
Development of BCR-ABL1 Transgenic Zebrafish Model Reproducing Chronic Myeloid Leukemia (CML) Like-Disease and Providing a New Insight into CML Mechanisms


 , , , , ,
, , , , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatic Analysis of bcr and abl1
2.2. Animals Husbandry
2.3. Microinjection
2.4. Generation of the pToL BCR-ABL1 pUAS-CFP/Hsp70-gal4 Transgenic Line
2.5. Activation of the Transgenic Promoter hsp70
2.6. Whole-Mount in Situ Hybridization (WISH) Analysis
2.7. RNA Extraction and Retrotrascription
2.8. Digital PCR BCR-ABL1 Transcript Quantification
2.9. Real Time PCR Transcripts Quantification
2.10. Acridine Orange Staining
2.11. Cell Proliferation Assays H3 Immunostaining and BrdU Immunostaining
2.12. Blood Extraction
2.13. Imaging
2.14. Histomorphology Analysis
2.15. Quantification of Fluorescence Intensity by ZF-Mapper Application
2.16. Statistical Analysis
3. Results
3.1. Generation of BCR-ABL1 Transgenic Line
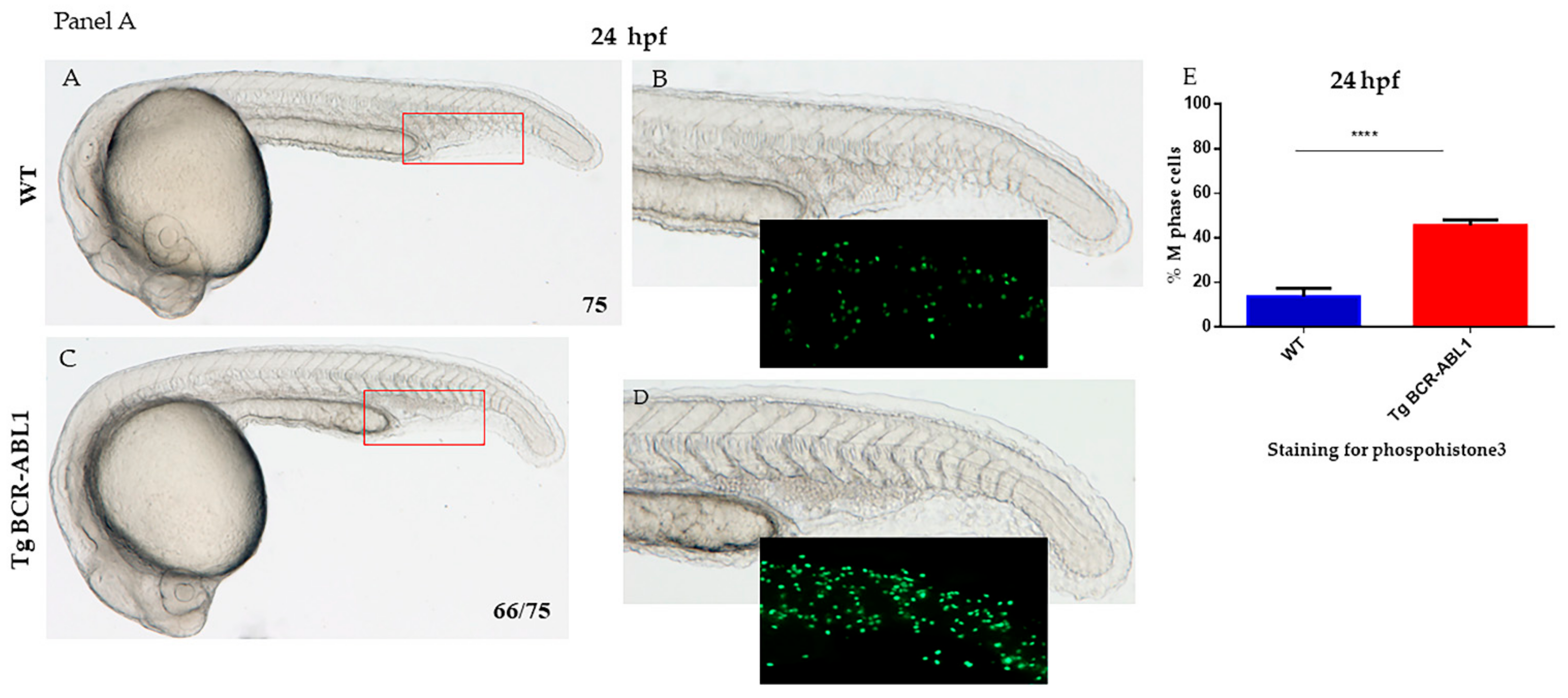
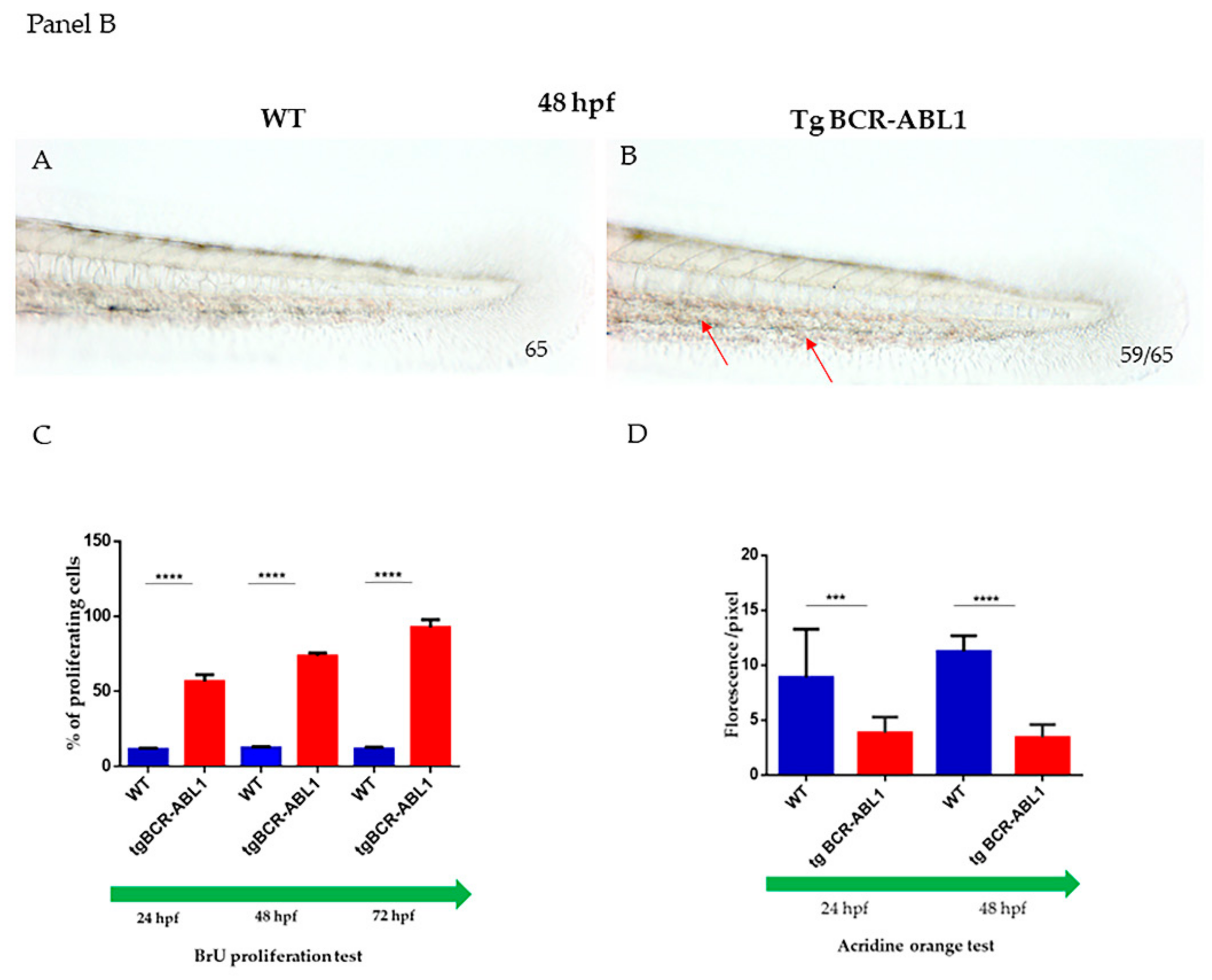
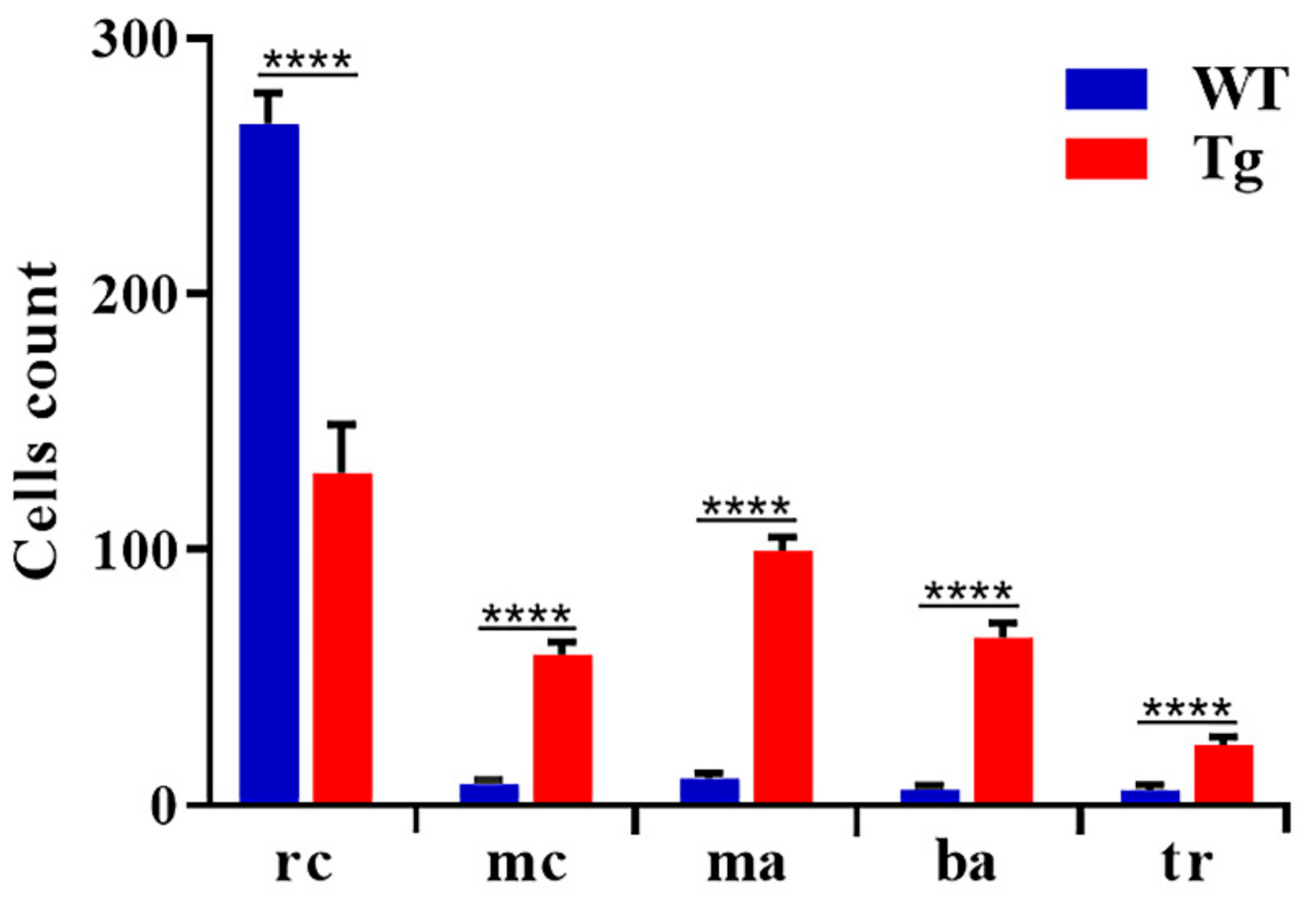
3.2. Induced Expression of BCR/ABL1 Increased the Number of Myeloid Cells during Embryonic Development
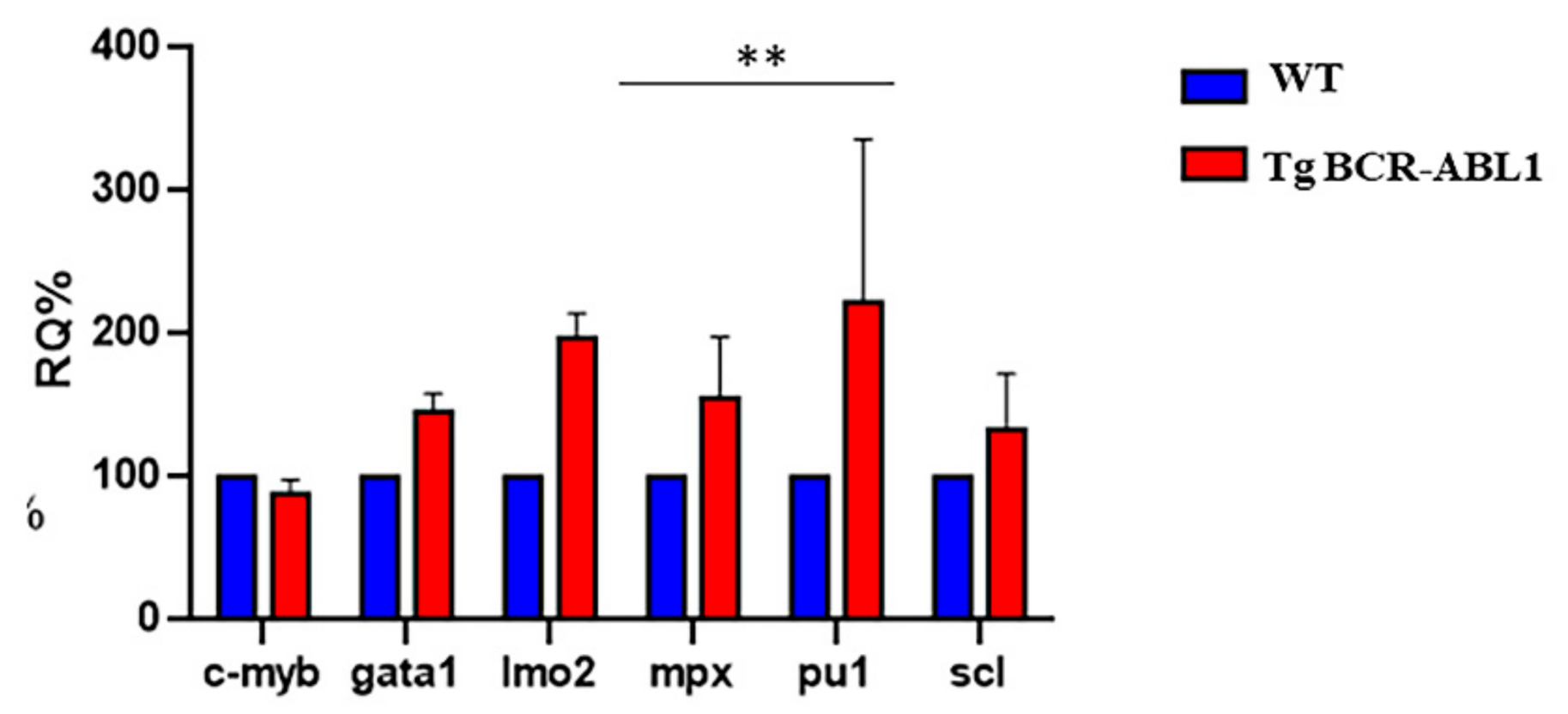
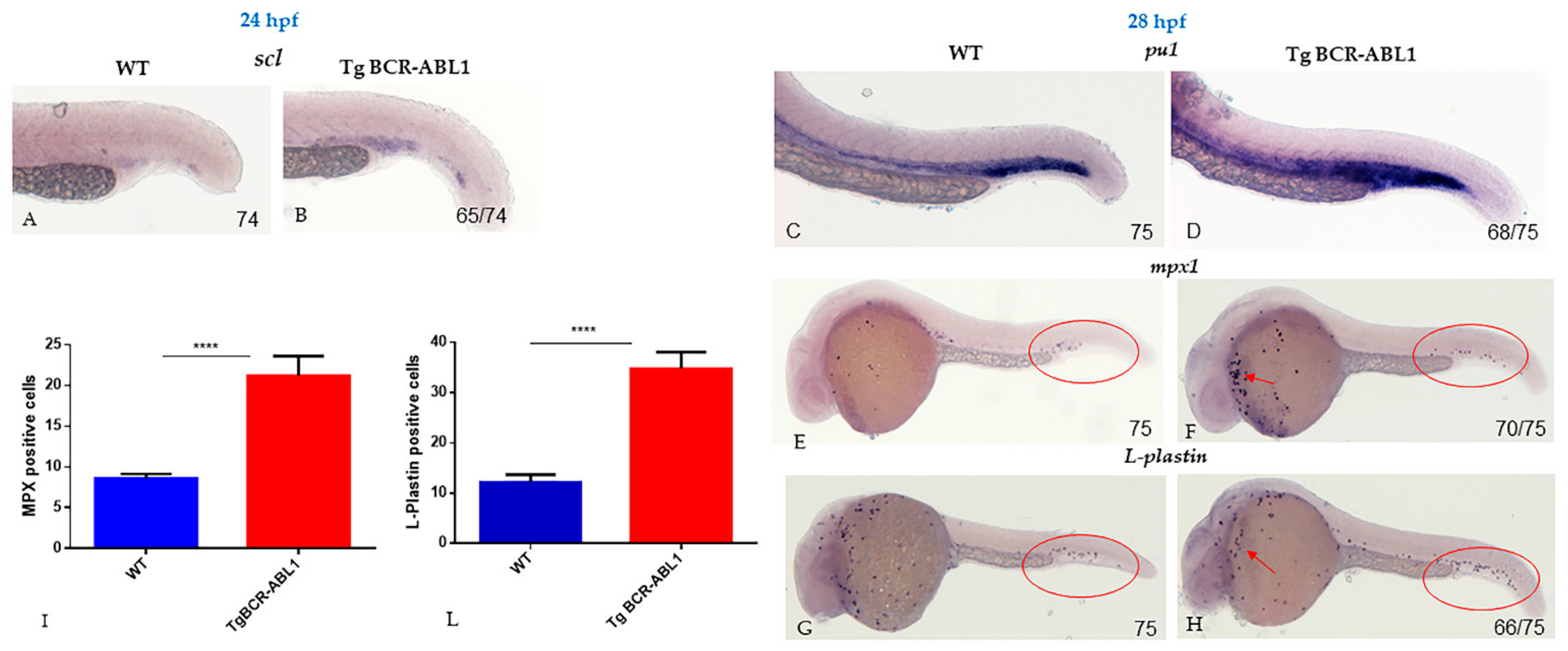
3.3. BCR-ABL Deregulates Lineage-Specific Hematopoietic Transcription Factors
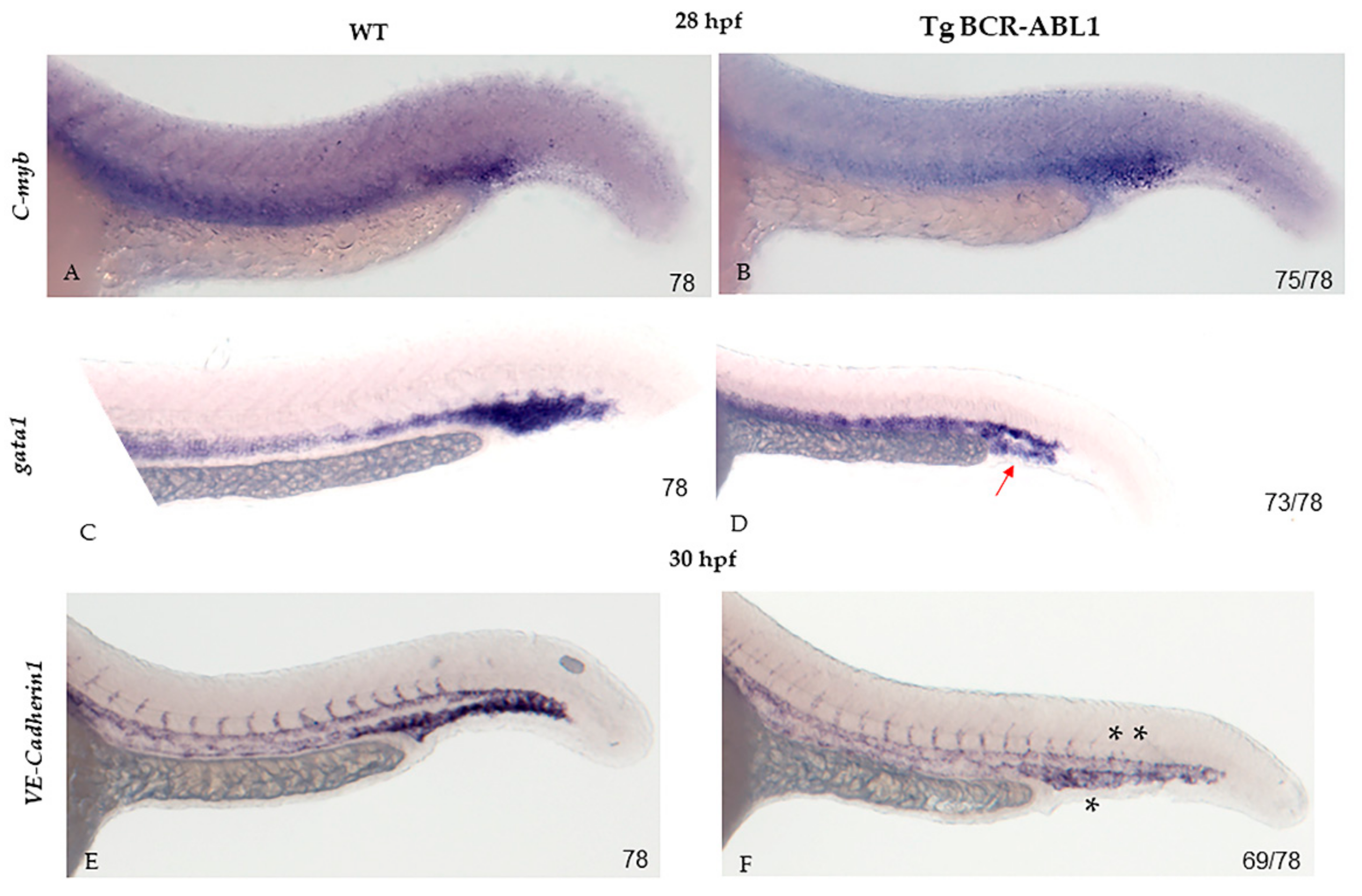
3.4. BCR-ABL1 Down-Regulates Erythropoiesis and Neovascularization
3.5. Peripheral Blood Analysis in Tg BCR-ABL1
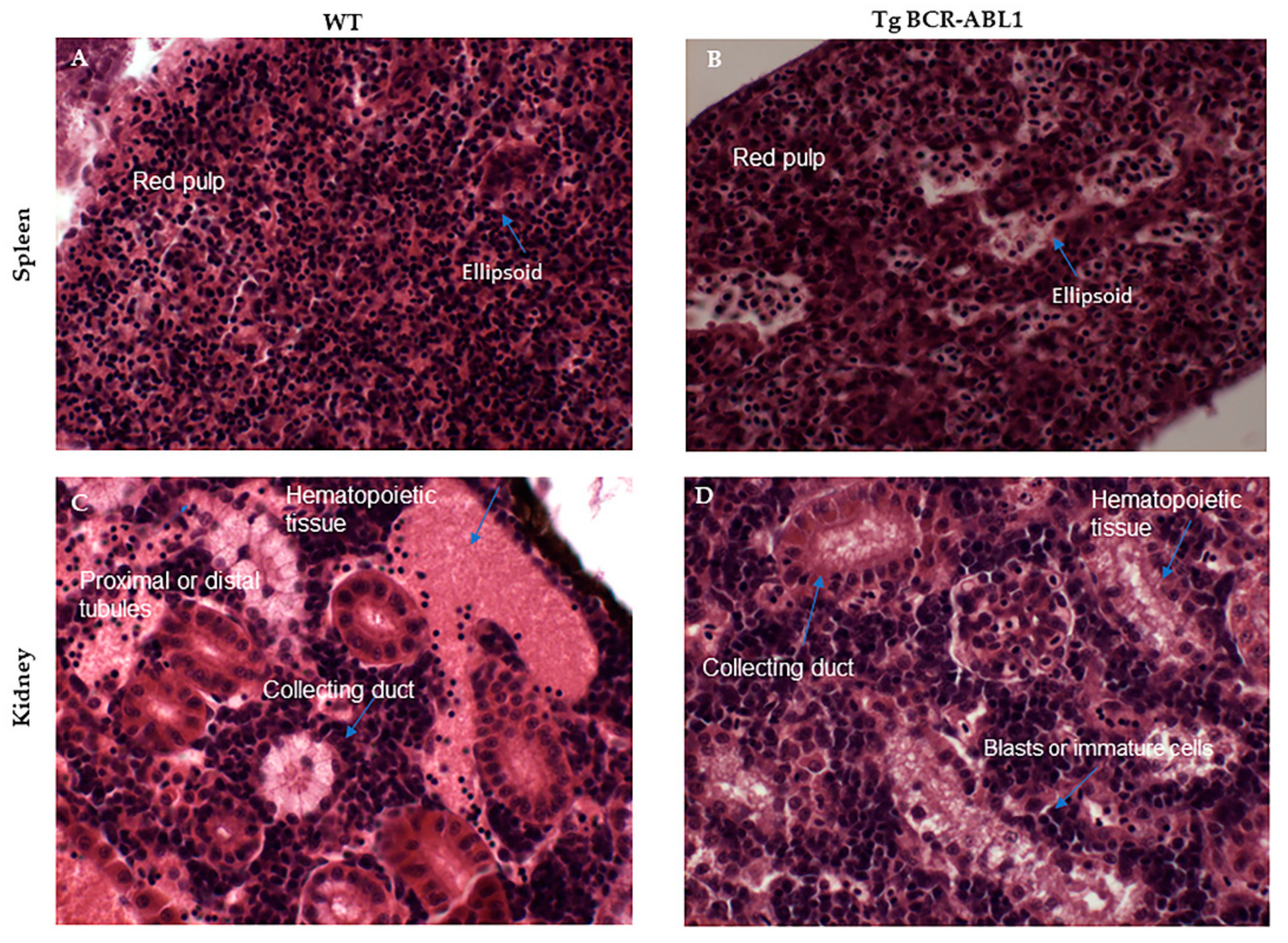
3.6. Histologic and Phenotypic Analysis of Hematopoietic Organs in Tg BCR-ABL1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Menke, A.L.; Spitsbergen, J.M.; Wolterbeek, A.P.M.; Woutersen, R.A. Normal Anatomy and Histology of the Adult Zebrafish. Toxicol. Pathol. 2011, 39, 759–775. [Google Scholar] [CrossRef] [PubMed]
- White, R.M. Cross-species oncogenomics using zebrafish models of cancer. Curr. Opin. Genet. Dev. 2015, 30, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Zizioli, D.; Mione, M.; Varinelli, M.; Malagola, M.; Bernardi, S.; Alghisi, E.; Borsani, G.; Finazzi, D.; Monti, E.; Presta, M.; et al. Zebrafish disease models in hematology: Highlights on biological and translational impact. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 620–633. [Google Scholar] [CrossRef]
- Robertson, A.L.; Avagyan, S.; Gansner, J.M.; Zon, L.I. Understanding the regulation of vertebrate hematopoiesis and blood disorders—big lessons from a small fish. FEBS Lett. 2016, 590, 4016–4033. [Google Scholar] [CrossRef] [Green Version]
- Gore, A.V.; Pillay, L.M.; Galanternik, M.V.; Weinstein, B.M. The zebrafish: A fintastic model for hematopoietic development and disease. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e312. [Google Scholar] [CrossRef] [PubMed]
- Skayneh, H.; Jishi, B.; Hleihel, R.; Hamieh, M.; Darwiche, N.; Bazarbachi, A.; El Sabban, M.; El Hajj, H. A Critical Review of Animal Models Used in Acute Myeloid Leukemia Pathophysiology. Genes 2019, 10, 614. [Google Scholar] [CrossRef] [Green Version]
