
Chiral Synthesis of 3-Amino-1-phenylbutane by a Multi-Enzymatic Cascade System


Department of Chemical, Biological and Environmental Engineering, Universitat Autònoma de Barcelona, 08193 Bellaterra, Spain
*
Author to whom correspondence should be addressed.
Catalysts 2021, 11(8), 973; https://doi.org/10.3390/catal11080973
Submission received: 20 July 2021
/
Revised: 11 August 2021
/
Accepted: 12 August 2021
/
Published: 14 August 2021
(This article belongs to the Special Issue Promising Industrial Enzymes)
Abstract
:Asymmetric synthesis of chiral amines from prochiral ketones using transaminases is an attractive biocatalytic strategy. Nevertheless, it is hampered by its unfavorable thermodynamic equilibrium. In the present work, an insitu by-product removal strategy was applied for the synthesis of 3-amino-1-phenylbutane (3-APB) by coupling a transaminase with a pyruvate decarboxylase (PDC), which does not require the use of any expensive additional cofactor. Using this strategy, the pyruvate obtained in the transamination reaction is transformed by PDC into acetaldehyde and CO2 which are of high volatility. Two different transaminases from Chromobacterium violaceum (CviTA) and Vibrio fluvialis (VflTA) were characterized to find out the appropriate pH conditions. In both cases, the addition of PDC dramatically enhanced 3-APB synthesis. Afterwards, different reaction conditions were tested to improve reaction conversion and yield. It was concluded that 30 °C and a 20-fold alanine excess lead to the best process metrics. Under the mentioned conditions, yields higher than 60% were reached with nearly 90% selectivity using both CviTA and VflTA. Moreover, high stereoselectivity for (S)-3-APB was obtained and ee of around 90% was achieved in both cases. For the first time, the asymmetric synthesis of 3-APB using PDC as by-product removal system using CviTA is reported.
1. Introduction
Chiral amines are target molecules of high industrial interest. These molecules are useful as intermediates and building blocks in pharmaceutical synthesis (e.g., for the production of codeine, morphine, and tropane alkaloids), as ligands in various asymmetric transformations, as chiral ligands in metal-complex catalysis, as chiral auxiliaries and resolving agents, and in agrochemical industries [1,2,3,4,5].
Chemical synthesis of chiral amines is usually performed by hydrogenation of a Schiff base. Other chemical processes could be also applied, such as nucleophilic addition, diastereoisomeric crystallization, and C-H insertion [6,7,8,9,10,11]. Alternatively, the biotechnological approaches for the synthesis of chiral amines have been described as a greener alternative to their chemical counterparts. In the literature, several enzymatic processes have been extensively reported using hydrolases, lyases, oxidoreductases, and transaminases (TAs) [7,8,12,13,14,15,16].
Transaminases (TAs) are pyridoxal-5′-phosphate (PLP) dependent enzymes that transfer an amino group from an amino donor onto a carbonyl group of an amino acceptor. These enzymes can be classified into α-TAs and ω-TAs, depending on the type of substrate they convert. α-TAs require a carboxylic group in the α-position to the keto and amine groups. Thus, α-TAs can only act on α-amino acids and α-keto acids. On the contrary, ω-TAs can transfer a −NH2 group from an amino donor to an amino acceptor in which at least one of the two molecules is not an α-amino acid or an α-keto acid [13,16]. Thus, ω-TAs are very useful in chiral amine synthesis as they allow the amination not only of keto acids, but also of ketones or aldehydes [13].
The most well-known and employed TAs are biologically active as dimers or higher oligomers, and they require the coordination of PLP in the active site to perform transamination [17,18]. The reaction is divided in two half reactions: oxidative deamination of the amino donor and reductive amination of the amine acceptor [19,20]. In both reactions, PLP acts as a molecular shuttle transporting the amino group, following a ping-pong bi-bi mechanism [13,17,21,22,23,24]. PLP, which forms a Schiff base with an active site lysine, is firstly aminated resulting in the formation of pyridoxamine-5′-phosphate (PMP), while the respective keto product of the amino donor is released [20,22,23]. Finally, the amino group from PMP is transferred to the acceptor molecule producing the aminated product and regenerating PLP for the next catalytic cycle [22,23].
The use of these enzymes for asymmetric synthesis of chiral amines from prochiral ketones is a very attractive strategy in biocatalysis. Nevertheless, although the reaction could theoretically reach 100% of conversion, the thermodynamic equilibrium is unfavorable. Two main strategies could be applied to overcome this issue: (i) the addition of an excess of the amino donor, and (ii) the in situ removal of the reaction by-product. Regarding the second approach, several amino donors can be used to obtain volatile ketones, (e.g., isopropyl amine to acetone) which can be easily removed from the media [25]. Another interesting strategy for in situ by-product removal is by coupling a secondary enzymatic reaction. If alanine is used as amino-donor, pyruvate is formed as by-product which can be easily transformed by a lactate dehydrogenase or amino acid dehydrogenase [8,13,16]. However, these oxidoreductases require expensive cofactors, demanding a system for its regeneration. The use of pyruvate decarboxylase (PDC) raises up as an alternative to oxidoreductases as it does not require NAD(H) [26,27]. PDC reaction generates acetaldehyde and CO2, which are of high volatility, leading to a promising system for in situ pyruvate removal. PDC only requires Mg2+ and TPP as cofactors which are regenerated in the catalytic mechanism.
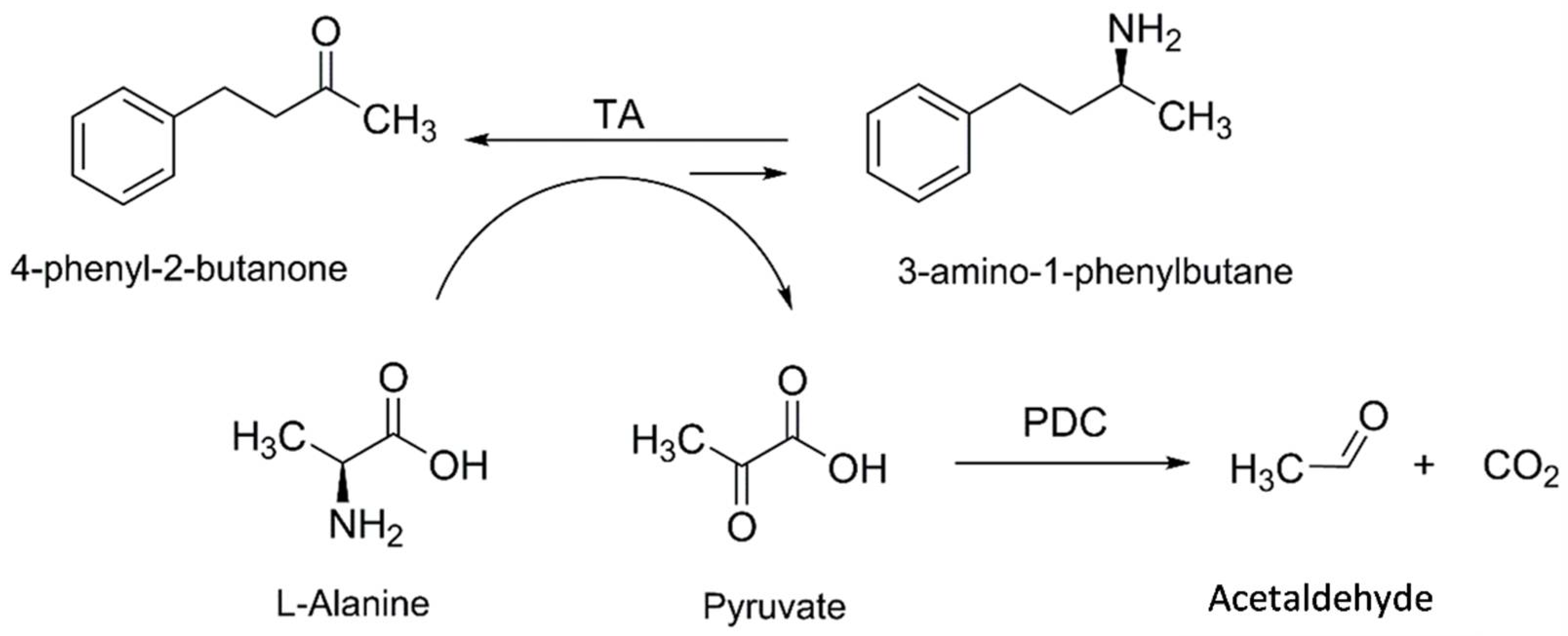
In the present work, the synthesis of 3-amino-1-phenylbutane (3-APB), has been studied using a one-pot multi-enzymatic system made up of an ω-TA and a PDC from Z. palmae (see Figure 1). Two different TAs from C. violaceum (CviTA) and V. fluvialis (VflTA) have been applied for the development of the cascade process. Finally, different reaction conditions were studied to maximize the process metrics.
