
Metallothionein 2A Expression in Cancer-Associated Fibroblasts and Cancer Cells Promotes Esophageal Squamous Cell Carcinoma Progression

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Preparation of CAF-Like Cells
2.3. Reverse Transcription Polymerase Chain Reaction (RT-PCR) and Quantitative Real-Time-PCR (qRT-PCR)
2.4. Western Blotting
2.5. Small Interfering (si)RNA in CAF-Like Cells and ESCC Cell Lines
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Antibody Array
2.8. Cell Survival and Growth Assay
2.9. Transwell Migration and Invasion Assay
2.10. Tissue Samples
2.11. Immunofluorescence
2.12. Immunohistochemistry
2.13. Statistical Analysis
3. Results
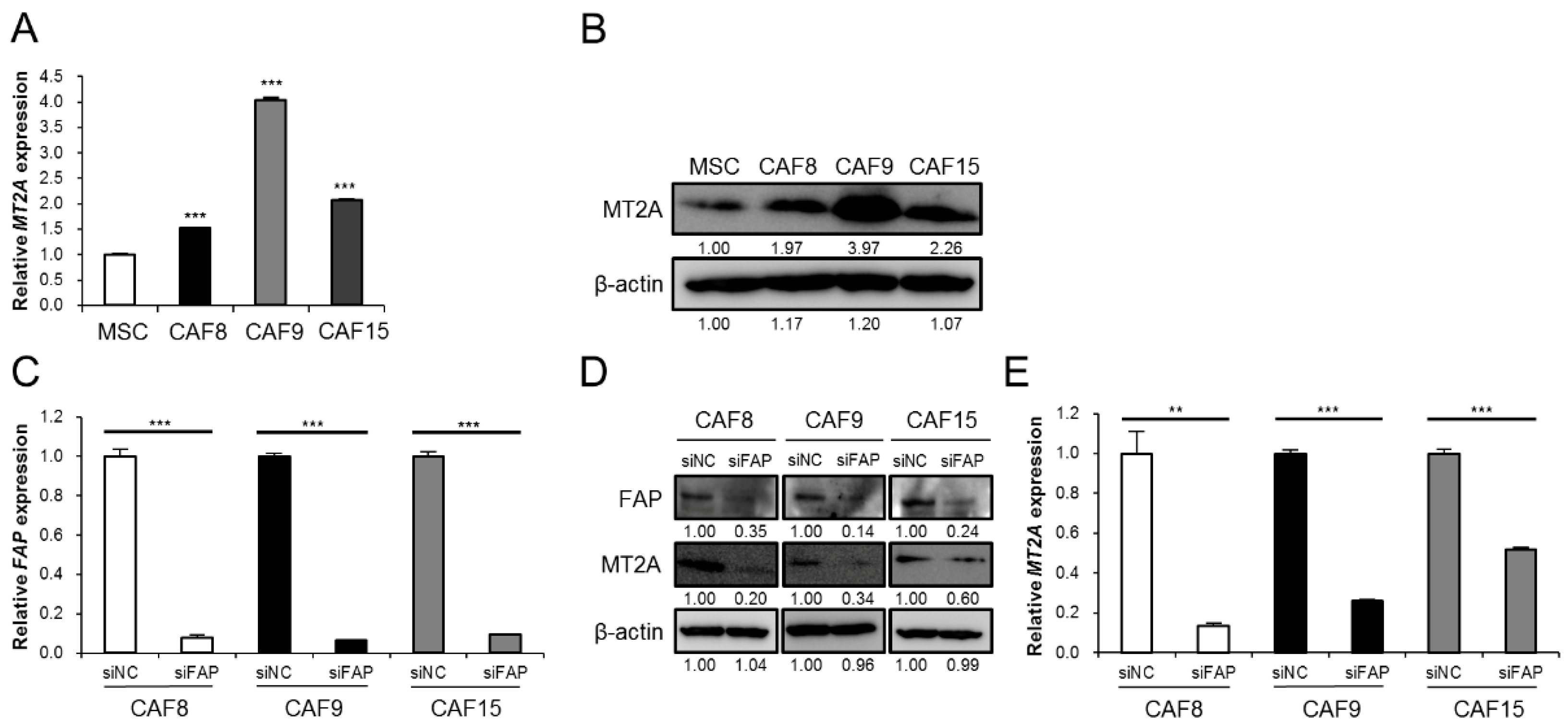
3.1. High Expression of MT2A in CAF-Like Cells
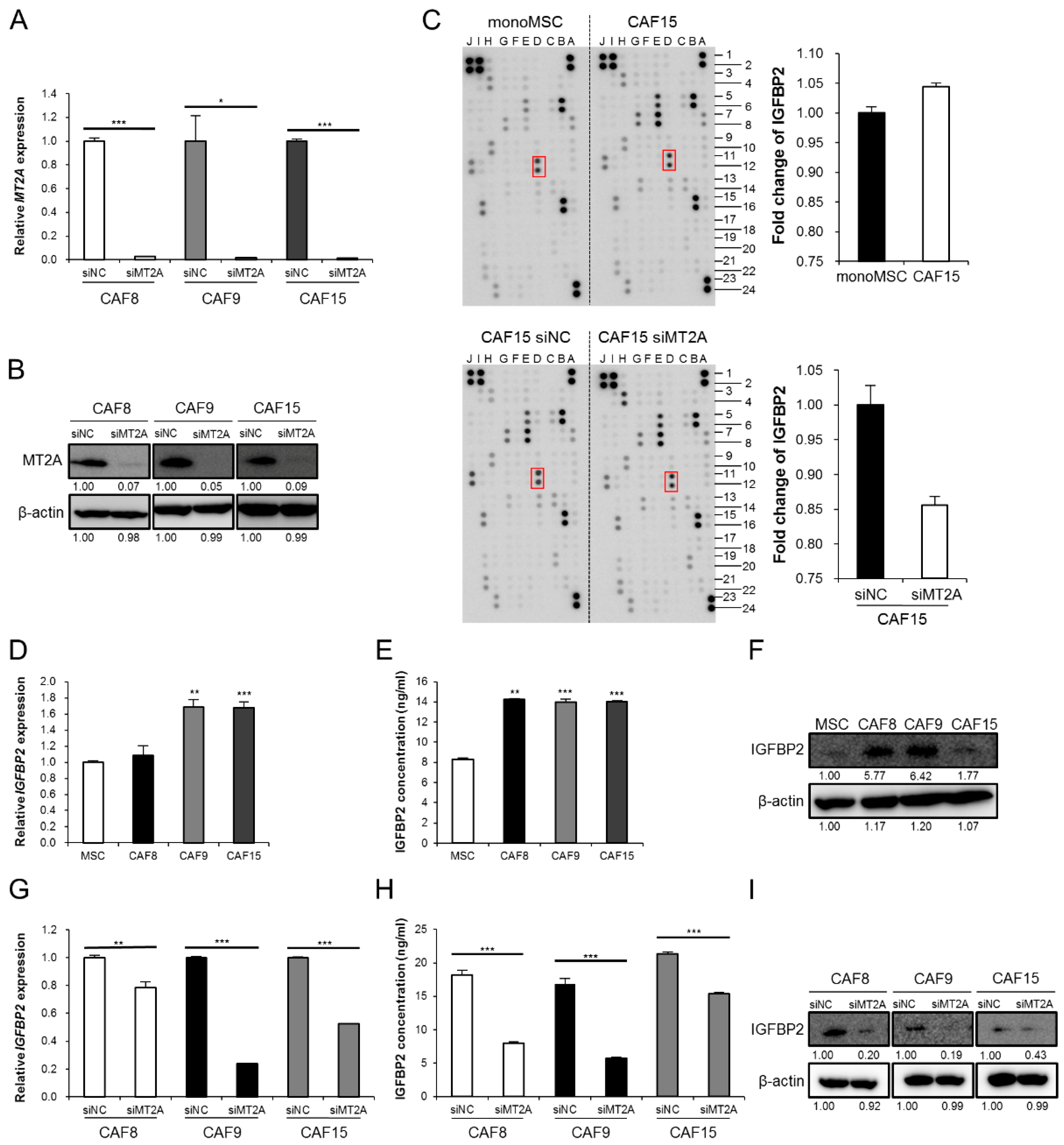
3.2. MT2A Induces Expression and Secretion of IGFBP2 in CAF-Like Cells
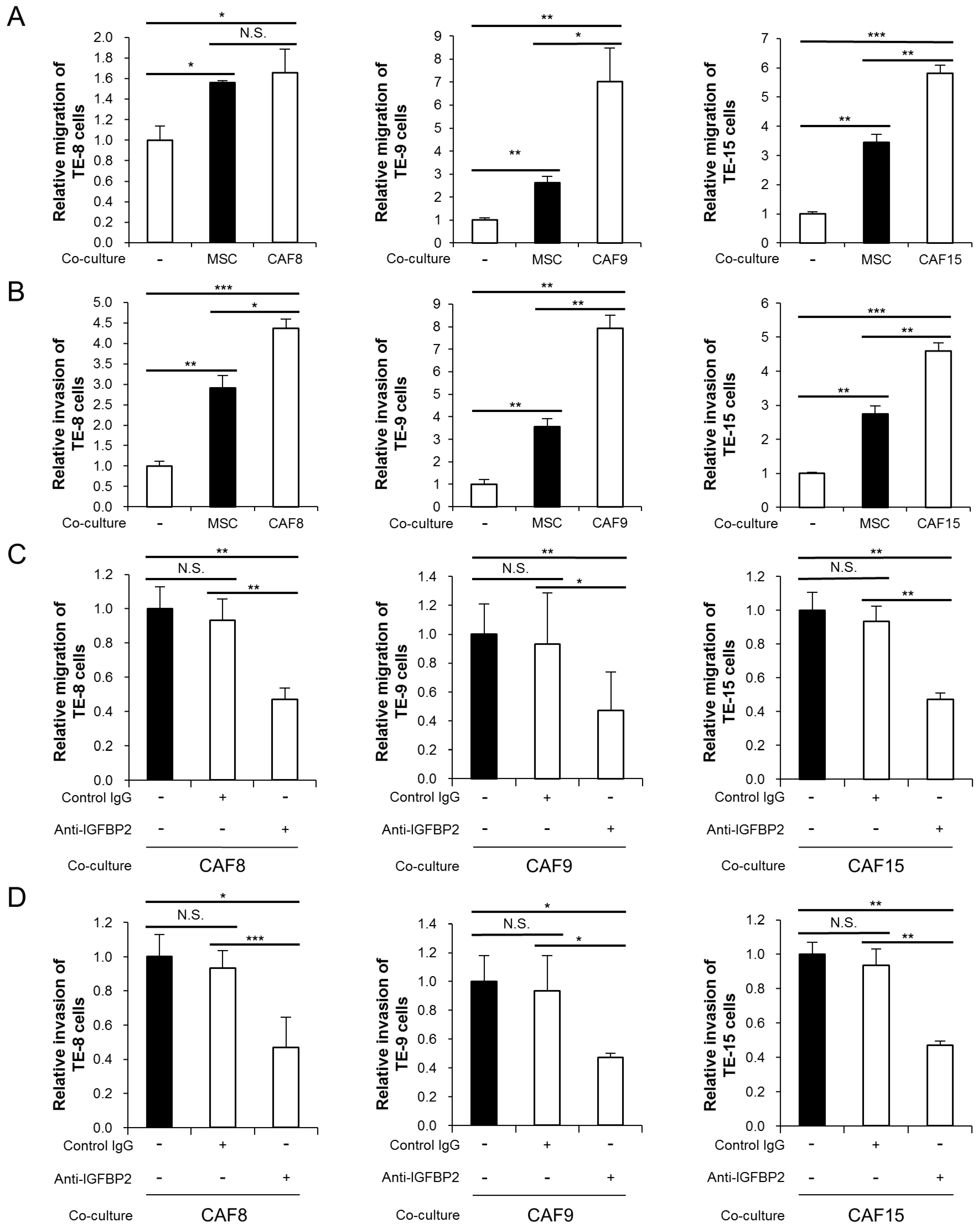
3.3. IGFBP2 Secreted by CAF-Like Cells Promotes Migration and Invasion of ESCC Cell Lines
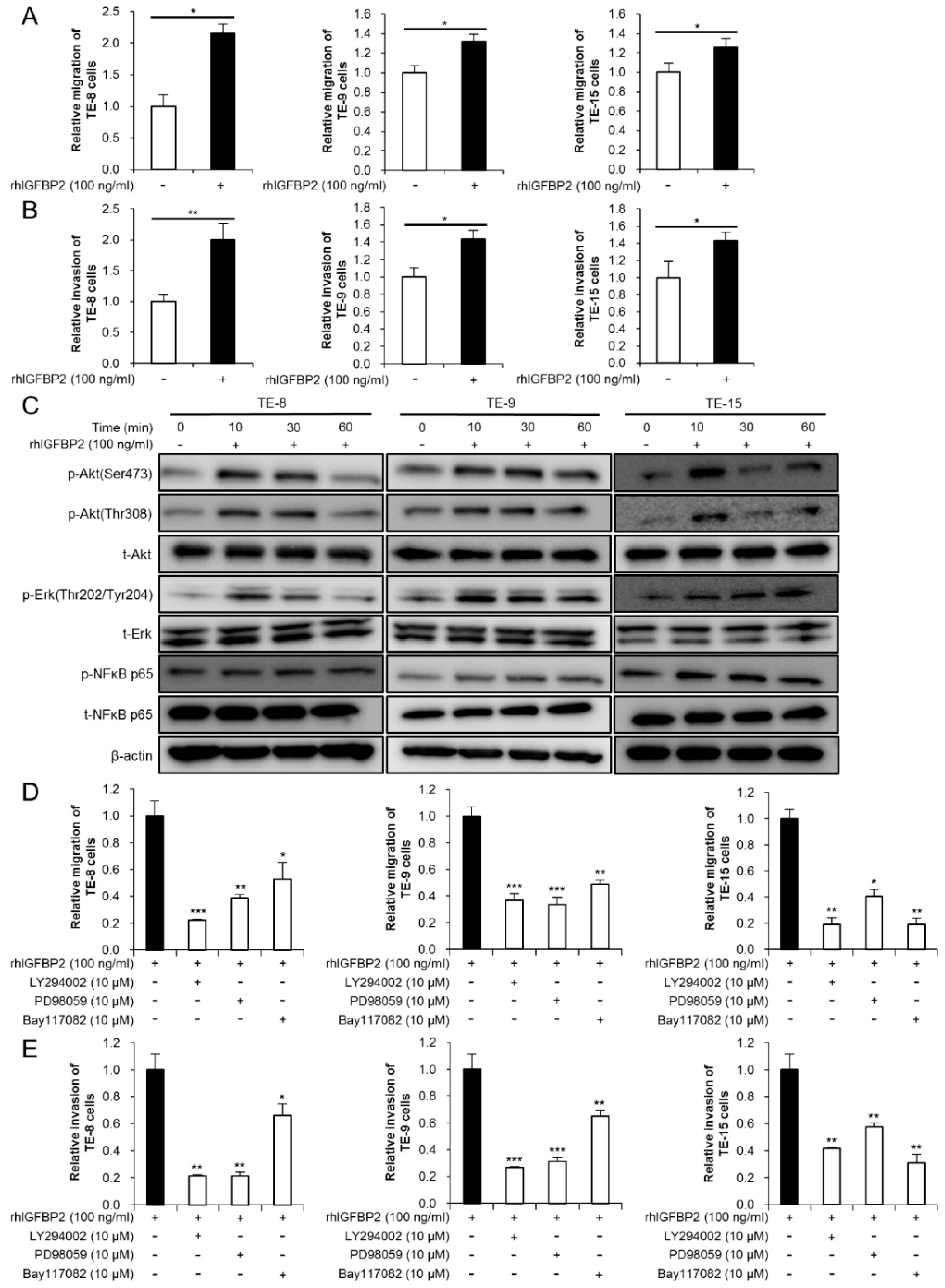
3.4. IGFBP2 Promotes Migration and Invasion through Akt, Erk, and NFκB Signaling Pathways in ESCC Cell Lines
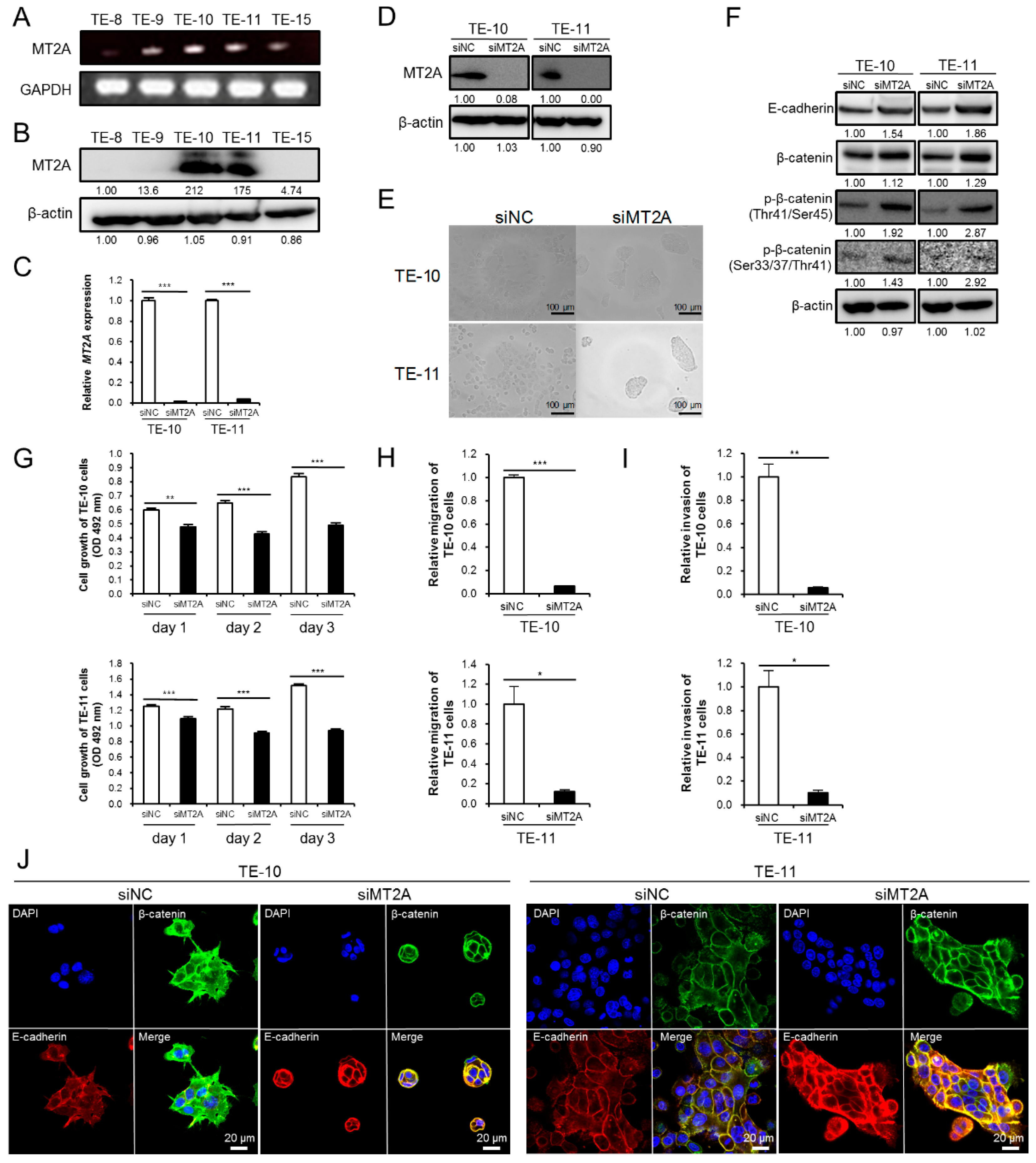
3.5. High Expression of MT2A in ESCC Cells Promotes Malignant Phenotype
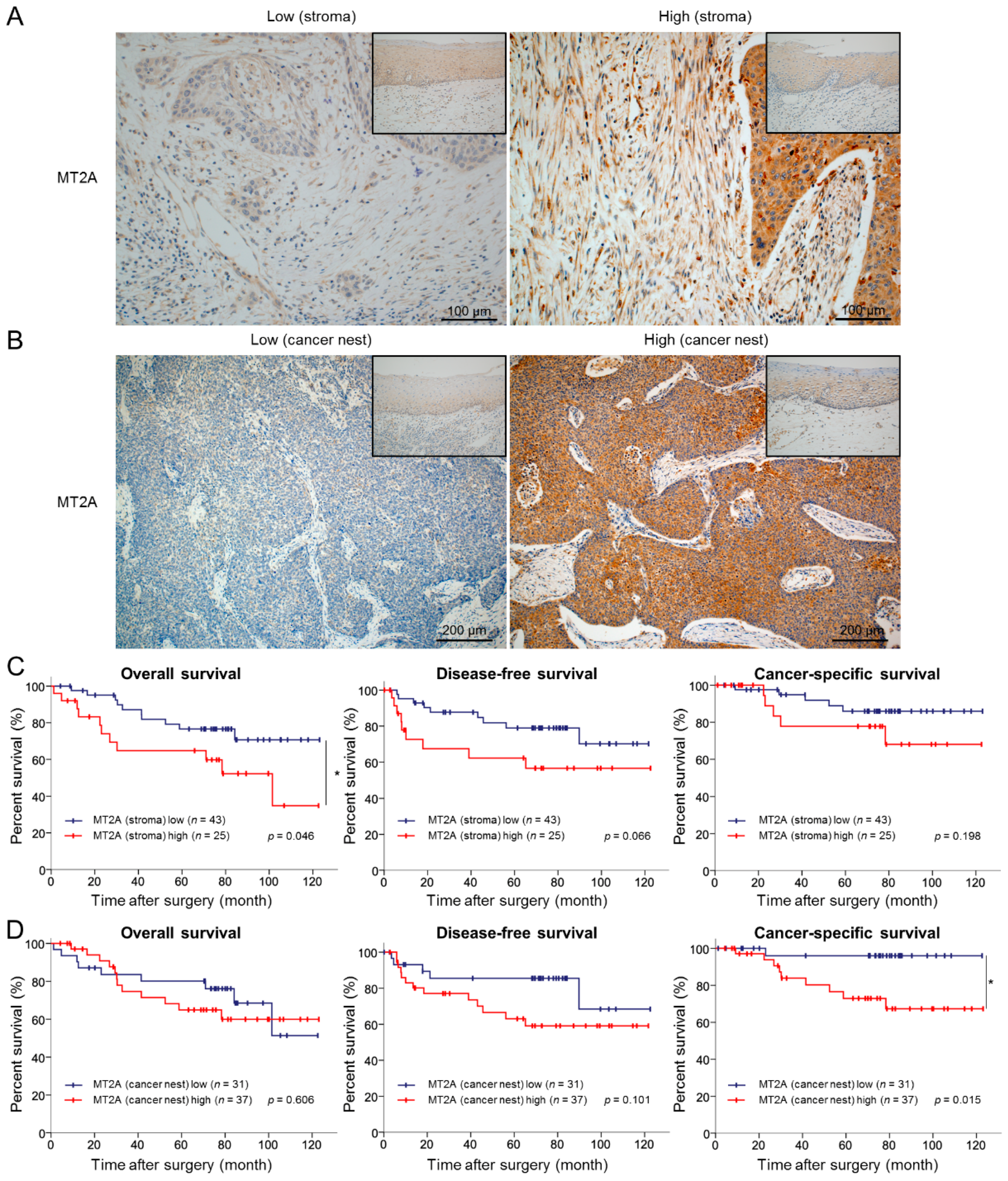
3.6. High Expression Levels of MT2A in the Cancer Stroma and Cancer Nest Correlate with Poor Prognosis of ESCC Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Short, M.W.; Burgers, K.G.; Fry, V.T. Esophageal Cancer. Am. Fam. Phys. 2017, 95, 22–28. [Google Scholar]
- Abnet, C.C.; Arnold, M.; Wei, W.-Q. Epidemiology of Esophageal Squamous Cell Carcinoma. Gastroenterology 2018, 154, 360–373. [Google Scholar] [CrossRef]
- Huang, F.-L.; Yu, S.-J. Esophageal cancer: Risk factors, genetic association, and treatment. Asian J. Surg. 2018, 41, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Koulaouzidis, A.; Marlicz, W.; Lok, V.; Chu, C.; Ngai, C.H.; Zhang, L.; Chen, P.; Wang, S.; Yuan, J.; et al. Global Burden, Risk Factors, and Trends of Esophageal Cancer: An Analysis of Cancer Registries from 48 Countries. Cancers 2021, 13, 141. [Google Scholar] [CrossRef] [PubMed]