- Alghisi, E.; Distel, M.; Malagola, M.; Anelli, V.; Santoriello, C.; Herwig, L.; Krudewig, A.; Henkel, C.V.; Russo, D.; Mione, M.C. Targeting oncogene expression to endothelial cells induces proliferation of the myelo-erythroid lineage by repressing the notch pathway. Leukemia 2013, 27, 2229–2241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, A.A.; Park, G.; Frazer, J.K. Tackling Acute Lymphoblastic Leukemia—One Fish at a Time. Int. J. Mol. Sci. 2019, 20, 5313. [Google Scholar] [CrossRef] [Green Version]
- Frazer, J.K.; Meeker, N.D.; Rudner, L.; Bradley, D.F.; Smith, A.C.H.; Demarest, B.; Joshi, D.; Locke, E.E.; Hutchinson, S.A.; Tripp, S.; et al. Heritable T-cell malignancy models established in a zebrafish phenotypic screen. Leukemia 2009, 23, 1825–1835. [Google Scholar] [CrossRef] [Green Version]
- Auer, R.L.; Riaz, S.; Cotter, F.E. The 13q and 11q B-cell chronic lymphocytic leukaemia-associated regions derive from a common ancestral region in the zebrafish. Br. J. Haematol. 2007, 137, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Langenau, D.M.; Madge, J.A.; Quinkertz, A.; Gutierrez, A.; Neuberg, D.S.; Kanki, J.P.; Look, A.T. Heat-shock induction of T-cell lymphoma/leukaemia in conditional Cre/lox-regulated transgenic zebrafish. Br. J. Haematol. 2007, 138, 169–175. [Google Scholar] [CrossRef]
- Simonetti, G.; Padella, A.; Valle, I.F.D.; Fontana, M.C.; Fonzi, E.; Bruno, S.; Baldazzi, C.; Guadagnuolo, V.; Manfrini, M.; Ferrari, A.; et al. Aneuploid acute myeloid leukemia exhibits a signature of genomic alterations in the cell cycle and protein degradation machinery. Cancer 2018, 125, 712–725. [Google Scholar] [CrossRef] [Green Version]
- Papaemmanuil, E.; Gerstung, M.; Bullinger, L.; Gaidzik, V.I.; Paschka, P.; Roberts, N.D.; Potter, N.E.; Heuser, M.; Thol, F.; Bolli, N.; et al. Genomic Classification and Prognosis in Acute Myeloid Leukemia. N. Engl. J. Med. 2016, 374, 2209–2221. [Google Scholar] [CrossRef]
- Russo, D.; Garcia-Gutierrez, J.V.; Soverini, S.; Baccarani, M. Chronic Myeloid Leukemia Prognosis and Therapy: Criticisms and Perspectives. J. Clin. Med. 2020, 9, 1709. [Google Scholar] [CrossRef]
- Faderl, S.; Talpaz, M.; Estrov, Z.; O’Brien, S.; Kurzrock, R.; Kantarjian, H.M. The Biology of Chronic Myeloid Leukemia. N. Engl. J. Med. 1999, 341, 164–172. [Google Scholar] [CrossRef]
- Shanmuganathan, N.; Branford, S. The Hidden Pathogenesis of CML: Is BCR-ABL1 the First Event? Curr. Hematol. Malign. Rep. 2019, 14, 501–506. [Google Scholar] [CrossRef]
- Kang, Z.-J.; Liu, Y.-F.; Xu, L.-Z.; Long, Z.-J.; Huang, D.; Yang, Y.; Liu, B.; Feng, J.-X.; Pan, Y.-J.; Yan, J.-S.; et al. The Philadelphia chromosome in leukemogenesis. Chin. J. Cancer 2016, 35, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Quintás-Cardama, A.; Cortes, J. Molecular biology of bcr-abl1–positive chronic myeloid leukemia. Blood 2009, 113, 1619–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.; Medeiros, L.J.; Hu, S. Chronic Myeloid Leukemia: Beyond BCR-ABL1. Curr. Hematol. Malign. Rep. 2018, 13, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Trojani, A.; Pungolino, E.; Molin, A.D.; Lodola, M.; Rossi, G.; D’Adda, M.; Perego, A.; Elena, C.; Turrini, M.; Borin, L.; et al. Nilotinib interferes with cell cycle, ABC transporters and JAK-STAT signaling pathway in CD34+/lin- cells of patients with chronic phase chronic myeloid leukemia after 12 months of treatment. PLoS ONE 2019, 14, e0218444. [Google Scholar] [CrossRef] [Green Version]