2. Results and Discussion
2.1. Enzyme Characterization
Aiming to find suitable conditions for the one-pot multi-enzymatic synthesis of 3-APB, enzyme characterization was performed to find the operational conditions where transaminase and pyruvate decarboxylase could be compatible. Thus, a study of the effect of pH on both enzymatic activity and kinetic stability was carried out.
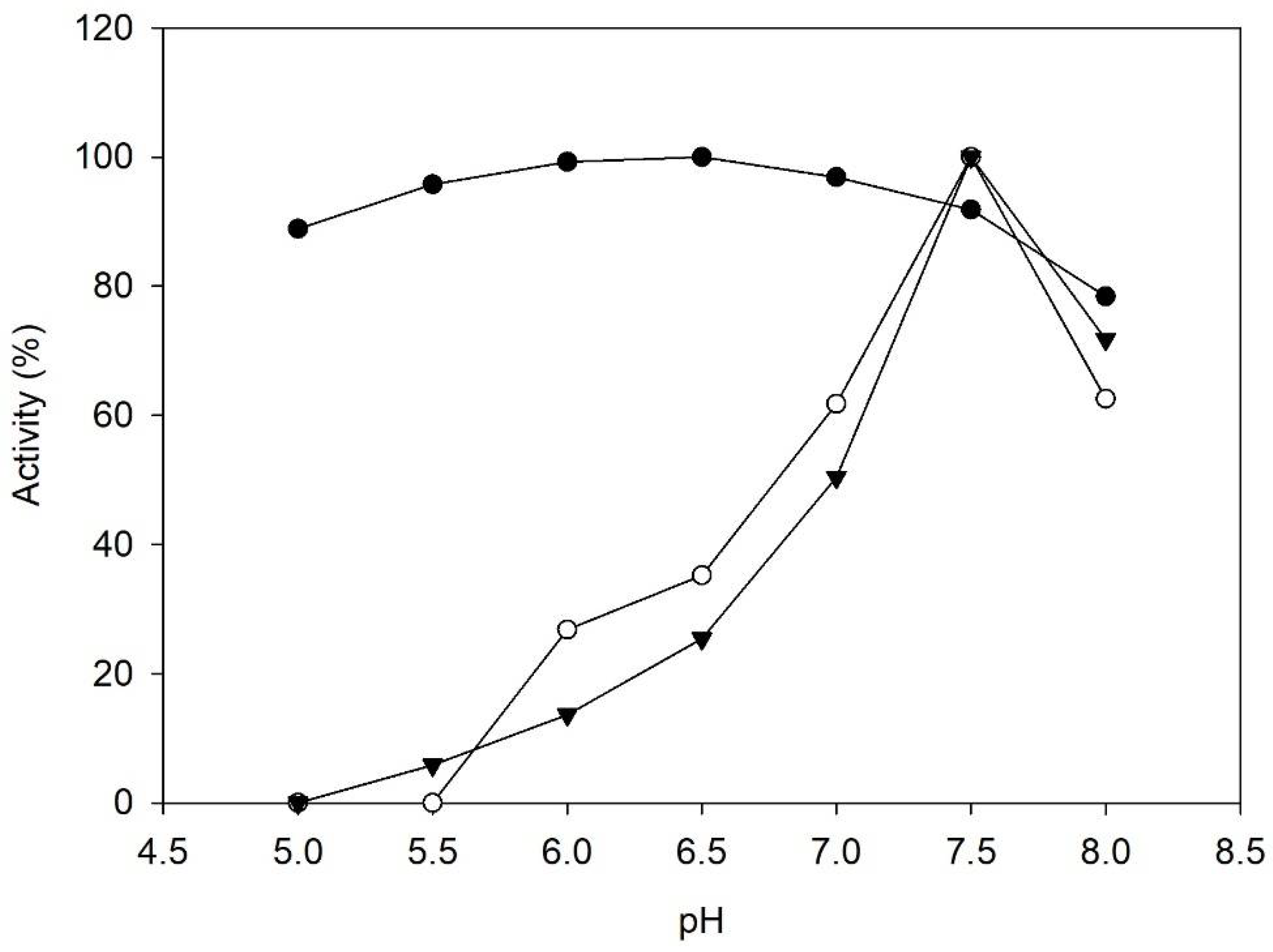
In Figure 2, pH activity profiles of PDC and TAs are shown. Regarding transaminases, CviTA and VflTA showed narrower profiles, both reaching the maximum activity under pH 7.5. Although it is usual to find optimum pHs higher than 8 for transaminases in the literature, especially for CviTA and VflTA [28,29], the optimum conditions found in this study have also been reported by other authors [30]. It should also be mentioned that VflTA precipitated under pH 5, while CviTA precipitation occurred under pH 5.5.
PDC reached its maximum activity under pH 6.5. Nevertheless, the PDC profile towards pH was not as dependent as transaminases, maintaining more than 80% of the maximum activity from pH 5 to 7.5. Previous characterizations of Z. palmae PDC revealed similar results, although slightly lower optimum pH values were reported [31].
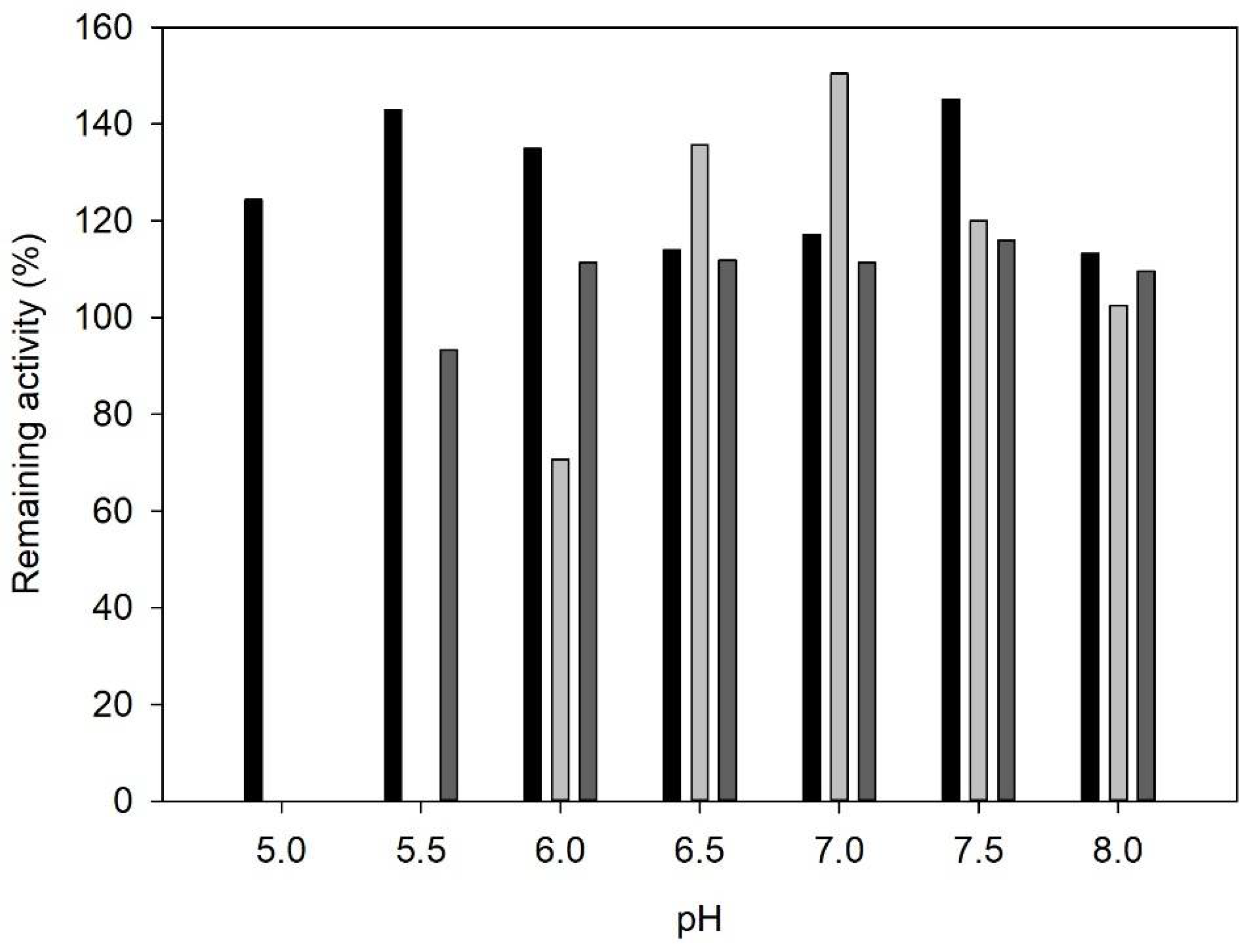
Stability towards pH was also checked under different pH (5–8) (Figure 3). Except for pH 5 and 5.5, all enzymes showed high stability. A hyper-activation was observed for PDC in all tested conditions. Activity values higher than 100% under pH 6 to 8 were also detected for VflTA, while CviTA was only hyperactivated under pHs from 6.5 to 8. Several authors also reported this effect on transaminases [32,33] which has been related to the enzyme 3D-structure. Most transaminases are homodimers whose active site is formed by both enzyme subunits. Thus, the hyper-activation phenomena could be caused by a complete dimerization to form active transaminase [32].
