
Arginine Signaling and Cancer Metabolism


1
Institute of Molecular and Genomic Medicine, National Health Research Institutes, Zhunan 350, Miaoli County, Taiwan
2
Institute of Biotechnology, National Tsing-Hua University, Hsinchu 30035, Taiwan
3
Institute of Cellular and System Medicine, National Health Research Institutes, Zhunan 350, Miaoli County, Taiwan
4
Department of Diabetes and Metabolic Diseases Research, Irell & Manella Graduate School of Biological Sciences, Beckman Research Institute, City of Hope, Duarte, CA 91010, USA
5
Ph.D. Program for Cancer Biology and Drug Discovery, College of Medical Science and Technology, Taipei Medical University, Taipei 110, Taiwan
6
Research Center of Cancer Translational Medicine, Taipei Medical University, Taipei 110, Taiwan
7
Comprehensive Cancer Center, Department of Biochemistry and Molecular Medicine, University of California at Davis, Sacramento, CA 95817, USA
*
Author to whom correspondence should be addressed.
Cancers 2021, 13(14), 3541; https://doi.org/10.3390/cancers13143541
Submission received: 10 June 2021
/
Revised: 1 July 2021
/
Accepted: 12 July 2021
/
Published: 15 July 2021
Abstract
:Simple Summary
In this review, we describe arginine’s role as a signaling metabolite, epigenetic regulator and mitochondrial modulator in cancer cells, and summarize recent progress in the application of arginine deprivation as a cancer therapy.
Abstract
Arginine is an amino acid critically involved in multiple cellular processes including the syntheses of nitric oxide and polyamines, and is a direct activator of mTOR, a nutrient-sensing kinase strongly implicated in carcinogenesis. Yet, it is also considered as a non- or semi-essential amino acid, due to normal cells’ intrinsic ability to synthesize arginine from citrulline and aspartate via ASS1 (argininosuccinate synthase 1) and ASL (argininosuccinate lyase). As such, arginine can be used as a dietary supplement and its depletion as a therapeutic strategy. Strikingly, in over 70% of tumors, ASS1 transcription is suppressed, rendering the cells addicted to external arginine, forming the basis of arginine-deprivation therapy. In this review, we will discuss arginine as a signaling metabolite, arginine’s role in cancer metabolism, arginine as an epigenetic regulator, arginine as an immunomodulator, and arginine as a therapeutic target. We will also provide a comprehensive summary of ADI (arginine deiminase)-based arginine-deprivation preclinical studies and an update of clinical trials for ADI and arginase. The different cell killing mechanisms associated with various cancer types will also be described.
1. Introduction
An important step in tumor development is a metabolic adaptation to cope with the demand of rapid cell division as well as a hypoxia, and nutritionally deprived microenvironment [1]. Different tumors utilize different strategies to reprogram their metabolic pathways. In so doing, tumor cells expose specific vulnerabilities, which can be exploited therapeutically. For instance, tumor cells, not their normal counterparts, are “addicted” to certain external nutrients including amino acids and amino acid starvation therapy has gained significant momentum in recent years [2]. One of the most common metabolic defects of tumor cells is the impaired intrinsic ability to synthesize arginine [3]. Targeting exogenous arginine by arginine-metabolizing enzymes such as arginase, arginine decarboxylase and arginine deiminase (ADI) has received increasing attention as therapies to treat a variety of cancers [4]. There are a number of excellent reviews on this topic [5,6,7]. In this review, we will focus on recent progress in understanding arginine’s role in cancer metabolism as a signaling metabolite, an epigenetic regulator and an immunomodulator. As much of the knowledge was derived from characterizing arginine-deprived cancer cells, we will also update the current status of arginine-deprivation therapy.
2. Arginine and Cancer Metabolism
Arginine is involved in numerous biological functions including cell proliferation, cell signaling, muscle contraction, immunity, neurotransmission, vasodilation, synthesis of growth factors and other amino acids [8]. Three major resources of arginine include (1) arginine-enriched nutrition supplement from dietary intake, such as chicken, pork loin, pumpkin seeds, peanuts, soybeans and so on (approximately 5 g of arginine per day), (2) endogenous synthesis from citrulline (15% of the total arginine production) and (3) protein catabolism (approximately 80% of the circulating arginine) [9]. Arginine is a direct resource for nitric oxide (NO), ornithine and agmatine through three enzyme reactions. Arginine is metabolized into NO and citrulline by nitric oxide synthase (NOS), into ornithine and urea by arginase, and into agmatine by arginine decarboxylase (ADC). NO serves multiple physiological functions.
