [62,92,93]. Among these, the hypothalamus is the initial stressor recognition site for both internal and external signals. In mammals, neurons localized in the paraventricular nucleus synthesize both corticotropin-releasing factor (CRF) and arginine vasopressin (AVP), which are released into hypophyseal portal vessels that access the anterior pituitary gland. On the other hand, in teleosts, there is a direct neuronal connection to endocrine cells through the hypophyseal stalk, since they lack a portal system between the hypothalamus and the pituitary gland [94]. Here, CRF binding to its receptor localized on pituitary corticotropes triggers the release of adrenocorticotropic hormone (ACTH) into the systemic circulation. In humans, ACTH derives by post translational modification of its precursor encoded by the proopiomelanocortin (POMC) gene. Of note, due to genome duplication, two pomc genes named pomca and pomcb have been identified in zebrafish [95,96,97]. However, only pomca seems to be expressed in the pituitary gland and is required for the inter-renal organ development [98,99]. The main target of ACTH is the adrenal cortex in humans and the inter-renal tissue in teleosts, where it binds to the melanocortin 2 receptor (MC2R) on the steroidogenic cells. Here, it stimulates cortisol biosynthesis and secretion starting from cholesterol [62,92,100,101].
Finally, once released into the systemic circulation, GCs can access target tissues (e.g., liver, heart, and vascular tissues) to exert metabolic and cardiovascular effects and the brain itself, in order to support cognitive processes required to tackle a threatening situation [102]. Under non stressful conditions, glucocorticoid levels in the serum are homeostatically controlled by the HPA monitoring activity, whereas glucocorticoid availability is further tuned at a tissue and cellular level. Circulating glucocorticoids are primarily bound to corticosteroid binding globulin (CBG) and just a small percentage (5–15%) is bound to albumin. As a result, the majority of GC is maintained in an inactive form, and only the remaining 5% of systemic GC is free and bioactive. Hence, CBG concentration constitutes a pivotal regulator of cortisol accessibility [103].
Importantly, GCs are also able to control their own biosynthesis and secretion by tuning the activity of the HPA/I axis itself. This is particularly important to stop the stress response and avoid an exacerbated reaction [104]. This is achieved via a GC-GR mediated negative feedback loop, which acts both at the hypothalamic and anterior pituitary levels, where GC-GR activity inhibits both CRH and POMC (ACTH precursor) biosynthesis and release [2,21,23,105]. This occurs via a mechanism that requires GC-GR binding to an nGRE within the pomca promoter [106]. For these reasons, pomca is a well-established and frequently used readout of GR activity. In addition, GCs may indirectly control the HPA axis activity through modulation of brain structures activity, such as the amygdala, the hippocampus and the prefrontal cortex, that can, in turn, influence the activity of the paraventricular nucleus [102,107,108,109].
2.2. The Glucocorticoid Receptor: Structure and Functions
GCs exert their systemic functions by binding to the glucocorticoid receptor (GR) and the mineralocorticoid receptor (MR). Due to their lipophilic nature, GCs can passively diffuse across the plasma membrane into the cytoplasm. Within the cells, their biological availability is then regulated by two enzymes of the 11β-Hydroxysteroid dehydrogenase (11β-HSD) family that work in an opposite fashion. 11β-HSD2 oxidizes cortisol into its inactive form cortisone, reducing GC availability. Vice versa, 11β-HSD1 transforms cortisone to cortisol, thereby increasing local GC activity. Inside the cell, GCs can bind to their specific receptors GR and MR [33,102,110,111]. Both receptors, in the absence of their ligands (unbound state) are associated in an inactive oligomeric complex with specific regulatory proteins. Among these, heat shock protein-90 kD (HSP90), which binds both GR and MR to the C-terminal domain, heat shock protein-70 kD (HSP70), p59 immunophilin, Fkbp51 and Fkbp52 and the small p23 phosphoprotein maintain correct protein folding of the receptor [102,112,113]. The GR, which belongs to the nuclear receptor transcription factor family, is composed of different conserved structural domains [114]. These include an N-terminal variable region (NTD) required for ligand-independent gene transactivation, which contains a transactivation domain named activation function 1 (AF1). The latter is responsible for the transcriptional activation and is involved in the association with coregulators and the basal transcription machinery. A central DNA-binding domain composed of two zinc fingers has been shown to be crucial both for GR homodimerization and DNA-binding specificity. This is followed by an adjacent flexible hinge region allowing proper DNA binding, dimerization, and nuclear translocation of the receptor [115]. Finally, the C-terminal region (LBD) contains the ligand binding domain and a secondary transactivation domain (AF2), regulated by hormone binding, which is essential for dimerization, interaction with cochaperones, coregulators, and other transcription factors [116,117]. The LBD also comprises a dimer interface which is fundamental for GR function and the binding of the heat shock protein 90 (Hsp 90) [118]. Both DBD and LBD include nuclear localization signals, which are required for GR nuclear translocation. Finally, DBD also incorporate the nuclear export signal sequence (NES) which targets it for export from the nucleus to the cytoplasm via the nuclear pore complex [113] (Figure 1).
Although the NTD is conserved, literature reviews and sequence alignments of human, monkey, rat, and mouse GRs have revealed that there are another eight conserved AUG start codons in the exon 2 (Figure 1). In humans, these were shown to produce various GR isoforms with progressively shorter N-terminal transactivation domains [113]. These are formed due to the presence of alternative Kozak translation initiation sequences which can cause either ribosomal shunting or ribosomal leaky scanning mechanisms. This allows the generation of different GR subtypes with truncated N-termini [119,120,121,122,123], which are likely to be fully active. This is consistent with data from zebrafish, where a GR mutant line (grsh551), characterized by a 1 bp deletion in the first coding exon (exon 2, Q48fsX3), proved not to have any detectable phenotype [124]. This was confirmed further by ISH analysis, which showed that both grsh551 mutants and wildtypes displayed an identical downregulated pomca expression after synthetic GC (Betamethasone 17,21-dipropionate) administration.
Synthetic GR agonists are supposed to trigger a potent GC response, which in turn elicits the GC-GR mediated negative feedback loop, aimed to shut down their own biosynthesis. As previously mentioned, this mainly occurs at the level of the pituitary gland via downregulation of pomca [2,106,111]. For this reason, if GR is not functional, the GC-GR negative feedback loop cannot occur and pomca expression should not be downregulated, as occurs in grsh551 mutants. Indeed, in grsh551 mutants the feedback occurs normally.
In addition to alternative starts, alternative splicing at exon 9 is responsible for generating two different GR splice variants, namely GRα (777 aa) and GRβ (742 aa) [25,125]. These two receptor isoforms share an identical amino acid sequence between 1–727 aa and then diverge. In particular, the human (h) GRα c C-terminal region contains 50 distinct amino acid residues that form two alpha-helical structures that play a key ligand binding role. In contrast, the hGRβ C-terminal is characterized by a shortened 15 non-homologous, specific amino acid sequence that prevents GC binding [20,126,127]. Hence, hGRβ does not bind traditional glucocorticoid agonists and lacks transactivational activity on GRE-containing promoters, whereas hGRα is the canonical GR isoform. Nevertheless, hGRβ is constitutively present in the nucleus where it has been shown to act as a dominant-negative inhibitor of hGRα’s transactivational properties [127,128,129]. The mechanism behind this inhibition is still uncertain, but several studies have suggested that competition between both hGR isoforms for transcriptional coactivator proteins and/or the formation of inactive GRα-GRβ heterodimers might be responsible for that [126,130,131,132].
Moreover, the function of hGRβ extends beyond antagonism of the hGRα isoform [133]; for instance, binding to the glucocorticoid antagonist mifepristone (RU486) has been also reported [134,135]. In addition, it has been shown that increased hGRβ expression is correlated both with the development of immune-related diseases (e.g., ulcerative colitis, leukemia and severe asthma) [136,137,138] and with glucocorticoid resistance in patients affected by these diseases [139,140,141]. Interestingly, previous studies have established the occurrence of a GR β-isoform in zebrafish larvae, which similarly to the hGRβ, confirmed the lack of a role in transcriptional regulation and a dominant-negative inhibitor activity on zGRα [19,20,142,143]. In this respect, zebrafish have been shown to be a reliable and useful model system both for GC resistance and glucocorticoid receptor research [2,85].
2.3. GCs Mechanisms of Action
The conventional view of GC mechanism of action has been recently revised and described as a more complicated multiprotein-regulated process. In this regard, it has been shown that in both humans and zebrafish, upon cortisol binding, GR undergoes a conformational change that involves an FKBP51-FKBP52 exchange. The latter triggers the translocation of the GC-GR active complex into the nucleus. FKBP51 is a cochaperone protein that binds HSP90 and decreases the affinity of GR for cortisol. For this reason, FKBP51 has been considered an inhibitor of GR transcriptional activity and its overexpression has been linked to GC resistance in autoimmune diseases [144,145,146]. After ligand binding, FKBP51 is replaced by FKBP52, which in turn recruits dynein to support translocation of the GC/GR complex to the nucleus [147,148]. This structural modification exposes the two GR nuclear localisation signals, allowing the hormone-activated GR to dimerize with another GC-GR molecule and to migrate into the nucleus via nuclear pores [149,150]. Interestingly, this transcription factor complex can also act nongenomically (in the cytoplasm), where it may interact via direct protein-protein interactions with other transcriptional regulators and/or kinases (e.g., basal transcription machinery (BTM); phosphoinositide 3-kinase (PI3K); signal transducer and activator of transcription (STAT)) [63,151,152,153] (Figure 2).
Inside the cell nucleus, GC-GR complexes can directly bind to specific GREs, as tetramers, to upregulate (transactivation) or downregulate (transrepression) the transcription of target genes. Generally, the preferred GRE motif (AGAACAnnnTGTTCT in humans, and GGAACAnnnTGTTCT in zebrafish) is an imperfect palindromic consensus sequence that consists of two 6 bp half sites. The three-nucleotide spacing in-between the two half sites is essential for the GR to tetramerize on this sequence. Previous genome-wide studies have shown that the same GRE can mediate both the GC-dependent induction of many genes (positive GRE) and the repression of others (negative GRE) [113,154]. Interestingly, the presence of specific inverted repeats negative GREs (IR nGRE), unrelated to simple GREs has also been reported both in mice and in humans. These DNA binding sequences are palindromic sequences consisting of two inverted repeated (IR) motifs separated by 1 bp. In particular, they bind GC-GR complexes to promote the assembly of cis-acting GR-SMRT/NCoR repressing complexes [155,156] (Figure 2).
In summary, these findings indicate that the broadly different GC effects on various tissues can be partially ascribed to cell type-specific differences in the chromatin landscape that affects the accessibility of specific GREs for GR binding [157,158]. Furthermore, the GC concentration at which the GR binds to GREs depends on the cell type and chromosomal context. Another important feature of the GC-GR complex that makes its effects even more versatile is that it can tune gene expression in different ways: by binding directly to DNA, by tethering itself to other transcription factors bound to DNA, or via direct binding to DNA and with neighbouring DNA-bound transcriptional regulators (composite manner, Figure 2) [113,159,160].
2.4. The Mineralocorticoid Receptor: Structure and Functions
As previously mentioned, cortisol can bind not only to GR, but also to MR. Both are members of the steroid receptor superfamily (corticosteroid receptors) of ligand-activated transcription factors that enhance or repress the transcription of target genes, as well as promote rapid nongenomic/extra-nuclear events via several cell signalling pathways [47]. The mineralocorticoid receptor is a 984-amino acid cytoplasmic protein that is characterized by three different domains: an N-terminal transcriptional regulator domain, a DNA-binding domain, and a ligand-binding domain responsible for the selectivity of hormone binding. Analogously to GR, in its unliganded state MR is associated with a number of chaperone proteins (HSP90, HSP70, FKBP51 and p23) that play a crucial role in trafficking and maintaining MR in a suitable conformation for ligand binding [161].
MR has a ten-fold higher affinity for cortisol than GR and is preferably activated under basal conditions, implying distinct roles for each receptor in the regulation of HPA axis activity [162,163] It has also been observed that cortisol, even at lower concentrations than those required to activate the GR, binds to MR and might enhance the activity of several kinases (i.e., protein kinase C (PKC), cyclic adenosine 3′,5′-monophosphate (cAMP), and phosphoinositide 3-kinases (PI3K)) involved in different signal transduction cascades [4,47,164]. On the other hand, GR is primarily activated as a result of stress, or at the diurnal peak when circulating cortisol levels are also peaking [162,165]. Cortisol also exerts broad effects on mood and behaviour via MRs and GRs that are expressed in different regions of the brain [166,167]. In particular, GR is widely expressed throughout the brain, primarily in the PVN (stress-regulating centre) and in the prefrontal cortex-hippocampal-amygdala circuitry (cognitive, emotional regulation and memory consolidation centre). Vice versa, MR is predominantly expressed in the hippocampus, amygdala, and the lateral septum (emotionality, social behaviour and feeding process hub) [168].
In mammals, the mineralocorticoid system is essential to regulate potassium and fluid homeostasis upon aldosterone activation of MR. Even though cortisol is a high-affinity ligand for MR, this steroid is deactivated in specific mineralocorticoid responsive tissues, such as the kidney, by the previously mentioned 11b-HSD-2 enzymatic activity. This allows aldosterone, a second corticosteroid present in mammals, to bind to this receptor. Surprisingly, teleosts do not synthesize aldosterone, and cortisol has been shown to mediate stress axis regulation, as well as the majority of the changes in iono-regulatory and osmo-regulatory functions, via GR and MR signalling [1,4].
Interestingly, previous work performed both in rats and in teleosts showed that while MR is involved both in basal and onset of stress-induced HPA/I axis activity, GR mainly controls its termination [4,169]. However, a recent zebrafish study highlighted that rapid locomotor responses to quick changes in light illumination or water salinity (environmental stressors) require GC-GR mediated HPI axis signalling, but not MR [170]. Finally, our recently published research suggested that not only GR, but also MR signalling, is involved in the GC-negative feedback regulation (HPI axis termination) and plays a key role in assuring a proper HIF response in teleosts [171]. In view of this, furthering the precise role of MR and mineralocorticoid modes of action in vivo, particularly in relation to in the HIF signalling pathway, is warranted.
2.5. The Role of Glucocorticoids in Inflammation
Glucocorticoids suppress most of the events early in the inflammatory response and subsequently facilitate inflammation resolution. They suppress both vasodilation and the enhanced vascular permeability that occurs as a consequence of an inflammatory challenge, and inhibit leukocyte migration from the inflamed region [10,172,173]. In addition, they tune both distribution and trafficking of leukocytes, promote death/survival in some cells and may affect cellular differentiation programmes [173,174,175,176,177,178]. It is well established that many of the anti-inflammatory and immunosuppressive glucocorticoid actions are referable either indirectly or directly to GC-GR mediated transcriptional regulation of numerous genes expressed in leukocytes [173]. On the other hand, even though MR expression has been reported in immune cells [179,180,181], its anti-inflammatory role has been considered negligible so far [181,182], but rather surprisingly, MR-dependent proinflammatory effects have also been noted [183,184]. It is presently unclear whether these effects are mediated by the glucocorticoid cortisol or the mineralocorticoid aldosterone [185].
Glucocorticoids, through GR, can modulate gene expression in different ways [112,117]. Among these, transactivation is the mechanism by which GC predominantly induce the transcription of numerous anti-inflammatory genes, such as GILZ, MKP-1 and IĸBα. This occurs through direct binding of single or multiple GC-GRs to palindromic glucocorticoid response elements (GREs) [186,187,188,189]. Importantly, this mode of action has also been shown to be responsible for several undesirable metabolic side effects linked to chronic GC treatment [148]. On the other hand, transrepression is the mechanism by which GC downregulate the transcription of inflammatory genes and requires direct protein-protein interaction of GR to other transcription factors. This mode of action is generally accepted to convey the beneficial GC anti-inflammatory effects, which are mainly implicated in rapid cellular responses [40,148,190,191,192]. In particular, transrepression is known to mainly occur via direct binding between the monomeric GC-GR complex and transcription factors (e.g., AP-1, NF-kB, c-Jun, and c-Fos) activated by cytokines and other pro-inflammatory stimuli, which synergistically coordinate the expression of several proinflammatory genes [27,109,160,193,194,195,196]. Of note, most of these genes are commonly overexpressed during chronic nonresolving inflammatory states. Interestingly, transrepression is not restricted to these transcription factors but also includes others such as CREB, STAT, and T-bet [160,197]. As a result, the mutual antagonism between transcription factors frequently impairs their transcriptional properties and prevents them from binding to their corresponding DNA response elements.
