
Aging-Related Cellular, Structural and Functional Changes in the Lymph Nodes: A Significant Component of Immunosenescence? An Overview


Abstract
:1. Introduction
2. Lymph Nodes Are Essential Components of the Immune System
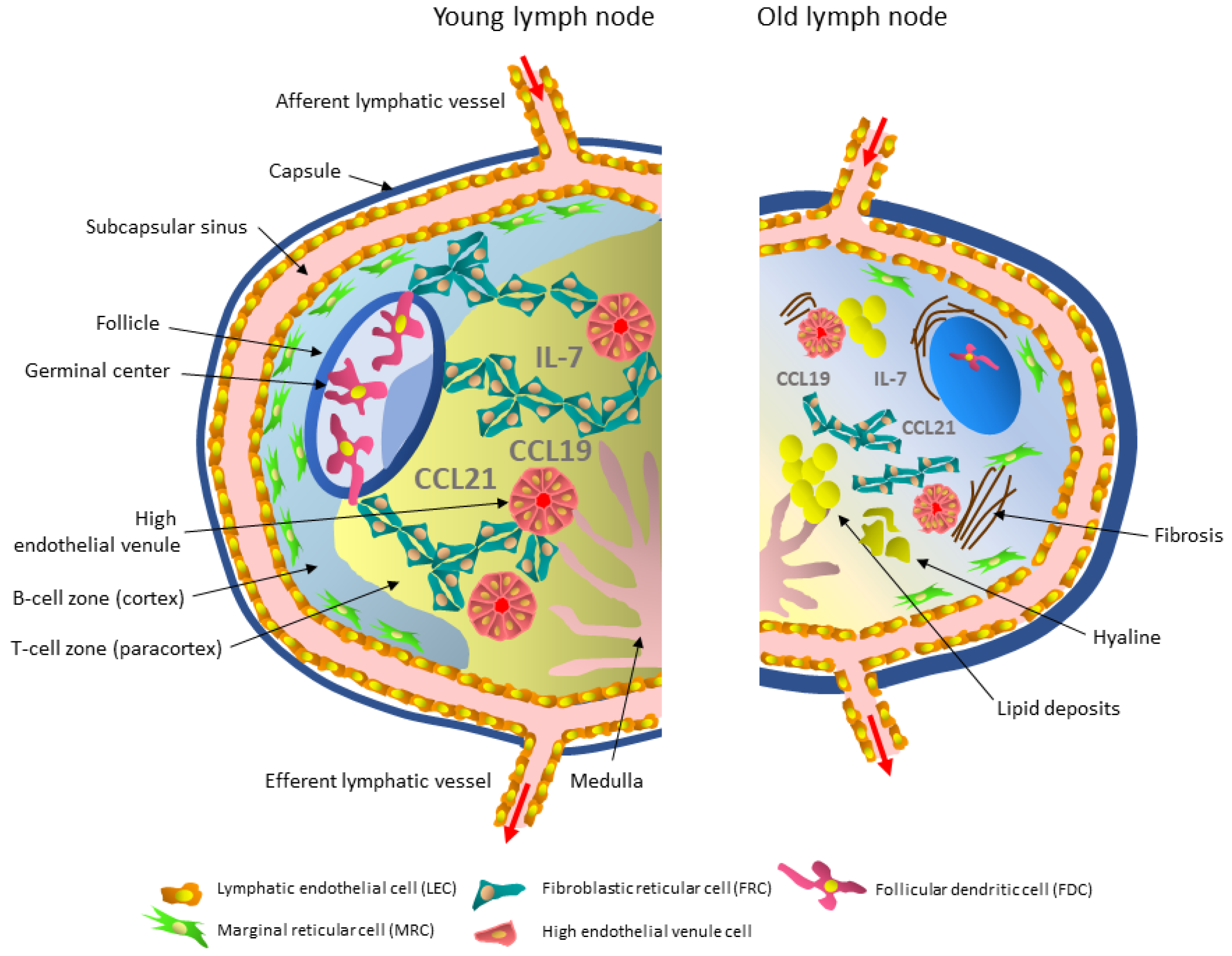
3. Aging-Related Structural, Cellular, and Functional Changes in Lymph Nodes
3.1. Stromal Cells
3.1.1. Lymphatic Endothelial Cells and Their Aging
3.1.2. High Endothelial Venule Cells and Their Aging
3.1.3. Marginal Reticular Cells and Their Aging
3.1.4. Follicular Dendritic Cells and Their Aging
3.1.5. Fibroblastic Reticular Cells and Their Aging

{kind=link}
{kind=link}
| Cells | Young Lymph Node | Old Lymph Node |
|---|---|---|
Lymphatic endothelial cells (LECs) |
|
|
| High endothelial venule cells  |
|
|
| Marginal reticular cells (MRCs)  |
|
|
Follicular dendritic cells (FDCs) |
|
|
Fibroblastic reticular cells (FRCs) |
|
|
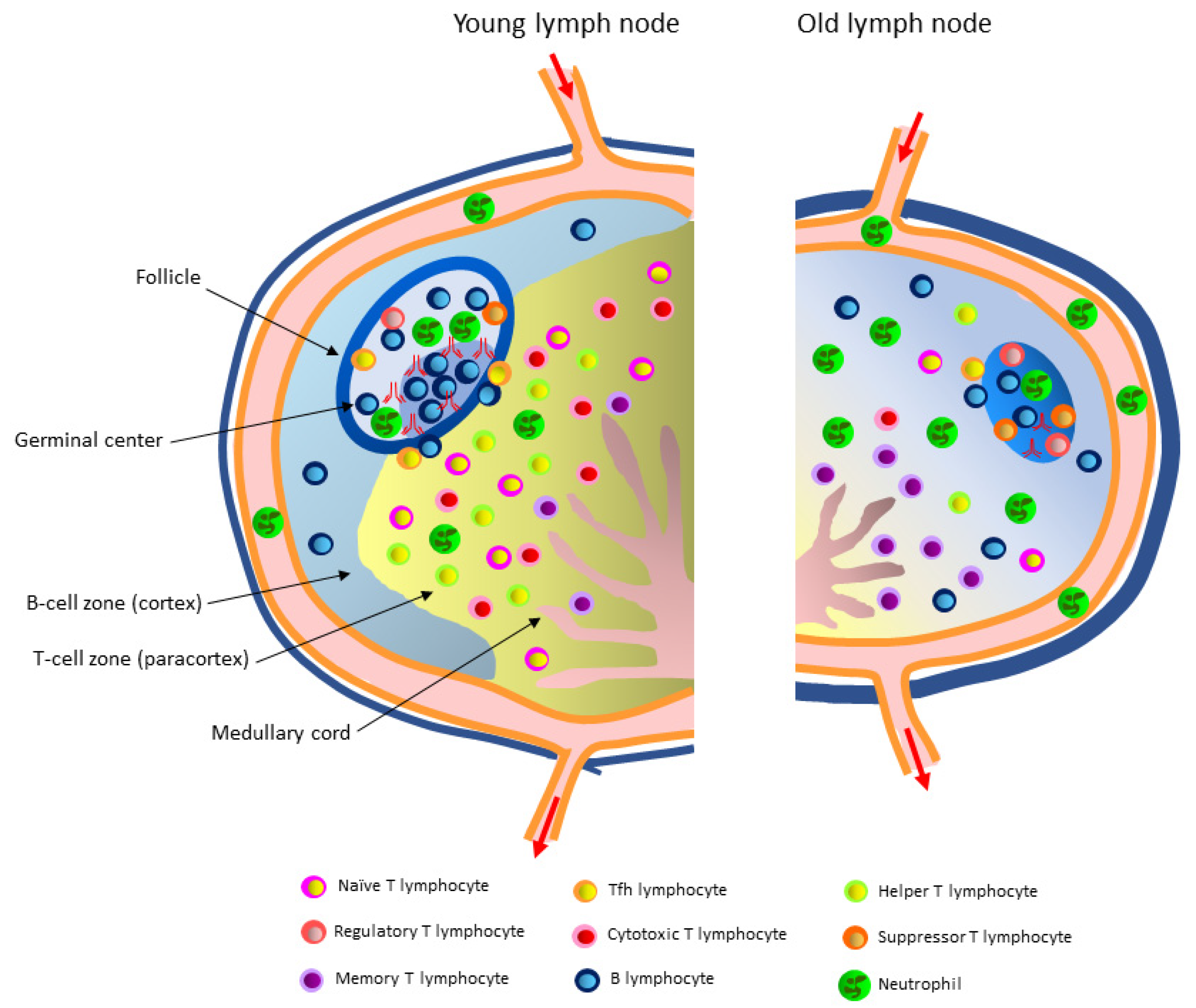
3.2. Immune Cells
3.2.1. T Lymphocytes in the Paracortex of the Lymph Node and Aging-Related Changes in the T Lymphocyte Microenvironment
3.2.2. B Lymphocytes in the Follicles of the Lymph Node and Aging-Related Changes in the B Lymphocyte Microenvironment
3.2.3. Neutrophils and Aging-Related Changes in Their Function
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yu, Y.; Zheng, S. Research progress on immune aging and its mechanisms affecting geriatric diseases. Aging Med. 2019, 2, 216–222. [Google Scholar] [CrossRef]
- Morrisette-Thomas, V.; Cohen, A.A.; Fülöp, T.; Riesco, É.; Legault, V.; Li, Q.; Milot, E.; Dusseault-Bélanger, F.; Ferrucci, L. Inflamm-aging does not simply reflect increases in pro-inflammatory markers. Mech. Ageing Dev. 2014, 139, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Fulop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.H.; Cohen, A.A.; Witkowski, J.M.; Franceschi, C. Immunosenescence and inflamm-aging as two sides of the same coin: Friends or foes? Front. Immunol. 2018, 8, 1960. [Google Scholar] [CrossRef] [Green Version]
- Thompson, H.L.; Smithey, M.J.; Surh, C.D.; Nikolich-Zugich, J. Functional and homeostatic impact of age-related changes in lymph node stroma. Front. Immunol. 2017, 8, 706. [Google Scholar] [CrossRef] [Green Version]
- Hazeldine, J.; Lord, J.M. The impact of ageing on natural killer cell function and potential consequences for health in older adults. Ageing Res. Rev. 2013, 12, 1069–1078. [Google Scholar] [CrossRef]
- Pritz, T.; Weinberger, B.; Grubeck-Loebenstein, B. The aging bone marrow and its impact on immune responses in old age. Immunol. Lett. 2014, 162, 310–315. [Google Scholar] [CrossRef]
- Pritz, T.; Lair, J.; Ban, M.; Keller, M.; Weinberger, B.; Krismer, M.; Grubeck-Loebenstein, B. Plasma cell numbers decrease in bone marrow of old patients. Eur. J. Immunol. 2015, 45, 738–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelmagid, S.M.; Barbe, M.F.; Safadi, F.F. Role of inflammation in the aging bones. Life Sci. 2015, 123, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Denkinger, M.D.; Leins, H.; Schirmbeck, R.; Florian, M.C.; Geiger, H. HSC aging and senescent immune remodeling. Trends Immunol. 2015, 36, 815–824. [Google Scholar] [CrossRef] [Green Version]
- Masters, A.R.; Haynes, L.; Su, D.M.; Palmer, D.B. Immune senescence: Significance of the stromal microenvironment. Clin. Exp. Immunol. 2017, 187, 6–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepletier, A.; Chidgey, A.P.; Savino, W. Perspectives for improvement of the thymic microenvironment through manipulation of thymic epithelial cells: A mini-review. Gerontology 2015, 61, 504–514. [Google Scholar] [CrossRef]
- Miranda, R.N.; Khoury, J.D.; Medeiros, L.J. Normal lymph node architecture and function. In Atlas of Lymph Node Pathology; Miranda, R.N., Khoury, J.D., Medeiros, L.J., Eds.; Springer: New York, NY, USA, 2013; pp. 3–17. [Google Scholar]
- Ruddle, N.H. High endothelial venules and lymphatic vessels in tertiary lymphoid organs: Characteristics, functions, and regulation. Front. Immunol. 2016, 7, 491. [Google Scholar] [CrossRef] [Green Version]
- Murakami, G.; Taniguchi, I. Histologic heterogeneity and intranodal shunt flow in lymph nodes from elderly subjects: A cadaveric study. Ann. Surg. Oncol. 2004, 11, 279S–284S. [Google Scholar] [CrossRef] [PubMed]
- Turner, V.M.; Mabbott, N.A. Structural and functional changes to lymph nodes in ageing mice. Immunology 2017, 151, 239–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briceño, O.; Lissina, A.; Wanke, K.; Afonso, G.; von Braun, A.; Ragon, K.; Miquel, T.; Gostick, E.; Papagno, L.; Stiasny, K.; et al. Reduced naïve CD8(+) T-cell priming efficacy in elderly adults. Aging Cell 2016, 15, 14–21. [Google Scholar] [CrossRef]
- Govaert, T.M.; Thijs, C.T.; Masurel, N.; Sprenger, M.J.; Dinant, G.J.; Knottnerus, J.A. The efficacy of influenza vaccination in elderly individuals. A randomized double-blind placebo-controlled trial. JAMA 1994, 272, 1661–1665. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, K.; Viboud, C.; Simonsen, L. Antibody response to influenza vaccination in the elderly: A quantitative review. Vaccine 2006, 24, 1159–1169. [Google Scholar] [CrossRef]
- Weinberger, B.; Grubeck-Loebenstein, B. Vaccines for the elderly. Clin. Microbiol. Infect. 2012, 18, 100–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciabattini, A.; Nardini, C.; Santoro, F.; Garagnani, P.; Franceschi, C.; Medaglini, D. Vaccination in the elderly: The challenge of immune changes with aging. Semin. Immunol. 2018, 40, 83–94. [Google Scholar] [CrossRef]