- Tiasto, V.; Mikhailova, V.; Gulaia, V.; Vikhareva, V.; Zorin, B.; Kalitnik, A.; Kagansky, A. Esophageal cancer research today and tomorrow: Lessons from algae and other perspectives. AIMS Genet. 2018, 05, 075–090. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Tsukagoshi, H.; Fujita, M.; Hosokawa, M.; Kato, M.; Asaka, M. Long-term outcome after endoscopic mucosal resection in patients with esophageal squamous cell carcinoma invading the muscularis mucosae or deeper. Gastrointest. Endosc. 2002, 56, 387–390. [Google Scholar] [CrossRef]
- Kodama, M.; Kakegawa, T. Treatment of Superficial Cancer of the Esophagus: A Summary of Responses to a Questionnaire on Superficial Cancer of the Esophagus in Japan. Surgery 1998, 123, 432–439. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, L.; Xia, W.; Wang, F. Anatomy of lymphatic drainage of the esophagus and lymph node metastasis of thoracic esophageal cancer. Cancer Manag. Res. 2018, 10, 6295–6303. [Google Scholar] [CrossRef] [Green Version]
- Barret, M.; Prat, F. Diagnosis and treatment of superficial esophageal cancer. Ann. Gastroenterol. 2018, 31, 256–265. [Google Scholar] [CrossRef]
- Weijs, T.J.; Ruurda, J.P.; Luyer, M.D.P.; Cuesta, M.A.; Van Hillegersberg, R.; Bleys, R.L.A.W. New insights into the surgical anatomy of the esophagus. J. Thorac. Dis. 2017, 9, S675–S680. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Dai, Y. Tumor microenvironment and therapeutic response. Cancer Lett. 2017, 387, 61–68. [Google Scholar] [CrossRef]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Ge, J.; Xiang, B.; Wu, X.; Ma, J.; Zhou, M.; Li, X.; et al. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol. Cancer 2019, 18, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raskov, H.; Orhan, A.; Gaggar, S.; Gögenur, I. Cancer-Associated Fibroblasts and Tumor-Associated Macrophages in Cancer and Cancer Immunotherapy. Front. Oncol. 2021, 11, 668731. [Google Scholar] [CrossRef]