As a consequence of the above considerations, optimal GC analogs should be characterized by a high inhibitory activity against inflammatory mediators, coupled with a low transactivation activity, in order to induce minimal side effects. Interestingly, different steroidal and nonsteroidal ligands have been reported to have this dual function (e.g., RU-24858 and ZK-216348) [148,191,198,199,200]. These compounds have been reported to suppress key inflammatory and immune transcription factor activity in vivo [198,199,200,201,202]. However, as stated before, since GCs can trigger gene expression via multiple routes, unexpected secondary side effects might occur. For this reason, further research is also warranted to elucidate the implications of the nongenomic GC-mediated activity both in the immune and inflammatory scenario.
Importantly, although the GC-GR complex is known for its anti-inflammatory effects, the picture is more complex. Indeed, contrary to expectations, GR loss of function was reported by Facchinello and coworkers to prevent the transcriptional activity linked to the inflammatory immune response (i.e., of cytokines Il6, Il1β, Il8 and Mmp-13) [8], corroborating the hypothesis of a GC-GR mediated dual action on the immune system [35,203]. However, it is clear that further research is warranted to better elucidate this aspect. In addition, GR was shown to synergistically induce proinflammatory genes by acting on other signalling pathways [204,205,206,207]. Finally, studies also demonstrated that GCs increase the transcription of numerous anti-inflammatory molecules such as interleukin-10 (IL-10), interleukin-1 receptor antagonist (IL-1RA), secretory leukocyte inhibitory protein and neutral endopeptidase [41,208].
Previous research also revealed that alterations in chromatin structure are important for modulating the outcome of GC activity. Indeed, GR can interact differently with histone acetyltransferases (HATs), with histone deacetylases (HDACs) and also kinases (i.e., MSK1, PKA and JNK) [160]. These may, in turn, modulate the chromatin environment by modifying chromatin accessibility and further tuning inflammatory and immune gene expression [160]. Furthermore, since chromatin accessibility can predetermine GR binding patterns and is crucial for cell-specific outcomes, it can provide novel molecular basis for tissue selectivity [209,210]. In addition, another study showed that GR may directly inhibit CREB binding protein (CBP)-associated HAT activity and may recruit HDAC2 to the p65-CBP HAT complex [211]. This novel glucocorticoid repression mechanism suggests that histone acetylation inhibition represents an additional level of control of inflammatory gene expression. Consequently, this further indicates that pharmacologically manipulating specific histone acetylation status could be an alternative approach for treating inflammatory diseases.
3. The HIF Signalling Pathway
The progression of the cell cycle is an energy-requiring process that demands a refined metabolic regulation to occur. Indeed, it is well known that cells must overtake an energy restriction checkpoint during G1 phase, in order to progress through the cell cycle. In this regard, as practically all metazoan organisms require oxygen for metabolically converting nutrients into energy, O2 represents a vital signalling molecule directing cellular functioning and survival. Consequently, assuring oxygen homeostasis is a critical task that must be precisely managed by cells in order to perform correctly and survive in a hostile environment [212].
In vertebrate cells, this is primarily carried out by Hypoxia-Inducible Factors (HIFs), which are a family of transcription factors that react both to environmental oxygen and cellular energy alterations (e.g., hypoxia) [213]. HIFs are obligate heterodimers consisting of three main α-subunits (HIF1α, HIF2α and HIF3α) and two constitutively stable aryl hydrocarbon receptor nuclear translocators or β-subunits (ARNT1 and ARNT2). Both HIF alpha and beta subunits are expressed in the cytoplasm and are basic helix-loop-helix (bHLH)/Per-Arnt-Sim (PAS) transcription factors containing transactivation domains (TADs) [214]. In particular, the initial bHLH domain, required for DNA binding, is followed by a Per-Arnt-Sim (PAS) domain that acts as a molecular sensor. The latter is followed by an oxygen-dependent degradation domain (ODD), which is targeted and hydroxylated by the VHL-E3-ubiquitin ligase complex, and by an N- and C-terminal transactivation domains (TAD-N and TAD-C). The latter is present in all the HIF-α subunits except for HIF-3α, which only has the TAD-N activation domain. On the other hand, HIF-β/ARNT being not targeted and degraded by the pVHL-E3 ligase complex lacks both the ODD and the TAD-N domains, and has a constitutively active N-terminal nuclear localization signal (NLS) [215,216] (Figure 3A).
As previously mentioned, HIF-α subunits are characterized by a very fast turnover that is post-translationally regulated by the PHD3-VHL-E3-ubiquitin ligase protein degradation complex. Under normoxic conditions (normal oxygen levels), a set of prolyl hydroxylases (PHD1, 2 and 3) directly exploit the available molecular oxygen to hydroxylate two prolyl residues (Pro402, Pro564) within the oxygen-dependent degradation domain (ODDD) of the HIF-α subunits (Figure 3A). Then, the hydroxylated HIF-α isoforms are recognized and targeted by the Von Hippel Lindau protein (pVHL), which acts as the substrate recognition component of the E3-ubiquitin ligase complex. The latter is characterized by a multiprotein complex which consists of Elongin B, Elongin C, Ringbox 1 and Cullin 2 [217]. Once this complex ubiquitinates the HIF-α subunits, it directs them towards proteasomal degradation to avoid an aberrant stabilization and activation of the HIF pathway when it is not necessary (Figure 3B, normoxia). In addition, HIF-α may be hydroxylated by factor-inhibiting HIF (FIH) on asparagine 803 (N803), which prevents the recruitment of the transcriptional coactivator p300/CREB-binding protein (p300/CBP) and reduces the effectiveness of HIF transcriptional activation [218].
Another level of complexity in the regulation of HIF response is represented by the fact that both FIH and PHDs are dioxygenases requiring O2, ferrous iron (Fe2+), 2-oxoglutarate and ascorbate, as cosubstrates, to hydroxylate HIF-α [219]. The ferrous iron is an essential cofactor for the enzyme to be assembled into its active conformation, as prolyl hydroxylases contain Fe2+ in their hydrophobic active centre. Moreover, the O2-binding event requires ascorbate to maintain this iron molecule into its ferrous state, whereas the transferring of one oxygen atom to 2-oxoglutarate (2-OG) is required to hydroxylate HIF-α subunit. This reaction yields both succinate and carbon dioxide as reaction products, making this catalytic process irreversible. During a complete reaction, Fe2+ is transiently oxidized to Fe4+ and then reduced to the Fe2+ state. Interestingly, when α-ketoglutarate is converted into succinate without hydroxylation of a peptide substrate, Fe2+ is oxidized to Fe3+. In this process, ascorbate is also necessary to reduce Fe3+ back to Fe2+ to allow the enzyme to be recycled [220]. Finally, mitochondrial reactive oxygen species (ROS), may also affect hydroxylase activity by post-translationally modifying these enzymes by oxidizing their cysteine residues, and/or by attacking ferrous iron (Fe2+) [221,222].
By contrast, the presence of reduced O2 levels impairs both PHD and FIH enzymatic activity and leads to HIF-α stabilization. Therefore, pVHL is no longer able to recognize and target HIF-α to proteasomal degradation when the latter is not hydroxylated. This allows HIF-α and β subunit heterodimer formation, followed by ARNT-mediated translocation in the nucleus. Here, p300/CBP may interact with the HIF-αβ transcription complex to further activate the hypoxic response. This implies the upregulation of target genes that are involved in decreasing oxygen consumption and increasing oxygen and nutrient delivery [31]. This occurs via direct recognition and binding of HIF to hypoxia-response elements (HREs). The latter are characterized by the presence of a consensus sequence G/ACGTG located within the promoter regions of target genes, such as phosphofructokinase, adrenomedullin, erythropoietin and vascular endothelial growth factor [73,74,78] (Figure 3B, hypoxia).
The Role of Hypoxia in Inflammation
Hypoxia and inflammation share an intimate relationship. Both in cells and tissues the hypoxic response plays a vital role in the metabolic changes that control cellular adaptation to low oxygen availability. In a hypoxic scenario, HIF exerts a proinflammatory role by stimulating multiple aspects of the host immune system, from enhancing the phagocyte microbicidal capacity, to driving T-cell differentiation and cytotoxic activity [223,224]. The HIF-mediated activation of the inflammatory response is a complex process, which is characterized by the simultaneous activation of both pathways in several pathological circumstances such as chronic inflammation, wound healing and solid tumours [225]. Hypoxia is able to activate monocytes, macrophages and dendritic cells by tuning their gene expression and cytokine secretion [226,227,228,229]. In addition, it triggers NF-κβ stabilisation, which acts as a master regulator of the inflammatory and anti-apoptotic response. This is achieved, as for HIF-α, via the oxygen dependent inhibition of prolyl hydroxylase activity. Then, this triggers the decrease in IκB kinase beta (IKKβ) hydroxylation, which leads to the activation of NF-κβ [87,230,231,232]. In turn, in response to hypoxia, LPS, or bacterial infection, NF-κB has been demonstrated to directly increase HIF-1α transcription [233,234,235]. This allows further confirmation of the extensive crosstalk between these two major molecular players involved in inflammation and hypoxia [236]. This interaction has been well studied in a zebrafish model of wound healing, where HIF-1α pathway activation was observed to delay neutrophil resolution [82]. This is believed to occur as a consequence of HIF activation inside the neutrophils themselves, and seems to be related to an augmented neutrophil apoptosis rate coupled with decreased trafficking away from the comorbid site [237].
Interestingly, several studies have also reported the presence of an interplay between hypoxia and glucocorticoid-dependent signalling pathways. Indeed, glucocorticoids have been observed via in vitro studies both to enhance and inhibit HIF pathway activation [77,78,79,80,81]. Analogously, hypoxia has been shown to attenuate the glucocorticoid anti-inflammatory response and to elicit corticosteroid insensitive inflammation [238,239,240].
In this regard, it is important to note that despite hypoxia being considered pro-inflammatory, HIF can also positively interact with pathways with apparently opposite effects. For instance, it was shown that HIF and GC signalling converge at the level of the promoter region of inflammatory factors to reciprocally tune their expression in a T-lymphocyte cell line model [241]. In relation to inflammation regulation, particular attention should be also placed on GILZ [242], which is an important mediator of the GC anti-inflammatory and immune-suppressive activity. Indeed, GILZ overexpression in T-cells has been demonstrated to suppress NF-κB activation by binding to its p65 subunit and preventing its nuclear translocation [243,244]. Moreover, it has been reported that hypoxia can strongly upregulate GILZ expression in rat macrophages, and that pretreatment with synthetic GC (Dexamethasone) amplifies these effects [242]. Additionally, in vitro studies revealed that under hypoxic conditions GILZ inhibition led to increased transcription and protein secretion both of proinflammatory mediators IL-1 and IL-6 and abolished the GC inhibitory effect on their expression. These findings indicate that GILZ plays a key role in tuning adaptive responses to hypoxic conditions by inhibiting pro-inflammatory responses and by mediating the GC anti-inflammatory responses [242].
Macrophages, which are broadly classified as M1 (pro-inflammatory), or M2 (anti-inflammatory) are a class of immune cells, residing in all tissues, that play a crucial role during inflammation [245]. In addition to their protective immunological function they also stimulate growth factors and pro-angiogenic cytokine expression such as basic fibroblast growth factor (FGF-2) and vascular endothelial growth factor (VEGF-A), thereby triggering angiogenesis [246]. Interestingly, under hypoxic conditions, HIF can also promote their polarization towards the M2 phenotype in order to modify the inflammatory microenvironment by decreasing the release of proinflammatory cytokines [247].
Conversely, GCs, being angiostatic, are often exploited to treat angiogenesis-related diseases, including solid tumours. In particular, GCs control angiogenesis by inhibiting proliferation, migration and sprouting of endothelial cells, and by decreasing both cytokines and pro-angiogenic factor expression [248]. Consequently, since both hypoxia and GC are specifically involved in inflammation, where they tune angiogenesis and affect macrophage function (i.e., by upregulating GILZ expression), understanding how precisely this interplay occurs in vivo, may have a wide physiological significance in health and disease, and may help researchers to develop more effective anti-inflammatory drugs in the future.
4. HIF-GC-Crosstalk: Previous Insights
Modulating the HIF pathway has the potential to be clinically exploited as therapeutic treatment for a variety of pathological conditions which include stroke, ischemia, spinal cord injury, inflammation, cancer, wounding, chronic anaemia and bone regeneration [99,224,249,250,251,252]. To this end, an unbiased chemical screen performed in our laboratory on zebrafish larvae discovered that HIF associated transcriptional responses are potently activated by GCs, particularly in the zebrafish liver [253].
Moreover, by translating these observations to human tissues, it has been possible to show that GCs promote HIF stabilization, without the need of the GR DNA binding domain (non-genomic action), in primary human hepatocytes and intact liver slices. In this regard, since c-src inhibitor PP2 treatment was able to rescue this effect, this suggested a role for GCs in promoting c-src–mediated proteasomal degradation of pVHL followed by stabilization of HIF-α subunit [253]. According to these data, since the liver is an important regulator of blood glucose levels, and both GC and HIF promote gluconeogenesis and glycogen storage in the liver, the crosstalk between these may contribute to GC effects on glucose metabolism and may have a wider physiological significance in health and disease than previously expected.
Indeed, both GCs and hypoxic transcriptional responses are mutually involved in assuring tissue homeostasis by controlling cellular responses to various forms of stress and inflammation, especially affecting glucose metabolism.
Synthetic GCs have been widely used for years as anti-inflammatory drugs for treating pathological conditions in which hypoxia plays a role in disease progression, such as rheumatoid arthritis and chronic obstructive pulmonary disease [31,32,35]. In humans, the adaptive GC release in response to atmospheric hypoxia has been shown to be linked to acclimatisation to high altitude, and the prophylactic treatment with GC has been broadly exploited to mitigate the related mountain sickness [254]. Additionally, GC administration protects different organs from ischemic injury, as was observed especially against experimental cerebral and hepatic ischemic/reperfusion injury [255,256,257,258].
The presence of an interplay between hypoxia and GC-dependent signalling pathways has been previously reported in different in vitro studies [78,79,80,259]. However, these studies reported conflicting results on the crosstalk between GC action and hypoxia, where the latter limits GR-mediated transactivation both in pulmonary endothelial and hepatic epithelial cells [78,80]. The first data about the interaction between HIF and GR were presented by Kodama et al., 2003 [79]. By exploiting an artificial approach using Gal4-fusion reporter assays, they found that the ligand-dependent activation of GR increases hypoxia-dependent gene expression and hypoxia response element (HRE) activity in HeLa cells.
Moreover, using dexamethasone-treated COS7 cells co-transfected with expression plasmids for either GR or GAL4-LBD and GFP-HIF-1α and exposed to hypoxic conditions, they showed colocalization of the GR and HIF-1α in the nucleus. For this reason, Kodama and colleagues postulated the presence of a direct protein-protein interaction between the GR LBD and HIF-1α as the main mechanism for GC-dependent enhancement of the HIF pathway but failed to demonstrate it via GST pulldown assays.
Leonard et al., 2005 [78], subsequently confirmed via microarray analysis that GR is upregulated (from 7-fold to 12-fold) by hypoxia over time (0–36 h) in human renal proximal tubular epithelial cells. Furthermore, using a cell-based GRE luciferase reporter system, they showed that hypoxic exposure can potentiate dexamethasone-stimulated GRE promoter-reporter activity.
Using ACTH secreting mouse pituitary tumor AtT-20 cells, Zhang et al., 2015 [259] demonstrated that GR expression levels were enhanced by HIF-1α under hypoxic conditions. However, dexamethasone treatment was able to cause the downregulation of GR expression in a HIF-1α dependent way. Finally, even if this was confirmed by transfecting AtT-20 cells with HIF-1α siRNA and culturing them under normoxia or hypoxic conditions, the involved underlying mechanism remains unclear. Unfortunately, the impact on downstream GR/GC pathway activity was not assessed.
By contrast, a dexamethasone-related inhibition of HIF-1α target gene expression in hypoxic HEPG2 cells was revealed by Wagner et al., 2008 [80]. In particular, via Western blot analysis they showed that dexamethasone reduces nuclear HIF-1α protein, as the HIF-1α amount was higher in cytosolic cell extracts than in the nuclear extracts upon DEX treatment. This cytoplasmic retention of HIF-1α suggested a blockage of nuclear import, via a still unknown mechanism, which resulted in a reduced HIF target gene expression. In addition, by exploiting a luciferase assay the author revealed that dexamethasone attenuates HIF-1 activity not only in a GR- dependent way but this effect depends on the presence of functional HREs. Importantly, Wagner et al. attributed these contradicting results (compared to Kodama’s) to the fact that they conducted all their cell-culture experiments in the presence of fetal bovine serum (FBS), avoiding not only growth and cell-cycle arrest, but also the synchronization of the cells. This might be an important issue, as many cellular processes depends on the cell cycle phase.