Taking into account that (i) PDC and all TAs showed high stability towards all tested pHs, and (ii) transaminases activity is strongly affected by pH compared to PDC, the conditions selected for the 3-APB synthesis were defined by the optimum pH of each transaminase, i.e., 7.5 for both CviTA and VflTA.
2.2. Transaminase Testing
A preliminary test was carried out, aiming to select which transaminase could be used to develop the enzymatic cascade. Reactions were performed following a double approach: (i) without adding pyruvate decarboxylase or (ii) by coupling the PDC as in situ by-product removal. Reactions were performed under the optimum pH for each transaminase (Table 1).
Results revealed that if no PDC is coupled, only slight amounts of 3-APB are detected. On the other hand, when the cascade system is applied, CviTA and VflTA led to significant amounts of 3-APB (Figure S1). These results suggested that the TA-PDC coupling favors an equilibrium shifting towards amine formation.
2.3. Synthesis of 3-APB Catalyzed by TA and PDC
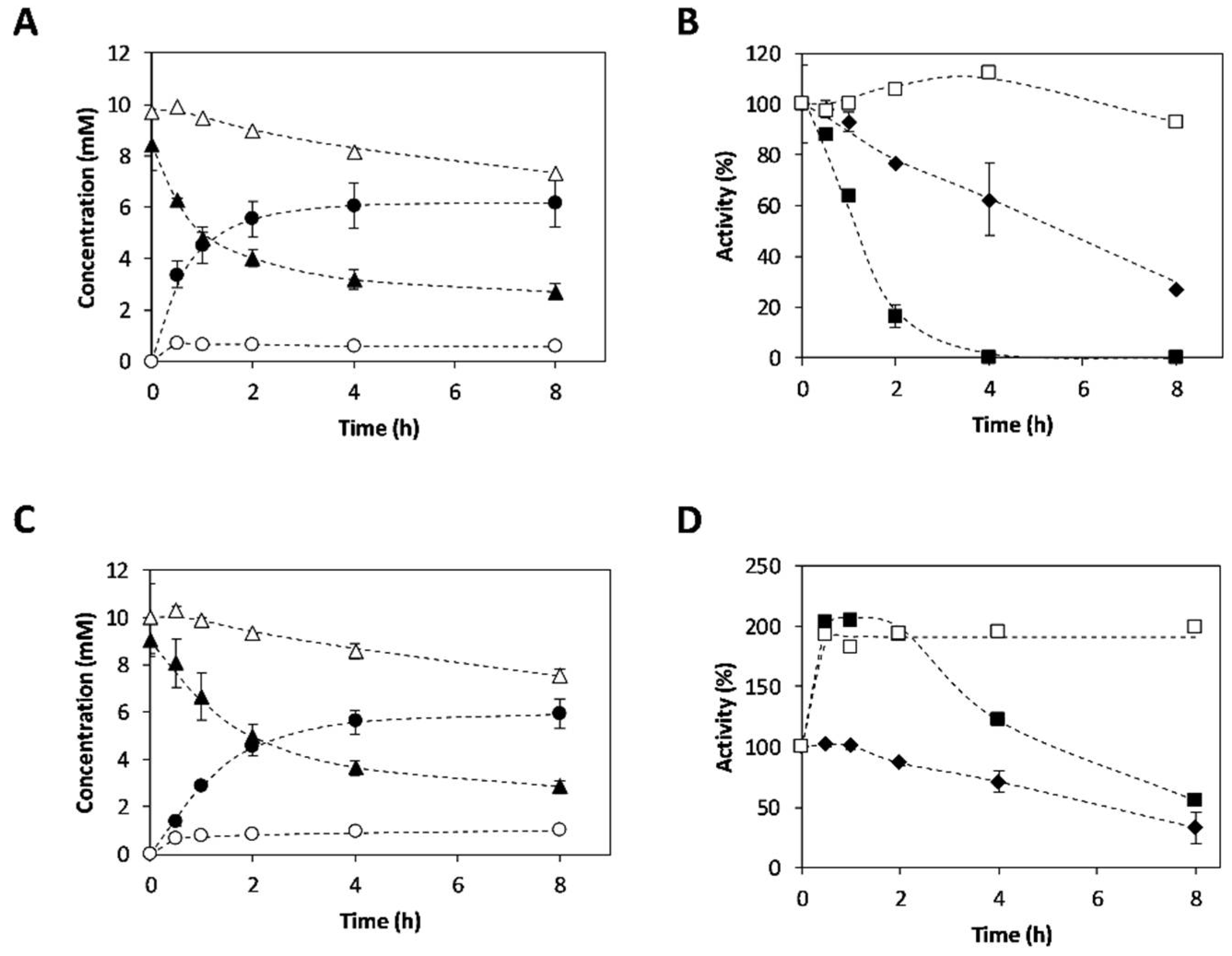
3-APB synthesis catalyzed by CviTA and VflTA was deeply studied. Reactions were performed using transaminases as the only biocatalysts, as well as coupled with PDC. These studies allow us to compare both approaches, single enzymatic system and multi-enzymatic system, for in situ product removal. Reaction time courses are presented in Figure 4A,C. Results showed that an effective amine synthesis only took place by the cascade systems (TA-PDC). In contrast, reactions performed without pyruvate removal led to 10-fold and 6-fold lower 3-APB concentration with CviTA-PDC and VflTA-PDC, respectively. Even though the same final amine concentration was achieved with both transaminases (≈6 mM), CviTA showed a 1.8-fold higher initial reaction rate (4.95 mM·h−1) than VflTA (2.83 mM·h−1). Regarding stability profiles (Figure 4B,D), while CviTA activity sharply decreased, suffering a 90% deactivation after two hours, VflTA showed a better operational stability. VflTA underwent a hyperactivation under reaction conditions, reaching up to a 2-fold increase in enzyme activity after 30 min from the reaction starting, maintaining 60% of the initial activity after 8 h.
It can be also observed that, when no PDC was coupled, and thus almost no amine was formed, the TAs showed higher stability. A low operational stability in TAs when transamination takes place has been widely reported by other authors [33] and recently related to an irreversible inactivation step that proceeds once the TA:PMP complex dissociates into free PMP and apoenzyme [17].
The same conversion (Table 2) was achieved with both transaminases when coupled with PDC, which were around 3-fold higher compared to the single enzyme approach. Moreover, the obtained yields in both coupled reactions were similar to the achieved conversion, which reflects a high reaction selectivity towards the target product (88.7 ± 10.2% and 86.1 ± 12.9% for the CviTA-PDC system and the VflTA-PDC system, respectively). When no PDC was added, similar conversions with both TAs were also reached (23.0 ± 9.5% and 22.9 ± 8.8% for the CviTA system and the VflTA system, respectively). However, yields were significantly lower compared to the conversions, resulting in very low selectivity: <35% and <60% (Table 2) for CviTA and VflTA, respectively. These results were an indicative of an undesired side-reaction emergence when decarboxylation reaction is not coupled. The unidentified side-reactions may be explained by the use of CFEs instead of the pure enzymes, which increases the system complexity.
Chiral amine synthesis using VflTA and Z. palmae PDC system was reported by Höhne et al. [26], who synthesized several amines including 3-APB. However, a conversion of 45% was obtained in that case [26]. Therefore, in the present work, process metrics of the TA-PDC system have been improved. Nevertheless, better conversions were achieved in studies where VflTA was coupled with other co-product removal systems, such as alanine dehydrogenase (AlaDH) or lactate dehydrogenase (LDH) [34]. The use of CviTA coupled with PDC has not been reported, thus the effectiveness of the system with this enzyme has been proven in the present study. However, CviTA has been applied in the mentioned AlaDH and LDH system, achieving conversions of 16% and 86%, respectively [35].
2.4. Effect of Temperature on 3-APB Synthesis
Temperature effect on reaction performance was studied to optimize the multi-enzymatic system. Increasing temperature to 40 °C was tested following a double objective: (i) to increase reaction rate and (ii) to enhanced volatile by-product elimination from the reaction medium (acetaldehyde and CO2). Decreasing the temperature to 15 °C was also tested, aiming to improve enzyme stability and, thus, prevent an early reaction stop due to enzyme deactivation.