- Bertacchini, J.; Ketabchi, N.; Mediani, L.; Capitani, S.; Marmiroli, S.; Saki, N. Inhibition of Ras-mediated signaling pathways in CML stem cells. Cell. Oncol. 2015, 38, 407–418. [Google Scholar] [CrossRef]
- Wagle, M.; Eiring, A.M.; Wongchenko, M.; Lu, S.; Guan, Y.; Wang, Y.; Lackner, M.; Amler, L.; Hampton, G.; Deininger, M.W.; et al. A role for FOXO1 in BCR–ABL1-independent tyrosine kinase inhibitor resistance in chronic myeloid leukemia. Leukemia 2016, 30, 1493–1501. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Dong, F.; Luan, C.; Zhang, X.; Shao, H.; Liu, J.; Sun, C. Overexpression of CCAAT Enhancer-Binding Protein α Inhibits the Growth of K562 Cells via the Foxo3a-Bim Pathway. Acta Haematol. 2016, 136, 65–70. [Google Scholar] [CrossRef]
- José-Enériz, E.S.; Román-Gómez, J.; Cordeu, L.; Ballestar, E.; Garate, L.; Andreu, E.J.; Isidro, I.; Guruceaga, E.; Jiménez-Velasco, A.; Heiniger, A.; et al. BCR-ABL1-induced expression of HSPA8 promotes cell survival in chronic myeloid leukaemia. Br. J. Haematol. 2008, 142, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Koschmieder, S.; Vetrie, D. Epigenetic dysregulation in chronic myeloid leukaemia: A myriad of mechanisms and therapeutic options. Semin. Cancer Biol. 2018, 51, 180–197. [Google Scholar] [CrossRef] [PubMed]
- Vardiman, J.W. Chronic Myelogenous Leukemia, BCR-ABL1+. Am. J. Clin. Pathol. 2009, 132, 250–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skorski, T. Genetic Mechanisms of Chronic Myeloid Leukemia Blastic Transformation. Curr. Hematol. Malign-Rep. 2012, 7, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Ma, N.; Chen, X.; Zhang, Y.; Zhang, W. An overview of chronic myeloid leukemia and its animal models. Sci. China Life Sci. 2015, 58, 1202–1208. [Google Scholar] [CrossRef]
- García-Gutiérrez, V.; Hernández-Boluda, J.C. Tyrosine Kinase Inhibitors Available for Chronic Myeloid Leukemia: Efficacy and Safety. Front. Oncol. 2019, 9, 603. [Google Scholar] [CrossRef] [Green Version]
- Braun, T.P.; Eide, C.A.; Druker, B.J. Response and Resistance to BCR-ABL1-Targeted Therapies. Cancer Cell 2020, 37, 530–542. [Google Scholar] [CrossRef]
- Saglio, G.; Jabbour, E. First-line therapy for chronic phase CML: Selecting the optimal BCR-ABL1-targeted TKI. Leuk. Lymphoma 2017, 59, 1523–1538. [Google Scholar] [CrossRef] [PubMed]
- Malagola, M.; Breccia, M.; Skert, C.; Cancelli, V.; Soverini, S.; Iacobucci, I.; Cattina, F.; Liberati, A.M.; Tiribelli, M.; Annunziata, M.; et al. Long term outcome of Ph+ CML patients achieving complete cytogenetic remission with interferon based therapy moving from interferon to imatinib era. Am. J. Hematol. 2013, 89, 119–124. [Google Scholar] [CrossRef]