- Taniguchi, I.; Murakami, G.; Sato, A.; Fujiwara, D.; Ichikawa, H.; Yajima, T.; Kohama, G. Lymph node hyalinization in elderly Japanese. Histol. Histopathol. 2003, 18, 1169–1180. [Google Scholar] [CrossRef]
- Ahmadi, O.; Mccall, J.L.; Stringer, M.D. Does senescence affect lymph node number and morphology? A systematic review. ANZ J. Surg. 2013, 83, 612–618. [Google Scholar] [CrossRef]
- Card, C.M.; Yu, S.S.; Swartz, M.A. Emerging roles of lymphatic endothelium in regulating adaptive immunity. J. Clin. Investig. 2014, 124, 943–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Andrian, U.; Mempel, T. Homing and cellular traffic in lymph nodes. Nat. Rev. Immunol. 2003, 3, 867–878. [Google Scholar] [CrossRef] [PubMed]
- Girard, J.; Moussion, C.; Förster, R. HEVs, lymphatics and homeostatic immune cell trafficking in lymph nodes. Nat. Rev. Immunol. 2012, 12, 762–773. [Google Scholar] [CrossRef]
- Miyasaka, M.; Tanaka, T. Lymphocyte trafficking across high endothelial venules: Dogmas and enigmas. Nat. Rev. Immunol. 2004, 4, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Rosen, S.D. Ligands for L-selectin: Homing, inflammation, and beyond. Annu. Rev. Immunol. 2004, 22, 129–156. [Google Scholar] [CrossRef] [PubMed]
- Hadamitzky, C.; Spohr, H.; Debertin, A.S.; Guddat, S.; Tsokos, M.; Pabst, R. Age-dependent histoarchitectural changes in human lymph nodes: An underestimated process with clinical relevance? J. Anat. 2010, 216, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Aydar, Y.; Balogh, P.; Tew, J.G.; Szakal, A.K. Age-related depression of FDC accessory functions and CD21 ligand-mediated repair of costimulation. Eur. J. Immunol. 2002, 32, 2817–2826. [Google Scholar] [CrossRef]
- Richner, J.M.; Gmyrek, G.B.; Govero, J.; Tu, Y.; van der Windt, G.J.; Metcalf, T.U.; Haddad, E.K.; Textor, J.; Miller, M.J.; Diamond, M.S. Age-dependent cell trafficking defects in draining lymph nodes impair adaptive immunity and control of West Nile virus infection. PLoS Pathog. 2015, 11, e1005027. [Google Scholar] [CrossRef] [Green Version]
- Becklund, B.R.; Purton, J.F.; Ramsey, C.; Favre, S.; Vogt, T.K.; Martin, C.E.; Spasova, D.S.; Sarkisyan, G.; LeRoy, E.; Tan, J.T.; et al. The aged lymphoid tissue environment fails to support naïve T cell homeostasis. Sci. Rep. 2016, 6, 30842. [Google Scholar] [CrossRef]
- Turner, V.M.; Mabbott, N.A. Ageing adversely affects the migration and function of marginal zone B cells. Immunology 2017, 151, 349–362. [Google Scholar] [CrossRef] [Green Version]
- Turner, V.M.; Mabbott, N.A. Influence of ageing on the microarchitecture of the spleen and lymph nodes. Biogerontology 2017, 18, 723–738. [Google Scholar] [CrossRef] [PubMed]
- Luscieti, P.; Hubschmid, T.; Cottier, H.; Hess, M.W.; Sobin, L.H. Human lymph node morphology as a function of age and site. J. Clin. Pathol. 1980, 33, 454–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denz, F.A. Age changes in lymph nodes. J. Pathol. Bacteriol. 1947, 59, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Erofeeva, L.M.; Mnikhovich, M.V. Structural and functional changes in the mesenteric lymph nodes in humans during aging. Bull. Exp. Biol. Med. 2020, 168, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.R.; Suami, H.; Taylor, G.I. Senile changes in human lymph nodes. Lymphat. Res. Biol. 2008, 6, 77–83. [Google Scholar] [CrossRef]
- Mueller, S.N.; Germain, R.N. Stromal cell contributions to the homeostasis and functionality of the immune system. Nat. Rev. Immunol. 2009, 9, 618–629. [Google Scholar] [CrossRef]
- Chang, J.E.; Turley, S.J. Stromal infrastructure of the lymph node and coordination of immunity. Trends Immunol. 2015, 36, 30–39. [Google Scholar] [CrossRef]
- Fletcher, A.L.; Acton, S.E.; Knoblich, K. Lymph node fibroblastic reticular cells in health and disease. Nat. Rev. Immunol. 2015, 15, 350–361. [Google Scholar] [CrossRef] [Green Version]
- Bajénoff, M.; Egen, J.G.; Qi, H.; Huang, A.Y.; Castellino, F.; Germain, R.N. Highways, byways and breadcrumbs: Directing lymphocyte traffic in the lymph node. Trends Immunol. 2007, 28, 346–352. [Google Scholar] [CrossRef]
- Link, A.; Vogt, T.K.; Favre, S.; Britschgi, M.R.; Acha-Orbea, H.; Hinz, B.; Cyster, J.G.; Luther, S.A. Fibroblastic reticular cells in lymph nodes regulate the homeostasis of naive T cells. Nat. Immunol. 2007, 8, 1255–1265. [Google Scholar] [CrossRef] [PubMed]
- Aw, D.; Palmer, D.B. The origin and implication of thymic involution. Aging Dis. 2011, 2, 437–443. [Google Scholar] [PubMed]
- Filelfi, S.L.; Onorato, A.; Brix, B.; Goswami, N. Lymphatic senescence: Current updates and perspectives. Biology 2021, 10, 293. [Google Scholar] [CrossRef]
- Coplen, C.; Jeftic, I.; Thompson, H.; Nikolich-Zugich, J. Reversal of age-associated lymph node atrophy requires improved stromal cell support and reduced fibrosis. Innov. Aging 2018, 2, 886–887. [Google Scholar] [CrossRef]
- Budamagunta, V.; Foster, T.C.; Zhou, D. Cellular senescence in lymphoid organs and immunosenescence. Aging 2021, 13, 19920–19941. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, B.; Herndler-Brandstetter, D.; Schwanninger, A.; Weiskopf, D.; Grubeck-Loebenstein, B. Biology of immune responses to vaccines in elderly persons. Clin. Infect. Dis. 2008, 46, 1078–1084. [Google Scholar] [CrossRef]
- Erofeeva, L.M.; Mnikhovich, M.V. Changes in the structure and cell composition of human carinal lymph nodes during aging. Bull. Exp. Biol. Med. 2018, 165, 702–706. [Google Scholar] [CrossRef]