In the following years, Gaber et al., 2011 [260] investigated the interaction between macrophage migration inhibitory factor (MIF), HIF and the effect of GCs in human primary nontumor CD4+ Th cells and Jurkat T cells. In contrast to the previous observations, this study showed the presence of a clear dexamethasone-dose dependent inhibition of HIF-1α protein expression, which resulted in decreased HIF-1 target gene expression. Therefore, as an alternative to the Wagner et al. 2008 hypothesis, they proposed a model based either on rapid DEX-mediated induction of HIF-1α inhibitors (e.g., PHD1–3, FIH, and pVHL) or fast DEX-mediated suppression of hypoxia-induced signalling. Cumulatively, these contradicting in vitro data reflect the diversity present among the cell types that were used in these studies and the different mechanisms by which these were conducted. Methods that are less cell line and cell culture-dependent would be helpful to understand the interaction better, and in vivo analysis would be valuable, in this respect.
5. HIF-GC-Crosstalk: HIF as Negative Regulator of GC Biosynthesis and Responsiveness
Zebrafish have been demonstrated to be informative organisms in which genetic mutations and chemical modulators of HIF and GC signalling pathways can be easily combined, not only in physiological, but also in pathophysiological conditions. Using such approaches [89,91,261,262], we demonstrated that the upregulation of HIF signalling repressed both GR responsiveness and cortisol levels, whereas GCs enhanced HIF activity. The latter effect was mediated both by the GR and MR receptors, and we speculated that GC can promote HIF-1 signalling via multiple routes [171].
With respect to the effect of HIF signalling on GC responsiveness, it was particularly interesting to note that a strong activation of HIF signalling can blunt GR transcriptional regulation, whereas reduction of HIF signalling via arnt1 loss of function is able to derepress it. Interestingly, HIF activity leads to a repression of GC biosynthetic genes, which is unusual since HIF is generally considered to be a transcriptional activator. However, the precise molecular mechanism behind this the strong negative HIF-mediated action remains to be resolved.
As stated before, GCs control a plethora of physiological processes, act on almost all the tissues and organs in the body and have a strong anti-inflammatory and immunosuppressive actions. For this reason, their production must be finely controlled by the HPA/I axis [113]. If the latter is disrupted by factors such as chronic stress, disease progression and prolonged exogenous GC treatment, a tissue-specific decrease in GRα functional pool coupled with an uncontrolled GC biosynthesis may result in the development of acquired GC resistance. This results in both excessive inflammation and HPA axis hyperactivity, which are known to contribute to the progression of numerous psychological and pathological conditions (e.g., cardiovascular disease, depression, schizophrenia, diabetes, and cancer, among others) [263,264].
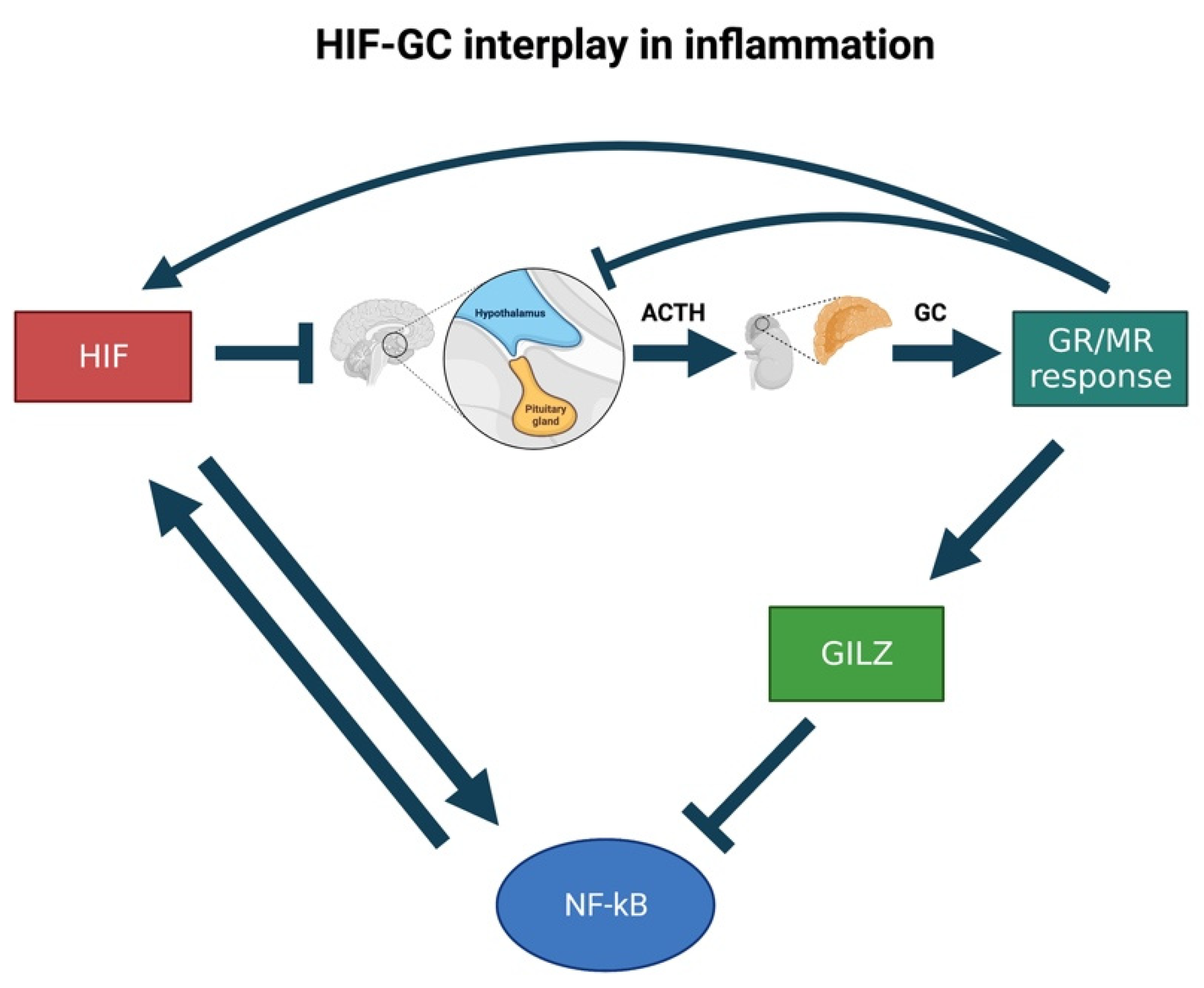
Cumulatively, the presence of high cortisol levels coupled to a significant decrease in GC sensitivity are seen as a hallmark of glucocorticoid resistance (e.g., in the gr mutant fish). Therefore, it is rare that a GC-resistant condition coincides with low cortisol levels. In addition, many studies have shown that if GC levels drop, pomca is upregulated to compensate for the reduced GC levels and feedback [2,87]. However, in the presence of upregulated HIF pathway (in vhl mutant fish), an unusual situation is seen where pomca is not upregulated, although GR-responsiveness is strongly attenuated, and GC levels are very low. Consequently, we speculate that HIF signalling can act as a second upstream controller of the GC-mediated stress response (in addition to cortisol levels themselves) (Figure 4).
Indeed, since previous work in our laboratory highlighted that GCs may also act as HIF activators [76,253] we inferred that HIF may, in turn, control cortisol levels by acting on pomca expression. This would allow HIF signalling not only to manage its own activity, but also to assure both stress resolution and homeostasis. The reason for hypothesising that HIF signalling would counteract anti-inflammatory GC activity resides in the fact that the simultaneous expression of both the upregulated HIF and GC pathways would be detrimental to homeostasis. This is because HIF is a master regulator of cellular pro-inflammatory responses to hypoxia, whereas GCs have a potent anti-inflammatory and immune suppressive activity [171]. In this regard, it is interesting to note that our proposed model effectively applies to the immune response context. Indeed, during inflammation, we speculate that the crosstalk between GC and HIF may influence the expression of downstream inflammatory effectors (e.g., GILZ and NF-κB) to eventually assure homeostasis (Figure 5).
This hypothesis is in accordance with a previous report showing that hypoxia exposure resulted in the downregulation of steroidogenic genes (StAR, cyp19b, cyp19a, cyp11c1, hsd17b2 and hmgcr) in 72 hpf larvae, whilst zHIF-α loss of function stimulated the upregulation specifically of StAR, cyp11b2 and cyp17a1 [265]. Importantly, the fact that cortisol levels were reduced in vhl-/- and upregulated in arnt1-/- is consistent with our assumption. More speculatively, the fact that in teleosts GC biosynthesis is finely regulated by hypothalamus-pituitary gland activity in a circadian way, and multiple studies have shown an evolutionary connection between HIF signalling and circadian rhythms [266,267,268,269], suggest that the interaction between GC and HIF may be tight and ancient.
Importantly, recent work from Watts et al., 2021 confirmed the role of HIF1α as a direct regulator of steroidogenesis in the adrenal gland. Indeed, mice deficient in HIF1α in adrenocortical cells exhibited both increased levels of steroidogenic enzymes, such as Star, Cyp11a1, Cyp21a1 and Cyp11b1, and an enhancement in circulatory steroid levels. These changes also resulted in cytokine alterations and modifications of the profile of circulatory mature hematopoietic cells. However, additional mechanisms are possible and indeed likely, one example being via miRNA103/107 [270]. Vice versa, HIF1α overexpression induced the opposite phenotype characterized by strongly downregulated steroid production as a result of impaired transcription of steroidogenic enzymes [271]. Therefore, the data derived from zebrafish research are in accordance with mice studies. As a consequence of the above considerations, the HIF-mediated pomca negative regulation seems to be a logic homeostatic response. Moreover, with respect to the stimulating effects of GCs on HIF signalling, it is important to point out that in vivo genetic analysis showed that functional GR is an essential prerequisite for high HIF signalling levels. Unfortunately, a molecular explanation for this is still elusive, and research towards this direction (e.g., ChIP seq, pull-down assay) is warranted.
Finally, the importance of GCs in the HIF pathway was further analysed by studying MR contribution to HIF signalling itself. Our data showed that in addition to the glucocorticoid receptor, the mineralocorticoid receptor acts partially redundantly to allow HIF signalling in the zebrafish. This hypothesis is consistent with work from Faught and Vijayan [4] showing that both Gr and Mr signalling are involved in the regulation of the GC negative feedback. It is also consistent with the finding from Rybnikova et al., that both GR and MR are involved in brain hypoxic tolerance induction [272]. If these results hold up in mammals, this suggests GR and MR inhibitors may be usable to reduce excessive HIF activity in certain situations; for instance, in vhl deficient tumours.
In addition, dexamethasone (GC agonist) has been recently shown to decrease short-term mortality and to reduce the need for mechanical ventilation in hospitalised patients with COVID-19 [273,274]. Since the latter show low blood oxygen saturation, and HIF is very likely to be overactivated in them, could it be that they have either too low GC levels or GR responsiveness? Since synthetic GC administration rescues these patients with upregulated HIF, is there a functional link between these two facts? Is this related to what we have seen in the zebrafish?
Future work that could be brought to any potential drug development would include additional preclinical testing both in vitro and in vivo, aiming to better elucidate how HIF-GC interaction take places at molecular level. Cumulatively, understanding the molecular mechanisms behind the HIF-GR crosstalk and their target gene activation is of undeniable importance and deserves significant research investment in the near future.
6. Conclusions and Future Directions
The development of novel therapeutic targets and of innovative GCs with a better benefits-risk ratio is of indisputable relevance to effectively treating immune and inflammatory responses. In this regard, the zebrafish has been shown to be an effective model organism that can be used for investigating not only the molecular mechanism behind glucocorticoid receptor modes of action, but also in drug discovery studies. To this end, it is important to define efficient readouts for both desired and undesired effects of GC.
In the last decades few in vitro studies have highlighted the potential for crosstalk between the hypoxia-inducible factor and glucocorticoid transcriptional responses. However, how this interplay takes place precisely in vivo has only been recently proposed, and use of the zebrafish as a model organism has shown that HIF can repress not only cortisol biosynthesis, but also GR responsiveness to synthetic GC. Conversely, research has uncovered the importance of the GC pathway in driving HIF signalling and highlighted a novel mineralocorticoid receptor contribution to the HIF-GC crosstalk. In addition, the interaction between HIF-GC and NF-kB signalling might be of high relevance. Indeed, investigating the putative mechanisms behind adaptation to severe hypoxia might help to unravel the hypoxia-related causes and effects in acute inflammatory disease. Finally understanding how MR precisely impacts on HIF signalling might be extremely important to provide a potential additional avenue to downregulate the HIF pathway in vivo, as it has been proven difficult to do so in the tumour microenvironment. Overall, new insights into the interaction between HIF and GR, both in vitro and in vivo, are promising directions for future research and might inspire new therapeutic approaches.
Author Contributions
Conceptualization, D.M. and F.J.M.v.E.; writing—original draft preparation, D.M.; writing—review and editing, D.M. and F.J.M.v.E.; visualization, D.M. and F.J.M.v.E. All authors have read and agreed to the published version of the manuscript.
Funding
DM was awarded by a University of Sheffield, Biomedical Science department studentship and supported by two BBRSC grants to FVE (BB/R015457/1; BB/M02332X/1). FVE was funded by two Biotechnology and Biological Sciences Research Council (BBRSC: https://bbsrc.ukri.org/ (accessed on 2 December 2021)) grants (code 1: BB/R015457/1; code 2: BB/M02332X/1). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
Authors report no conflict of interest.