- Wolff, N.C.; Veach, D.R.; Tong, W.P.; Bornmann, W.G.; Clarkson, B.; Ilaria, R.L. PD166326, a novel tyrosine kinase inhibitor, has greater antileukemic activity than imatinib mesylate in a murine model of chronic myeloid leukemia. Blood 2005, 105, 3995–4003. [Google Scholar] [CrossRef] [Green Version]
- Schubert, C.; Chatain, N.; Braunschweig, T.; Schemionek, M.; Feldberg, K.; Hoffmann, M.; Dufva, O.; Mustjoki, S.; Brummendorf, T.H.; Koschmieder, S. The SCLtTAxBCR-ABL transgenic mouse model closely reflects the differential effects of dasatinib on normal and malignant hematopoiesis in chronic phase-CML patients. Oncotarget 2017, 8, 34736–34749. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Hao, S.; Liu, Y.; Pang, Y.; Ma, S.; Dong, F.; Xu, J.; Zheng, G.; Li, S.; Yuan, W.; et al. Leukemic marrow infiltration reveals a novel role for Egr3 as a potent inhibitor of normal hematopoietic stem cell proliferation. Blood 2015, 126, 1302–1313. [Google Scholar] [CrossRef] [PubMed]
- Dash, A.B.; Williams, I.R.; Kutok, J.L.; Tomasson, M.H.; Anastasiadou, E.; Lindahl, K.; Li, S.; Van Etten, R.A.; Borrow, J.; Housman, D.; et al. A murine model of CML blast crisis induced by cooperation between BCR/ABL and NUP98/HOXA9. Proc. Natl. Acad. Sci. USA 2002, 99, 7622–7627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, C.; Chen, Y.; Yang, Z.; Zhang, H.; Osterby, L.; Rosmarin, A.G.; Li, S. PTEN is a tumor suppressor in CML stem cells and BCR-ABL–induced leukemias in mice. Blood 2010, 115, 626–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Outa, A.; Abubaker, D.; Bazarbachi, A.; El Sabban, M.; Shirinian, M.; Nasr, R. Validation of a drosophila model of wild-type and T315I mutated BCR-ABL1 in chronic myeloid leukemia: An effective platform for treatment screening. Haematologica 2020, 105, 387–397. [Google Scholar] [CrossRef]
- Bernardoni, R.; Giordani, G.; Signorino, E.; Monticelli, S.; Messa, F.; Pradotto, M.; Rosso, V.; Bracco, E.; Giangrande, A.; Perini, G.; et al. A new BCR-ABL1 Drosophila model as a powerful tool to elucidate the pathogenesis and progression of chronic myeloid leukemia. Haematologica 2018, 104, 717–728. [Google Scholar] [CrossRef]
- Xu, M.; Ye, Y.; Ye, Z.; Xu, S.; Liu, W.; Xu, J.; Zhang, Y.; Liu, Q.; Huang, Z.; Zhang, W. Human BCR/ABL1 induces chronic myeloid leukemia-like disease in zebrafish. Haematologica 2019, 105, 674–686. [Google Scholar] [CrossRef] [PubMed]
- Russo, D.; Malagola, M.; Skert, C.; Cancelli, V.; Turri, D.; Pregno, P.; Bergamaschi, M.; Fogli, M.; Testoni, N.; De Vivo, A.; et al. Managing chronic myeloid leukaemia in the elderly with intermittent imatinib treatment. Blood Cancer J. 2015, 5, e347. [Google Scholar] [CrossRef] [Green Version]
- Hochhaus, A.; Baccarani, M.; Silver, R.T.; Schiffer, C.; Apperley, J.F.; Cervantes, F.; Clark, R.E.; Cortes, J.E.; Deininger, M.W.; Guilhot, F.; et al. European LeukemiaNet 2020 recommendations for treating chronic myeloid leukemia. Leukemia 2020, 34, 966–984. [Google Scholar] [CrossRef] [Green Version]