- Julg, B.; Alter, G. Lymph node fibrosis: A structural barrier to unleashing effective vaccine immunity. J. Clin. Investig. 2018, 128, 2743–2745. [Google Scholar] [CrossRef]
- Demchenko, G.A.; Abdreshov, S.N.; Nurmakhanova, B.A. Contractile activity of lymph nodes in young, middle-aged, and old rats. Bull. Exp. Biol. Med. 2019, 167, 194–197. [Google Scholar] [CrossRef] [PubMed]
- Shang, T.; Liang, J.; Kapron, C.M.; Liu, J. Pathophysiology of aged lymphatic vessels. Aging 2019, 11, 6602–6613. [Google Scholar] [CrossRef]
- Wiig, H.; Swartz, M.A. Interstitial fluid and lymph formation and transport: Physiological regulation and roles in inflammation and cancer. Physiol. Rev. 2012, 92, 1005–1060. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.C.; D’Alessandro, A.; Clement, C.C.; Santambrogio, L. Lymph formation, composition and circulation: A proteomics perspective. Int. Immunol. 2015, 27, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randolph, G.J.; Ivanov, S.; Zinselmeyer, B.H.; Scallan, J.P. The lymphatic system: Integral roles in immunity. Annu. Rev. Immunol. 2017, 35, 31–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sainte-Marie, G.; Peng, F.S.; Belisle, C. Overall architecture and pattern of lymph flow in the rat lymph node. Am. J. Anat. 1982, 164, 275–309. [Google Scholar] [CrossRef]
- Sainte-Marie, G.; Peng, F.S. Evidence for the existence of a subsinus layer of the peripheral cortex in the lymph node of the rat. Cell Tissue Res. 1985, 239, 37–42. [Google Scholar] [CrossRef]
- Nikles, S.A.; Heath, T.J. Pathways of lymph flow through intestinal lymph nodes in the horse. Anat. Rec. 1992, 232, 126–132. [Google Scholar] [CrossRef]
- Clement, C.C.; Wang, W.; Dzieciatkowska, M.; Cortese, M.; Hansen, K.C.; Becerra, A.; Thangaswamy, S.; Nizamutdinova, I.; Moon, J.Y.; Stern, L.J.; et al. Quantitative profiling of the lymph node clearance capacity. Sci. Rep. 2018, 8, 11253. [Google Scholar] [CrossRef]
- Onder, L.; Mörbe, U.; Pikor, N.; Novkovic, M.; Cheng, H.W.; Hehlgans, T.; Pfeffer, K.; Becher, B.; Waisman, A.; Rülicke, T.; et al. Lymphatic endothelial cells control initiation of lymph node organogenesis. Immunity 2017, 47, 80–92.e4. [Google Scholar] [CrossRef] [Green Version]
- Jalkanen, S.; Salmi, M. Lymphatic endothelial cells of the lymph node. Nat. Rev. Immunol. 2020, 20, 566–578. [Google Scholar] [CrossRef]
- Cohen, J.N.; Guidi, C.J.; Tewalt, E.F.; Qiao, H.; Rouhani, S.J.; Ruddell, A.; Farr, A.G.; Tung, K.S.; Engelhard, V.H. Lymph node-resident lymphatic endothelial cells mediate peripheral tolerance via Aire-independent direct antigen presentation. J. Exp. Med. 2010, 207, 681–688. [Google Scholar] [CrossRef] [Green Version]
- Hirosue, S.; Vokali, E.; Raghavan, V.R.; Rincon-Restrepo, M.; Lund, A.W.; Corthésy-Henrioud, P.; Capotosti, F.; Halin Winter, C.; Hugues, S.; Swartz, M.A. Steady-state antigen scavenging, cross-presentation, and CD8+ T cell priming: A new role for lymphatic endothelial cells. J. Immunol. 2014, 192, 5002–5011. [Google Scholar] [CrossRef] [Green Version]
- Lukacs-Kornek, V.; Malhotra, D.; Fletcher, A.L.; Acton, S.E.; Elpek, K.G.; Tayalia, P.; Collier, A.R.; Turley, S.J. Regulated release of nitric oxide by nonhematopoietic stroma controls expansion of the activated T cell pool in lymph nodes. Nat. Immunol. 2011, 12, 1096–1104. [Google Scholar] [CrossRef]
- Nörder, M.; Gutierrez, M.G.; Zicari, S.; Cervi, E.; Caruso, A.; Guzmán, C.A. Lymph node-derived lymphatic endothelial cells express functional costimulatory molecules and impair dendritic cell-induced allogenic T-cell proliferation. FASEB J. 2012, 26, 2835–2846. [Google Scholar] [CrossRef]
- Hara, T.; Shitara, S.; Imai, K.; Miyachi, H.; Kitano, S.; Yao, H.; Tani-ichi, S.; Ikuta, K. Identification of IL-7-producing cells in primary and secondary lymphoid organs using IL-7-GFP knock-in mice. J. Immunol. 2012, 189, 1577–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceschi, C.; Bonafe, M.; Valensin, S. Human immunosenescence: The prevailing of innate immunity, the failing of clonotypic immunity, and the filling of immunological space. Vaccine 2000, 18, 1717–1720. [Google Scholar] [CrossRef]
- Panda, A.; Arjona, A.; Sapey, E.; Bai, F.; Fikrig, E.; Montgomery, R.R.; Lord, J.M.; Shaw, A.C. Human innate immunosenescence: Causes and consequences for immunity in old age. Trends Immunol. 2009, 30, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Masters, A.R.; Hall, A.; Bartley, J.M.; Keilich, S.R.; Lorenzo, E.C.; Jellison, E.R.; Puddington, L.; Haynes, L. Assessment of lymph node stromal cells as an underlying factor in age-related immune impairment. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 1734–1743. [Google Scholar] [CrossRef] [PubMed]
- Thangaswamy, S.; Bridenbaugh, E.A.; Gashev, A.A. Evidence of increased oxidative stress in aged mesenteric lymphatic vessels. Lymphat. Res. Biol. 2012, 10, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Zolla, V.; Nizamutdinova, I.T.; Scharf, B.; Clement, C.C.; Maejima, D.; Akl, T.; Nagai, T.; Luciani, P.; Leroux, J.C.; Halin, C.; et al. Aging-related anatomical and biochemical changes in lymphatic collectors impair lymph transport, fluid homeostasis, and pathogen clearance. Aging Cell 2015, 14, 582–594. [Google Scholar] [CrossRef]
- Ahn, S.M.; Simpson, R.J. Body fluid proteomics: Prospects for biomarker discovery. Proteom. Clin. Appl. 2007, 1, 1004–1015. [Google Scholar] [CrossRef]