References
- Alsop, D.; Vijayan, M. The zebrafish stress axis: Molecular fallout from the teleost-specific genome duplication event. Gen. Comp. Endocrinol. 2009, 161, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, B.B.; Schoonheim, P.J.; Ziv, L.; Voelker, L.; Baier, H.; Gahtan, E. A zebrafish model of glucocorticoid resistance shows serotonergic modulation of the stress response. Front. Behav. Neurosci. 2012, 6, 68. [Google Scholar] [CrossRef] [Green Version]
- Tokarz, J.; Möller, G.; de Angelis, M.H.; Adamski, J. Zebrafish and steroids: What do we know and what do we need to know? J. Steroid Biochem. Mol. Biol. 2013, 137, 165–173. [Google Scholar] [CrossRef]
- Faught, E.; Vijayan, M.M. The mineralocorticoid receptor is essential for stress axis regulation in zebrafish larvae. Sci. Rep. 2018, 8, 18081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munck, A.; Guyre, P.M.; Holbrook, N.J. Physiological Functions of Glucocorticoids in Stress and Their Relation to Pharmacological Actions. Endocr. Rev. 1984, 5, 25–44. [Google Scholar] [CrossRef]
- Macfarlane, D.P.; Forbes, S.; Walker, B.R. Glucocorticoids and fatty acid metabolism in humans: Fuelling fat redistribution in the metabolic syndrome. J. Endocrinol. 2008, 197, 189–204. [Google Scholar] [CrossRef] [Green Version]
- Kuo, T.; McQueen, A.; Chen, T.-C.; Wang, J.-C. Regulation of Glucose Homeostasis by Glucocorticoids. In Glucocorticoid Signaling: From Molecules to Mice to Man; Wang, J.-C., Harris, C., Eds.; Springer: New York, NY, USA, 2015; pp. 99–126. [Google Scholar] [CrossRef]
- Facchinello, N.; Skobo, T.; Meneghetti, G.; Colletti, E.; Dinarello, A.; Tiso, N.; Costa, R.; Gioacchini, G.; Carnevali, O.; Argenton, F.; et al. nr3c1 null mutant zebrafish are viable and reveal DNA-binding-independent activities of the glucocorticoid receptor. Sci. Rep. 2017, 7, 4371. [Google Scholar] [CrossRef] [Green Version]
- Hench, P.S.; Kendall, E.C.; Slocumb, C.H.; Polley, H.F. The effect of a hormone of the adrenal cortex (17-hydroxy-11-dehydrocorticosterone: Compound E) and of pituitary Adrenocortical Hormone in Arthritis: Preliminary Report. Ann. Rheum. Dis. 1949, 8, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Perretti, M.; Ahluwalia, A. The microcirculation and inflammation: Site of action for glucocorticoids. Microcirculation 2000, 7, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Smoak, K.A.; Cidlowski, J.A. Mechanisms of glucocorticoid receptor signaling during inflammation. Mech. Ageing Dev. 2004, 125, 697–706. [Google Scholar] [CrossRef]
- Newton, R.; Holden, N. Separating Transrepression and Transactivation: A Distressing Divorce for the Glucocorticoid Receptor? Mol. Pharmacol. 2007, 72, 799–809. [Google Scholar] [CrossRef]
- Necela, B.M.; Cidlowski, J.A. Crystallization of the human glucocorticoid receptor ligand binding domain: A step towards selective glucocorticoids. Trends Pharmacol. Sci. 2003, 24, 58–61. [Google Scholar] [CrossRef]
- Rhen, T.; Cidlowski, J. Antiinflammatory Action of Glucocorticoids—New Mechanisms for Old Drugs. N. Engl. J. Med. 2005, 353, 1711–1723. [Google Scholar] [CrossRef] [Green Version]
- Yeager, M.P.; Guyre, P.M.; Munck, A.U. Glucocorticoid regulation of the inflammatory response to injury. Acta Anaesthesiol. Scand. 2004, 48, 799–813. [Google Scholar] [CrossRef]
- Kleiman, A.; Tuckermann, J.P. Glucocorticoid receptor action in beneficial and side effects of steroid therapy: Lessons from conditional knockout mice. Mol. Cell. Endocrinol. 2007, 275, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Newton, R.; Shah, S.; Altonsy, M.O.; Gerber, A.N. Glucocorticoid and cytokine crosstalk: Feedback, feedforward, and co-regulatory interactions determine repression or resistance. J. Biol. Chem. 2017, 292, 7163–7172. [Google Scholar] [CrossRef] [Green Version]
- De Bosscher, K. Selective Glucocorticoid Receptor modulators. J. Steroid Biochem. Mol. Biol. 2010, 120, 96–104. [Google Scholar] [CrossRef]
- Chatzopoulou, A.; Schoonheim, P.J.; Torraca, V.; Meijer, A.H.; Spaink, H.; Schaaf, M.J. Functional analysis reveals no transcriptional role for the glucocorticoid receptor β-isoform in zebrafish. Mol. Cell. Endocrinol. 2017, 447, 61–70. [Google Scholar] [CrossRef]
- Schaaf, M.J.M.; Champagne, D.; van Laanen, I.H.C.; van Wijk, D.C.W.A.; Meijer, A.H.; Meijer, O.C.; Spaink, H.P.; Richardson, M.K. Discovery of a Functional Glucocorticoid Receptor β-Isoform in Zebrafish. Endocrinology 2007, 149, 1591–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.T.; Hillhouse, E.W.; Burden, J.L. Dynamics and mechanics of corticosteroid feedback at the hypothalamus and anterior pituitary gland. J. Endocrinol. 1977, 73, 405–417. [Google Scholar] [CrossRef]
- Dallman, M.F.; Akana, S.F.; Jacobson, L.; Levin, N.; Cascio, C.S.; Shinsako, J. Characterization of Corticosterone Feedback Regulation of ACTH Secretion. Ann. N. Y. Acad. Sci. 1987, 512, 402–414. [Google Scholar] [CrossRef]
- Alsop, D.; Vijayan, M.M. Development of the corticosteroid stress axis and receptor expression in zebrafish. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R711–R719. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.R. Steroid receptor regulated transcription of specific genes and gene networks. Annu. Rev. Genet. 1985, 19, 209–252. [Google Scholar] [CrossRef]
- Hollenberg, S.M.; Weinberger, C.; Ong, E.S.; Cerelli, G.; Oro, A.; Lebo, R.; Thompson, E.B.; Rosenfeld, M.G.; Evans, R. Primary structure and expression of a functional human glucocorticoid receptor cDNA. Nature 1985, 318, 635–641. [Google Scholar] [CrossRef]
- Miesfeld, R.; Rusconi, S.; Godowski, P.J.; Maler, B.A.; Okret, S.; Wikström, A.-C.; Gustafsson, J.; Yamamoto, K.R. Genetic complementation of a glucocorticoid receptor deficiency by expression of cloned receptor cDNA. Cell 1986, 46, 389–399. [Google Scholar] [CrossRef]
- Jonat, C.; Rahmsdorf, H.J.; Park, K.-K.; Cato, A.; Gebel, S.; Ponta, H.; Herrlich, P. Antitumor promotion and antiinflammation: Down-modulation of AP-1 (Fos/Jun) activity by glucocorticoid hormone. Cell 1990, 62, 1189–1204. [Google Scholar] [CrossRef]
- Heck, S.; Kullmann, M.; Gast, A.; Ponta, H.; Rahmsdorf, H.; Herrlich, P.; Cato, A. A distinct modulating domain in glucocorticoid receptor monomers in the repression of activity of the transcription factor AP-1. EMBO J. 1994, 13, 4087–4095. [Google Scholar] [CrossRef]
- Heck, S.; Bender, K.; Kullmann, M.; Göttlicher, M.; Herrlich, P.; Cato, A.C. Ikappa Balpha -independent downregulation of NF-kappa B activity by glucocorticoid receptor. EMBO J. 1997, 16, 4698–4707. [Google Scholar] [CrossRef] [Green Version]
- Yang-Yen, H.-F.; Chambard, J.C.; Sun, Y.-L.; Smeal, T.; Schmidt, T.; Drouin, J.; Karin, M. Transcriptional interference between c-Jun and the glucocorticoid receptor: Mutual inhibition of DNA binding due to direct protein-protein interaction. Cell 1990, 62, 1205–1215. [Google Scholar] [CrossRef]
- Nikolaus, S.; Fölscn, U.; Schreiber, S. Immunopharmacology of 5-aminosalicylic acid and of glucocorticoids in the therapy of inflammatory bowel disease. Hepatogastroenterology 2000, 47, 71–82. [Google Scholar]
- Neeck, G.; Renkawitz, R.; Eggert, M. Molecular aspects of glucocorticoid hormone action in rheumatoid arthritis. Cytokines Cell. Mol. Ther. 2002, 7, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P.; Kino, T. Intracellular Glucocorticoid Signaling: A Formerly Simple System Turns Stochastic. Sci. STKE 2005, 2005, pe48. [Google Scholar] [CrossRef]
- Revollo, J.R.; Cidlowski, J.A. Mechanisms Generating Diversity in Glucocorticoid Receptor Signaling. Ann. N. Y. Acad. Sci. 2009, 1179, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Busillo, J.M.; Cidlowski, J.A. The five Rs of glucocorticoid action during inflammation: Ready, reinforce, repress, resolve, and restore. Trends Endocrinol. Metab. 2013, 24, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Reichardt, S.D.; Amouret, A.; Muzzi, C.; Vettorazzi, S.; Tuckermann, J.P.; Lühder, F.; Reichardt, H.M. The Role of Glucocorticoids in Inflammatory Diseases. Cells 2021, 10, 2921. [Google Scholar] [CrossRef]
- Van Staa, T.P.; Leufkens, H.G.M.; Abenhaim, L.; Begaud, B.; Zhang, B.; Cooper, C. Use of oral corticosteroids in the United Kingdom. QJM Int. J. Med. 2000, 93, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Moghadam-Kia, S.; Werth, V.P. Prevention and treatment of systemic glucocorticoid side effects. Int. J. Dermatol. 2010, 49, 239–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, P.J.; Adcock, I. Glucocorticoid resistance in inflammatory diseases. Lancet 2009, 373, 1905–1917. [Google Scholar] [CrossRef]
- Barnes, P.J. Glucocorticosteroids: Current and future directions. Br. J. Pharmacol. 2010, 163, 29–43. [Google Scholar] [CrossRef] [Green Version]
- Schaaf, M.J.; Cidlowski, J.A. Molecular mechanisms of glucocorticoid action and resistance. J. Steroid Biochem. Mol. Biol. 2002, 83, 37–48. [Google Scholar] [CrossRef]
- Spies, L.-M.L.; Verhoog, N.J.D.; Louw, A. Acquired Glucocorticoid Resistance Due to Homologous Glucocorticoid Receptor Downregulation: A Modern Look at an Age-Old Problem. Cells 2021, 10, 2529. [Google Scholar] [CrossRef]
- Howell, M.P.; Muglia, L.J. Effects of genetically altered brain glucocorticoid receptor action on behavior and adrenal axis regulation in mice. Front. Neuroendocrinol. 2006, 27, 275–284. [Google Scholar] [CrossRef]
- Zappia, C.D.; Torralba-Agu, V.; Echeverria, E.; Fitzsimons, C.P.; Fernández, N.; Monczor, F. Antihistamines Potentiate Dexamethasone Anti-Inflammatory Effects. Impact on Glucocorticoid Receptor-Mediated Expression of Inflammation-Related Genes. Cells 2021, 10, 3026. [Google Scholar] [CrossRef] [PubMed]
- Bamberger, C.M.; Schulte, H.M.; Chrousos, G.P. Molecular Determinants of Glucocorticoid Receptor Function and Tissue Sensitivity to Glucocorticoids. Endocr. Rev. 1996, 17, 245–261. [Google Scholar] [CrossRef]
- Faught, E.; Vijayan, M.M. Maternal stress and fish reproduction: The role of cortisol revisited. Fish Fish. 2018, 19, 1016–1030. [Google Scholar] [CrossRef]
- Gomez-Sanchez, E.; Gomez-Sanchez, C.E. The Multifaceted Mineralocorticoid Receptor. Compr. Physiol. 2014, 4, 965–994. [Google Scholar] [CrossRef] [Green Version]
- Spach, C.; Streeten, D.H.P. Retardation of Sodium Exchange in Dog Erythrocytes by Physiological Concentrations of Aldosterone, in Vitro. J. Clin. Investig. 1964, 43, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Moura, A.M.; Worcel, M. Direct action of aldosterone on transmembrane 22Na efflux from arterial smooth muscle. Rapid and delayed effects. Hypertension 1984, 6, 425–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttgereit, F.; Wehling, M.; Burmester, G.R. A new hypothesis of modular glucocorticoid actions: Steroid treatment of rheumatic diseases revisited. Arthritis Rheum. 1998, 41, 761–767. [Google Scholar] [CrossRef]
- Buttgereit, F.; Scheffold, A. Rapid glucocorticoid effects on immune cells. Steroids 2002, 67, 529–534. [Google Scholar] [CrossRef]
- Pereda, M.P.; Lohrer, P.; Kovalovsky, D.; Castro, C.P.; Goldberg, V.; Losa, M.; Chervin, A.; Berner, S.; Molina, H.; Stalla, G.K.; et al. Interleukin-6 is inhibited by glucocorticoids and stimulates ACTH secretion and POMC expression in human corticotroph pituitary adenomas. Exp. Clin. Endocrinol. Diabetes 2000, 108, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Topete, D.; Cidlowski, J. One Hormone, Two Actions: Anti- and Pro-Inflammatory Effects of Glucocorticoids. Neuroimmunomodulation 2015, 22, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnett, M.G.; Muglia, L.M.; Laryea, G.; Muglia, L.J. Genetic Approaches to Hypothalamic-Pituitary-Adrenal Axis Regulation. Neuropsychopharmacology 2015, 41, 245–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertini, R.; Bianchi, M.; Ghezzi, P. Adrenalectomy sensitizes mice to the lethal effects of interleukin 1 and tumor necrosis factor. J. Exp. Med. 1988, 167, 1708–1712. [Google Scholar] [CrossRef]
- Edwards, C.K., 3rd; Yunger, L.M.; Lorence, R.M.; Dantzer, R.; Kelley, K.W. The pituitary gland is required for protection against lethal effects of Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 1991, 88, 2274–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macphee, I.A.M.; Antoni, F.A.; Mason, D.W. Spontaneous recovery of rats from experimental allergic encephalomyelitis is dependent on regulation of the immune system by endogenous adrenal corticosteroids. J. Exp. Med. 1989, 169, 431–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandra, R.N. Neuro-hormonal host defence in endotoxin shock. Brain Behav. Immun. 1992, 6, 157–169. [Google Scholar] [CrossRef]
- Refojo, D.; Liberman, A.C.; Giacomini, D.; Nagashima, A.C.; Graciarena, M.; Echenique, C.; Pereda, M.P.; Stalla, G.; Holsboer, F.; Arzt, E. Integrating Systemic Information at the Molecular Level: Cross-talk between steroid receptors and cytokine signaling on different target cells. Ann. N. Y. Acad. Sci. 2003, 992, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Ruzek, M.C.; Pearce, B.D.; Miller, A.H.; Biron, C.A. Endogenous glucocorticoids protect against cytokine-mediated lethality during viral infection. J. Immunol. 1999, 162, 3527–3533. [Google Scholar]
- Silverman, M.N.; Sternberg, E.M. Glucocorticoid regulation of inflammation and its functional correlates: From HPA axis to glucocorticoid receptor dysfunction. Ann. N. Y. Acad. Sci. 2012, 1261, 55–63. [Google Scholar] [CrossRef]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialog. Clin. Neurosci. 2006, 8, 383–395. [Google Scholar]