- Higashino, N.; Koma, Y.-I.; Hosono, M.; Takase, N.; Okamoto, M.; Kodaira, H.; Nishio, M.; Shigeoka, M.; Kakeji, Y.; Yokozaki, H. Fibroblast activation protein-positive fibroblasts promote tumor progression through secretion of CCL2 and interleukin-6 in esophageal squamous cell carcinoma. Lab. Investig. 2019, 99, 777–792. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, H.; Koma, Y.-I.; Higashino, N.; Kodama, T.; Tanigawa, K.; Shimizu, M.; Fujikawa, M.; Nishio, M.; Shigeoka, M.; Kakeji, Y.; et al. PAI-1 derived from cancer-associated fibroblasts in esophageal squamous cell carcinoma promotes the invasion of cancer cells and the migration of macrophages. Lab. Investig. 2021, 101, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Yan, M.; Wang, X.; Xu, Q.; Wang, X.; Zhu, X.; Shi, J.; Li, Z.; Zhang, J.; Chen, W. Cancer-associated Fibroblast-derived IL-6 Promotes Head and Neck Cancer Progression via the Osteopontin-NF-kappa B Signaling Pathway. Theranostics 2018, 8, 921–940. [Google Scholar] [CrossRef]
- Tsuyada, A.; Chow, A.; Wu, J.; Somlo, G.; Chu, P.; Loera, S.; Luu, T.; Li, A.X.; Wu, X.; Ye, W.; et al. CCL2 Mediates Cross-talk between Cancer Cells and Stromal Fibroblasts That Regulates Breast Cancer Stem Cells. Cancer Res. 2012, 72, 2768–2779. [Google Scholar] [CrossRef] [Green Version]
- Che, Y.; Wang, J.; Li, Y.; Lu, Z.; Huang, J.; Sun, S.; Mao, S.; Lei, Y.; Zang, R.; Sun, N.; et al. Cisplatin-activated PAI-1 secretion in the cancer-associated fibroblasts with paracrine effects promoting esophageal squamous cell carcinoma progression and causing chemoresistance. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Japan Esophageal Society. Japanese Classification of Esophageal Cancer, 10th ed.; Kanehara & Co.: Tokyo, Japan, 2008. [Google Scholar]
- Sobin, L.H.; Gospodarowicz, M.K.; Wittekind, C. TNM Classification of Malignant Tumours, 7th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2011. [Google Scholar]
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 127, 469–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigeoka, M.; Urakawa, N.; Nakamura, T.; Nishio, M.; Watajima, T.; Kuroda, D.; Komori, T.; Kakeji, Y.; Semba, S.; Yokozaki, H. Tumor associated macrophage expressing CD204 is associated with tumor aggressiveness of esophageal squamous cell carcinoma. Cancer Sci. 2013, 104, 1112–1119. [Google Scholar] [CrossRef]
- Thirumoorthy, N.; Manisenthil Kumar, K.T.; Shyam Sundar, A.; Panayappan, L.; Chatterjee, M. Metallothionein: An overview. World J. Gastroenterol. 2007, 13, 993–996. [Google Scholar] [CrossRef] [Green Version]