- Meng, Z.; Veenstra, T.D. Proteomic analysis of serum, plasma, and lymph for the identification of biomarkers. Proteom. Clin. Appl. 2007, 1, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Clement, C.C.; Cannizzo, E.S.; Nastke, M.D.; Sahu, R.; Olszewski, W.; Miller, N.E.; Stern, L.J.; Santambrogio, L. An expanded self-antigen peptidome is carried by the human lymph as compared to the plasma. PLoS ONE 2010, 5, e9863. [Google Scholar] [CrossRef]
- Fang, J.F.; Shih, L.Y.; Yuan, K.C.; Fang, K.Y.; Hwang, T.L.; Hsieh, S.Y. Proteomic analysis of post-hemorrhagic shock mesenteric lymph. Shock 2010, 34, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Clement, C.C.; Rotzschke, O.; Santambrogio, L. The lymph as a pool of self-antigens. Trends Immunol. 2011, 32, 6–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, C.C.; Santambrogio, L. The lymph self-antigen repertoire. Front. Immunol. 2013, 4, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, J.P.; Springer, T.A. High endothelial venules (HEVs): Specialized endothelium for lymphocyte migration. Immunol. Today 1995, 16, 449–457. [Google Scholar] [CrossRef]
- Moussion, C.; Girard, J.P. Dendritic cells control lymphocyte entry to lymph nodes through high endothelial venules. Nature 2011, 479, 542–546. [Google Scholar] [CrossRef]
- Katakai, T. Marginal reticular cells: A stromal subset directly descended from the lymphoid tissue organizer. Front. Immunol. 2012, 3, 200. [Google Scholar] [CrossRef] [Green Version]
- Katakai, T.; Suto, H.; Sugai, M.; Gonda, H.; Togawa, A.; Suematsu, S.; Ebisuno, Y.; Katagiri, K.; Kinashi, T.; Shimizu, A. Organizer-like reticular stromal cell layer common to adult secondary lymphoid organs. J. Immunol. 2008, 181, 6189–6200. [Google Scholar] [CrossRef] [Green Version]
- Jarjour, M.; Jorquera, A.; Mondor, I.; Wienert, S.; Narang, P.; Coles, M.C.; Klauschen, F.; Bajénoff, M. Fate mapping reveals origin and dynamics of lymph node follicular dendritic cells. J. Exp. Med. 2014, 211, 1109–1122. [Google Scholar] [CrossRef] [Green Version]
- Fasnacht, N.; Huang, H.Y.; Koch, U.; Favre, S.; Auderset, F.; Chai, Q.; Onder, L.; Kallert, S.; Pinschewer, D.D.; MacDonald, H.R.; et al. Specific fibroblastic niches in secondary lymphoid organs orchestrate distinct Notch-regulated immune responses. J. Exp. Med. 2014, 211, 2265–2279. [Google Scholar] [CrossRef] [Green Version]
- Holmes, K.L.; Schnizlein, C.T.; Perkins, E.H.; Tew, J.G. The effect of age on antigen retention in lymphoid follicles and in collagenous tissue of mice. Mech. Ageing Dev. 1984, 25, 243–255. [Google Scholar] [CrossRef]
- Hoorweg, K.; Narang, P.; Li, Z.; Thuery, A.; Papazian, N.; Withers, D.R.; Coles, M.C.; Cupedo, T. A stromal cell niche for human and mouse type 3 innate lymphoid cells. J. Immunol. 2015, 195, 4257–4263. [Google Scholar] [CrossRef] [Green Version]
- Magri, G.; Miyajima, M.; Bascones, S.; Mortha, A.; Puga, I.; Cassis, L.; Barra, C.M.; Comerma, L.; Chudnovskiy, A.; Gentile, M.; et al. Innate lymphoid cells integrate stromal and immunological signals to enhance antibody production by splenic marginal zone B cells. Nat. Immunol. 2014, 15, 354–364. [Google Scholar] [CrossRef] [Green Version]
- Denton, A.E.; Silva-Cayetano, A.; Dooley, J.; Hill, D.L.; Carr, E.J.; Robert, P.A.; Meyer-Hermann, M.; Liston, A.; Linterman, M.A. Intrinsic defects in lymph node stromal cells underpin poor germinal center responses during aging. bioRxiv 2020. [Google Scholar] [CrossRef]
- Wols, H.A.; Johnson, K.M.; Ippolito, J.A.; Birjandi, S.Z.; Su, Y.; Le, P.T.; Witte, P.L. Migration of immature and mature B cells in the aged microenvironment. Immunology 2010, 129, 278–290. [Google Scholar] [CrossRef]
- Allen, C.D.; Cyster, J.G. Follicular dendritic cell networks of primary follicles and germinal centers: Phenotype and function. Semin. Immunol. 2008, 20, 14–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannard, O.; Horton, R.M.; Allen, C.D.; An, J.; Nagasawa, T.; Cyster, J.G. Germinal center centroblasts transition to a centrocyte phenotype according to a timed program and depend on the dark zone for effective selection. Immunity 2013, 39, 912–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodda, L.B.; Bannard, O.; Ludewig, B.; Nagasawa, T.; Cyster, J.G. Phenotypic and morphological properties of germinal center dark zone Cxcl12-expressing reticular cells. J. Immunol. 2015, 195, 4781–4791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pikor, N.B.; Mörbe, U.; Lütge, M.; Gil-Cruz, C.; Perez-Shibayama, C.; Novkovic, M.; Cheng, H.W.; Nombela-Arrieta, C.; Nagasawa, T.; Linterman, M.A.; et al. Remodeling of light and dark zone follicular dendritic cells governs germinal center responses. Nat. Immunol. 2020, 21, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S. Follicular helper CD4 T cells (Tfh). Annu. Rev. Immunol. 2011, 29, 621–663. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Adams, J.C.; Steinman, R.M. Anatomy of germinal centers in mouse spleen, with special reference to “follicular dendritic cells”. J. Cell Biol. 1978, 77, 148–164. [Google Scholar] [CrossRef] [PubMed]
- Kranich, J.; Krautler, N.J. How follicular dendritic cells shape the B-cell antigenome. Front. Immunol. 2016, 7, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandel, T.E.; Phipps, R.P.; Abbot, A.; Tew, J.G. The follicular dendritic cell: Long term antigen retention during immunity. Immunol. Rev. 1980, 53, 29–59. [Google Scholar] [CrossRef] [PubMed]
- Heesters, B.A.; Myers, R.C.; Carroll, M.C. Follicular dendritic cells: Dynamic antigen libraries. Nat. Rev. Immunol. 2014, 14, 495–504. [Google Scholar] [CrossRef]