Both ornithine and agmatine are the main resources for putrescine, which is a crucial precursor for polyamines [10] (Figure 1). These metabolites play key functions in cell physiology and human health so does arginine. Yet, arginine is also considered a “semi-essential” or “conditional essential” amino acid. This is due to normal cells’ intrinsic ability to synthesize this molecule from citrulline and aspartate via argininosuccinate synthase 1 (ASS1) and argininosuccinate lyase (ASL) in the urea cycle [11]. Citrulline can be synthesized from glutamate [12], proline [13] and glutamine [14]. As such, normal cells do not completely depend on external arginine. Yet, many cancer cells are “addicted” to external arginine.
Much of our current understanding of the arginine effects on cancer metabolism is derived from arginine-deprivation studies of ASS1-low cancer cells. The responses depend on the cell types studied but generally can be put into two categories.
For breast and prostate cancer cells, arginine starvation causes global transcriptional suppression of metabolic genes including those involved in oxidative phosphorylation (OXPHOS) and mitochondrial functions, glycolysis, purine and pyrimidine synthesis, DNA repair genes [15,16,17,18], due to epigenetic remodeling [19,20]. Removal of arginine causes fragmentation of mitochondria and impairment of mitochondrial functions as measured by OCR (oxygen consumption rate) and membrane potential [15,17,20,21]. The suppressed transcription of genes involved in mitochondria functions is reflected by the metabolomics studies which show the general depletion of TCA cycle metabolites such αKG, malate, fumarate and succinate [17,20]. It was suggested that at least in breast cancer cells, such depletions are in part due to the ER-stress mediated activation of ASNS (asparagine synthetase), which depletes aspartate and diminishes aspartate-malate shuttle, negatively impacting TCA [17]. The perturbation of OXPHOS reaction due to metabolite depletion, perpetuated by the transcriptional suppression of nuclear-encoded OXPHOS genes, generates copious amount of mtROS which leads to DNA damage and eventual cell death [15,16,17,20]. Indeed, in these cells, functional knockout of mitochondria prevented DNA damage and cell death [17]. Additional evidence for impairment of mitochondria by arginine deprivation includes the morphological changes from hyperfusion at early stage to fragmentation at late state [17,20]. This transition is caused by the reduced expression of Mfn2 (mitofusin2) mediated by arginine deprivation-induced p38 activation and KAP-1 phosphorylation [21].
By contrast, arginine deprivation of ASS1-low melanoma and sarcoma cells leads to downmodulation of glycolysis pathway with increased glutamine anaplerosis and serine synthesis to sustain the TCA cycle [22,23]. The authors suggest the inhibition of the Warburg effect is one reason how cancer cells adapt to the stress environment and eventually become resistant to the treatment. A characteristic of these cells is the activation of c-myc upon arginine deprivation [24,25], which is not necessarily the case in prostate and breast cancers described above and may explain the differences in their responses to arginine deprivation. c-myc is known to upregulate glutaminase and render cells addicted to glutamine [26]. Accordingly, these types of tumors are synthetic lethal with glutaminase inhibitor [22].
In either of the above cases, mitochondria are targeted by arginine deprivation and arginine represents a major regulator of mitochondrial activities in cancer metabolism.
3. Arginine and Signal Transduction
3.1. Arginine Mediated Signals
There are at least two ways arginine can transmit signals to the cells. The first is through transporters, solute carriers (SLCs) (Figure 2). As a cationic amino acid, arginine is mainly imported by two types of SLCs, the cationic amino acid transporters and the system y+L amino acid transporters [27,28]. It is noteworthy that arginine activates its downstream mTOR signal via lysosomal SLC38A9 [29]. Arginine is the most consumed amino acid in the inner necrotic core of tumor mass, indicating its high demand for the survival of tumor cells [30]. Accordingly, tumor cells frequently overexpress specific types of SLCs such as SLC6A14, SLC7A3, SLC7A9, etc. to meet their high arginine demand (Table 1). It should be noted that T cells up-regulate distinct types of SLCs to increase arginine uptake for T cell activation and anti-tumor functions [31,32] (Table 1). Thus, targeting the tumor specifically with SCLs, and avoiding those expressed in T-cell and macrophage (e.g., SLC7A1, and A2) could be a potential strategy for cancer therapy.