- Atrian, S.; Capdevila, M. Metallothionein-protein interactions. Biomol. Concepts 2013, 4, 143–160. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, G.; Li, W. MT2A Promotes Oxaliplatin Resistance in Colorectal Cancer Cells. Cell Biochem. Biophys. 2020, 78, 475–482. [Google Scholar] [CrossRef]
- Borchert, S.; Suckrau, P.-M.; Walter, R.F.H.; Wessolly, M.; Mairinger, E.; Steinborn, J.; Hegedus, B.; Hager, T.; Herold, T.; Eberhardt, W.E.E.; et al. Impact of metallothionein-knockdown on cisplatin resistance in malignant pleural mesothelioma. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tekur, S.; Ho, S.M. Ribozyme-mediated downregulation of human metallothionein II(a) induces apoptosis in human prostate and ovarian cancer cell lines. Mol. Carcinog. 2002, 33, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Huang, J.; Xing, R.; Yin, X.; Cui, J.; Li, W.; Yu, J.; Lu, Y. Metallothionein 2A inhibits NF-κB pathway activation and predicts clinical outcome segregated with TNM stage in gastric cancer patients following radical resection. J. Transl. Med. 2013, 11, 173. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Peng, W.; Wu, B.; Liu, H.; Zhang, R.; Zhou, R.; Yao, L.; Ye, A.L. Metallothionein MT1M Suppresses Carcinogenesis of Esophageal Carcinoma Cells through Inhibition of the Epithelial-Mesenchymal Transition and the SOD1/PI3K Axis. Mol. Cells 2021, 44, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Mageed, A.B.; Agrawal, K.C. Activation of nuclear factor kappaB: Potential role in metallothionein-mediated mitogenic response. Cancer Res. 1998, 58, 2335–2338. [Google Scholar]
- Kim, H.G.; Kim, J.Y.; Han, E.H.; Hwang, Y.P.; Choi, J.H.; Park, B.H.; Jeong, H.G. Metallothionein-2A overexpression increases the expression of matrix metalloproteinase-9 and invasion of breast cancer cells. FEBS Lett. 2010, 585, 421–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marikar, F.M.; Jin, G.; Sheng, W.; Ma, D.; Hua, Z. Metallothionein 2A an interactive protein linking phosphorylated FADD to NF-κB pathway leads to colorectal cancer formation. Chin. Clin. Oncol. 2016, 5, 76. [Google Scholar] [CrossRef]