Synthesis profiles and operational stabilities are shown in Figure 5. When temperature was increased to 40 °C, a 1.45-fold higher initial reaction rate was obtained for both TAs-PDC systems compared to cascade reactions performed at 30 °C (7.2 mM·h−1 CviTA and 5.8 mM·h−1 VflTA). However, lower 3-APB concentrations were achieved (around 4.5 mM in both cases). In addition, a decrease in product concentration was detected after 4 h of reaction, which was slightly more pronounced with VflTA. Thus, temperature increase may have led to a boost in undesired side reactions involving the target product. Temperature increase also negatively affected the enzyme operational stabilities (Figure 5B,D), especially in Vfl-TA which, after 8 h under 40 °C, was practically deactivated, while at 30 °C it still conserved around 60% of the initial activity. Other authors have reported similar results in the synthesis of chiral amine from pro-chiral ketones using commercial transaminases. When ATA 103, ATA-113, ATA-114, and ATA 117 were applied for asymmetric amination of 4-phenyl 2-butanone, lower conversions were obtained with ATA 114, ATA-117, and ATA-103 after 24 h when temperature was increased from 30 to 40 °C, probably due to a decrease in enzyme stability. Only with commercial ATA-113 conversion increased from 61 to 82% after temperature increasing, which could be related to a higher operational stability of this transaminase towards temperature [36].
As expected, both CviTA and VflTA increased their operational stability when the temperature was decreased to 15 °C. Nevertheless, this stability enhanced was associated to lower reaction rates (2.1 mM·h−1 CviTA and 0.28 mM·h−1 Vfl-TA), which increased reaction times to 24 h and 48 h, respectively.
As it is shown in Table 3, temperature shift was only favorable for the CviTA at 15 °C in terms of selectivity, the high value of which indicates that undesired side reactions may have been suppressed. However, only a 10% increase in reaction yield was obtained compared to the results at 30 °C, while space time yield suffered a 60% decrease. Regarding VflTA, STY loss at 15 °C was even more severe due to the high reaction time combined with a decrease in terms of conversion, yield, and selectivity. Regarding the results concerning 40 °C, low selectivity percentages, which did not surpass 50%, in addition to the significant difference between conversions and yields, indicates again the presence of undesired side reactions. Altogether, this lead to the conclusion that 30 °C was the best option to perform the cascade reactions.
2.5. Effect of Amine Donor Concentration on 3-APB Synthesis
Amine donor concentration on reaction yield was also studied. Although certain limitations, such as enzyme inhibition, substrate solubility, and downstream complexity must be taken into account [19,27,37], offering an excess of amino donor is a widely reported strategy to force an equilibrium shift in transaminations [6,15,19,27,37]. When L-Alanine is used as amine donor, at least 10-fold excess is recommendable [15]. A study on alanine excess effect on reaction yield has been reported for the case of 1-N-Boc-3-aminopyrrolidine synthesis by the TA-PDC cascade system [27]. In the mentioned work, a significant yield increase was obtained when an alanine excess higher than 22-fold was used. For this reason, in this study, initial lower amino donor concentrations than 200 mM (20-fold excess) were not tested and reactions were performed with a 40-fold and a 60-fold excess.
Figure 6 shows reaction time courses at different initial alanine concentrations (200 mM, 400 mM, and 600 mM) both using CviTA-PDC and VflTA-PDC cascade systems. In both cases, initial reaction rate is higher when alanine concentration increases from 200 to 600 mM thus, showing no inhibitory effect on transaminases. Nevertheless, no significant changes were observed in operational stabilities (Figure 6B,D). Final 3-APB concentrations achieved at 400 mM and 600 mM alanine concentration were not significantly higher than the ones obtained with 200 mM (see Table 4). Around 10% higher yield was obtained when amine donor excess was increased to 60-fold (600 mM alanine) in comparison with applying a 20-fold excess (200 mM alanine), which agrees with the previously reported 1-N-Boc-3-aminopyrrolidine synthesis [26]. The similarity between yield and conversion percentages, as well as the high selectivity values, indicates that the alanine increase did not favor side reactions. Although higher alanine concentrations did not show a negative effect on reaction, the low yield increase obtained lead to the conclusion that amine donor excess of 20-fold is the most suitable strategy.
It should be considered that in the proposed system acetaldehyde is the by-product of the reaction catalyzed by PDC. This compound could be used by transaminase as substrate, thus reducing the efficiency of the system due to a substrate competition between 4-phenyl 2-butanone and acetaldehyde.
2.6. Enantiomeric Excess Determination
Enantiomeric excess of 3-APB obtained in the most suitable conditions (30 °C and 20-fold excess of amine donor) was determined by NMR using BHP as chiral solvating agent (Figure S2). Both CviTA-PDC and VflTA-PDC systems showed high stereoselectivity reaching enantiomeric excesses of 92.0% and 89.3% for (S)-3-APB, respectively. Similar ee was reported for the VflTA-PDC cascade by Höhne et al. [26], reaching an ee of 88.5%. In other reported cases in which VflTA was coupled with AlaDH and LDH as pyruvate removing systems, similar ee values were also obtained (84% (S) and 89% (S), respectively) [34]. Finally, as mentioned before, CviTA has never been reported in the asymmetric chiral amine system using PDC. In addition, the use of this transaminase coupled with AlaDH and LDH lead to low stereoselectivity results in the case of 3-APB [35]. Therefore, ee percentages higher than 90% have been obtained for the first time with CviTA.
3. Materials and Methods
3.1. Chemicals and Enzymes
All chemicals were purchased from Sigma Aldrich (St. Louis, MO, USA). Transaminases from C. violaceum (CviTA) and V. fluvialis (VflTA) were kindly donated by DSM/InnoSyn in the form of E. coli cell free extracts (CFE), containing between 40 and 55 mg·mL−1 of total protein. For CviTA, 47% of the total protein was transaminase, while this percentage was 35% for VflTA. Pyruvate decarboxylase (PDC) from Z. palmae was produced by high cell density cultures of a recombinant E. coli strain obtained by the research group [38]. CFEs of this strain were obtained by sonication with a Vibracell® VC50 (Sonic and Materials®, Newton, CT, USA) with 4 × 15 s pulses (50 W) and 2 min intervals in ice between each pulse. Centrifugation was performed (10,000× g, 10 min) to remove the resulting cell debris. ZpPDC CFEs contained 10 mg·mL−1 of total protein, from which around 40% was PDC.
3.2. Transaminase Activity Assay
Transaminase activity assay was based on the acetophenone formation, measured at 340 nm, from α-Methylbenzylamine and pyruvic acid. The assay was performed at 30 °C in 1 mL reaction mixture, consisting of potassium phosphate buffer 100 mM and pH 7.5 containing 0.1 mM PLP, 22.5 mM α-Methylbenzylamine and 5 mM sodium pyruvate. Molar extinction coefficient (ε) of acetophenone is 0.28 mM−1·cm−1 and one transaminase activity unit corresponds to the amount of enzyme that produces 1 μmol acetophenone per minute at 30 °C. Absorbance measurements were performed using a SPECORD® 200 PLUS (Analytik Jena, Jena, Germany) spectrophotometer.
3.3. Pyruvate Decarboxylase Activity Assay
PDC activity was determined by coupling the pyruvate decarboxylation with alcohol dehydrogenase (ADH) and following NADH oxidation at 340 nm and 25 °C, whose molar extinction coefficient (ε) is 6.22 mM−1·cm−1. The reaction mixture contained 33 mM sodium pyruvate, 0.11 mM NADH, 3.5 UA mL−1 ADH from S. cerevisiae (Sigma Aldrich, St. Louis, MO, USA), 0.1 mM TPP, and 0.1 mM MgCl2 in citrate buffer 200 mM and pH 6. One unit of PDC activity corresponds to the amount of pyruvate decarboxylase that converts 1 μmol of pyruvate to acetaldehyde per minute. Absorbance measurements were performed using a SPECORD® 200 PLUS (Analytik Jena, Jena, Germany) spectrophotometer.
3.4. Influence of pH on Enzyme Activity and Stability
The optimum pH for each enzyme was found by performing enzymatic activity assays, replacing the assay buffer with citrate buffer 100 mM (pH from 5 to 6), and potassium phosphate buffer 100 mM (pH 6.5–8). Enzyme stability was determined by preparing a 5% v·v−1 solution of the cell lysates containing the enzymes in the mentioned buffers, and maintaining the solutions at 25 °C and mild agitation. Aliquots were taken at different times and enzyme activity was measured.
3.5. Transaminase Screening
Reactions were performed in 15 mL Falcon tubes with 3 mL reaction mixture consisting of 5% v·v−1 transaminase cell lysate, 25% v·v−1 PDC cell lysate, 1 mM pyridoxal-5′-phospate (PLP), 0.1 mM thiamine pyrophosphate (TPP), 0.1 mM MgCl2, 200 mM alanine, and 10 mM 4-phenyl-2-butanone (4-PB) in 150 mM of potassium phosphate buffer, pH 7.5. Samples were placed in a Multitherm shaker (Benchmark Scientific, Sayreville, NJ, USA) at 30 °C and 1000 rpm, and after 24 h the presence or absence of 3-APB was detected by HPLC. Control reactions were performed substituting PDC cell lysate with distillate water.