Subsequent to its transport, arginine is able to activate several signal pathways. Chief among them is mTOR kinase. Arginine is one of only three amino acids that can directly activate mTOR pathway, a major cellular sensor of nutritional state [47]. The other two are glutamine and leucine. As such, arginine has profound impacts on protein synthesis, lipid synthesis and nucleotide synthesis, three anabolic pathways mediated by mTOR [48]. Indeed, nutrients [49,50]. are as important as growth factors in the activation of mTOR. Upon growth factor stimulation, mTOR can be activated through either PI3K (phosphatylinositol 3-kinase) pathway or MAPK pathway, via the inactivation of TSC (tuberous sclerosis complex), an mTOR negative regulator [51]. Inhibition of TSC converts the Rheb (RAS homolog enriched in brain) into active form, resulting in the activation of mTORC1 (mTOR complex 1). There are at least three ways, arginine can activate mTOR. (1) arginine disrupts the interaction between TSC and mTORC1, thereby activating mTOR [52]. (2) in the lysosome, arginine interacts with SLC38A9 and v-ATPase, upstream regulators of mTORC1, leading to the activation of Rag GTPase that is required for recruitment of mTORC1 complex to the lysosomal surface [53,54]. (3) in the cytosol, arginine interacts with CASTOR1(cytosolic arginine sensor for mTORC1 subunit 1) to disrupt the CASTOR complex, which is a negative regulator of Rag A [55], allowing Rag A to bind mTORC1 component RAPTOR (Regulatory-associated protein of mTOR) and redistributes mTORC1 to the lysosome. This may explain why arginine is such a potent activator of mTOR, and arginine deprivation leads to immediate inactivation of mTOR.
The second means arginine can transmit signal is through binding to L-amino acid receptor, G-protein coupled receptor GPRCA6 [56]. In human fibroblast, GPRCA6 was shown to activate PKA (activate protein kinase A), RAS/ERK (extracellular regulated protein kinases) and PI3K/Akt/mTOR pathways [57,58] and in human prostate cancer cell line, arginine/GPRCA6 pathway stimulates ERK pathway to mediate its growth and progression [59].
There are additional ways that arginine may modulate cellular signal pathways. As mentioned, arginine is a precursor to NO, by the action of NO synthase (NOS) [60]. The duality of NO functions as a signaling messenger in cancer has been noted [61]. NO was regarded as an anti-tumor reagent due to its antioxidant capacity and radical scavenging property [62]. Emerging evidence, however, shows a diverse role of NO in tumorigenesis, including angiogenesis, metastasis, anti-apoptosis, anti-host immune response [63]. The increased level of NO and NOS expression has been observed in cancer patients, which is highly correlated with VEGF expression, angiogenesis [64] and metastasis [65]. Induction of NO and NOS leads to the inactivation of tumor suppressors, such as p53 [66] and pRb [67]. It has also been shown that induction of NOS and cyclooxygenase (COX-2) increased the level of NO and prostaglandins, which leads to angiogenesis in cancers [68,69,70]. Another way NO exerts its effect is through S-nitrosylation of key molecules involved in cancer induction such as EGFR and TSC2 which impacts mTOR pathway [71,72].
In addition to cell growth, NO affects immune cells in tumor microenvironment and again, in a dual mode. The dual roles of NO in carcinogenesis notwithstanding, arginine is generally considered as a tumor-promoting metabolite, very much needed for tumor growth [73].
3.2. Arginine Deprivation Induced Signals
Consistent with arginine being an activator of mTOR and PI3K pathways, numerous studies showed that arginine-deprivation suppresses mTOR and p70S6K activation with consequent inactivation of PI3K/Akt pathway [74,75] (Figure 3). In addition, arginine-deprivation activates AMPK (5’ adenosine monophosphate-activated protein kinase), due to the reduced mitochondrial OXPHOS activities and ATP production, which further suppresses mTOR activities through inactivating phosphorylation [75]. The consequences of severe mTOR suppression include aggressive autophagy and diminished synthesis of proteins, lipids and nucleotides [75,76].
Arginine deprivation also activates stress-response kinase p38, which impacts mitochondria functions, by (1) activating KAP1 (KRAB-associated protein 1) and mitochondrial fission [21] and (2) translocates nuclear TEAD4 (TEA Domain Transcription Factor 4) to cytosol, resulting in the reduced nuclear OXPHOS gene expression and mitochondrial functions [19].