- Sakurai, A.; Hara, S.; Okano, N.; Kondo, Y.; Inoue, J.-I.; Imura, N. Regulatory role of metallothionein in NF-κB activation. FEBS Lett. 1999, 455, 55–58. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.-P.; Merin, J.P.; Nakano, T.; Kato, T.; Kitade, Y.; Okamoto, T. Inhibition of the DNA-binding activity of NF-κB by gold compounds in vitro. FEBS Lett. 1995, 361, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Penkowa, M.; Carrasco, J.; Giralt, M.; Molinero, A.; Hernández, J.; Campbell, I.L.; Hidalgo, J. Altered Central Nervous System Cytokine-Growth Factor Expression Profiles and Angiogenesis in Metallothionein-I+II Deficient Mice. J. Cereb. Blood Flow Metab. 2000, 20, 1174–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemmons, D.R. Insulinlike growth factor binding proteins. Trends Endocrinol. Metab. 1990, 1, 412–417. [Google Scholar] [CrossRef]
- Baxter, R.C. Circulating binding proteins for the insulinlike growth factors. Trends Endocrinol. Metab. 1993, 4, 91–96. [Google Scholar] [CrossRef]
- Li, T.; Forbes, M.E.; Fuller, G.N.; Li, J.; Yang, X.; Zhang, W. IGFBP2: Integrative hub of developmental and oncogenic signaling network. Oncogene 2020, 39, 2243–2257. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Sun, Y.; Zhang, X.; Hu, L.; Liu, Y.; Chua, C.Y.; Phillips, L.M.; Ren, H.; Fleming, J.B.; Wang, H.; et al. IGFBP2 Activates the NF-κB Pathway to Drive Epithelial–Mesenchymal Transition and Invasive Character in Pancreatic Ductal Adenocarcinoma. Cancer Res. 2016, 76, 6543–6554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Zhang, X.; Song, Q.; Liu, L.; Forbes, E.; Tian, W.; Zhang, Z.; Kang, Y.; Wang, H.; Fleming, J.B.; et al. IGFBP2 promotes tumor progression by inducing alternative polarization of macrophages in pancreatic ductal adenocarcinoma through the STAT3 pathway. Cancer Lett. 2021, 500, 132–146. [Google Scholar] [CrossRef]
- Ma, Y.; Cui, D.; Zhang, Y.; Han, C.-C.; Wei, W. Insulin-Like Growth Factor Binding Protein-2 Promotes Proliferation and Predicts Poor Prognosis in Hepatocellular Carcinoma. OncoTargets Ther. 2020, 13, 5083–5092. [Google Scholar] [CrossRef]