- Sage, P.T.; Tan, C.L.; Freeman, G.J.; Haigis, M.; Sharpe, A.H. Defective TFH cell function and increased TFR cells contribute to defective antibody production in aging. Cell Rep. 2015, 12, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Szakal, A.K.; Aydar, Y.; Balogh, P.; Tew, J.G. Molecular interactions of FDCs with B cells in aging. Semin. Immunol. 2002, 14, 267–274. [Google Scholar] [CrossRef]
- Aydar, Y.; Balogh, P.; Tew, J.G.; Szakal, A.K. Altered regulation of Fc gamma RII on aged follicular dendritic cells correlates with immunoreceptor tyrosine-based inhibition motif signaling in B cells and reduced germinal center formation. J. Immunol. 2003, 171, 5975–5987. [Google Scholar] [CrossRef] [Green Version]
- Aydar, Y.; Balogh, P.; Tew, J.G.; Szakal, A.K. Follicular dendritic cells in aging, a “bottle-neck” in the humoral immune response. Ageing Res. Rev. 2004, 3, 15–29. [Google Scholar] [CrossRef]
- Fletcher, A.L.; Malhotra, D.; Acton, S.E.; Lukacs-Kornek, V.; Bellemare-Pelletier, A.; Curry, M.; Armant, M.; Turley, S.J. Reproducible isolation of lymph node stromal cells reveals site-dependent differences in fibroblastic reticular cells. Front. Immunol. 2011, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.Y.; Vogt, T.K.; Favre, S.; Scarpellino, L.; Huang, H.Y.; Tacchini-Cottier, F.; Luther, S.A. Trapping of naive lymphocytes triggers rapid growth and remodeling of the fibroblast network in reactive murine lymph nodes. Proc. Natl. Acad. Sci. USA 2014, 111, E109–E118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onder, L.; Danuser, R.; Scandella, E.; Firner, S.; Chai, Q.; Hehlgans, T.; Stein, J.V.; Ludewig, B. Endothelial cell-specific lymphotoxin-β receptor signaling is critical for lymph node and high endothelial venule formation. J. Exp. Med. 2013, 210, 465–473. [Google Scholar] [CrossRef]
- Junt, T.; Scandella, E.; Ludewig, B. Form follows function: Lymphoid tissue microarchitecture in antimicrobial immune defense. Nat. Rev. Immunol. 2008, 8, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Katakai, T.; Hara, T.; Sugai, M.; Gonda, H.; Shimizu, A. Lymph node fibroblastic reticular cells construct the stromal reticulum via contact with lymphocytes. J. Exp. Med. 2004, 200, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Chyou, S.; Benahmed, F.; Chen, J.; Kumar, V.; Tian, S.; Lipp, M.; Lu, T.T. Coordinated regulation of lymph node vascular-stromal growth first by CD11c+ cells and then by T and B cells. J. Immunol. 2011, 187, 5558–5567. [Google Scholar] [CrossRef] [Green Version]
- Acton, S.E.; Farrugia, A.J.; Astarita, J.L.; Mourão-Sá, D.; Jenkins, R.P.; Nye, E.; Hooper, S.; van Blijswijk, J.; Rogers, N.C.; Snelgrove, K.J.; et al. Dendritic cells control fibroblastic reticular network tension and lymph node expansion. Nature 2014, 514, 498–502. [Google Scholar] [CrossRef]
- Mori, S.; Nakano, H.; Aritomi, K.; Wang, C.R.; Gunn, M.D.; Kakiuchi, T. Mice lacking expression of the chemokines CCL21-ser and CCL19 (plt mice) demonstrate delayed but enhanced T cell immune responses. J. Exp. Med. 2001, 193, 207–218. [Google Scholar] [CrossRef]
- Bajénoff, M.; Egen, J.G.; Koo, L.Y.; Laugier, J.P.; Brau, F.; Glaichenhaus, N.; Germain, R.N. Stromal cell networks regulate lymphocyte entry, migration, and territoriality in lymph nodes. Immunity 2006, 25, 989–1001. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, D.; Fletcher, A.L.; Astarita, J.; Lukacs-Kornek, V.; Tayalia, P.; Gonzalez, S.F.; Elpek, K.G.; Chang, S.K.; Knoblich, K.; Hemler, M.E.; et al. Transcriptional profiling of stroma from inflamed and resting lymph nodes defines immunological hallmarks. Nat. Immunol. 2012, 13, 499–510. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.Y.; Rivas-Caicedo, A.; Renevey, F.; Cannelle, H.; Peranzoni, E.; Scarpellino, L.; Hardie, D.L.; Pommier, A.; Schaeuble, K.; Favre, S.; et al. Identification of a new subset of lymph node stromal cells involved in regulating plasma cell homeostasis. Proc. Natl. Acad. Sci. USA 2018, 115, E6826–E6835. [Google Scholar] [CrossRef] [Green Version]
- Denton, A.E.; Roberts, E.W.; Linterman, M.A.; Fearon, D.T. Fibroblastic reticular cells of the lymph node are required for retention of resting but not activated CD8+ T cells. Proc. Natl. Acad. Sci. USA 2014, 111, 12139–12144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thannickal, V.J.; Zhou, Y.; Gaggar, A.; Duncan, S.R. Fibrosis: Ultimate and proximate causes. J. Clin. Investig. 2014, 124, 4673–4677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borthwick, L.A.; Wynn, T.A. IL-13 and TGF-β1: Core mediators of fibrosis. Curr. Pathobiol. Rep. 2015, 3, 273–282. [Google Scholar] [CrossRef]
- Thompson, H.L.; Smithey, M.J.; Uhrlaub, J.L.; Jeftić, I.; Jergović, M.; White, S.E.; Currier, N.; Lang, A.M.; Okoye, A.; Park, B.; et al. Lymph nodes as barriers to T-cell rejuvenation in aging mice and nonhuman primates. Aging Cell 2019, 18, e12865. [Google Scholar] [CrossRef] [Green Version]
- Textor, J.; Mandl, J.N.; de Boer, R.J. The reticular cell network: A robust backbone for immune responses. PLoS Biol. 2016, 14, e2000827. [Google Scholar] [CrossRef]
- D’Souza, W.N.; Hedrick, S.M. Cutting edge: Latecomer CD8 T cells are imprinted with a unique differentiation program. J. Immunol. 2006, 177, 777–781. [Google Scholar] [CrossRef] [Green Version]