- De Guia, R.; Rose, A.J.; Herzig, S. Glucocorticoid hormones and energy homeostasis. Horm. Mol. Biol. Clin. Investig. 2014, 19, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Katsu, Y.; Iguchi, T. Subchapter 95D—Cortisol. In Handbook of Hormones; Takei, Y., Ando, H., Tsutsui, K., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 532–533. [Google Scholar] [CrossRef]
- Cummins, E.P.; Taylor, C.T. Hypoxia-responsive transcription factors. Pflügers Arch. 2005, 450, 363–371. [Google Scholar] [CrossRef]
- Bertout, J.A.; Patel, S.A.; Simon, M.C. The impact of O2 availability on human cancer. Nat. Rev. Cancer 2008, 8, 967–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dougherty, E.J.; Pollenz, R.S. ARNT: A Key bHLH/PAS Regulatory Protein Across Multiple Pathways. Compr. Toxicol. 2010, 2, 231–252. [Google Scholar] [CrossRef]
- Pelster, B.; Egg, M. Hypoxia-inducible transcription factors in fish: Expression, function and interconnection with the circadian clock. J. Exp. Biol. 2018, 221, jeb163709. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.J.; Heiden, T.C.K.; Heideman, W.; Peterson, R.E. Potential Roles of Arnt2 in Zebrafish Larval Development. Zebrafish 2009, 6, 79–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasch, A.L.; Tanguay, R.L.; Mehta, V.; Heideman, W.; Peterson, R.E. Identification of Zebrafish ARNT1 Homologs: 2,3,7,8-Tetrachlorodibenzo-p-dioxin Toxicity in the Developing Zebrafish Requires ARNT1. Mol. Pharmacol. 2005, 69, 776–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elks, P.M.; Renshaw, S.; Meijer, A.H.; Walmsley, S.; van Eeden, F.J. Exploring the HIFs, buts and maybes of hypoxia signalling in disease: Lessons from zebrafish models. Dis. Model. Mech. 2015, 8, 1349–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L. Hypoxia-Inducible Factors in Physiology and Medicine. Cell 2012, 148, 399–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lando, D.; Peet, D.J.; Gorman, J.J.; Whelan, D.A.; Whitelaw, M.L.; Bruick, R.K. FIH-1 is an asparaginyl hydroxylase enzyme that regulates the transcriptional activity of hypoxia-inducible factor. Genes Dev. 2002, 16, 1466–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Rooijen, E.; Voest, E.E.; Logister, I.; Korving, J.; Schwerte, T.; Schulte-Merker, S.; Giles, R.H.; van Eeden, F.J. Zebrafish mutants in the von Hippel-Lindau tumor suppressor display a hypoxic response and recapitulate key aspects of Chuvash polycythemia. Blood 2009, 113, 6449–6460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pescador, N.; Cuevas, Y.; Naranjo, S.; Alcaide, M.; Villar, D.; Landázuri, M.O.; del Peso, L. Identification of a functional hypoxia-responsive element that regulates the expression of the egl nine homologue 3 (egln3/phd3) gene. Biochem. J. 2005, 390, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santhakumar, K.; Judson, E.C.; Elks, P.M.; McKee, S.; Elworthy, S.; Van Rooijen, E.; Walmsley, S.S.; Renshaw, S.A.; Cross, S.S.; Van Eeden, F.J. A Zebrafish Model to Study and Therapeutically Manipulate Hypoxia Signaling in Tumorigenesis. Cancer Res. 2012, 72, 4017–4027. [Google Scholar] [CrossRef] [Green Version]
- Basu, M.; Sawhney, R.C.; Kumar, S.; Pal, K.; Prasad, R.; Selvamurthy, W. Glucocorticoids as prophylaxis against acute mountain sickness. Clin. Endocrinol. 2002, 57, 761–767. [Google Scholar] [CrossRef]
- Leonard, M.O.; Godson, C.; Brady, H.R.; Taylor, C. Potentiation of Glucocorticoid Activity in Hypoxia through Induction of the Glucocorticoid Receptor. J. Immunol. 2005, 174, 2250–2257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodama, T.; Shimizu, N.; Yoshikawa, N.; Makino, Y.; Ouchida, R.; Okamoto, K.; Hisada, T.; Nakamura, H.; Morimoto, C.; Tanaka, H. Role of the Glucocorticoid Receptor for Regulation of Hypoxia-dependent Gene Expression. J. Biol. Chem. 2003, 278, 33384–33391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, A.; Huck, G.; Stiehl, D.; Jelkmann, W.; Hellwig-Bürgel, T. Dexamethasone impairs hypoxia-inducible factor-1 function. Biochem. Biophys. Res. Commun. 2008, 372, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-Y.; Wang, C.-Y.; Hsu, M.-F.; Juan, S.-H.; Chang, C.-Y.; Chou, C.-M.; Yang, L.-Y.; Hung, K.-S.; Xu, J.; Lee, Y.-H.; et al. Glucocorticoid Protection of Oligodendrocytes against Excitotoxin Involving Hypoxia-Inducible Factor-1 in a Cell-Type-Specific Manner. J. Neurosci. 2010, 30, 9621–9630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elks, P.; Van Eeden, F.J.; Dixon, G.; Wang, X.; Reyes-Aldasoro, C.C.; Ingham, P.W.; Whyte, M.K.B.; Walmsley, S.; Renshaw, S.A. Activation of hypoxia-inducible factor-1α (Hif-1α) delays inflammation resolution by reducing neutrophil apoptosis and reverse migration in a zebrafish inflammation model. Blood 2011, 118, 712–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Rooijen, E.; Santhakumar, K.; Logister, I.; Voest, E.; Schulte-Merker, S.; Giles, R.; van Eeden, F. A Zebrafish Model for VHL and Hypoxia Signaling. Apoptosis 2011, 105, 163–190. [Google Scholar] [CrossRef]
- Schoonheim, P.J.; Chatzopoulou, A.; Schaaf, M.J. The zebrafish as an in vivo model system for glucocorticoid resistance. Steroids 2010, 75, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Schaaf, M.; Chatzopoulou, A.; Spaink, H. The zebrafish as a model system for glucocorticoid receptor research. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 153, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Hammond, F.R.; Lewis, A.; Elks, P.M. If it’s not one thing, HIF’s another: Immunoregulation by hypoxia inducible factors in disease. FEBS J. 2020, 287, 3907–3916. [Google Scholar] [CrossRef] [PubMed]
- Ziv, L.; Muto, A.; Schoonheim, P.J.; Meijsing, S.H.; Strasser, D.; Ingraham, H.A.; Schaaf, M.J.M.; Yamamoto, K.R.; Baier, H. An affective disorder in zebrafish with mutation of the glucocorticoid receptor. Mol. Psychiatry 2012, 18, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.L.; Holmes, G.R.; Bojarczuk, A.N.; Burgon, J.; Loynes, C.A.; Chimen, M.; Sawtell, A.K.; Hamza, B.; Willson, J.; Walmsley, S.R.; et al. A Zebrafish Compound Screen Reveals Modulation of Neutrophil Reverse Migration as an Anti-Inflammatory Mechanism. Sci. Transl. Med. 2014, 6, 225ra29. [Google Scholar] [CrossRef] [Green Version]
- Hruscha, A.; Krawitz, P.; Rechenberg, A.; Heinrich, V.; Hecht, J.; Haass, C.; Schmid, B. Efficient CRISPR/Cas9 genome editing with low off-target effects in zebrafish. Development 2013, 140, 4982–4987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varshney, G.K.; Pei, W.; LaFave, M.C.; Idol, J.; Xu, L.; Gallardo, V.; Carrington, B.; Bishop, K.; Jones, M.; Li, M.; et al. High-throughput gene targeting and phenotyping in zebrafish using CRISPR/Cas9. Genome Res. 2015, 25, 1030–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Nesan, D.; Vijayan, M.M. Maternal Cortisol Mediates Hypothalamus-Pituitary-Interrenal Axis Development in Zebrafish. Sci. Rep. 2016, 6, 22582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsigos, C.; Chrousos, G.P. Hypothalamic–pituitary–adrenal axis, neuroendocrine factors and stress. J. Psychosom. Res. 2002, 53, 865–871. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, F.; Braunbeck, T. Alterations along the Hypothalamic-Pituitary-Thyroid Axis of the Zebrafish (Danio rerio) after Exposure to Propylthiouracil. J. Thyroid. Res. 2011, 2011, 376243. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Nunez, V. Identification of two proopiomelanocortin genes in zebrafish (Danio rerio). Mol. Brain Res. 2003, 120, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hansen, I.A.; To, T.T.; Wortmann, S.; Burmester, T.; Winkler, C.; Meyer, S.R.; Neuner, C.; Fassnacht, M.; Allolio, B. The pro-opiomelanocortin gene of the zebrafish (Danio rerio). Biochem. Biophys. Res. Commun. 2003, 303, 1121–1128. [Google Scholar] [CrossRef]
- To, T.T.; Hahner, S.; Nica, G.; Rohr, K.B.; Hammerschmidt, M.; Winkler, C.; Allolio, B. Pituitary-Interrenal Interaction in Zebrafish Interrenal Organ Development. Mol. Endocrinol. 2007, 21, 472–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagle, M.; Mathur, P.; Guo, S. Corticotropin-Releasing Factor Critical for Zebrafish Camouflage Behavior Is Regulated by Light and Sensitive to Ethanol. J. Neurosci. 2011, 31, 214–224. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Lu, Y.; Zhai, G.; Huang, J.; Shang, G.; Lou, Q.; Li, D.; Jin, X.; He, J.; Du, Z.; et al. Hyperandrogenism in POMCa-deficient zebrafish enhances somatic growth without increasing adiposity. J. Mol. Cell Biol. 2019, 12, 291–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mommsen, T.P.; Vijayan, M.M.; Moon, T.W. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fish. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Nesan, D.; Vijayan, M.M. Role of glucocorticoid in developmental programming: Evidence from zebrafish. Gen. Comp. Endocrinol. 2013, 181, 35–44. [Google Scholar] [CrossRef]
- Spiga, F.; Walker, J.J.; Terry, J.R.; Lightman, S.L. HPA Axis-Rhythms. Compr. Physiol. 2014, 4, 1273–1298. [Google Scholar] [CrossRef]
- Ramamoorthy, S.; Cidlowski, J.A. Corticosteroids. Mechanisms of Action in Health and Disease. Rheum. Dis. Clin. N. Am. 2016, 42, 15–31. [Google Scholar] [CrossRef] [Green Version]
- Gjerstad, J.K.; Lightman, S.L.; Spiga, F. Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility. Stress 2018, 21, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Dallman, M.F.; Akana, S.F.; Cascio, C.S.; Darlington, D.N.; Jacobson, L.; Levin, N. Regulation of ACTH Secretion: Variations on a Theme of B. Recent Prog. Horm. Res. 1987, 43, 113–173. [Google Scholar] [CrossRef] [PubMed]
- Drouin, J.; Trifiro, M.A.; Plante, R.K.; Nemer, M.; Eriksson, P.; Wrange, O. Glucocorticoid receptor binding to a specific DNA sequence is required for hormone-dependent repression of pro-opiomelanocortin gene transcription. Mol. Cell. Biol. 1989, 9, 5305–5314. [Google Scholar] [CrossRef]
- Ulrich-Lai, Y.; Herman, J. Neural regulation of endocrine and autonomic stress responses. Nat. Rev. Neurosci. 2009, 10, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Nicolaides, N.C.; Galata, Z.; Kino, T.; Chrousos, G.P.; Charmandari, E. The human glucocorticoid receptor: Molecular basis of biologic function. Steroids 2010, 75, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Nicolaides, N.C.; Kyratzi, E.; Lamprokostopoulou, A.; Chrousos, G.P.; Charmandari, E. Stress, the Stress System and the Role of Glucocorticoids. Neuroimmunomodulation 2015, 22, 6–19. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schütz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The nuclear receptor superfamily: The second decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef] [Green Version]
- De Kloet, E.R.; Joels, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef]
- Schoneveld, O.J.; Gaemers, I.C.; Lamers, W.H. Mechanisms of glucocorticoid signalling. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 2004, 1680, 114–128. [Google Scholar] [CrossRef]
- Oakley, R.H.; Cidlowski, J.A. The Biology of the Glucocorticoid Receptor: New Signaling Mechanism in Health and Disease. J. Allergy Clin. Inmunol. 2013, 132, 1033–1044. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.M. The steroid and thyroid hormone receptor superfamily. Science 1988, 240, 889–895. [Google Scholar] [CrossRef]
- Kumar, R.; Thompson, E.B. Gene regulation by the glucocorticoid receptor: Structure:function relationship. J. Steroid Biochem. Mol. Biol. 2005, 94, 383–394. [Google Scholar] [CrossRef]
- Glass, C.K.; Rose, D.W.; Rosenfeld, M.G. Nuclear receptor coactivators. Curr. Opin. Cell Biol. 1997, 9, 222–232. [Google Scholar] [CrossRef]
- Beck, I.; Berghe, W.V.; Vermeulen, L.; Yamamoto, K.R.; Haegeman, G.; De Bosscher, K. Crosstalk in Inflammation: The Interplay of Glucocorticoid Receptor-Based Mechanisms and Kinases and Phosphatases. Endocr. Rev. 2009, 30, 830–882. [Google Scholar] [CrossRef]
- Bledsoe, R.K.; Montana, V.G.; Stanley, T.B.; Delves, C.J.; Apolito, C.J.; McKee, D.D.; Consler, T.G.; Parks, D.J.; Stewart, E.L.; Willson, T.M.; et al. Crystal Structure of the Glucocorticoid Receptor Ligand Binding Domain Reveals a Novel Mode of Receptor Dimerization and Coactivator Recognition. Cell 2002, 110, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Yudt, M.R.; Cidlowski, J.A. Molecular Identification and Characterization of A and B Forms of the Glucocorticoid Receptor. Mol. Endocrinol. 2001, 15, 1093–1103. [Google Scholar] [CrossRef]
- Yudt, M.R.; Cidlowski, J. The Glucocorticoid Receptor: Coding a Diversity of Proteins and Responses through a Single Gene. Mol. Endocrinol. 2002, 16, 1719–1726. [Google Scholar] [CrossRef] [Green Version]
- Oakley, R.H.; Cidlowski, J.A. Cellular Processing of the Glucocorticoid Receptor Gene and Protein: New Mechanisms for Generating Tissue-specific Actions of Glucocorticoids. J. Biol. Chem. 2011, 286, 3177–3184. [Google Scholar] [CrossRef] [Green Version]
- Merkulov, V.M.; Merkulova, T.I. Glucocorticoid receptor isoforms generated by alternative splicing and alternative translation initiation. Russ. J. Genet. Appl. Res. 2012, 2, 205–213. [Google Scholar] [CrossRef]
- Rafacho, A.; Ortsäter, H.; Nadal, A.; Quesada, I. Glucocorticoid treatment and endocrine pancreas function: Implications for glucose homeostasis, insulin resistance and diabetes. J. Endocrinol. 2014, 223, R49–R62. [Google Scholar] [CrossRef]
- Marchi, D. Bidirectional Crosstalk between Hypoxia-Inducible Factor and Glucocorticoid Signalling in Zebrafish Larvae. University of Sheffield. 2020. Available online: https://etheses.whiterose.ac.uk/28224/ (accessed on 2 December 2021).