3.6. Asymmetric Synthesis of APB with Transaminase and Pyruvate Decarboxylase
All biocatalytic reactions were performed in potassium phosphate buffer 150 mM and pH 7.5 containing 1 mM pyridoxal-5′-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2. A transaminase cell lysate concentration of 5% v·v−1 in volume was coupled with 25% v·v−1 PDC cell lysate in 15 mL Falcon tubes containing 3 mL working volume and kept at 30 °C and 1000 rpm for 8 h in a Multitherm shaker (Benchmark Scientific, Sayreville, NJ, USA). Reactant concentrations were 10 mM of 4-phenyl 2-butanone, which corresponds to the solubility limit, and 200 mM L-Alanine.
3.7. Temperature and Amine Donor Concentration Effect on 3-APB Synthesis
To study the temperature effect on 3-APB synthesis, all reaction conditions mentioned were maintained, while temperature was shifted to 15 °C or 40 °C. In the alanine concentration effect studies, reactions were performed at 30 °C, increasing the amine donor concentration to 400 mM and 600 mM.
3.8. Quantification of 3-APB and 4-PB
3-amino 1-phenylbutane and its pro chiral ketone (4-phenyl 2-butanone) concentrations were measured by HPLC analysis in an UltiMate 3000 (Dionex) equipped with a variable wavelength detector. Compounds were separated on a reversed-phase CORTECS C18 + 2.7 μm 4.6 × 150 mm column (Waters Milford, MA, USA). After acidifying reaction samples with 3 M hydrochloric acid to deactivate the enzymes, 15 μL were injected at a 0.7 mL min−1 flow rate and 30 °C. Samples were eluted using a gradient from 30 to 55% solvent B—consisting of 0.095% (v·v−1) in MeCN/H2O 4:1 (v·v−1)—to solvent A—consisting of 0.1% (v·v−1) TFA in H2O—over 13 min. Compounds were detected at a 254 nm wavelength.
3.9. Enantiomeric Excess Determination
At the end of the reaction, the entire volume was acidified with 3 M hydrochloric acid and centrifuged to remove the precipitate proteins. After that, a first extraction step was performed with dichloromethane and the organic phase was discarded. After basifying the aqueous phase with NaOH, a second extraction was carried out with the same solvent. The new organic phase was dried with anhydrous sodium sulphate and concentrated by distillation in a Heidolph VV 2000 rotary evaporator. APB was separated from the remaining impurities by a silica column using 1:10 methanol in dichloromethane with 2% triethylamine as mobile phase.
The enantiomeric excess was determined by nuclear magnetic resonance (NMR) using (R)-(−)-1,1′-binaphtyl-2,2′-hydrogenphosphate (BHP) as chiral solvating agent (CSA). An equimolar quantity of the CSA was used if not stated otherwise. The 1H-NMR spectra were acquired with a Bruker AVANCE-III 600 MHz NMR Spectrometer (Bruker Biospin, Rheinstetten, Germany) in CDCl3 at 298 K.
4. Conclusions
Coupling pyruvate removal system has been proved to be required for the asymmetric synthesis of 3-amino-1-phenylbutane. The target reaction was efficiently carried out by the cascade reaction of C. violaceum and V. fluvialis transaminases and PDC. PDC coupling dramatically enhanced 3-APB synthesis in comparison with a single enzyme approach. After reaction optimization, it was concluded that 30 °C and a 20-fold excess of amine donor were the best conditions obtaining yields higher than 60% with nearly 90% selectivity. Moreover, reactions took place with high stereoselectivity for (S)-APB and with up to 90% ee in both cases. For the first time, asymmetric synthesis of 3-APB using PDC as by-product removal system is reported using CviTA.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/catal11080973/s1, Figure S1: HPLC chromatogram of (A) 1 mM commercial 3-APB from Sigma Aldrich (St. Louis, MO, USA), (B) final sample (24 h) of 3-APB synthesis catalyzed by CviTA and PDC and (C) final sample (24 h) of 3-APB synthesis catalyzed by CviTA and PDC, Table S1: 4-PB and 3-APB concentrations used for Figure 4A plotting, Table S2: 4-PB and 3-APB concentrations used for Figure 4A plotting, Table S3: PB and 3-APB concentrations used for Figure 4C plotting, Table S4: PB and 3-APB concentrations used for Figure 4C plotting, Table S5: 3-APB concentration used for Figure 5A plotting, Table S6: 3-APB concentration used for Figure 5A plotting, Table S7: 3-APB concentration used for Figure 5A plotting, Table S8: 3-APB concentration used for Figure 5C plotting, Table S9: 3-APB concentration used for Figure 5C plotting, Table S10: 3-APB concentration used for Figure 5C plotting, Table S11: 3-APB concentration used for Figure 6A plotting, Table S12: 3-APB concentration used for Figure 6A plotting, Table S13: 3-APB concentration used for Figure 6A plotting, Table S14: 3-APB concentration used for Figure 6C plotting, Table S15: 3-APB concentration used for Figure 6C plotting, Table S16: 3-APB concentration used for Figure 6C plotting, Figure S2: Results on nuclear magnetic resonance (NMR) for 3-APB obtained from reactions catalyzed by (A) CviTA-PDC and (B) VflTA-PDC before adding chiral solvating agent (red line) and after adding chiral solvating agent (green line).
Author Contributions
Conceptualization, M.G. and G.Á.; methodology, N.A.; validation, M.G., G.Á. and N.A.; formal analysis, N.A.; investigation, N.A.; data curation, N.A.; writing—original draft preparation, N.A.; writing—review and editing, G.Á. and MG; supervision, M.G. and G.Á.; project administration, G.Á.; and funding acquisition, G.Á. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Spanish MINECO, project grant number CTQ2014-53114R and Ph.D. scholarship (FPI: BES 2015 072542) of N.A. It has been also funded by AGAUR-Generalitat de Catalunya (research group 2017 SGR 1462) and by European Regional Development Fund.
Data Availability Statement
All data is included in this article.
Acknowledgments
Z. palmae PDC gene was kindly donated by the Microbiology and Cell Science Department of the University of Florida (Gainesville, FL, USA). Transaminases were kindly donated by DSM/InnoSyn.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jacques, J.; Collet, A.; Wilen, S.H. Enantiomers, Racemates and Resolutions; John Wiley & Sons, Ltd.: New York, NY, USA, 1981; ISBN 0471080586. [Google Scholar]
- Whitesell, J.K. C2 symmetry and asymmetric induction. Chem. Rev. 1989, 89, 1581–1590. [Google Scholar] [CrossRef]
- Castagnetto, J.; Canary, J.W. A chiroptically enhanced fluorescent chemosensor. Chem. Commun. 1998, 203–204. [Google Scholar] [CrossRef]
- Arrasate, S.; Lete, E.; Sotomayor, N. Synthesis of enantiomerically enriched amines by chiral ligand mediated addition of organolithium reagents to imines. Tetrahedron Asymmetry 2001, 12, 2077–2082. [Google Scholar] [CrossRef]
- Nugent, T.; El-Shazly, M. Chiral Amine Synthesis—Recent Developments and Trends for Enamide Reduction, Reductive Amination, and Imine Reduction. Adv. Synth. Catal. 2010, 352, 753–819. [Google Scholar] [CrossRef]
- Breuer, M.; Ditrich, K.; Habicher, T.; Hauer, B.; Keßeler, M.; Stürmer, R.; Zelinski, T. Industrial Methods for the Production of Optically Active Intermediates. Angew. Chem. Int. Ed. 2004, 43, 788–824. [Google Scholar] [CrossRef] [PubMed]
- Ghislieri, D.; Turner, N.J. Biocatalytic Approaches to the Synthesis of Enantiomerically Pure Chiral Amines. Top. Catal. 2013, 57, 284–300. [Google Scholar] [CrossRef]
- Kelly, S.A.; Pohle, S.; Wharry, S.; Mix, S.; Allen, C.; Moody, T.S.; Gilmore, B.F. Application of ω-Transaminases in the Pharmaceutical Industry. Chem. Rev. 2017, 118, 349–367. [Google Scholar] [CrossRef]