- Baccarani, M.; Abruzzese, E.; Accurso, V.; Albano, F.; Annunziata, M.; Barulli, S.; Beltrami, G.; Bergamaschi, M.; Binotto, G.; Bocchia, M.; et al. Managing chronic myeloid leukemia for treatment-free remission: A proposal from the GIMEMA CML WP. Blood Adv. 2019, 3, 4280–4290. [Google Scholar] [CrossRef] [Green Version]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Takeda, H.; Kawakami, N.; Kobayashi, M.; Matsuda, N.; Mishina, M. A Transposon-Mediated Gene Trap Approach Identifies Developmentally Regulated Genes in Zebrafish. Dev. Cell 2004, 7, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Moro, E.; Vettori, A.; Porazzi, P.; Schiavone, M.; Rampazzo, E.; Casari, A.; Ek, O.; Facchinello, N.; Astone, M.; Zancan, I.; et al. Generation and application of signaling pathway reporter lines in zebrafish. Mol. Genet. Genom. 2013, 288, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Facchinello, N.; Schiavone, M.; Vettori, A.; Argenton, F.; Tiso, N. Monitoring Wnt Signaling in Zebrafish Using Fluorescent Biosensors. Wnt Signal. 2016, 1481, 81–94. [Google Scholar]
- Kawakami, K.; Asakawa, K.; Hibi, M.; Itoh, M.; Muto, A.; Wada, H. Gal4 Driver Transgenic Zebrafish: Powerful Tools to Study Developmental Biology, Organogenesis, and Neuroscience. Adv. Genet. 2016, 95, 65–87. [Google Scholar]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2007, 3, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Soverini, S.; Bernardi, S.; Galimberti, S. Molecular Testing in CML between Old and New Methods: Are We at a Turning Point? J. Clin. Med. 2020, 9, 3865. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, S.; Malagola, M.; Zanaglio, C.; Polverelli, N.; Dereli Eke, E.; D’Adda, M.; Farina, M.; Bucelli, C.; Scaffidi, L.; Toffoletti, E.; et al. Digital PCR improves the quantitation of DMR and the selection of CML candidates to TKIs discontinuation. Cancer Med. 2019, 8, 2041–2055. [Google Scholar] [CrossRef] [PubMed]
- Zanaglio, C.; Bernardi, S.; Gandolfi, L.; Farina, M.; Re, F.; Polverelli, N.; Zollner, T.; Turra, A.; Morello, E.; Malagola, M.; et al. RT-qPCR versus Digital PCR: How Do They Impact Differently on Clinical Management of Chronic Myeloid Leukemia Patients? Case Rep. Oncol. 2020, 13, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, S.; Bonifacio, M.; Iurlo, A.; Zanaglio, C.; Tiribelli, M.; Binotto, G.; Abruzzese, E.; Russo, D. “Variant-specific discrepancy when quantitating BCR-ABL1 e13a2 and e14a2 transcripts using the Europe Against Cancer qPCR assay.” Is dPCR the key? Eur. J. Haematol. 2019, 103, 272–273. [Google Scholar] [CrossRef] [Green Version]
- Bernardi, S.; Ruggieri, G.; Malagola, M.; Cancelli, V.; Cattina, F.; Polverelli, N.; Zanaglio, C.; Perucca, S.; Re, F.; Montanelli, A. Digital PCR (Dpcr) a Step Forward to Detection and Quantification of Minimal Residual Disease (MRD) in Ph+/BCR-ABL1 Chronic Myeloid Leukemia (CML). J. Mol. Biomarkers Diagn. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Pagani, F.; Trivedi, A.; Khatri, D.; Zizioli, D.; Garrafa, E.; Mitola, S.; Finazzi, D. Silencing of pantothenate kinase 2 reduces endothelial cell angiogenesis. Mol. Med. Rep. 2018, 18, 4739–4746. [Google Scholar] [CrossRef] [Green Version]
- Perkins, B.D.; Nicholas, C.S.; Baye, L.M.; Link, B.A.; Dowling, J.E. dazed gene is necessary for late cell type development and retinal cell maintenance in the zebrafish retina. Dev. Dyn. 2005, 233, 680–694. [Google Scholar] [CrossRef]