- Encío, I.J.; Detera-Wadleigh, S.D. The genomic structure of the human glucocorticoid receptor. J. Biol. Chem. 1991, 266, 7182–7188. [Google Scholar] [CrossRef]
- Yudt, M.R.; Jewell, C.M.; Bienstock, R.; Cidlowski, J.A. Molecular Origins for the Dominant Negative Function of Human Glucocorticoid Receptor Beta. Mol. Cell. Biol. 2003, 23, 4319–4330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oakley, R.H.; Sar, M.; Cidlowski, J.A. The Human Glucocorticoid Receptor β Isoform: Expression, biochemical properties, and putative function. J. Biol. Chem. 1996, 271, 9550–9559. [Google Scholar] [CrossRef] [Green Version]
- Bamberger, C.M.; Bamberger, A.M.; de Castro, M.; Chrousos, G.P. Glucocorticoid receptor beta, a potential endogenous inhibitor of glucocorticoid action in humans. J. Clin. Investig. 1995, 95, 2435–2441. [Google Scholar] [CrossRef] [Green Version]
- Oakley, R.H.; Jewell, C.M.; Yudt, M.R.; Bofetiado, D.M.; Cidlowski, J. The Dominant Negative Activity of the Human Glucocorticoid Receptor β Isoform. Specificity and mechanisms of action. J. Biol. Chem. 1999, 274, 27857–27866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charmandari, E.; Chrousos, G.P.; Ichijo, T.; Bhattacharyya, N.; Vottero, A.; Souvatzoglou, E.; Kino, T. The Human Glucocorticoid Receptor (hGR) β Isoform Suppresses the Transcriptional Activity of hGRα by Interfering with Formation of Active Coactivator Complexes. Mol. Endocrinol. 2005, 19, 52–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauk, P.J.; Goleva, E.; Strickland, I.; Vottero, A.; Chrousos, G.P.; Kisich, K.O.; Leung, D.Y.M. Increased Glucocorticoid ReceptorβExpression Converts Mouse Hybridoma Cells to a Corticosteroid-Insensitive Phenotype. Am. J. Respir. Cell Mol. Biol. 2002, 27, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Strickland, I.; Kisich, K.; Hauk, P.J.; Vottero, A.; Chrousos, G.P.; Klemm, D.J.; Leung, D.Y. High Constitutive Glucocorticoid Receptor β in Human Neutrophils Enables Them to Reduce Their Spontaneous Rate of Cell Death in Response to Corticosteroids. J. Exp. Med. 2001, 193, 585–594. [Google Scholar] [CrossRef] [Green Version]
- Timmermans, S.; Souffriau, J.; Libert, C. A General Introduction to Glucocorticoid Biology. Front. Immunol. 2019, 10, 1545. [Google Scholar] [CrossRef] [Green Version]
- Lewis-Tuffin, L.J.; Jewell, C.M.; Bienstock, R.; Collins, J.B.; Cidlowski, J. Human Glucocorticoid Receptor β Binds RU-486 and Is TranscriptionallyActive. Mol. Cell. Biol. 2007, 27, 2266–2282. [Google Scholar] [CrossRef] [Green Version]
- Kino, T.; Manoli, I.; Kelkar, S.; Wang, Y.; Su, Y.A.; Chrousos, G.P. Glucocorticoid receptor (GR) β has intrinsic, GRα-independent transcriptional activity. Biochem. Biophys. Res. Commun. 2009, 381, 671–675. [Google Scholar] [CrossRef] [Green Version]
- Honda, M.; Orii, F.; Ayabe, T.; Imai, S.; Ashida, T.; Obara, T.; Kohgo, Y. Expression of glucocorticoid receptor β in lymphocytes of patients with glucocorticoid-resistant ulcerative colitis. Gastroenterology 2000, 118, 859–866. [Google Scholar] [CrossRef]
- Shahidi, H.; Vottero, A.; Stratakis, C.A.; Taymans, S.E.; Karl, M.; Longui, C.A.; Chrousos, G.P.; Daughaday, W.H.; Gregory, S.A.; Plate, J.M. Imbalanced Expression of the Glucocorticoid Receptor Isoforms in Cultured Lymphocytes from a Patient with Systemic Glucocorticoid Resistance and Chronic Lymphocytic Leukemia. Biochem. Biophys. Res. Commun. 1999, 254, 559–565. [Google Scholar] [CrossRef]
- Bergeron, C.; Fukakusa, M.; Olivenstein, R.; Lemiere, C.; Shannon, J.; Ernst, P.; Martin, J.G.; Hamid, Q. Increased glucocorticoid receptor–β expression, but not decreased histone deacetylase 2, in severe asthma. J. Allergy Clin. Immunol. 2006, 117, 703–705. [Google Scholar] [CrossRef]
- Hamid, Q.A.; Wenzel, S.E.; Hauk, P.J.; Tsicopoulos, A.; Wallaert, B.; Lafitte, J.-J.; Chrousos, G.P.; Szefler, S.J.; Leung, D.Y.M. Increased Glucocorticoid Receptor β in Airway Cells of Glucocorticoid-insensitive Asthma. Am. J. Respir. Crit. Care Med. 1999, 159, 1600–1604. [Google Scholar] [CrossRef]
- Goleva, E.; Li, L.-B.; Eves, P.T.; Strand, M.; Martin, R.J.; Leung, D.Y.M. Increased Glucocorticoid Receptor β Alters Steroid Response in Glucocorticoid-insensitive Asthma. Am. J. Respir. Crit. Care Med. 2006, 173, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Leung, D.Y.; Hamid, Q.; Vottero, A.; Szefler, S.J.; Surs, W.; Minshall, E.; Chrousos, G.P.; Klemm, D.J. Association of Glucocorticoid Insensitivity with Increased Expression of Glucocorticoid Receptor β. J. Exp. Med. 1997, 186, 1567–1574. [Google Scholar] [CrossRef]
- Chatzopoulou, A.; Roy, U.; Meijer, A.H.; Alia, A.; Spaink, H.; Schaaf, M.J.M. Transcriptional and Metabolic Effects of Glucocorticoid Receptor α and β Signaling in Zebrafish. Endocrinology 2015, 156, 1757–1769. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Ramírez, P.; Tliba, O. Glucocorticoid Receptor β (GRβ): Beyond Its Dominant-Negative Function. Int. J. Mol. Sci. 2021, 22, 3649. [Google Scholar] [CrossRef]
- Holownia, A.; Mroz, R.M.; Kolodziejczyk, A.; Chyczewska, E.; Braszko, J.J. Increased FKBP51 in induced sputum cells of chronic obstructive pulmonary disease patients after therapy. Eur. J. Med. Res. 2009, 14 (Suppl. 4), 108–111. [Google Scholar] [CrossRef] [Green Version]
- Storer, C.L.; Dickey, C.A.; Galigniana, M.D.; Rein, T.; Cox, M.B. FKBP51 and FKBP52 in signaling and disease. Trends Endocrinol. Metab. 2011, 22, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Woodruff, P.G.; Boushey, H.A.; Dolganov, G.M.; Barker, C.S.; Yang, J.; Donnelly, S.; Ellwanger, A.; Sidhu, S.S.; Dao-Pick, T.P.; Pantoja, C.; et al. Genome-wide profiling identifies epithelial cell genes associated with asthma and with treatment response to corticosteroids. Proc. Natl. Acad. Sci. USA 2007, 104, 15858–15863. [Google Scholar] [CrossRef] [Green Version]
- Binder, E.B. The role of FKBP5, a co-chaperone of the glucocorticoid receptor in the pathogenesis and therapy of affective and anxiety disorders. Psychoneuroendocrinology 2009, 34, S186–S195. [Google Scholar] [CrossRef]
- Liberman, A.C.; Budziñski, M.L.; Sokn, C.; Gobbini, R.P.; Steininger, A.; Arzt, E. Regulatory and Mechanistic Actions of Glucocorticoids on T and Inflammatory Cells. Front. Endocrinol. 2018, 9, 235. [Google Scholar] [CrossRef]
- Vandevyver, S.; Dejager, L.; Tuckermann, J.; Libert, C. New Insights into the Anti-inflammatory Mechanisms of Glucocorticoids: An Emerging Role for Glucocorticoid-Receptor-Mediated Transactivation. Endocrinology 2013, 154, 993–1007. [Google Scholar] [CrossRef] [Green Version]
- Presman, D.M.; Hager, G.L. More than meets the dimer: What is the quaternary structure of the glucocorticoid receptor? Transcription 2016, 8, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Reichardt, H.M.; Kaestner, K.H.; Tuckermann, J.; Kretz, O.; Wessely, O.; Bock, R.; Gass, P.; Schmid, W.; Herrlich, P.; Angel, P.; et al. DNA Binding of the Glucocorticoid Receptor Is Not Essential for Survival. Cell 1998, 93, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Prager, E.M.; Johnson, L.R. Stress at the Synapse: Signal Transduction Mechanisms of Adrenal Steroids at Neuronal Membranes. Sci. Signal. 2009, 2, re5. [Google Scholar] [CrossRef]
- Groeneweg, F.L.; Karst, H.; de Kloet, R.; Joels, M. Rapid non-genomic effects of corticosteroids and their role in the central stress response. J. Endocrinol. 2011, 209, 153–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlenhaut, H.; Barish, G.D.; Yu, R.T.; Downes, M.; Karunasiri, M.; Liddle, C.; Schwalie, P.C.; Hübner, N.; Evans, R.M. Insights into Negative Regulation by the Glucocorticoid Receptor from Genome-wide Profiling of Inflammatory Cistromes. Mol. Cell 2012, 49, 158–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guenther, M.G.; Barak, O.; Lazar, M.A. The SMRT and N-CoR Corepressors Are Activating Cofactors for Histone Deacetylase 3. Mol. Cell. Biol. 2001, 21, 6091–6101. [Google Scholar] [CrossRef] [Green Version]
- Surjit, M.; Ganti, K.P.; Mukherji, A.; Ye, T.; Hua, G.; Metzger, D.; Li, M.; Chambon, P. Widespread Negative Response Elements Mediate Direct Repression by Agonist- Liganded Glucocorticoid Receptor. Cell 2011, 145, 224–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramamoorthy, S.; Cidlowski, J.A. Exploring the Molecular Mechanisms of Glucocorticoid Receptor Action from Sensitivity to Resistance. Endocr. Dev. 2013, 24, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Escoter-Torres, L.; Caratti, G.; Mechtidou, A.; Tuckermann, J.; Uhlenhaut, N.H.; Vettorazzi, S. Fighting the Fire: Mechanisms of Inflammatory Gene Regulation by the Glucocorticoid Receptor. Front. Immunol. 2019, 10, 1859. [Google Scholar] [CrossRef]
- Oh, K.-S.; Patel, H.; Gottschalk, R.A.; Lee, W.S.; Baek, S.; Fraser, I.; Hager, G.L.; Sung, M.-H. Anti-Inflammatory Chromatinscape Suggests Alternative Mechanisms of Glucocorticoid Receptor Action. Immunity 2017, 47, 298–309.e5. [Google Scholar] [CrossRef]
- Ratman, D.; Berghe, W.V.; Dejager, L.; Libert, C.; Tavernier, J.; Beck, I.; De Bosscher, K. How glucocorticoid receptors modulate the activity of other transcription factors: A scope beyond tethering. Mol. Cell. Endocrinol. 2013, 380, 41–54. [Google Scholar] [CrossRef]
- Baudrand, R.; Pojoga, L.; Romero, J.R. Chapter 10—Aldosterone’s Mechanism of Action: Genomic and Nongenomic Signaling. In Textbook of Nephro-Endocrinology, 2nd ed.; Singh, A.K., Williams, G.H., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 173–188. [Google Scholar] [CrossRef]
- Reul, J.M.H.M.; de Kloet, R. Two Receptor Systems for Corticosterone in Rat Brain: Microdistribution and Differential Occupation. Endocrinology 1985, 117, 2505–2511. [Google Scholar] [CrossRef]
- Reul, J.M.; Collins, A.; Saliba, R.S.; Mifsud, K.; Carter, S.D.; Gutierrez-Mecinas, M.; Qian, X.; Linthorst, A.C. Glucocorticoids, epigenetic control and stress resilience. Neurobiol. Stress 2014, 1, 44–59. [Google Scholar] [CrossRef] [Green Version]
- Grossmann, C.; Gekle, M. Non-classical actions of the mineralocorticoid receptor: Misuse of EGF receptors? Mol. Cell. Endocrinol. 2007, 277, 6–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sireeni, J.; Bakker, N.; Jaikumar, G.; Obdam, D.; Slabbekoorn, H.; Tudorache, C.; Schaaf, M. Profound effects of glucocorticoid resistance on anxiety-related behavior in zebrafish adults but not in larvae. Gen. Comp. Endocrinol. 2020, 292, 113461. [Google Scholar] [CrossRef]
- Müller, M.B.; Holsboer, F.; Keck, M.E. Genetic modification of corticosteroid receptor signalling: Novel insights into pathophysiology and treatment strategies of human affective disorders. Neuropeptides 2002, 36, 117–131. [Google Scholar] [CrossRef]
- Van Eekelen, J.; Bohn, M.; De Kloet, E. Postnatal ontogeny of mineralocorticoid and glucocorticoid receptor gene expression in regions of the rat tel- and diencephalon. Dev. Brain Res. 1991, 61, 33–43. [Google Scholar] [CrossRef]
- Hartmann, J.; Bajaj, T.; Klengel, C.; Chatzinakos, C.; Ebert, T.; Dedic, N.; McCullough, K.M.; Lardenoije, R.; Joëls, M.; Meijer, O.C.; et al. Mineralocorticoid receptors dampen glucocorticoid receptor sensitivity to stress via regulation of FKBP5. Cell Rep. 2021, 35, 109185. [Google Scholar] [CrossRef]
- De Kloet, E.; Meijer, O.; de Nicola, A.; de Rijk, R.; Joëls, M. Importance of the brain corticosteroid receptor balance in metaplasticity, cognitive performance and neuro-inflammation. Front. Neuroendocrinol. 2018, 49, 124–145. [Google Scholar] [CrossRef]
- Lee, H.B.; Schwab, T.L.; Sigafoos, A.N.; Gauerke, J.L.; Ii, R.G.K.; Serres, M.R.; Jacobs, D.C.; Cotter, R.P.; Das, B.; Petersen, M.O.; et al. Novel zebrafish behavioral assay to identify modifiers of the rapid, nongenomic stress response. Genes Brain Behav. 2018, 18, e12549. [Google Scholar] [CrossRef]
- Marchi, D.; Santhakumar, K.; Markham, E.; Li, N.; Storbeck, K.-H.; Krone, N.; Cunliffe, V.T.; van Eeden, F.J.M. Bidirectional crosstalk between Hypoxia-Inducible Factor and glucocorticoid signalling in zebrafish larvae. PLoS Genet. 2020, 16, e1008757. [Google Scholar] [CrossRef]
- Ince, L.M.; Weber, J.; Scheiermann, C. Control of Leukocyte Trafficking by Stress-Associated Hormones. Front. Immunol. 2019, 9, 3143. [Google Scholar] [CrossRef] [Green Version]
- Coutinho, A.E.; Chapman, K.E. The anti-inflammatory and immunosuppressive effects of glucocorticoids, recent developments and mechanistic insights. Mol. Cell. Endocrinol. 2011, 335, 2–13. [Google Scholar] [CrossRef]
- Schmidt, S.; Rainer, J.; Ploner, C.; Presul, E.; Riml, S.; Kofler, R. Glucocorticoid-induced apoptosis and glucocorticoid resistance: Molecular mechanisms and clinical relevance. Cell Death Differ. 2004, 11, S45–S55. [Google Scholar] [CrossRef] [Green Version]
- Planey, S.L.; Litwack, G. Glucocorticoid-Induced Apoptosis in Lymphocytes. Biochem. Biophys. Res. Commun. 2000, 279, 307–312. [Google Scholar] [CrossRef]
- Ashwell, J.D.; Lu, F.W.M.; Vacchio, M.S. Glucocorticoids in T Cell Development and Function. Annu. Rev. Immunol. 2000, 18, 309–345. [Google Scholar] [CrossRef]
- Herold, M.J.; McPherson, K.G.; Reichardt, H.M. Glucocorticoids in T cell apoptosis and function. Cell. Mol. Life Sci. 2005, 63, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McColl, A.; Bournazos, S.; Franz, S.; Perretti, M.; Morgan, B.P.; Haslett, C.; Dransfield, I. Glucocorticoids Induce Protein S-Dependent Phagocytosis of Apoptotic Neutrophils by Human Macrophages. J. Immunol. 2009, 183, 2167–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, A.H.; Spencer, R.L.; Stein, M.; McEwen, B.S. Adrenal steroid receptor binding in spleen and thymus after stress or dexamethasone. Am. J. Physiol. Metab. 1990, 259, E405. [Google Scholar] [CrossRef] [PubMed]
- Barish, G.D.; Downes, M.; Alaynick, W.A.; Yu, R.T.; Ocampo, C.B.; Bookout, A.L.; Mangelsdorf, D.J.; Evans, R.M. A Nuclear Receptor Atlas: Macrophage Activation. Mol. Endocrinol. 2005, 19, 2466–2477. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.-Y.; Müller, N.; Herold, M.J.; Brandt, J.V.D.; Reichardt, H.M. Glucocorticoids exert opposing effects on macrophage function dependent on their concentration. Immunology 2007, 122, 47–53. [Google Scholar] [CrossRef]
- Harizi, H.; Mormede, P.; Corcuff, J. Inter-strain differences in glucocorticoid and mineralocorticoid effects on macrophage and lymphocyte functions in mice. J. Neuroimmunol. 2008, 204, 38–42. [Google Scholar] [CrossRef]
- Bene, N.C.; Alcaide, P.; Wortis, H.H.; Jaffe, I.Z. Mineralocorticoid receptors in immune cells: Emerging role in cardiovascular disease. Steroids 2014, 91, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Durango, N.; Vecchiola, A.; Gonzalez-Gomez, L.M.; Simon, F.; Riedel, C.; Fardella, C.E.; Kalergis, A.M. Modulation of Immunity and Inflammation by the Mineralocorticoid Receptor and Aldosterone. BioMed Res. Int. 2015, 2015, 652738. [Google Scholar] [CrossRef] [Green Version]
- Van Der Heijden, C.D.C.C.; Deinum, J.; Joosten, L.A.B.; Netea, M.G.; Riksen, N.P. The mineralocorticoid receptor as a modulator of innate immunity and atherosclerosis. Cardiovasc. Res. 2018, 114, 944–953. [Google Scholar] [CrossRef] [Green Version]
- Vandevyver, S.; Dejager, L.; Libert, C. On the Trail of the Glucocorticoid Receptor: Into the Nucleus and Back. Traffic 2011, 13, 364–374. [Google Scholar] [CrossRef]
- Ronchetti, S.; Migliorati, G.; Riccardi, C. GILZ as a Mediator of the Anti-Inflammatory Effects of Glucocorticoids. Front. Endocrinol. 2015, 6, 170. [Google Scholar] [CrossRef] [Green Version]