- Hayashi, H.; Uchida, T. Nitrene Transfer Reactions for Asymmetric C-H Amination: Recent Development. Eur. J. Org. Chem. 2019, 2020, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.-S.; Gong, L.-Z. Palladium-Catalyzed Asymmetric Allylic C–H Functionalization: Mechanism, Stereo- and Regioselectivities, and Synthetic Applications. Acc. Chem. Res. 2020, 53, 2841–2854. [Google Scholar] [CrossRef]
- Abdine, R.A.A.; Hedouin, G.; Colobert, F.; Wencel-Delord, J. Metal-Catalyzed Asymmetric Hydrogenation of C═N Bonds. ACS Catal. 2020, 11, 215–247. [Google Scholar] [CrossRef]
- Busto, E.; Gotor-Fernández, V.; Gotor, V. Hydrolases in the Stereoselective Synthesis of N-Heterocyclic Amines and Amino Acid Derivatives. Chem. Rev. 2011, 111, 3998–4035. [Google Scholar] [CrossRef]
- Mathew, S.; Yun, H. ω-Transaminases for the Production of Optically Pure Amines and Unnatural Amino Acids. ACS Catal. 2012, 2, 993–1001. [Google Scholar] [CrossRef]
- Kroutil, W.; Fischereder, E.-M.; Fuchs, C.S.; Lechner, H.; Mutti, F.G.; Pressnitz, D.; Rajagopalan, A.; Sattler, J.H.; Simon, R.C.; Siirola, E. Asymmetric preparation of prim-, sec-, and tert-amines employing selected biocatalysts. Org. Process. Res. Dev. 2013, 17, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Green, A.P.; Turner, N.J.; O’Reilly, E. Chiral Amine Synthesis Using ω-Transaminases: An Amine Donor that Displaces Equilibria and Enables High-Throughput Screening. Angew. Chem. Int. Ed. 2014, 53, 10714–10717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomm, A.; O’Reilly, E. Transaminases for chiral amine synthesis. Curr. Opin. Chem. Biol. 2018, 43, 106–112. [Google Scholar] [CrossRef]
- Börner, T.; Rämisch, S.; Reddem, E.R.; Bartsch, S.; Vogel, A.; Thunnissen, A.-M.; Adlercreutz, P.; Grey, C. Explaining Operational Instability of Amine Transaminases: Substrate-Induced Inactivation Mechanism and Influence of Quaternary Structure on Enzyme–Cofactor Intermediate Stability. ACS Catal. 2017, 7, 1259–1269. [Google Scholar] [CrossRef]
- Chen, S.; Brocal, J.C.C.; Berglund, P.; Humble, M.S. Characterization of the stability of Vibrio fluvialis JS17 amine transaminase. J. Biotechnol. 2018, 282, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Berglund, P. Transaminase biocatalysis: Optimization and application. Green Chem. 2016, 19, 333–360. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.S.; Park, E.-S.; Shin, J.-S. Features and technical applications of ω-transaminases. Appl. Microbiol. Biotechnol. 2012, 94, 1163–1171. [Google Scholar] [CrossRef]
- Slabu, I.; Galman, J.; Lloyd, R.C.; Turner, N.J. Discovery, Engineering, and Synthetic Application of Transaminase Biocatalysts. ACS Catal. 2017, 7, 8263–8284. [Google Scholar] [CrossRef]
- Patil, M.D.; Grogan, G.; Bommarius, A.; Yun, H. Recent Advances in ω-Transaminase-Mediated Biocatalysis for the Enantioselective Synthesis of Chiral Amines. Catalysts 2018, 8, 254. [Google Scholar] [CrossRef] [Green Version]
- Koszelewski, D.; Tauber, K.; Faber, K.; Kroutil, W. ω-Transaminases for the synthesis of non-racemic α-chiral primary amines. Trends Biotechnol. 2010, 28, 324–332. [Google Scholar] [CrossRef]
- Kaulmann, U.; Smithies, K.; Smith, M.E.; Hailes, H.; Ward, J. Substrate spectrum of ω-transaminase from Chromobacterium violaceum DSM30191 and its potential for biocatalysis. Enzym. Microb. Technol. 2007, 41, 628–637. [Google Scholar] [CrossRef]
- Petri, A.; Colonna, V.; Piccolo, O. Asymmetric synthesis of a high added value chiral amine using immobilized ω-transaminases. Beilstein J. Org. Chem. 2019, 15, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Höhne, M.; Kühl, S.; Robins, K.; Bornscheuer, U. Efficient Asymmetric Synthesis of Chiral Amines by Combining Transaminase and Pyruvate Decarboxylase. ChemBioChem 2008, 9, 363–365. [Google Scholar] [CrossRef]
- Tufvesson, P.; Lima-Ramos, J.; Jensen, J.S.; Al-Haque, N.; Neto, W.L.A.; Woodley, J. Process considerations for the asymmetric synthesis of chiral amines using transaminases. Biotechnol. Bioeng. 2011, 108, 1479–1493. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Kim, B. Asymmetric synthesis of chiral amines with ω-transaminase. Biotechnol. Bioeng. 1999, 65, 206. [Google Scholar] [CrossRef]
- Schell, U.; Wohlgemuth, R.; Ward, J.M. Synthesis of pyridoxamine 5′-phosphate using an MBA:pyruvate transaminase as biocatalyst. J. Mol. Catal. B Enzym. 2009, 59, 279–285. [Google Scholar] [CrossRef]
- Humble, M.S.; Cassimjee, K.E.; Håkansson, M.; Kimbung, Y.R.; Walse, B.; Abedi, V.; Federsel, H.-J.; Berglund, P.; Logan, D.T. Crystal structures of the Chromobacterium violaceum ω-transaminase reveal major structural rearrangements upon binding of coenzyme PLP. FEBS J. 2012, 279, 779–792. [Google Scholar] [CrossRef] [PubMed]
- Raj, K.C.; Talarico, L.A.; Ingram, L.O.; Maupin-Furlow, J.A. Cloning and Characterization of the Zymobacter palmae Pyruvate Decarboxylase Gene (pdc) and Comparison to Bacterial Homologues. Appl. Environ. Microbiol. 2002, 68, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassimjee, K.E.; Humble, M.S.; Miceli, V.; Colomina, C.G.; Berglund, P. Active Site Quantification of an ω-Transaminase by Performing a Half Transamination Reaction. ACS Catal. 2011, 1, 1051–1055. [Google Scholar] [CrossRef]
- Chen, S.; Land, H.; Berglund, P.; Humble, M.S. Stabilization of an amine transaminase for biocatalysis. J. Mol. Catal. B Enzym. 2016, 124, 20–28. [Google Scholar] [CrossRef]
- Mutti, F.G.; Fuchs, C.S.; Pressnitz, D.; Turrini, N.G.; Sattler, J.H.; Lerchner, A.; Skerra, A.; Kroutil, W. Amination of Ketones by Employing Two New (S)-Selective ω-Transaminases and the His-Tagged ω-TA from Vibrio fluvialis. Eur. J. Org. Chem. 2011, 2012, 1003–1007. [Google Scholar] [CrossRef]
- Koszelewski, D.; Göritzer, M.; Clay, D.; Seisser, B.; Kroutil, W. Synthesis of optically active amines employing recombinant ω-transaminases in E. coli cells. ChemCatChem 2010, 2, 73–77. [Google Scholar] [CrossRef]
- Koszelewski, D.; Lavandera, I.; Clay, D.; Rozzell, D.; Kroutil, W. Asymmetric Synthesis of Optically Pure Pharmacologically Relevant Amines Employing ω-Transaminases. Adv. Synth. Catal. 2008, 350, 2761–2766. [Google Scholar] [CrossRef]
- Gundersen, M.T.; Abu, R.; Schürmann, M.; Woodley, J.M. Amine donor and acceptor influence on the thermodynamics of ω-transaminase reactions. Tetrahedron Asymmetry 2015, 26, 567–570. [Google Scholar] [CrossRef]
- Alcover, N.; Carceller, A.; Alvaro, G.; Guillén, M. Zymobacter palmae pyruvate decarboxylase production process development: Cloning in Escherichia coli, fed-batch culture and purification. Eng. Life Sci. 2019, 19, 502–512. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Asymmetric synthesis of 3-amino 1-phenylbutane (3-APB) from 4-phenyl 2-butanone (4-PB) and L-Alanine catalyzed by the TA-PDC cascade process.
Figure 1.
Asymmetric synthesis of 3-amino 1-phenylbutane (3-APB) from 4-phenyl 2-butanone (4-PB) and L-Alanine catalyzed by the TA-PDC cascade process.

Figure 2.
pH activity profile of PDC (●), Cvi-TA (○), and Vfl-TA (▼). Assays were performed with citrate buffer 100 mM (pH 5–6), potassium phosphate buffer 100 mM (pH 6.5–8).
Figure 2.
pH activity profile of PDC (●), Cvi-TA (○), and Vfl-TA (▼). Assays were performed with citrate buffer 100 mM (pH 5–6), potassium phosphate buffer 100 mM (pH 6.5–8).

Figure 3.
pH stability profile of PDC (black bars), CviTA (light grey bars), and VflTA (dark grey bars). Assay conditions: citrate buffer 100 mM (pH 5–6), potassium phosphate buffer 100 mM (pH 6.5–8), 25 °C, mild agitation, 8 h.
Figure 3.
pH stability profile of PDC (black bars), CviTA (light grey bars), and VflTA (dark grey bars). Assay conditions: citrate buffer 100 mM (pH 5–6), potassium phosphate buffer 100 mM (pH 6.5–8), 25 °C, mild agitation, 8 h.