- Jagadeeswaran, P.; Sheehan, J.P.; Craig, F.E.; Troyer, D. Identification and characterization of zebrafish thrombocytes. Br. J. Haematol. 1999, 107, 731–738. [Google Scholar] [CrossRef] [Green Version]
- Jagadeeswaran, P.; Sheehan, J.P. Analysis of Blood Coagulation in the Zebrafish. Blood Cells Mol. Dis. 1999, 25, 239–249. [Google Scholar] [CrossRef]
- Yamamoto, D.; Sato, D.; Nakayama, H.; Nakagawa, Y.; Shimada, Y. ZF-Mapper: Simple and Complete Freeware for Fluorescence Quantification in Zebrafish Images. Zebrafish 2019, 16, 233–239. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Davidson, A.J.; Zon, L.I. The “definitive” (and ’primitive’) guide to zebrafish hematopoiesis. Oncogene 2004, 23, 7233–7246. [Google Scholar] [CrossRef] [Green Version]
- Findakly, D.; Arslan, W. Clinical Features and Outcomes of Patients with Chronic Myeloid Leukemia Presenting With Isolated Thrombocytosis: A Systematic Review and a Case from Our Institution. Cureus 2020, 12. [Google Scholar] [CrossRef]
- Traver, D.; Herbomel, P.; Patton, E.E.; Murphey, R.D.; Yoder, J.A.; Litman, G.W.; Catic, A.; Amemiya, C.T.; Zon, L.I.; Trede, N.S. The Zebrafish as a Model Organism to Study Development of the Immune System. Adv. Immunol. 2003, 81, 253–330. [Google Scholar] [PubMed]
- Danilova, N.; Steiner, L.A. B cells develop in the zebrafish pancreas. Proc. Natl. Acad. Sci. USA 2002, 99, 13711–13716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gjini, E.; Mansour, M.; He, S.; Nguyen, A.; Ko, M.; Sander, J.D.; Joung, J.; Zhou, Y.I.; Zon, L.; Rao, A.; et al. A zebrafish model of myelodysplastic syndrome produced through tet2 genomic editing. Leuk. Res. 2015, 39, S11. [Google Scholar] [CrossRef] [Green Version]
- Payne, E.; Look, T. Zebrafish modelling of leukaemias. Br. J. Haematol. 2009, 146, 247–256. [Google Scholar] [CrossRef]
- Rasighaemi, P.; Basheer, F.; Liongue, C.; Ward, A.C. Zebrafish as a model for leukemia and other hematopoietic disorders. J. Hematol. Oncol. 2015, 8. [Google Scholar] [CrossRef] [Green Version]
- Kurzrock, R.; Kantarjian, H.M.; Druker, B.J.; Talpaz, M. Philadelphia ChromosomePositive Leukemias: From Basic Mechanisms to Molecular Therapeutics. Ann. Intern. Med. 2003, 138, 819–830. [Google Scholar] [CrossRef]
- Rowley, P.T.; Keng, P.C.; Kosciolek, B.A. The effect of bcr-abl antisense oligonucleotide on DNA synthesis and apoptosis in K562 chronic myeloid leukemia cells. Leuk. Res. 1996, 20, 473–480. [Google Scholar] [CrossRef]
- Lin, C.W.; Manshouri, T.; Jilani, I.; Neuberg, D.; Patel, K.; Kantarjian, H.; Andreeff, M.; Estrov, Z.; Beran, M.; Keating, M.; et al. Proliferation and apoptosis in acute and chronic leukemias and myelodysplastic syndrome. Leuk. Res. 2002, 26, 551–559. [Google Scholar] [CrossRef]