- Scheinman, R.I.; Gualberto, A.; Jewell, C.M.; Cidlowski, J.A.; Baldwin, A.S. Characterization of mechanisms involved in transrepression of NF-kappa B by activated glucocorticoid receptors. Mol. Cell. Biol. 1995, 15, 943–953. [Google Scholar] [CrossRef] [Green Version]
- Escoter-Torres, L.; Greulich, F.; Quagliarini, F.; Wierer, M.; Uhlenhaut, N.H. Anti-inflammatory functions of the glucocorticoid receptor require DNA binding. Nucleic Acids Res. 2020, 48, 8393–8407. [Google Scholar] [CrossRef]
- Stellato, C. Post-transcriptional and Nongenomic Effects of Glucocorticoids. Proc. Am. Thorac. Soc. 2004, 1, 255–263. [Google Scholar] [CrossRef]
- De Bosscher, K.; Van Craenenbroeck, K.; Meijer, O.C.; Haegeman, G. Selective transrepression versus transactivation mechanisms by glucocorticoid receptor modulators in stress and immune systems. Eur. J. Pharmacol. 2008, 583, 290–302. [Google Scholar] [CrossRef] [PubMed]
- De Bosscher, K.; Haegeman, G.; Elewaut, D. Targeting inflammation using selective glucocorticoid receptor modulators. Curr. Opin. Pharmacol. 2010, 10, 497–504. [Google Scholar] [CrossRef]
- Davies, T.H.; Ning, Y.-M.; Sánchez, E.R. Differential Control of Glucocorticoid Receptor Hormone-Binding Function by Tetratricopeptide Repeat (TPR) Proteins and the Immunosuppressive Ligand FK506. Biochemistry 2005, 44, 2030–2038. [Google Scholar] [CrossRef] [Green Version]
- Liberman, A.C.; Druker, J.; Perone, M.J.; Arzt, E. Glucocorticoids in the regulation of transcription factors that control cytokine synthesis. Cytokine Growth Factor Rev. 2007, 18, 45–56. [Google Scholar] [CrossRef]
- Pascual, G.; Glass, C.K. Nuclear receptors versus inflammation: Mechanisms of transrepression. Trends Endocrinol. Metab. 2006, 17, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Limbourg, F.P.; Liao, J.K. Nontranscriptional actions of the glucocorticoid receptor. J. Mol. Med. 2003, 81, 168–174. [Google Scholar] [CrossRef] [Green Version]
- Liberman, A.C.; Refojo, D.; Druker, J.; Toscano, M.; Rein, T.; Holsboer, F.; Arzt, E. The activated glucocorticoid receptor inhibits the transcription factor T-bet by direct protein-protein interaction. FASEB J. 2007, 21, 1177–1188. [Google Scholar] [CrossRef]
- Vayssière, B.M.; Dupont, S.; Choquart, A.; Petit, F.; Garcia, T.; Marchandeau, C.; Gronemeyer, H.; Resche-Rigon, M. Synthetic Glucocorticoids That Dissociate Transactivation and AP-1 Transrepression Exhibit Antiinflammatory Activity in Vivo. Mol. Endocrinol. 1997, 11, 1245–1255. [Google Scholar] [CrossRef] [Green Version]
- Belvisi, M.G.; Wicks, S.L.; Battram, C.H.; Bottoms, S.E.W.; Redford, J.E.; Woodman, P.; Brown, T.J.; Webber, S.E.; Foster, M.L. Therapeutic Benefit of a Dissociated Glucocorticoid and the Relevance of In Vitro Separation of Transrepression from Transactivation Activity. J. Immunol. 2001, 166, 1975–1982. [Google Scholar] [CrossRef] [Green Version]
- Schacke, H.; Schottelius, A.; Docke, W.-D.; Strehlke, P.; Jaroch, S.; Schmees, N.; Rehwinkel, H.; Hennekes, H.; Asadullah, K. Dissociation of transactivation from transrepression by a selective glucocorticoid receptor agonist leads to separation of therapeutic effects from side effects. Proc. Natl. Acad. Sci. USA 2003, 101, 227–232. [Google Scholar] [CrossRef] [Green Version]
- De Bosscher, K.; Beck, I.M.; Dejager, L.; Bougarne, N.; Gaigneaux, A.; Chateauvieux, S.; Ratman, D.; Bracke, M.; Tavernier, J.; Berghe, W.V.; et al. Selective modulation of the glucocorticoid receptor can distinguish between transrepression of NF-κB and AP-1. Cell. Mol. Life Sci. 2013, 71, 143–163. [Google Scholar] [CrossRef] [Green Version]
- De Bosscher, K.; Berghe, W.V.; Beck, I.; Van Molle, W.; Hennuyer, N.; Hapgood, J.; Libert, C.; Staels, B.; Louw, A.; Haegeman, G. A fully dissociated compound of plant origin for inflammatory gene repression. Proc. Natl. Acad. Sci. USA 2005, 102, 15827–15832. [Google Scholar] [CrossRef] [Green Version]
- Duque, E.D.A.; Munhoz, C.D. The Pro-inflammatory Effects of Glucocorticoids in the Brain. Front. Endocrinol. 2016, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Langlais, D.; Couture, C.; Balsalobre, A.; Drouin, J. Regulatory Network Analyses Reveal Genome-Wide Potentiation of LIF Signaling by Glucocorticoids and Define an Innate Cell Defense Response. PLoS Genet. 2008, 4, e1000224. [Google Scholar] [CrossRef]
- Langlais, D.; Couture, C.; Balsalobre, A.; Drouin, J. The Stat3/GR Interaction Code: Predictive Value of Direct/Indirect DNA Recruitment for Transcription Outcome. Mol. Cell 2012, 47, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Dittrich, A.; Khouri, C.; Sackett, S.D.; Ehlting, C.; Böhmer, O.; Albrecht, U.; Bode, J.G.; Trautwein, C.; Schaper, F. Glucocorticoids increase interleukin-6-dependent gene induction by interfering with the expression of the suppressor of cytokine signaling 3 feedback inhibitor. Hepatology 2011, 55, 256–266. [Google Scholar] [CrossRef]
- Xie, Y.; Tolmeijer, S.; Oskam, J.M.; Tonkens, T.; Meijer, A.H.; Schaaf, M.J.M. Glucocorticoids inhibit macrophage differentiation towards a pro-inflammatory phenotype upon wounding without affecting their migration. Dis. Model. Mech. 2019, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bos, R.V.D.; Cromwijk, S.; Tschigg, K.; Althuizen, J.; Zethof, J.; Whelan, R.; Flik, G.; Schaaf, M. Early Life Glucocorticoid Exposure Modulates Immune Function in Zebrafish (Danio rerio) Larvae. Front. Immunol. 2020, 11, 727. [Google Scholar] [CrossRef] [PubMed]
- John, S.; Sabo, P.J.; Thurman, R.E.; Sung, M.-H.; Biddie, S.; Johnson, T.A.; Hager, G.L.; Stamatoyannopoulos, J.A. Chromatin accessibility pre-determines glucocorticoid receptor binding patterns. Nat. Genet. 2011, 43, 264–268. [Google Scholar] [CrossRef]
- John, S.; Sabo, P.J.; Johnson, T.A.; Sung, M.-H.; Biddie, S.C.; Lightman, S.L.; Voss, T.C.; Davis, S.R.; Meltzer, P.S.; Stamatoyannopoulos, J.A.; et al. Interaction of the Glucocorticoid Receptor with the Chromatin Landscape. Mol. Cell 2008, 29, 611–624. [Google Scholar] [CrossRef]
- Ito, K.; Barnes, P.J.; Adcock, I.M. Glucocorticoid Receptor Recruitment of Histone Deacetylase 2 Inhibits Interleukin-1β-Induced Histone H4 Acetylation on Lysines 8 and 12. Mol. Cell. Biol. 2000, 20, 6891–6903. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Hypoxia. Cross talk between oxygen sensing and the cell cycle machinery. Am. J. Physiol. Physiol. 2011, 301, C550–C552. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. HIF-1 and mechanisms of hypoxia sensing. Curr. Opin. Cell Biol. 2001, 13, 167–171. [Google Scholar] [CrossRef]
- Webb, J.D.; Coleman, M.L.; Pugh, C.W. Hypoxia, hypoxia-inducible factors (HIF), HIF hydroxylases and oxygen sensing. Cell. Mol. Life Sci. 2009, 66, 3539. [Google Scholar] [CrossRef]
- Pollenz, R.S.; Sattler, C.A.; Poland, A. The aryl hydrocarbon receptor and aryl hydrocarbon receptor nuclear translocator protein show distinct subcellular localizations in Hepa 1c1c7 cells by immunofluorescence microscopy. Mol. Pharmacol. 1994, 45, 428–438. [Google Scholar] [PubMed]
- Eguchi, H.; Ikuta, T.; Tachibana, T.; Yoneda, Y.; Kawajiri, K. A Nuclear Localization Signal of Human Aryl Hydrocarbon Receptor Nuclear Translocator/Hypoxia-inducible Factor 1β Is a Novel Bipartite Type Recognized by the Two Components of Nuclear Pore-targeting Complex. J. Biol. Chem. 1997, 272, 17640–17647. [Google Scholar] [CrossRef] [Green Version]
- Iwai, K.; Yamanaka, K.; Kamura, T.; Minato, N.; Conaway, R.C.; Conaway, J.; Klausner, R.D.; Pause, A. Identification of the von Hippel-Lindau tumor-suppressor protein as part of an active E3 ubiquitin ligase complex. Proc. Natl. Acad. Sci. USA 1999, 96, 12436–12441. [Google Scholar] [CrossRef] [Green Version]
- Shay, J.E.; Simon, M.C. Hypoxia-inducible factors: Crosstalk between inflammation and metabolism. Semin. Cell Dev. Biol. 2012, 23, 389–394. [Google Scholar] [CrossRef]
- Siddiq, A.; Aminova, L.R.; Ratan, R.R. Hypoxia Inducible Factor Prolyl 4-Hydroxylase Enzymes: Center Stage in the Battle Against Hypoxia, Metabolic Compromise and Oxidative Stress. Neurochem. Res. 2007, 32, 931–946. [Google Scholar] [CrossRef]
- Pan, Y.; Mansfield, K.D.; Bertozzi, C.C.; Rudenko, V.; Chan, D.A.; Giaccia, A.J.; Simon, M.C. Multiple Factors Affecting Cellular Redox Status and Energy Metabolism Modulate Hypoxia-Inducible Factor Prolyl Hydroxylase Activity In Vivo and In Vitro. Mol. Cell. Biol. 2007, 27, 912–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masson, N.; Singleton, R.S.; Sekirnik, R.; Trudgian, D.C.; Ambrose, L.J.; Miranda, M.X.; Tian, Y.; Kessler, B.M.; Schofield, C.J.; Ratcliffe, P.J. The FIH hydroxylase is a cellular peroxide sensor that modulates HIF transcriptional activity. EMBO Rep. 2012, 13, 251–257. [Google Scholar] [CrossRef]
- Rhee, S.G.; Woo, H.A.; Kil, I.S.; Bae, S.H. Peroxiredoxin Functions as a Peroxidase and a Regulator and Sensor of Local Peroxides. J. Biol. Chem. 2012, 287, 4403–4410. [Google Scholar] [CrossRef] [Green Version]
- Palazon, A.; Goldrath, A.W.; Nizet, V.; Johnson, R.S. HIF Transcription Factors, Inflammation, and Immunity. Immunity 2014, 41, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Oxygen Sensing, Homeostasis, and Disease. N. Engl. J. Med. 2011, 365, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Safronova, O.; Morita, I. Transcriptome Remodeling in Hypoxic Inflammation. J. Dent. Res. 2010, 89, 430–444. [Google Scholar] [CrossRef]
- Bosco, M.C.; Puppo, M.; Blengio, F.; Fraone, T.; Cappello, P.; Giovarelli, M.; Varesio, L. Monocytes and dendritic cells in a hypoxic environment: Spotlights on chemotaxis and migration. Immunobiology 2008, 213, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, C.; Muthana, M.; Lewis, C.E. Hypoxia Regulates Macrophage Functions in Inflammation. J. Immunol. 2005, 175, 6257–6263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, C.; Murdoch, C. Macrophage Responses to Hypoxia: Implications for Tumor Progression and Anti-Cancer Therapies. Am. J. Pathol. 2005, 167, 627–635. [Google Scholar] [CrossRef]
- Safronova, O.; Pluemsampant, S.; Nakahama, K.-I.; Morita, I. Regulation of chemokine gene expression by hypoxia via cooperative activation of NF-κB and histone deacetylase. Int. J. Biochem. Cell Biol. 2009, 41, 2270–2280. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Yin, M.-J.; Gaynor, R.B. IκB Kinase α (IKKα) Regulation of IKKβ Kinase Activity. Mol. Cell. Biol. 2000, 20, 3655–3666. [Google Scholar] [CrossRef]
- Cummins, E.P.; Berra, E.; Comerford, K.M.; Ginouves, A.; Fitzgerald, K.T.; Seeballuck, F.; Godson, C.; Nielsen, J.E.; Moynagh, P.; Pouyssegur, J.; et al. Prolyl hydroxylase-1 negatively regulates IκB kinase-β, giving insight into hypoxia-induced NFκB activity. Proc. Natl. Acad. Sci. USA 2006, 103, 18154–18159. [Google Scholar] [CrossRef] [Green Version]
- Cummins, E.; Comerford, K.M.; Scholz, C.; Bruning, U.; Taylor, C. Hypoxic Regulation of NF-κB Signaling. Methods Enzymol. 2007, 435, 479–492. [Google Scholar] [CrossRef]
- van Uden, P.; Kenneth, N.S.; Rocha, S. Regulation of hypoxia-inducible factor-1α by NF-κB. Biochem. J. 2008, 412, 477–484. [Google Scholar] [CrossRef] [Green Version]
- van Uden, P.; Kenneth, N.S.; Webster, R.; Müller, H.A.; Mudie, S.; Rocha, S. Evolutionary Conserved Regulation of HIF-1β by NF-κB. PLoS Genet. 2011, 7, e1001285. [Google Scholar] [CrossRef] [Green Version]
- Rius, J.; Guma, M.; Schachtrup, C.; Akassoglou, K.; Zinkernagel, A.S.; Nizet, V.; Johnson, R.S.; Haddad, G.G.; Karin, M. NF-κB links innate immunity to the hypoxic response through transcriptional regulation of HIF-1α. Nature 2008, 453, 807–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Ignazio, L.; Bandarra, D.; Rocha, S. NF-κB and HIF crosstalk in immune responses. FEBS J. 2015, 283, 413–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schild, Y.; Mohamed, A.; Wootton, E.J.; Lewis, A.; Elks, P.M. Hif-1alpha stabilisation is protective against infection in zebrafish comorbid models. FEBS J. 2020, 287, 3925–3943. [Google Scholar] [CrossRef] [PubMed]
- Charron, C.E.; Chou, P.-C.; Coutts, D.J.C.; Kumar, V.; To, M.; Akashi, K.; Pinhu, L.; Griffiths, M.; Adcock, I.; Barnes, P.J.; et al. Hypoxia-inducible Factor 1α Induces Corticosteroid-insensitive Inflammation via Reduction of Histone Deacetylase-2 Transcription. J. Biol. Chem. 2009, 284, 36047–36054. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Zhao, J.-J.; Lv, Y.-Y.; Ding, P.-S.; Liu, R.-Y. Hypoxia down-regulates glucocorticoid receptor alpha and attenuates the anti-inflammatory actions of dexamethasone in human alveolar epithelial A549 cells. Life Sci. 2009, 85, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Vanderhaeghen, T.; Timmermans, S.; Watts, D.; Paakinaho, V.; Eggermont, M.; Vandewalle, J.; Wallaeys, C.; Van Wyngene, L.; Van Looveren, K.; Nuyttens, L.; et al. Reprogramming of glucocorticoid receptor function by hypoxia. EMBO Rep. 2021, e53083. [Google Scholar] [CrossRef] [PubMed]
- Elsby, L.M.; Donn, R.; Alourfi, Z.; Green, L.M.; Beaulieu, E.; Ray, D.W. Hypoxia and glucocorticoid signaling converge to regulate macrophage migration inhibitory factor gene expression. Arthritis Rheum. 2009, 60, 2220–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Ma, Y.-Y.; Song, X.-L.; Cai, H.-Y.; Chen, J.-C.; Song, L.-N.; Yang, R.; Lu, J. Upregulations of Glucocorticoid-Induced Leucine Zipper by Hypoxia and Glucocorticoid Inhibit Proinflammatory Cytokines under Hypoxic Conditions in Macrophages. J. Immunol. 2011, 188, 222–229. [Google Scholar] [CrossRef] [Green Version]
- Ayroldi, E.; Riccardi, C. Glucocorticoid-induced leucine zipper (GILZ): A new important mediator of glucocorticoid action. FASEB J. 2009, 23, 3649–3658. [Google Scholar] [CrossRef] [Green Version]
- Cannarile, L.; Delfino, D.V.; Adorisio, S.; Riccardi, C.; Ayroldi, E. Implicating the Role of GILZ in Glucocorticoid Modulation of T-Cell Activation. Front. Immunol. 2019, 10, 1823. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-H.; Choi, E.Y. Macrophages and Inflammation. J. Rheum. Dis. 2018, 25, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Xue, L.; Greisler, H.P. Angiogenic effect of fibroblast growth factor-1 and vascular endothelial growth factor and their synergism in a novel in vitro quantitative fibrin-based 3-dimensional angiogenesis system. Surgery 2002, 132, 259–267. [Google Scholar] [CrossRef]
- Ke, X.; Chen, C.; Song, Y.; Cai, Q.; Li, J.; Tang, Y.; Han, X.; Qu, W.; Chen, A.; Wang, H.; et al. Hypoxia modifies the polarization of macrophages and their inflammatory microenvironment, and inhibits malignant behavior in cancer cells. Oncol. Lett. 2019, 18, 5871–5878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Goodwin, J.E. The Effect of Glucocorticoids on Angiogenesis in the Treatment of Solid Tumors. J. Cell. Signal. 2020, 1, 42–49. [Google Scholar] [CrossRef]
- Bernhardt, W.M.; Câmpean, V.; Kany, S.; Jürgensen, J.-S.; Weidemann, A.; Warnecke, C.; Arend, M.; Klaus, S.; Günzler, V.; Amann, K.; et al. Preconditional Activation of Hypoxia-Inducible Factors Ameliorates Ischemic Acute Renal Failure. J. Am. Soc. Nephrol. 2006, 17, 1970–1978. [Google Scholar] [CrossRef]
- Wan, C.; Gilbert, S.R.; Wang, Y.; Cao, X.; Shen, X.; Ramaswamy, G.; Jacobsen, K.A.; Alaql, Z.S.; Eberhardt, A.W.; Gerstenfeld, L.C.; et al. Activation of the hypoxia-inducible factor-1 pathway accelerates bone regeneration. Proc. Natl. Acad. Sci. USA 2008, 105, 686–691. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 2003, 3, 721–732. [Google Scholar] [CrossRef]