- Chua, C.Y.; Liu, Y.; Granberg, K.J.; Hu, L.; Haapasalo, H.; Annala, M.J.; Cogdell, D.E.; Verploegen, M.; Moore, L.M.; Fuller, G.; et al. IGFBP2 potentiates nuclear EGFR–STAT3 signaling. Oncogene 2016, 35, 738–747. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Chen, Q.; Cui, Y.; Dong, J.; Chen, M.; Wu, P.; Jiang, C. Immune heterogeneity and clinicopathologic characterization of IGFBP2 in 2447 glioma samples. OncoImmunology 2018, 7, e1426516. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.K.; Hu, L.; Fuller, G.N.; Zhang, W. An Interaction between Insulin-like Growth Factor-binding Protein 2 (IGFBP2) and Integrin α5 Is Essential for IGFBP2-induced Cell Mobility. J. Biol. Chem. 2006, 281, 14085–14091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Li, Z.; Master, L.M.; Master, Z.W.; Wu, A. Exogenous IGFBP-2 promotes proliferation, invasion, and chemoresistance to temozolomide in glioma cells via the integrin β1-ERK pathway. Br. J. Cancer 2014, 111, 1400–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azar, W.J.; Zivkovic, S.; Werther, G.A.; Russo, V.C. IGFBP-2 nuclear translocation is mediated by a functional NLS sequence and is essential for its pro-tumorigenic actions in cancer cells. Oncogene 2014, 33, 578–588. [Google Scholar] [CrossRef] [Green Version]
- Aquime, J.R.H.S.; Zampieri, L.C.D.P.; Kataoka, M.S.D.S.; Ribeiro, N.A.B.; Jaeger, R.G.; Da Silva, A.L.; Ramos, R.T.J.; Júnior, S.D.M.A.; Pinheiro, J.D.J.V. Metallothionein Expression and its Influence on the In Vitro Biological Behavior of Mucoepidermoid Carcinoma. Cells 2020, 9, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, T.-S.; Gao, W.; Chan, J.Y.-W. Transcription Regulation of E-Cadherin by Zinc Finger E-Box Binding Homeobox Proteins in Solid Tumors. BioMed Res. Int. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Mendonsa, A.M.; Na, T.-Y.; Gumbiner, B.M. E-cadherin in contact inhibition and cancer. Oncogene 2018, 37, 4769–4780. [Google Scholar] [CrossRef]
- Stamos, J.L.; Weis, W.I. The β-Catenin Destruction Complex. Cold Spring Harb. Perspect. Biol. 2012, 5, a007898. [Google Scholar] [CrossRef]