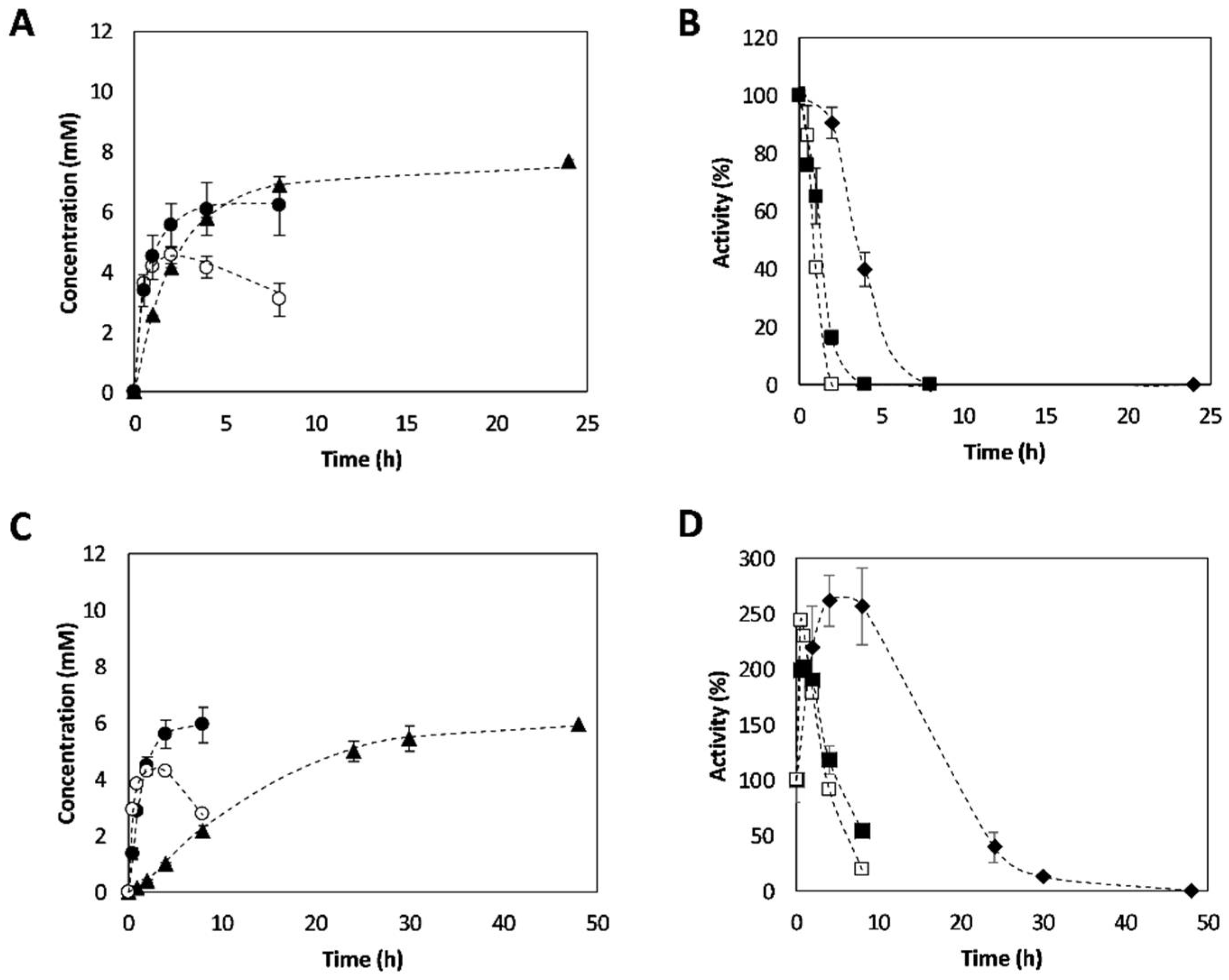
Figure 4.
Reaction time courses of 3-APB synthesis and operational enzymatic stability. (A) Reaction time courses performed with CviTA and (C) VflTA: (●) 3-APB concentration in cascade reactions; (◯) 3-APB concentration in reactions without PDC; (▲) 4-PB concentration in cascade reactions; and (△) 4-PB concentration in reactions without PDC. (B) Operational stability of CviTA and (D) VflTA, as well as PDC when cascade reactions were performed: (■) Transaminase activity in cascade reactions. (☐) Transaminase activity in reactions without PDC. (♦) PDC activity. Reaction conditions: 30 °C; pH 7.5; 4-PB 10 mM; L-Alanine 200 mM; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2. Concentration values are reported in Tables S1–S4.
Figure 4.
Reaction time courses of 3-APB synthesis and operational enzymatic stability. (A) Reaction time courses performed with CviTA and (C) VflTA: (●) 3-APB concentration in cascade reactions; (◯) 3-APB concentration in reactions without PDC; (▲) 4-PB concentration in cascade reactions; and (△) 4-PB concentration in reactions without PDC. (B) Operational stability of CviTA and (D) VflTA, as well as PDC when cascade reactions were performed: (■) Transaminase activity in cascade reactions. (☐) Transaminase activity in reactions without PDC. (♦) PDC activity. Reaction conditions: 30 °C; pH 7.5; 4-PB 10 mM; L-Alanine 200 mM; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2. Concentration values are reported in Tables S1–S4.

Figure 5.
Temperature effect on 3-APB synthesis and operational enzymatic stability. (A) Reaction time courses performed with CviTA-PDC and (C) VflTA-PDC at 15 °C (▲), 30°C (●), and 40 °C (◯). (B) Operational stability of Cvi-TA and (D) Vfl-TA at 15 °C (♦), 30 °C (■), and 40 °C (☐). Reaction conditions: pH 7.5; 4-PB 10 mM; L-Alanine 200 mM; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal-5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2. Concentration values are reported in Tables S5–S10.
Figure 5.
Temperature effect on 3-APB synthesis and operational enzymatic stability. (A) Reaction time courses performed with CviTA-PDC and (C) VflTA-PDC at 15 °C (▲), 30°C (●), and 40 °C (◯). (B) Operational stability of Cvi-TA and (D) Vfl-TA at 15 °C (♦), 30 °C (■), and 40 °C (☐). Reaction conditions: pH 7.5; 4-PB 10 mM; L-Alanine 200 mM; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal-5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2. Concentration values are reported in Tables S5–S10.

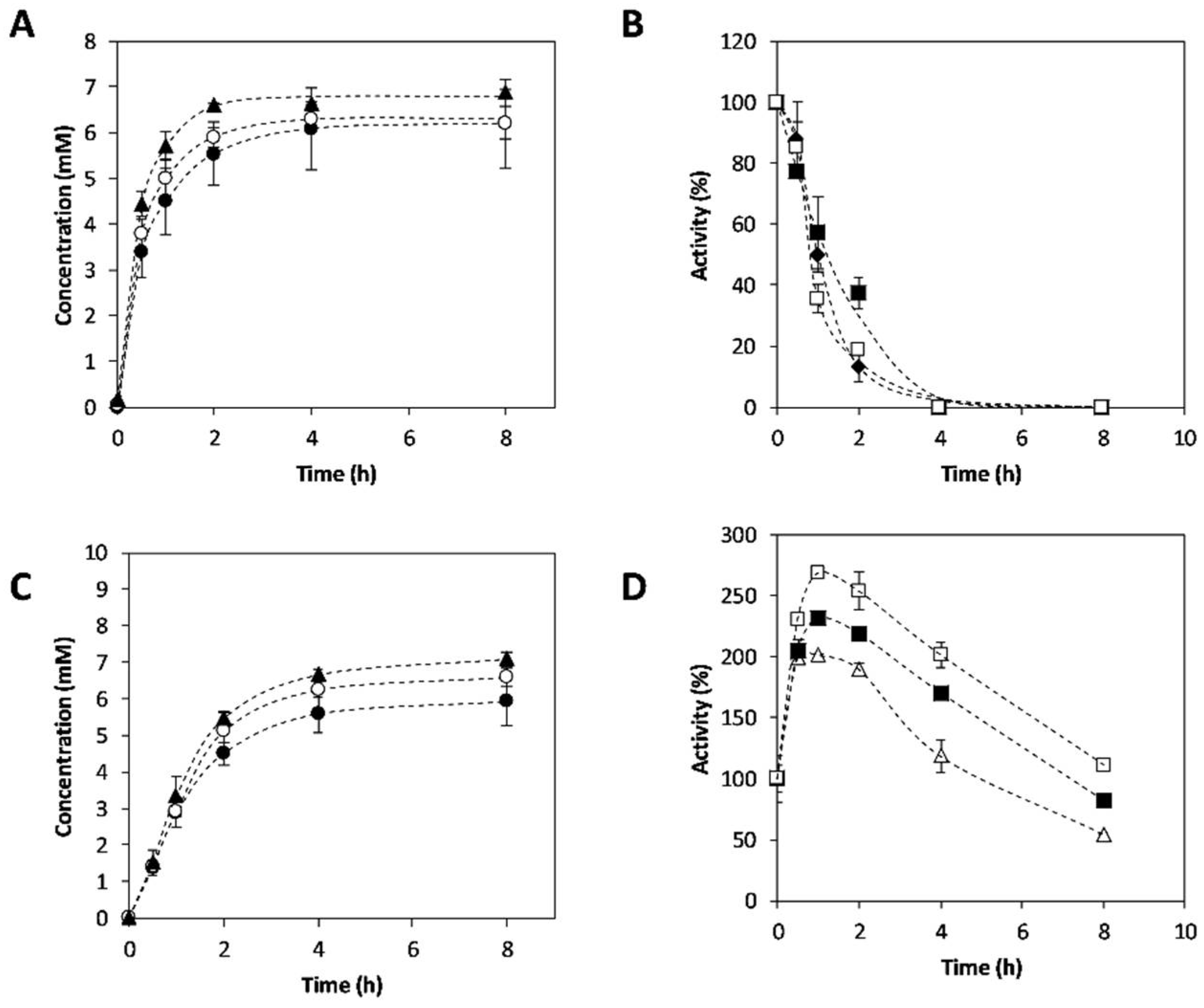
Figure 6.