Arginine deprivation in the form of nutritional stress-mediated GCN2 (general control nonderepressible 2) activation and ER stress pathway, leading to ATF4 (activating transcription factor 4) and ASNS activation and increased consumption of aspartate. Aspartate exhaustion is one reason arginine-deprivation causes the death of ASS1-low cancer cells [17].
4. Arginine and Epigenetic Regulation
Recent studies showed that arginine can act as an effective epigenetic modulator [19]. In cancer cells, arginine is a strong inducer of histone acetylation, globally enhancing the expression of metabolic, mitochondrial and DNA repair genes. Histone acetylation involves the transfer of acetyl group from acetyl-CoA to histone mediated by HATs (histone acetyltransferases) and KATs (lysine acetyltransferases), which is counteracted by deacetylation enzymes such as HDACs (histone deacetylases) and SIRTs (sirtuins). Several enzymes including ACLY (ATP citrate synthase), ACSS1 (Acyl-coA synthetase short-chain family member 1) and ACSS2 (Acyl-coA synthetase short-chain family member 2) contribute to the synthesis of acetyl-CoA. In arginine stimulated cells, the acetyl-CoA level significantly increases so do the expression levels of ACLY, ACSS2 and the majority of HATs and KATs. By contrast, the expressions of several of the HDACs and SIRTs are decreased. These results together could account for the increased global histone acetylation observed. Since mTOR is known to activate ACLY and ACSS [77,78], arginine stimulation of histone acetylation is in part attributed to the activation of mTOR. The global increase of histone acetylation however is not random but has region specificity, which is dictated by several transcription factors including TEAD4, STAT3 (signal transducer and activator of transcription 3), WT1 (Wilms’ tumor 1) and TFAM (mitochondrial transcription factor A).
Conversely, arginine deprivation leads to depletion of α-KG, which has profound effects on epigenetic regulation. As described above, arginine deprivation immediately affects mitochondrial functions [15,17,20,21] with consequent depletion of mitochondrial metabolites including α-KG. Alpha-KG is a cofactor of jumonji domain C containing histone demethylases (KDMs). As such, histone methylation generally increases during arginine deprivation. Most prominent are H3K9me3 and H3K27me3, two repressive marks contributing to gene silencing. These marks decorate genes involved in mitochondrial functions including OXPHOS, purine and pyrimidine synthesis, DNA repair, etc. (Figure 4). The consequence of such epigenetic repression is mitochondrial dysfunction, generation of reactive oxygen species (ROS), DNA damage and slow DNA repair, features which figure prominently in arginine-deprived tumor cells [15,17,20].
5. Arginine and Genome Integrity
Sufficient arginine is required for maintaining nucleotide pool and DNA repair capacity. Although arginine is not directly involved in the synthesis of nucleotide, arginine can be converted to glutamine, proline and serine, precursors of pyrimidine and arginine abundance affects genome integrity. As described above, arginine augments the transcription of genes involved in purine and pyrimidine synthesis. In addition, by virtue of activating mTOR/S6K pathway, arginine promotes the phosphorylation and oligomerization of CAD complex (carbamoyl-phosphate synthetase 2, aspartate transcarbamylase, and dihydroorotase) to enhance pyrimidine synthesis [79,80]. On the other hand, both arginine and pyrimidine syntheses require aspartate and they “compete” for this metabolite. Tumor cells often augment pyrimidine synthesis by suppressing arginine synthesis via epigenetic silencing of ASS1 [81,82], the basis of arginine-deprivation therapy. As described above, arginine deprivation also activates ATF4/ASAN which converts aspartate to asparagine causing depletion of nucleotide pool. In this scenario, the cell death caused by arginine-deprivation can be partially rescued by the addition of aspartate or nucleotide precursors [17].
In addition to arginine’s ability to epigenetically regulate the transcription of DNA repair genes, arginine affects DNA repair through the synthesis of polyamines. Polyamines interact with negatively charged DNA and plays a key role in maintaining the genome stability [83]. Polyamine depletion impairs DNA repair [84] and sensitizes cancer cells to genotoxic reagents [85,86]. Consistently, arginine deprivation which significantly reduces the polyamine levels [87] synergizes with polyamine inhibitors in the killing of cancer cells [87].