- Onder, T.T.; Gupta, P.B.; Mani, S.A.; Yang, J.; Lander, E.S.; Weinberg, R.A. Loss of E-Cadherin Promotes Metastasis via Multiple Downstream Transcriptional Pathways. Cancer Res. 2008, 68, 3645–3654. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Liu, Y.; Li, M.-Y.; Ng, C.S.H.; Yang, S.-L.; Wang, S.; Zou, C.; Dong, Y.; Du, J.; Long, X.; et al. FOXP3 promotes tumor growth and metastasis by activating Wnt/β-catenin signaling pathway and EMT in non-small cell lung cancer. Mol. Cancer 2017, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hishikawa, Y.; Koji, T.; Dhar, D.K.; Kinugasa, S.; Yamaguchi, M.; Nagasue, N. Metallothionein expression correlates with metastatic and proliferative potential in squamous cell carcinoma of the oesophagus. Br. J. Cancer 1999, 81, 712–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, L.M.; Zhou, X.; Cogdell, D.E.; Chua, C.Y.; Huisinga, A.; Hess, K.R.; Fuller, G.N.; Zhang, W. Glioma progression is mediated by an addiction to aberrant IGFBP2 expression and can be blocked using anti-IGFBP2 strategies. J. Pathol. 2016, 239, 355–364. [Google Scholar] [CrossRef]
- Patil, S.S.; Railkar, R.; Swain, M.; Atreya, H.S.; Dighe, R.R.; Kondaiah, P. Novel anti IGFBP2 single chain variable fragment inhibits glioma cell migration and invasion. J. Neuro-Oncol. 2015, 123, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Tschammer, N.; Mohammed, S.I.; Guo, P. Specific Delivery of Therapeutic RNAs to Cancer Cells via the Dimerization Mechanism of phi29 Motor pRNA. Hum. Gene Ther. 2005, 16, 1097–1110. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Siegwart, D.J.; Anderson, D.G. Strategies, design, and chemistry in siRNA delivery systems. Adv. Drug Deliv. Rev. 2019, 144, 133–147. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Expression of MT2A in Cancer Stroma 1 | Expression of MT2A in Cancer Nest 1 | ||||||
|---|---|---|---|---|---|---|---|---|
| Number of Cases | Low | High | Number of Cases | Low | High | |||
| (n = 43) | (n = 26) | p-Value | (n = 31) | (n = 38) | p-Value | |||
| Age | ||||||||
| <65 | 32 | 20 | 12 | 0.997 | 32 | 13 | 19 | 0.504 |
| ≥65 | 37 | 23 | 14 | 37 | 18 | 19 | ||
| Sex | ||||||||
| Male | 14 | 8 | 6 | 0.654 | 14 | 6 | 8 | 0.862 |
| Female | 55 | 35 | 20 | 55 | 25 | 30 | ||
| Histological grade 2 | ||||||||
| HGIEN + WDSCC | 15 | 11 | 4 | 0.320 | 15 | 8 | 7 | 0.459 |
| MDSCC + PDSCC | 54 | 32 | 22 | 54 | 23 | 31 | ||
| Depth of tumor invasion 2 | ||||||||
| T1 | 48 | 36 | 12 | 0.001 ** | 48 | 25 | 23 | 0.071 |
| T2+T3 | 21 | 7 | 14 | 21 | 6 | 15 | ||