- Semenza, G.L. HIF-1 mediates metabolic responses to intratumoral hypoxia and oncogenic mutations. J. Clin. Investig. 2013, 123, 3664–3671. [Google Scholar] [CrossRef] [Green Version]
- Vettori, A.; Greenald, D.; Wilson, G.K.; Peron, M.; Facchinello, N.; Markham, E.; Sinnakaruppan, M.; Matthews, L.C.; McKeating, J.A.; Argenton, F.; et al. Glucocorticoids promote Von Hippel Lindau degradation and Hif-1α stabilization. Proc. Natl. Acad. Sci. USA 2017, 114, 9948–9953. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.D.; Brearey, S.P.; Imray, C.H.E. High hopes at high altitudes: Pharmacotherapy for acute mountain sickness and high-altitude cerebral and pulmonary oedema. Expert Opin. Pharmacother. 2007, 9, 119–127. [Google Scholar] [CrossRef]
- Dardzinski, B.J.; Smith, S.L.; Towfighi, J.; Williams, G.D.; Vannucci, R.C.; Smith, M.B. Increased Plasma Beta-Hydroxybutyrate, Preserved Cerebral Energy Metabolism, and Amelioration of Brain Damage During Neonatal Hypoxia Ischemia with Dexamethasone Pretreatment. Pediatr. Res. 2000, 48, 248–255. [Google Scholar] [CrossRef] [Green Version]
- Limbourg, F.P.; Huang, Z.; Plumier, J.-C.; Simoncini, T.; Fujioka, M.; Tuckermann, J.; Schutz, G.; Moskowitz, M.A.; Liao, J.K. Rapid nontranscriptional activation of endothelial nitric oxide synthase mediates increased cerebral blood flow and stroke protection by corticosteroids. J. Clin. Investig. 2002, 110, 1729–1738. [Google Scholar] [CrossRef]
- Glanemann, M.; Strenziok, R.; Kuntze, R.; Münchow, S.; Dikopoulos, N.; Lippek, F.; Langrehr, J.M.; Dietel, M.; Neuhaus, P.; Nussler, A.K. Ischemic preconditioning and methylprednisolone both equally reduce hepatic ischemia/reperfusion injury. Surgery 2004, 135, 203–214. [Google Scholar] [CrossRef]
- Tokudome, S.; Sano, M.; Shinmura, K.; Matsuhashi, T.; Morizane, S.; Moriyama, H.; Tamaki, K.; Hayashida, K.; Nakanishi, H.; Yoshikawa, N.; et al. Glucocorticoid protects rodent hearts from ischemia / reperfusion injury by activating lipocalin-type prostaglandin D synthase—Derived PGD 2 biosynthesis. J. Clin. Investig. 2009, 119, 1477–1488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Qiang, Q.; Jiang, Y.; Hu, L.; Ding, X.; Lu, Y.; Hu, G. Effects of hypoxia inducible factor-1α on apoptotic inhibition and glucocorticoid receptor downregulation by dexamethasone in AtT-20 cells. BMC Endocr. Disord. 2015, 15, 24. [Google Scholar] [CrossRef] [Green Version]
- Gaber, T.; Schellmann, S.; Erekul, K.B.; Fangradt, M.; Tykwinska, K.; Hahne, M.; Maschmeyer, P.; Wagegg, M.; Stahn, C.; Kolar, P.; et al. Macrophage Migration Inhibitory Factor Counterregulates Dexamethasone-Mediated Suppression of Hypoxia-Inducible Factor-1α Function and Differentially Influences Human CD4+ T Cell Proliferation under Hypoxia. J. Immunol. 2010, 186, 764–774. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.S.; Lam, I.I.; Clay, H.; Duong, D.N.; Deo, R.C.; Coughlin, S.R. A Rapid Method for Directed Gene Knockout for Screening in G0 Zebrafish. Dev. Cell 2018, 46, 112–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burger, A.; Lindsay, H.; Felker, A.; Hess, C.; Anders, C.; Chiavacci, E.; Zaugg, J.; Weber, L.M.; Catena, R.; Jinek, M.; et al. Maximizing mutagenesis with solubilized CRISPR-Cas9 ribonucleoprotein complexes. Development 2016, 143, 2025–2037. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, L.; Verhoog, N.; Louw, A. Novel role for receptor dimerization in post-translational processing and turnover of the GRα. Sci. Rep. 2018, 8, 14266. [Google Scholar] [CrossRef] [Green Version]
- Pace, T.W.; Hu, F.; Miller, A.H. Cytokine-effects on glucocorticoid receptor function: Relevance to glucocorticoid resistance and the pathophysiology and treatment of major depression. Brain Behav. Immun. 2007, 21, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.; Yu, R.M.K.; Wu, R.S.S.; Kong, R.Y.C. Overexpression and Knockdown of Hypoxia-Inducible Factor 1 Disrupt the Expression of Steroidogenic Enzyme Genes and Early Embryonic Development in Zebrafish. Gene Regul. Syst. Biol. 2017, 11. [Google Scholar] [CrossRef] [Green Version]
- Egg, M.; Köblitz, L.; Hirayama, J.; Schwerte, T.; Folterbauer, C.; Kurz, A.; Fiechtner, B.; Möst, M.; Salvenmoser, W.; Sassone-Corsi, P.; et al. Linking Oxygen to Time: The Bidirectional Interaction Between the Hypoxic Signaling Pathway and the Circadian Clock. Chronobiol. Int. 2013, 30, 510–529. [Google Scholar] [CrossRef] [PubMed]
- Egg, M.; Paulitsch, M.; Ennemoser, Y.; Wüstenhagen, A.; Schwerte, T.; Sandbichler, A.M.; Fiechtner, B.; Köblitz, L.; Prem, C.; Pelster, B. Chronodisruption increases cardiovascular risk in zebrafish via reduced clearance of senescent erythrocytes. Chronobiol. Int. 2014, 31, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Pelster, B.; Egg, M. Multiplicity of Hypoxia-Inducible Transcription Factors and Their Connection to the Circadian Clock in the Zebrafish. Physiol. Biochem. Zool. 2015, 88, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Peek, C.B.; Levine, D.; Cedernaes, J.; Taguchi, A.; Kobayashi, Y.; Tsai, S.J.; Bonar, N.A.; McNulty, M.R.; Ramsey, K.M.; Bass, J. Circadian Clock Interaction with HIF1α Mediates Oxygenic Metabolism and Anaerobic Glycolysis in Skeletal Muscle. Cell Metab. 2016, 25, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N.; Berry, A.; Sauer, C.; Baxter, M.; Donaldson, I.J.; Forbes, K.; Donn, R.; Matthews, L.; Ray, D. Hypoxia regulates GR function through multiple mechanisms involving microRNAs 103 and 107. Mol. Cell. Endocrinol. 2020, 518, 111007. [Google Scholar] [CrossRef]
- Watts, D.; Stein, J.; Meneses, A.; Bechmann, N.; Neuwirth, A.; Kaden, D.; Krüger, A.; Sinha, A.; Alexaki, V.I.; Perez-Rivas, L.G.; et al. HIF1α is a direct regulator of steroidogenesis in the adrenal gland. Cell. Mol. Life Sci. 2021, 78, 3577–3590. [Google Scholar] [CrossRef] [PubMed]
- Rybnikova, E.; Glushchenko, T.; Churilova, A.; Pivina, S.; Samoilov, M. Expression of glucocorticoid and mineralocorticoid receptors in hippocampus of rats exposed to various modes of hypobaric hypoxia: Putative role in hypoxic preconditioning. Brain Res. 2011, 1381, 66–77. [Google Scholar] [CrossRef]
- Robinson, P.C.; Morand, E. Divergent effects of acute versus chronic glucocorticoids in COVID-19. Lancet Rheumatol. 2021, 3, e168–e170. [Google Scholar] [CrossRef]
- van Paassen, J.; Vos, J.S.; Hoekstra, E.M.; Neumann, K.M.I.; Boot, P.C.; Arbous, S.M. Corticosteroid use in COVID-19 patients: A systematic review and meta-analysis on clinical outcomes. Crit. Care 2020, 24, 696. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Glucocorticoid receptor domain structure and translational isoforms. The N-terminal domain (NTD), which is required for ligand-independent gene transactivation, includes a transcriptional activation function region (AF1). The latter, which interacts with coregulators and with the basal transcriptional machinery, is the main posttranslational modifications site. The LBD, which is made up of 12 α-helices and 4 β-sheets, forms a hydrophobic pocket needed for GC binding and includes an AF2 domain. The latter allows interaction with coregulators in a ligand-dependent way. Finally, two nuclear localization signals, named NL1 and NL2, are localized in the DBD-hinge region junction and within the LBD, respectively. Asterisks indicate the location of the starting amino acid (aa position: 1, 27, 86, 90, 98, 316, 331, 336) of the eight different GRα translational isoforms, which are characterised by progressively shorter NTDs.
Figure 1.
Glucocorticoid receptor domain structure and translational isoforms. The N-terminal domain (NTD), which is required for ligand-independent gene transactivation, includes a transcriptional activation function region (AF1). The latter, which interacts with coregulators and with the basal transcriptional machinery, is the main posttranslational modifications site. The LBD, which is made up of 12 α-helices and 4 β-sheets, forms a hydrophobic pocket needed for GC binding and includes an AF2 domain. The latter allows interaction with coregulators in a ligand-dependent way. Finally, two nuclear localization signals, named NL1 and NL2, are localized in the DBD-hinge region junction and within the LBD, respectively. Asterisks indicate the location of the starting amino acid (aa position: 1, 27, 86, 90, 98, 316, 331, 336) of the eight different GRα translational isoforms, which are characterised by progressively shorter NTDs.

Figure 2.
Representative picture of the canonical GR signalling pathway. After binding to GC, GR undergoes a FKBP51-FKBP52 mediated conformational change, becomes hyper-phosphorylated, dissociates from accessory proteins (chaperone complex) and finally translocates into the nucleus. Here, after dimerization with other GRs, it regulates the transcription of target genes by binding to DNA. Interestingly, GR may enhance or repress transcription of target genes by directly binding to palindromic GC response elements (GRE), or by tethering itself to other transcription factors apart from DNA binding, or in a composite manner by both directly binding GRE and interacting with transcription factors bound to neighbouring sites. Created with BioRender.com.
Figure 2.
Representative picture of the canonical GR signalling pathway. After binding to GC, GR undergoes a FKBP51-FKBP52 mediated conformational change, becomes hyper-phosphorylated, dissociates from accessory proteins (chaperone complex) and finally translocates into the nucleus. Here, after dimerization with other GRs, it regulates the transcription of target genes by binding to DNA. Interestingly, GR may enhance or repress transcription of target genes by directly binding to palindromic GC response elements (GRE), or by tethering itself to other transcription factors apart from DNA binding, or in a composite manner by both directly binding GRE and interacting with transcription factors bound to neighbouring sites. Created with BioRender.com.

Figure 3.
HIF-α isoforms and their receptor HIF-1β/ARNT structural domains. (A) In vertebrates both HIF-1α and HIF-2α, but not HIF-3α, contain a basic helix-loop-helix (bHLH) domain, a Per-Arnt-Sim domain (PAS), an oxygen dependent degradation (ODD) domain, an N-terminal transactivation domain (N-TAD) located in the ODD and a C-TAD localized in the C-terminal region. (B) The HIF signalling pathway. Under normoxic conditions, PHDs hydroxylate HIF-α subunits on two specific prolyl residues within the ODDD. In turn, VHL recognizes and binds to hydroxylated HIF-α and then recruits the other components of the E3-ubiquitin ligase complex. The latter promotes the ubiquitin-mediated proteasomal degradation of HIF-α subunits. Conversely, hypoxic conditions inhibit PHD activity, and the subsequent degradation of HIF-α, which can, in turn, be stabilized in the cytoplasm, can dimerize with HIF-β-subunit and migrate into the nucleus. Here, the HIF-αβ active complex enhance the expression of target genes such as PHD3, VEGF, GLUT1 and EPO involved in restoring oxygen homeostasis. Created with BioRender.com.
Figure 3.
HIF-α isoforms and their receptor HIF-1β/ARNT structural domains. (A) In vertebrates both HIF-1α and HIF-2α, but not HIF-3α, contain a basic helix-loop-helix (bHLH) domain, a Per-Arnt-Sim domain (PAS), an oxygen dependent degradation (ODD) domain, an N-terminal transactivation domain (N-TAD) located in the ODD and a C-TAD localized in the C-terminal region. (B) The HIF signalling pathway. Under normoxic conditions, PHDs hydroxylate HIF-α subunits on two specific prolyl residues within the ODDD. In turn, VHL recognizes and binds to hydroxylated HIF-α and then recruits the other components of the E3-ubiquitin ligase complex. The latter promotes the ubiquitin-mediated proteasomal degradation of HIF-α subunits. Conversely, hypoxic conditions inhibit PHD activity, and the subsequent degradation of HIF-α, which can, in turn, be stabilized in the cytoplasm, can dimerize with HIF-β-subunit and migrate into the nucleus. Here, the HIF-αβ active complex enhance the expression of target genes such as PHD3, VEGF, GLUT1 and EPO involved in restoring oxygen homeostasis. Created with BioRender.com.

Figure 4.
Speculative scheme of how putative HIF-GC crosstalk occurs in the zebrafish. Previous research highlighted that GCs may enhance both GR and MR-mediated responsiveness and stimulate the HIF signalling. Conversely, upregulated HIF levels may inhibit GC biosynthesis by negatively regulating pomca at the level of the pituitary gland. Consequently, this is believed to occur as a logic homeostatic response exerted by HIF signalling to control not only its own activity but that of glucocorticoids to facilitate inflammation resolution and homeostasis. Created with BioRender.com.
Figure 4.
Speculative scheme of how putative HIF-GC crosstalk occurs in the zebrafish. Previous research highlighted that GCs may enhance both GR and MR-mediated responsiveness and stimulate the HIF signalling. Conversely, upregulated HIF levels may inhibit GC biosynthesis by negatively regulating pomca at the level of the pituitary gland. Consequently, this is believed to occur as a logic homeostatic response exerted by HIF signalling to control not only its own activity but that of glucocorticoids to facilitate inflammation resolution and homeostasis. Created with BioRender.com.

Figure 5.
Speculative scheme of how the putative HIF-GC crosstalk occurs in inflammation. We speculate that HIF can negatively regulate pomca expression to control not only its own activity but also that of glucocorticoids throughout the inflammatory process. Indeed, if not properly controlled, glucocorticoids could upregulate GILZ, whose overexpression has been demonstrated to suppress NF-κB activation. Since the latter can, in turn, increase the transcription of HIF-1α in response to hypoxic and inflammatory stimuli, its downregulation would hamper HIF activation and avoid essential pro-inflammatory HIF-mediated effects to occur.
Figure 5.
Speculative scheme of how the putative HIF-GC crosstalk occurs in inflammation. We speculate that HIF can negatively regulate pomca expression to control not only its own activity but also that of glucocorticoids throughout the inflammatory process. Indeed, if not properly controlled, glucocorticoids could upregulate GILZ, whose overexpression has been demonstrated to suppress NF-κB activation. Since the latter can, in turn, increase the transcription of HIF-1α in response to hypoxic and inflammatory stimuli, its downregulation would hamper HIF activation and avoid essential pro-inflammatory HIF-mediated effects to occur.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marchi, D.; van Eeden, F.J.M. Homeostatic Regulation of Glucocorticoid Receptor Activity by Hypoxia-Inducible Factor 1: From Physiology to Clinic. Cells 2021, 10, 3441. https://doi.org/10.3390/cells10123441
AMA Style
Marchi D, van Eeden FJM. Homeostatic Regulation of Glucocorticoid Receptor Activity by Hypoxia-Inducible Factor 1: From Physiology to Clinic. Cells. 2021; 10(12):3441. https://doi.org/10.3390/cells10123441
Chicago/Turabian StyleMarchi, Davide, and Fredericus J. M. van Eeden. 2021. "Homeostatic Regulation of Glucocorticoid Receptor Activity by Hypoxia-Inducible Factor 1: From Physiology to Clinic" Cells 10, no. 12: 3441. https://doi.org/10.3390/cells10123441
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.