- Chai, Q.; Onder, L.; Scandella, E.; Gil-Cruz, C.; Perez-Shibayama, C.; Cupovic, J.; Danuser, R.; Sparwasser, T.; Luther, S.A.; Thiel, V.; et al. Maturation of lymph node fibroblastic reticular cells from myofibroblastic precursors is critical for antiviral immunity. Immunity 2013, 38, 1013–1024. [Google Scholar] [CrossRef] [Green Version]
- Aebischer, D.; Iolyeva, M.; Halin, C. The inflammatory response of lymphatic endothelium. Angiogenesis. 2013, 17, 383–393. [Google Scholar] [CrossRef]
- Förster, R.; Davalos-Misslitz, A.C.; Rot, A. CCR7 and its ligands: Balancing immunity and tolerance. Nat. Rev. Immunol. 2008, 8, 362–371. [Google Scholar] [CrossRef]
- Pepper, M.; Jenkins, M.K. Origins of CD4+ effector and central memory T cells. Nat. Immunol. 2011, 12, 467–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahalan, M.D.; Parker, I. Imaging the choreography of lymphocyte trafficking and the immune response. Curr. Opin. Immunol. 2006, 18, 476–482. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, J.S.; Maue, A.C.; Eaton, S.M.; Lanthier, P.A.; Tighe, M.; Haynes, L. The aged microenvironment contributes to the age-related functional defects of CD4 T cells in mice. Aging Cell 2012, 11, 732–740. [Google Scholar] [CrossRef] [Green Version]
- Lazuardi, L.; Jenewein, B.; Wolf, A.M.; Pfister, G.; Tzankov, A.; Grubeck-Loebenstein, B. Age-related loss of naïve T cells and dysregulation of T-cell/B-cell interactions in human lymph nodes. Immunology 2005, 114, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Martinet, K.Z.; Bloquet, S.; Bourgeois, C. Ageing combines CD4 T cell lymphopenia in secondary lymphoid organs and T cell accumulation in gut associated lymphoid tissue. Immun. Ageing 2014, 11, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goronzy, J.J.; Weyand, C.M. Understanding immunosenescence to improve responses to vaccines. Nat. Immunol. 2013, 14, 428–436. [Google Scholar] [CrossRef] [Green Version]
- Davies, J.S.; Thompson, H.L.; Pulko, V.; Padilla Torres, J.; Nikolich-Zugich, J. Role of cell-intrinsic and environmental age-related changes in altered maintenance of murine T cells in lymphoid organs. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 1018–1026. [Google Scholar] [CrossRef]
- Lerner, A.; Yamada, T.; Miller, R.A. Pgp-1hi T lymphocytes accumulate with age in mice and respond poorly to concanavalin A. Eur. J. Immunol. 1989, 19, 977–982. [Google Scholar] [CrossRef]
- Ernst, D.N.; Hobbs, M.V.; Torbett, B.E.; Glasebrook, A.L.; Rehse, M.A.; Bottomly, K.; Hayakawa, K.; Hardy, R.R.; Weigle, W.O. Differences in the expression profiles of CD45RB, Pgp-1, and 3G11 membrane antigens and in the patterns of lymphokine secretion by splenic CD4+ T cells from young and aged mice. J. Immunol. 1990, 145, 1295–1302. [Google Scholar]
- Weisel, F.J.; Zuccarino-Catania, G.V.; Chikina, M.; Shlomchik, M.J. A temporal switch in the germinal center determines differential output of memory B and plasma cells. Immunity 2016, 44, 116–130. [Google Scholar] [CrossRef] [Green Version]
- Cyster, J.G. B cell follicles and antigen encounters of the third kind. Nat. Immunol. 2010, 11, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Baldazzi, V.; Paci, P.; Bernaschi, M.; Castiglione, F. Modeling lymphocyte homing and encounters in lymph nodes. BMC Bioinform. 2009, 10, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meffre, E.; Wardemann, H. B-cell tolerance checkpoints in health and autoimmunity. Curr. Opin. Immunol. 2008, 20, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Scheid, J.F.; Mouquet, H.; Kofer, J.; Yurasov, S.; Nussenzweig, M.C.; Wardemann, H. Differential regulation of self-reactivity discriminates between IgG+ human circulating memory B cells and bone marrow plasma cells. Proc. Natl. Acad. Sci. USA 2011, 108, 18044–18048. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.S.; Hussell, T.; Gilliland, S.M.; Holden, D.W.; Paton, J.C.; Ehrenstein, M.R.; Walport, M.J.; Botto, M. The classical pathway is the dominant complement pathway required for innate immunity to streptococcus pneumoniae infection in mice. Proc. Natl. Acad. Sci. USA 2002, 99, 16969–16974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn-Walters, D.K.; Silva O’Hare, J. Older human B cells and antibodies. In Handbook of Immunosenescence: Basic Understanding and Clinical Implications; Fulop, T., Franceschi, C., Hirokawa, K., Pawelec, G., Eds.; Springer: New York, NY, USA, 2019; pp. 785–819. [Google Scholar]
- Shankwitz, K.; Pallikkuth, S.; Sirupangi, T.; Kirk Kvistad, D.; Russel, K.B.; Pahwa, R.; Gama, L.; Koup, R.A.; Pan, L.; Villinger, F.; et al. Compromised steady-state germinal center activity with age in nonhuman primates. Aging Cell 2020, 19, e13087. [Google Scholar] [CrossRef]
- Kosco, M.H.; Burton, G.F.; Kapasi, Z.F.; Szakal, A.K.; Tew, J.G. Antibody-forming cell induction during an early phase of germinal centre development and its delay with ageing. Immunology 1989, 68, 312–318. [Google Scholar]
- Szakal, A.K.; Taylor, J.K.; Smith, J.P.; Kosco, M.H.; Burton, G.F.; Tew, J.J. Kinetics of germinal center development in lymph nodes of young and aging immune mice. Anat. Rec. 1990, 227, 475–485. [Google Scholar] [CrossRef]
- Boraschi, D.; Aguado, M.T.; Dutel, C.; Goronzy, J.; Louis, J.; Grubeck-Loebenstein, B.; Rappuoli, R.; Del Giudice, G. The gracefully aging immune system. Sci. Transl. Med. 2013, 5, 185ps8. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Stedra, J.; Cerny, J. Relative contribution of T and B cells to hypermutation and selection of the antibody repertoire in germinal centers of aged mice. J. Exp. Med. 1996, 183, 959–970. [Google Scholar] [CrossRef] [Green Version]