- Liao, E.C.; Paw, B.H.; Oates, A.C.; Pratt, S.J.; Postlethwait, J.H.; Zon, L.I. SCL/Tal-1 transcription factor acts downstream of cloche to specify hematopoietic and vascular progenitors in zebrafish. Genes Dev. 1998, 12, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, J.; Hagen, A.; Hsu, K.; Deng, M.; Liu, T.X.; Look, A.; Kanki, J.P. Interplay of Pu.1 and Gata1 Determines Myelo-Erythroid Progenitor Cell Fate in Zebrafish. Dev. Cell 2005, 8, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.A.; Ransom, D.G.; Pratt, S.J.; MacLennan, H.; Kieran, M.W.; Detrich, H.W., III; Vail, B.; Huber, T.L.; Paw, B.; Brownlie, A.J.; et al. The cloche and spadetail genes differentially affect hematopoiesis and vasculogenesis. Dev. Biol. 1998, 197, 248–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, C.M.; Kanki, J.P.; Rhodes, J.; Liu, T.X.; Paw, B.H.; Kieran, M.W.; Langenau, D.M.; Delahaye-Brown, A.; Zon, L.I.; Fleming, M.D.; et al. Myelopoiesis in the zebrafish, Danio rerio. Blood 2001, 98, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Oates, A.C.; Crowhurst, M.O.; Ward, A.C.; Layton, J.E. Morphologic and functional characterization of granulocytes and macrophages in embryonic and adult zebrafish. Blood 2001, 98, 3087–3096. [Google Scholar] [CrossRef]
- Cofre, J.; Menezes, J.R.L.; Pizzatti, L.; Abdelhay, E. Knock-down of Kaiso induces proliferation and blocks granulocytic differentiation in blast crisis of chronic myeloid leukemia. Cancer Cell Int. 2012, 12, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimamoto, T.; Ohyashiki, K.; Ohyashiki, J.H.; Kawakubo, K.; Fujimura, T.; Iwama, H.; Nakazawa, S.; Toyama, K. The expression pattern of erythrocyte/megakaryocyte-related transcription factors GATA-1 and the stem cell leukemia gene correlates with hematopoietic differentiation and is associated with outcome of acute myeloid leukemia. Blood 1995, 86, 3173–3180. [Google Scholar] [CrossRef] [Green Version]
- Bavaro, L.; Martelli, M.; Cavo, M.; Soverini, S. Mechanisms of Disease Progression and Resistance to Tyrosine Kinase Inhibitor Therapy in Chronic Myeloid Leukemia: An Update. Int. J. Mol. Sci. 2019, 20, 6141. [Google Scholar] [CrossRef] [Green Version]
- Bonifacio, M.; Stagno, F.; Scaffidi, L.; Krampera, M.; Di Raimondo, F. Management of Chronic Myeloid Leukemia in Advanced Phase. Front. Oncol. 2019, 9, 1132. [Google Scholar] [CrossRef] [Green Version]
- Iurlo, A.; Cattaneo, D.; Gianelli, U. Blast Transformation in Myeloproliferative Neoplasms: Risk Factors, Biological Findings, and Targeted Therapeutic Options. Int. J. Mol. Sci. 2019, 20, 1839. [Google Scholar] [CrossRef] [Green Version]
- Sangiorgio, V.F.I.; Arber, D.A.; Orazi, A. How I investigate chronic myelomonocytic leukemia. Int. J. Lab. Hematol. 2019, 42, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Eaves, C.; Cashman, J.; Eaves, A. Defective regulation of leukemic hematopoiesis in chronic myeloid leukemia. Leuk. Res. 1998, 22, 1085–1096. [Google Scholar] [CrossRef]
- Tang, Q.; Iyer, S.; Lobbardi, R.; Moore, J.C.; Chen, H.; Lareau, C.; Hebert, C.; Shaw, M.L.; Neftel, C.; Suva, M.L.; et al. Dissecting hematopoietic and renal cell heterogeneity in adult zebrafish at single-cell resolution using RNA sequencing. J. Exp. Med. 2017, 214, 2875–2887. [Google Scholar] [CrossRef] [PubMed]