As arginine deficiency both depletes nucleotide pool and slows down DNA repair in tumor cells, it is no surprise that arginine starved tumor cells exhibit extensive DNA damages [16,17,20]. In ASS1-low pancreatic ductal adenocarcinoma, arginine deprivation exacerbates the HDAC inhibition-induced downregulation of C-terminal-binding protein-interacting protein (CtIP), a key protein for homologous recombination, leading to DNA damage and cell death [82]. In prostate and pancreatic cancer cells, arginine-starvation induced caspase-independent autophagic cell death with the appearance of nuclear DNA leakage and chromatin-autophagy (chromatophagy) [16]. This is caused by mitochondrial dysfunction and ROS production in the presence of excessive autophagy. Depletion of mitochondria or removal of ROS by NAC attenuates the DNA leakage phenotype and cell death [17]. In a study of ASS1-low melanomas, regardless of the BRAF status, arginine deprivation down-modulates FANCD2 and p-ATM, which are important initiators for DNA double strand break repair [88]. Although in this cell type, arginine deprivation alone does not induce DNA damage, combined treatment with cisplatin increases DNA double breaks, possibly due to persistent downregulation of DNA repair machinery caused by arginine deprivation. Taken together, arginine affects nucleotide synthesis/DNA repair in a complex way. The nucleotide insufficiency and down-modulated DNA repair machinery may underlie the arginine deprivation-induced DNA damage and its deficiency impairs this process and causes death of tumor cells.
6. Arginine and Immunomodulation
Arginine is a crucial immune-modulating amino acid for both innate and adaptive immunity [89,90]. It is involved in the activation of T-cell via the upregulation of T-cell receptor [91] and accelerating cell cycle progression [32]. Depletion of arginine has been used by tumor cells to generate an immunosuppressive micro-environment. Cancer cells release factors (G-CSF, GM-CSF, CCl2, etc.) to convert myeloid cells into immunosuppressive phenotypes (e.g., MDSC, myeloid-derived suppressive cells, or M2 macrophages) [92]. These cells are characterized by the expression and release of arginase [93]. Arginase produced by macrophages and MDSCs depletes extracellular arginine and hence suppression of T cell proliferation [94,95]. Supplement of arginine rewires T cell metabolism from glycolysis to OXPHOS and promotes its survival and anti-tumor ability [96]. Arginase inhibitors thus have been developed and under clinical trials as cancer therapy to counteract such immune suppression [97]. This presents an interesting duality of arginase as cancer therapy. On the one hand, arginase itself can effectively starve ASS1-low tumor cells to death. On the other, arginase inhibitor could help boost the immune defense system to suppress the growth of cancer cells. Arginine also affects innate immunity primarily via the synthesis of NO. Besides acting as a neurotransmitter and vasodilator, at high concentration, NO’s radical property enables spontaneous reactions with oxygen to form reactive nitrogen oxide species (RNOS) [98]. These RNOS reacts with DNA, protein and lipid causing DNA damage, protein dysfunction, and lipid peroxidation, which contributes to the antimicrobial and tumoricidal activities [99]. NO’s synthesis is limited by arginine availability. NO is synthesized by inducible nitric oxide synthase (iNOS) in M1 macrophages, which converts arginine into citrulline and NO (Figure 1). NO induces metabolic rewiring in M1 macrophages [100], and inhibition of iNOS induces M1 polarization into M2 type [101]. Indeed, the two arginine-metabolizing enzymes, iNOS from M1 macrophage and arginase from M2 macrophage compete for arginine to modulate innate immune responses [90]. In addition to cancer cells, several infectious bacteria and parasites also utilize the same strategy to create immunosuppressive niche by enhancing arginase expression in M2 macrophages, such as mycobacteria, Helicobacter pylori and Leishmania major [102,103,104].
Although the correlation between arginine removal by arginase and T cell suppression has been well established, how systemic removal of arginine affects the tumor microenvironment and tumor growth remains poorly understood. In a study of the effect of autophagy on tumor growth, it was found that autophagy-negative (Atg-/-) mouse released abundant arginase in the serum with consequent systemic depletion of arginine [73]. In these mice, ASS1-low syngeneic murine melanoma cells failed to grow with the infiltration of CD8 positive cells, indicating the T cell immune response is not severely affected by systemic depletion of arginine.