- Eaton, S.M.; Burns, E.M.; Kusser, K.; Randall, T.D.; Haynes, L. Age-related defects in CD4 T cell cognate helper function lead to reductions in humoral responses. J. Exp. Med. 2004, 200, 1613–1622. [Google Scholar] [CrossRef] [Green Version]
- Stebegg, M.; Bignon, A.; Hill, D.L.; Silva-Cayetano, A.; Krueger, C.; Vanderleyden, I.; Innocentin, S.; Boon, L.; Wang, J.; Zand, M.S.; et al. Rejuvenating conventional dendritic cells and T follicular helper cell formation after vaccination. eLife 2020, 9, e52473. [Google Scholar] [CrossRef]
- Sayin, I.; Radtke, A.J.; Vella, L.A.; Jin, W.; Wherry, E.J.; Buggert, M.; Betts, M.R.; Herati, R.S.; Germain, R.N.; Canaday, D.H. Spatial distribution and function of T follicular regulatory cells in human lymph nodes. J. Exp. Med. 2018, 215, 1531–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linterman, M.A.; Pierson, W.; Lee, S.K.; Kallies, A.; Kawamoto, S.; Rayner, T.F.; Srivastava, M.; Divekar, D.P.; Beaton, L.; Hogan, J.J.; et al. Foxp3+ follicular regulatory T cells control the germinal center response. Nat. Med. 2011, 17, 975–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lages, C.S.; Suffia, I.; Velilla, P.A.; Huang, B.; Warshaw, G.; Hildeman, D.A.; Belkaid, Y.; Chougnet, C. Functional regulatory T cells accumulate in aged hosts and promote chronic infectious disease reactivation. J. Immunol. 2008, 181, 1835–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strzepa, A.; Pritchard, K.A.; Dittel, B.N. Myeloperoxidase: A new player in autoimmunity. Cell Immunol. 2017, 317, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arulraj, T.; Binder, S.C.; Robert, P.A.; Meyer-Hermann, M. Germinal centre shutdown. Front. Immunol. 2021, 12, 705240. [Google Scholar] [CrossRef]
- Uhrlaub, J.L.; Pulko, V.; DeFilippis, V.R.; Broeckel, R.; Streblow, D.N.; Coleman, G.D.; Park, B.S.; Lindo, J.F.; Vickers, I.; Anzinger, J.J.; et al. Dysregulated TGF-β production underlies the age-related vulnerability to Chikungunya virus. PLoS Pathog. 2016, 12, e1005891. [Google Scholar] [CrossRef] [Green Version]
- Frasca, D.; Diaz, A.; Romero, M.; Landin, A.M.; Blomberg, B.B. High TNF-α levels in resting B cells negatively correlate with their response. Exp. Gerontol. 2014, 54, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Frasca, D.; Blomberg, B.B. Aging, cytomegalovirus (CMV) and influenza vaccine responses. Hum. Vaccin. Immunother. 2016, 12, 682–690. [Google Scholar] [CrossRef]
- Gibson, K.L.; Wu, Y.C.; Barnett, Y.; Duggan, O.; Vaughan, R.; Kondeatis, E.; Nilsson, B.O.; Wikby, A.; Kipling, D.; Dunn-Walters, D.K. B-cell diversity decreases in old age and is correlated with poor health status. Aging Cell 2009, 8, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabibian-Keissar, H.; Hazanov, L.; Schiby, G.; Rosenthal, N.; Rakovsky, A.; Michaeli, M.; Shahaf, G.L.; Hickman, Y.; Rosenblatt, K.; Melamed, D.; et al. Aging affects B-cell antigen receptor repertoire diversity in primary and secondary lymphoid tissues. Eur. J. Immunol. 2016, 46, 480–492. [Google Scholar] [CrossRef]
- Khurana, S.; Frasca, D.; Blomberg, B.; Golding, H. AID activity in B cells strongly correlates with polyclonal antibody affinity maturation in-vivo following pandemic 2009-H1N1 vaccination in humans. PLoS Pathog. 2012, 8, e1002920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.T.; Correia-Neves, M. Neutrophils and macrophages: The main partners of phagocyte cell systems. Front. Immunol. 2012, 3, 174. [Google Scholar] [CrossRef] [Green Version]
- Lok, L.S.C.; Clatworthy, M.R. Neutrophils in secondary lymphoid organs. Immunology 2021. epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Chtanova, T.; Schaeffer, M.; Han, S.J.; van Dooren, G.G.; Nollmann, M.; Herzmark, P.; Chan, S.W.; Satija, H.; Camfield, K.; Aaron, H.; et al. Dynamics of neutrophil migration in lymph nodes during infection. Immunity 2008, 29, 487–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abadie, V.; Badell, E.; Douillard, P.; Ensergueix, D.; Leenen, P.J.; Tanguy, M.; Fiette, L.; Saeland, S.; Gicquel, B.; Winter, N. Neutrophils rapidly migrate via lymphatics after Mycobacterium bovis BCG intradermal vaccination and shuttle live bacilli to the draining lymph nodes. Blood 2005, 106, 1843–1850. [Google Scholar] [CrossRef] [Green Version]
- Lämmermann, T.; Afonso, P.V.; Angermann, B.R.; Wang, J.M.; Kastenmüller, W.; Parent, C.A.; Germain, R.N. Neutrophil swarms require LTB4 and integrins at sites of cell death in vivo. Nature 2013, 498, 371–375. [Google Scholar] [CrossRef]
- John, A.L.S.; Ang, W.X.G.; Huang, M.N.; Kunder, C.A.; Chan, E.W.; Gunn, M.D.; Abraham, S.N. S1P-Dependent trafficking of intracellular yersinia pestis through lymph nodes establishes Buboes and systemic infection. Immunity 2014, 41, 440–450. [Google Scholar] [CrossRef] [Green Version]
- Hampton, H.R.; Bailey, J.; Tomura, M.; Brink, R.; Chtanova, T. Microbe-dependent lymphatic migration of neutrophils modulates lymphocyte proliferation in lymph nodes. Nat. Commun. 2015, 6, 7139. [Google Scholar] [CrossRef]
- Kamenyeva, O.; Boularan, C.; Kabat, J.; Cheung, G.Y.; Cicala, C.; Yeh, A.J.; Chan, J.L.; Periasamy, S.; Otto, M.; Kehrl, J.H. Neutrophil recruitment to lymph nodes limits local humoral response to Staphylococcus aureus. PLoS Pathog. 2015, 11, e1004827. [Google Scholar] [CrossRef] [PubMed]
- de Castro Pinho, J.; Förster, R. Lymph-derived neutrophils primarily locate to the subcapsular and medullary sinuses in resting and inflamed lymph nodes. Cells 2021, 10, 1486. [Google Scholar] [CrossRef] [PubMed]