L-Alanine concentration effect on 3-APB synthesis and on operational enzymatic stability. (A) Reaction time courses performed with CviTA-PDC or (C) VflTA-PDC using 200 mM (●), 400 mM (◯), and 600 mM (▲) initial L-alanine concentration. (B) Operational stability of CviTA and (D) VflTA (D) at 200 mM (♦), 400 mM (■), and 600 mM (☐) initial alanine concentration. Reaction conditions: 30 °C; pH 7.5; 4-PB 10 mM; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal-5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2. Concentration values are reported in Tables S11–S16.
Figure 6.
L-Alanine concentration effect on 3-APB synthesis and on operational enzymatic stability. (A) Reaction time courses performed with CviTA-PDC or (C) VflTA-PDC using 200 mM (●), 400 mM (◯), and 600 mM (▲) initial L-alanine concentration. (B) Operational stability of CviTA and (D) VflTA (D) at 200 mM (♦), 400 mM (■), and 600 mM (☐) initial alanine concentration. Reaction conditions: 30 °C; pH 7.5; 4-PB 10 mM; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal-5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2. Concentration values are reported in Tables S11–S16.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Transaminase screening for 3-APB synthesis. Reaction conditions: 24 h; 30 °C; 4-PB 10 mM; L-Alanine 200 mM, potassium phosphate buffer 150 mM, 25% v·v−1 PDC (30 U·mL−1), 5% v·v−1 transaminase, 1 mM pyridoxal-5′-phospate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2.
Table 1.
Transaminase screening for 3-APB synthesis. Reaction conditions: 24 h; 30 °C; 4-PB 10 mM; L-Alanine 200 mM, potassium phosphate buffer 150 mM, 25% v·v−1 PDC (30 U·mL−1), 5% v·v−1 transaminase, 1 mM pyridoxal-5′-phospate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2.
| Transaminase | PDC | pH | [APB] (mM) |
|---|---|---|---|
| CviTA | + | 7.5 | 6.2 |
| − | 0.6 | ||
| VflTA | + | 7.5 | 5.9 |
| − | 1.0 |
Table 2.
Process metrics of 3-APB synthesis. Reaction conditions: reaction time 8 h; temperature 30 °C; pH 7.5; 4-PB 10 mM; L-Alanine 200 mM; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal-5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2.
Table 2.
Process metrics of 3-APB synthesis. Reaction conditions: reaction time 8 h; temperature 30 °C; pH 7.5; 4-PB 10 mM; L-Alanine 200 mM; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal-5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2.
| Transaminase | PDC | Final [APB] (mM) | Conversion (%) | Yield (%) | Selectivity (%) | STY (mmol·L−1·h−1) |
|---|---|---|---|---|---|---|
| CviTA | Yes | 6.20 ± 0.98 | 72.0 ± 3.1 | 64.2 ± 10.1 | 88.7 ± 10.2 | 0.78 ± 0.12 |
| No | 0.58 ± 0.00 | 23.0 ± 9.5 | 6.1 ± 1.0 | 34.1 ± 18.3 | 0.07 ± 0.00 | |
| VflTA | Yes | 5.92 ± 0.64 | 71.0 ± 3.0 | 60.8 ± 6.6 | 86.1 ± 12.9 | 0.74 ± 0.08 |
| No | 1.01 ± 0.05 | 22.9 ± 8.8 | 10.5 ± 2.0 | 57.4 ± 30.8 | 0.13 ± 0.01 |
Table 3.
Process metrics of 3-APB synthesis applying the multi-enzymatic system TA-PDC at different temperatures. Reaction conditions: pH 7.5; 4-PB 10 mM; L-Alanine 200 mM; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal-5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2.
Table 3.
Process metrics of 3-APB synthesis applying the multi-enzymatic system TA-PDC at different temperatures. Reaction conditions: pH 7.5; 4-PB 10 mM; L-Alanine 200 mM; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal-5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2.
| Transaminase | Temperature (°C) | Reaction Time (h) | Final APB (mM) | Conversion (%) | Yield (%) | Selectivity (%) | STY (mmol·L−1·h−1) |
|---|---|---|---|---|---|---|---|
| CviTA | 15 | 24 | 7.68 ± 0.05 | 73.7 ± 0.5 | 75.4 ± 0.5 | 102.3 ± 1.4 | 0.32 ± 0.00 |
| 30 | 8 | 6.20 ± 0.98 | 72.0 ± 3.1 | 64.2 ± 10.1 | 88.7 ± 10.2 | 0.78 ± 0.12 | |
| 40 | 8 | 3.08 ± 0.54 | 61.7 ± 1.3 | 30.2 ± 5.3 | 48.8 ± 7.6 | 0.38 ± 0.07 | |
| VflTA | 15 | 48 | 5.92 ± 0.50 | 71.8 ± 1.3 | 52.2 ± 4.4 | 80.9 ± 5.4 | 0.12 ± 0.01 |
| 30 | 8 | 5.92 ± 0.64 | 71.0 ± 3.0 | 60.8 ± 6.6 | 86.1 ± 12.9 | 0.74 ± 0.08 | |
| 40 | 8 | 2.30 ± 0.45 | 48.4 ± 2.1 | 24.1 ± 4.7 | 46.5 ± 7.1 | 0.29 ± 0.06 |
Table 4.
Process metrics of 3-APB applying the multi-enzymatic system TA-PDC at different initial L-alanine concentrations. Reaction conditions: pH 7.5; 4-PB 10 mM; 30 °C; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal-5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2.
Table 4.
Process metrics of 3-APB applying the multi-enzymatic system TA-PDC at different initial L-alanine concentrations. Reaction conditions: pH 7.5; 4-PB 10 mM; 30 °C; TA 5% v·v−1; PDC 25% v·v−1, 1 mM pyridoxal-5’-phosphate (PLP), 0.1 mM thiamine pyrophosphate (TPP), and 0.1 mM MgCl2.
| Transaminase | Initial L-Alanine (mM) | Final APB (mM) | Conversion (%) | Yield (%) | Selectivity (%) | STY (mmol·L−1·h−1) |
|---|---|---|---|---|---|---|
| CviTA | 200 | 6.20 ± 0.98 | 72.0 ± 3.1 | 64.2 ± 10.1 | 88.7 ± 10.2 | 0.78 ± 0.12 |
| 400 | 6.22 ± 0.35 | 76.4 ± 3.6 | 59.5 ± 3.3 | 88.7 ± 10.2 | 0.78 ± 0.12 | |
| 600 | 6.88 ± 0.06 | 77.8 ± 1.6 | 71.7 ± 1.6 | 48.8 ± 7.6 | 0.38 ± 0.07 | |
| VflTA | 200 | 5.92 ± 0.64 | 71.0 ± 3.0 | 60.8 ± 6.6 | 86.1 ± 12.9 | 0.74 ± 0.08 |
| 400 | 6.60 ± 0.25 | 78.4 ± 4.4 | 68.4 ± 2.6 | 87.8 ± 8.2 | 0.82 ± 0.03 | |
| 600 | 7.08 ± 0.18 | 80.0 ± 6.2 | 73.2 ± 2.0 | 91.8 ± 4.6 | 0.89 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alcover, N.; Álvaro, G.; Guillén, M. Chiral Synthesis of 3-Amino-1-phenylbutane by a Multi-Enzymatic Cascade System. Catalysts 2021, 11, 973. https://doi.org/10.3390/catal11080973
AMA Style
Alcover N, Álvaro G, Guillén M. Chiral Synthesis of 3-Amino-1-phenylbutane by a Multi-Enzymatic Cascade System. Catalysts. 2021; 11(8):973. https://doi.org/10.3390/catal11080973
Chicago/Turabian StyleAlcover, Natàlia, Gregorio Álvaro, and Marina Guillén. 2021. "Chiral Synthesis of 3-Amino-1-phenylbutane by a Multi-Enzymatic Cascade System" Catalysts 11, no. 8: 973. https://doi.org/10.3390/catal11080973
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.