- Stachura, D.L.; Reyes, J.R.; Bartunek, P.; Paw, B.H.; Zon, L.I.; Traver, D. Zebrafish kidney stromal cell lines support multilineage hematopoiesis. Blood 2009, 114, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Helbig, G.; Janikowska, A.; Kyrcz-Krzemien, S. Aggregates of pseudo-Gaucher cells after treatment of chronic myeloid leukemia in blastic phase. Int. J. Hematol. 2015, 101, 3–4. [Google Scholar] [CrossRef]
- Ridzuan Mohd Said, M.; Yap, E.; Fariza Wan Jamaluddin, W.; Abdul Wahid, F.S.; Shuib, S. A case of chronic myeloid leukaemia in blast transformation with leukemic ascites. Med. J. Malaysia 2016, 71, 85–86. [Google Scholar]
- Horny, H.-P.; Parwaresch, M.; Lennert, K. Bone marrow findings in systemic mastocytosis. Hum. Pathol. 1985, 16, 808–814. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligo | Sequence | Purpose |
|---|---|---|
| for1 BA cDNA | AGACTGTCCACAGCATTCCG | tg screening |
| rev1 BA cDNA | GCAACGAAAAGGTTGGGGTC | tg screening |
| for2 BA cDNA | GATGCTGACCAACTCGTGTG | tg screening |
| rev2 BA cDNA | GACCCGGAGCTTTTCACCTT | tg screening |
| for scl | CCGCTCGCCACTATTAACAG | real-time |
| rev scl | GTTCGTGAAAATCCGTCGC | real-time |
| for lmo2 | ACTACAAACTCGGCAGAAAGC | real-time |
| rev lmo2 | CACGCATGGTCATTTCAAAGG | real-time |
| for gata1 | TGAATGTGTGAATTGTGGTG | real-time |
| rev gata1 | ATTGCGTCTCCATAGTGTTG | real-time |
| for pu.1 | CCATTAGAGGTGTCCGATGAG | real-time |
| rev pu.1 | ACCAGATGCTGTCCTTCATG | real-time |
| for runx1 | CCCCGCCCACAGCCAGATTC | real-time |
| rev runx1 | GACGGGCGTGGGGGTGTAGGT | real-time |
| for c-myb | GGAAAGTGGAGCAAAGAAGGTTA | real-time |
| rev c-myb | TCGTGTAGTGTCTCTGGATAG | real-time |
| for mpx | TGATGTTTGGTTAGGAGGTG | real-time |
| rev mpx | GAGCTGTTTTCTGTTTGGTG | real-time |
| Classification | Mature Red Cells | Myeloid Cells | Macrophages | Blasts Aggregates | Trombocytes |
|---|---|---|---|---|---|
| WT | 57.6 ± 0.99 | 1.7 ± 0.98 | 2.5 ± 0.96 | 1.5 ± 0.07 | 0.13 ± 0.05 |
| Tg BCR-ABL1 | 28.4 ± 0.97 | 10.4 ± 1.5 | 23.9 ± 1.6 | 15.2 ± 0.98 | 1.7 ± 1.3 |
| p values | p < 0.0001 | p < 0.0001 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zizioli, D.; Bernardi, S.; Varinelli, M.; Farina, M.; Mignani, L.; Bosio, K.; Finazzi, D.; Monti, E.; Polverelli, N.; Malagola, M.; et al. Development of BCR-ABL1 Transgenic Zebrafish Model Reproducing Chronic Myeloid Leukemia (CML) Like-Disease and Providing a New Insight into CML Mechanisms. Cells 2021, 10, 445. https://doi.org/10.3390/cells10020445
Zizioli D, Bernardi S, Varinelli M, Farina M, Mignani L, Bosio K, Finazzi D, Monti E, Polverelli N, Malagola M, et al. Development of BCR-ABL1 Transgenic Zebrafish Model Reproducing Chronic Myeloid Leukemia (CML) Like-Disease and Providing a New Insight into CML Mechanisms. Cells. 2021; 10(2):445. https://doi.org/10.3390/cells10020445
Chicago/Turabian StyleZizioli, Daniela, Simona Bernardi, Marco Varinelli, Mirko Farina, Luca Mignani, Katia Bosio, Dario Finazzi, Eugenio Monti, Nicola Polverelli, Michele Malagola, and et al. 2021. "Development of BCR-ABL1 Transgenic Zebrafish Model Reproducing Chronic Myeloid Leukemia (CML) Like-Disease and Providing a New Insight into CML Mechanisms" Cells 10, no. 2: 445. https://doi.org/10.3390/cells10020445