- Abadie, V.; Bonduelle, O.; Duffy, D.; Parizot, C.; Verrier, B.; Combadière, B. Original encounter with antigen determines antigen-presenting cell imprinting of the quality of the immune response in mice. PLoS ONE 2009, 4, e8159. [Google Scholar] [CrossRef] [PubMed]
- Brackett, C.M.; Muhitch, J.B.; Evans, S.S.; Gollnick, S.O. IL-17 promotes neutrophil entry into tumor-draining lymph nodes following induction of sterile inflammation. J. Immunol. 2013, 191, 4348–4357. [Google Scholar] [CrossRef] [Green Version]
- Brubaker, A.L.; Rendon, J.L.; Ramirez, L.; Choudhry, M.A.; Kovacs, E.J. Reduced neutrophil chemotaxis and infiltration contributes to delayed resolution of cutaneous wound infection with advanced age. J. Immunol. 2013, 190, 1746–1757. [Google Scholar] [CrossRef] [Green Version]
- Sapey, E.; Greenwood, H.; Walton, G.; Mann, E.; Love, A.; Aaronson, N.; Insall, R.H.; Stockley, R.A.; Lord, J.M. Phosphoinositide 3-kinase inhibition restores neutrophil accuracy in the elderly: Toward targeted treatments for immunosenescence. Blood 2014, 123, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Nomellini, V.; Brubaker, A.L.; Mahbub, S.; Palmer, J.L.; Gomez, C.R.; Kovacs, E.J. Dysregulation of neutrophil CXCR2 and pulmonary endothelial ICAM-1 promotes age-related pulmonary inflammation. Aging Dis. 2012, 3, 234–247. [Google Scholar]
- Shaw, A.C.; Joshi, S.; Greenwood, H.; Panda, A.; Lord, J.M. Aging of the innate immune system. Curr. Opin. Immunol. 2010, 22, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Tomay, F.; Wells, K.; Duong, L.; Tsu, J.W.; Dye, D.E.; Radley-Crabb, H.G.; Grounds, M.D.; Shavlakadze, T.; Metharom, P.; Nelson, D.J.; et al. Aged neutrophils accumulate in lymphoid tissues from healthy elderly mice and infiltrate T- and B-cell zones. Immunol. Cell Biol. 2018, 96, 831–840. [Google Scholar] [CrossRef]
- Buckley, C.D.; Ross, E.A.; McGettrick, H.M.; Osborne, C.E.; Haworth, O.; Schmutz, C.; Stone, P.C.; Salmon, M.; Matharu, N.M.; Vohra, R.K.; et al. Identification of a phenotypically and functionally distinct population of long-lived neutrophils in a model of reverse endothelial migration. J. Leukoc. Biol. 2006, 79, 303–311. [Google Scholar] [CrossRef]
- Kuni-Eda, Y.; Okabe, M.; Kurosawa, M.; Maekawa, I.; Higuchi, M.; Kawamura, M.; Morioka, M.; Suzuki, S.; Ohmura, T.; Fujimoto, N.; et al. Effects of rhG-CSF on infection complications and impaired function of neutrophils secondary to chemotherapy for non-Hodgkin’s lymphoma. Leuk. Lymphoma 1995, 16, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Wenisch, C.; Patruta, S.; Daxböck, F.; Krause, R.; Hörl, W. Effect of age on human neutrophil function. J. Leukoc. Biol. 2000, 67, 40–45. [Google Scholar] [CrossRef]
- Andrews, T.; Sullivan, K.E. Infections in patients with inherited defects in phagocytic function. Clin. Microbiol. Rev. 2003, 16, 597–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukamoto, K.; Machida, K. Effects of life events and stress on neutrophil functions in elderly men. Immun. Ageing 2012, 9, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stebegg, M.; Kumar, S.D.; Silva-Cayetano, A.; Fonseca, V.R.; Linterman, M.A.; Graca, L. Regulation of the germinal center response. Front. Immunol. 2018, 9, 2469. [Google Scholar] [CrossRef] [Green Version]
- Denton, A.E.; Linterman, M.A. Stromal networking: Cellular connections in the germinal centre. Curr. Opin. Immunol. 2017, 45, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, I.; Sakurada, A.; Murakami, G.; Suzuki, D.; Sato, M.; Kohama, G.I. Comparative histology of lymph nodes from aged animals and humans with special reference to the proportional areas of the nodal cortex and sinus. Ann. Anat. 2004, 186, 337–347. [Google Scholar] [CrossRef]
- Lefebvre, J.S.; Masters, A.R.; Hopkins, J.W.; Haynes, L. Age-related impairment of humoral response to influenza is associated with changes in antigen specific T follicular helper cell responses. Sci. Rep. 2016, 6, 25051. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurty, A.T.; Turley, S.J. Lymph node stromal cells: Cartographers of the immune system. Nat. Immunol. 2020, 21, 369–380. [Google Scholar] [CrossRef]
- Duan, Y.Q.; Xia, M.H.; Ren, L.; Zhang, Y.F.; Ao, Q.L.; Xu, S.P.; Kuang, D.; Liu, Q.; Yan, B.; Zhou, Y.W.; et al. Deficiency of Tfh cells and germinal center in deceased COVID-19 patients. Curr. Med. Sci. 2020, 40, 618–624. [Google Scholar] [CrossRef]


| Cells | Young Lymph Node | Old Lymph Node |
|---|---|---|
T lymphocytes |
|
|
B lymphocytes |
|
|
Neutrophils |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cakala-Jakimowicz, M.; Kolodziej-Wojnar, P.; Puzianowska-Kuznicka, M. Aging-Related Cellular, Structural and Functional Changes in the Lymph Nodes: A Significant Component of Immunosenescence? An Overview. Cells 2021, 10, 3148. https://doi.org/10.3390/cells10113148
Cakala-Jakimowicz M, Kolodziej-Wojnar P, Puzianowska-Kuznicka M. Aging-Related Cellular, Structural and Functional Changes in the Lymph Nodes: A Significant Component of Immunosenescence? An Overview. Cells. 2021; 10(11):3148. https://doi.org/10.3390/cells10113148
Chicago/Turabian StyleCakala-Jakimowicz, Marta, Paulina Kolodziej-Wojnar, and Monika Puzianowska-Kuznicka. 2021. "Aging-Related Cellular, Structural and Functional Changes in the Lymph Nodes: A Significant Component of Immunosenescence? An Overview" Cells 10, no. 11: 3148. https://doi.org/10.3390/cells10113148