| Lymphatic vessel invasion 2 | ||||||||
| Negative | 37 | 31 | 6 | <0.001 *** | 37 | 19 | 18 | 0.249 |
| Positive | 32 | 12 | 20 | 32 | 12 | 20 | ||
| Blood vessel invasion 2 | ||||||||
| Negative | 43 | 32 | 11 | 0.008 ** | 43 | 20 | 23 | 0.734 |
| Positive | 26 | 11 | 15 | 26 | 11 | 15 | ||
| Lymph node metastasis 2 | ||||||||
| Negative | 43 | 31 | 12 | 0.031 * | 43 | 22 | 21 | 0.181 |
| Positive | 26 | 12 | 14 | 26 | 9 | 17 | ||
| Stage 3 | ||||||||
| 0 + I | 38 | 28 | 10 | 0.031 * | 38 | 19 | 19 | 0.348 |
| II + III + IV | 31 | 15 | 16 | 31 | 12 | 19 | ||
| Expression of αSMA 4 | ||||||||
| Low | 36 | 31 | 5 | <0.001 *** | 36 | 16 | 20 | 0.933 |
| High | 33 | 12 | 21 | 33 | 15 | 18 | ||
| Expression of FAP 4 | ||||||||
| Low | 39 | 33 | 6 | <0.001 *** | 39 | 19 | 20 | 0.470 |
| High | 30 | 10 | 20 | 30 | 12 | 18 | ||
| Expression of CD163 5 | ||||||||
| Low | 34 | 27 | 7 | 0.004 ** | 34 | 16 | 18 | 0.726 |
| High | 35 | 16 | 19 | 35 | 15 | 20 | ||
| Expression of CD204 5 | ||||||||
| Low | 34 | 31 | 3 | <0.001 *** | 34 | 16 | 18 | 0.726 |
| High | 35 | 12 | 23 | 35 | 15 | 20 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimizu, M.; Koma, Y.-i.; Sakamoto, H.; Tsukamoto, S.; Kitamura, Y.; Urakami, S.; Tanigawa, K.; Kodama, T.; Higashino, N.; Nishio, M.; et al. Metallothionein 2A Expression in Cancer-Associated Fibroblasts and Cancer Cells Promotes Esophageal Squamous Cell Carcinoma Progression. Cancers 2021, 13, 4552. https://doi.org/10.3390/cancers13184552
Shimizu M, Koma Y-i, Sakamoto H, Tsukamoto S, Kitamura Y, Urakami S, Tanigawa K, Kodama T, Higashino N, Nishio M, et al. Metallothionein 2A Expression in Cancer-Associated Fibroblasts and Cancer Cells Promotes Esophageal Squamous Cell Carcinoma Progression. Cancers. 2021; 13(18):4552. https://doi.org/10.3390/cancers13184552
Chicago/Turabian StyleShimizu, Masaki, Yu-ichiro Koma, Hiroki Sakamoto, Shuichi Tsukamoto, Yu Kitamura, Satoshi Urakami, Kohei Tanigawa, Takayuki Kodama, Nobuhide Higashino, Mari Nishio, and et al. 2021. "Metallothionein 2A Expression in Cancer-Associated Fibroblasts and Cancer Cells Promotes Esophageal Squamous Cell Carcinoma Progression" Cancers 13, no. 18: 4552. https://doi.org/10.3390/cancers13184552