In another study, when human peripheral blood mononuclear cells (PBMCs) are stimulated by anti-CD3/CD28 antibodies, co-treatment with ADI-PEG20 does not block T cell activation; instead, arginine deprivation sustains the CD69+ T cells up to 72 h [105]. In the meantime, the induction of CTLA4 and PD-1 in activated T cells is blunted by arginine deprivation, and arginine deprivation also prevents Treg cells differentiation. Although arginine deprivation decreases cell proliferation of activated T cell as shown by previous studies, T cell infiltration is not compromised in syngeneic models [73,105].
Furthermore, using murine MC38 colon cell model, it was shown that ADI-PEG20 is able to induce apoptotic and immunogenic cell death with the phenotypes of cell exposure of calreticulin, the release of HMGA1 and ATP, which is enhanced by N-acetylcystein [106]. These “eat me” signals resulted in increased phagocytosis by bone marrow-derived dendritic cells. Another intriguing study shows that upon ADI-PEG20 treatment, the surrounding human T cells, which express ASS1, are still able to proliferate by taking up citrulline from the byproduct of ADI reaction and generate arginine intracellularly [107]. These studies, taken together, suggest arginine deprivation, despite its ability to suppress T-cells, are able to elicit immune responses.
More and more studies reveal that innate immunity is also crucial for anti-tumor responses [108], such as NK cell-mediated immunosurveillance [109]. In this regard, it is interesting that a recent study showed that arginine-deprivation activated innate immune response and turned “cold” tumors “hot” [20]. This is caused by cGAS-STING activation triggered by nuclear DNA leakage caused by arginine-starvation induced chromatin-autophagy. Preclinical models further showed the enhanced infiltration of immune cells in dietary arginine restricted animal model and this process is dependent on the activity of cGAS. Thus, arginine’s role in immunomodulation is complex and under certain circumstances, it may enhance the anti-tumor immunity.
Increasing evidence shows that arginine deprivation induces autonomous cancer cell death and enhances immune response. Dietary arginine-restriction offers a promising option for prevention and intervention.
Hitherto, we narrated the significance of arginine from the perspective of its role in tumorigenesis which reveals the potential of arginine deprivation as cancer therapy. In the ensuing sections, we will focus on arginine-deprivation therapy. We will first describe how arginine deprivation induces cancer cell death via different mechanisms and comprehensively summarize the current applications of arginine deprivation therapy on various cancers and discuss the associated challenges.
7. Arginine Deprivation and Cell Killing
Arginine deprivation suppresses the growth and induces cell death of ASS1-low cancer cells. The general mechanisms associated with cell killing have been studied in a number of systems (Figure 5).
7.1. Caspase-Dependent Apoptosis
This is the major mechanism associated with arginine-deprivation induced cell death, which operates in many cancer types including pleural mesothelioma cells [110], lymphoma cells [111], pancreatic cancer cells [82,112], ovarian cancers [113], sarcoma cells [114], T-lymphoblastic leukemia cells [115], liver cancer cells [116] and melanoma [117].
7.2. Caspase-Independent Apoptosis
7.3. Caspase-Independent Autophagic Death
Arginine deprivation inhibits mTOR, which is a negative regulator of autophagy. Accordingly, arginine deprivation is often accompanied by aggressive autophagy. Autophagy is a major means to regenerate arginine, which protects cells from nutrient stress. However, prolonged arginine deprivation leads to excessive and aberrant autophagy. This, coupled with ROS-induced DNA damage, leads to chromatin-autophagy or chromatophagy, where autolysosome fused with nuclear membrane and “extracts” broken chromatin out of nucleus [16], eventually leading to caspase-independent cell death. This was observed in prostate cancer cells [16], breast cancer cells [15]; hepatocellular carcinoma cells [120] and pancreatic cells [121].
In general, arginine deprivation initially induces autophagy to protect cells from starvation and at the same time, generates ROS (due to mitochondria impairment) and DNA damages which trigger apoptosis. During this early phase, an autophagy inhibitor such as chloroquine would increase cell death and enhances the drug efficacies [121]. For some cancer cells, however, autophagy persists and captures damaged broken DNA, leading to nuclear DNA leakage and cell death [20].
7.4. Necroptosis
As described above, arginine-deprivation induces autophagy which initially exerts a protective role and co-treatment with the autophagy inhibitor, chloroquine, and can facilitate the cell death [75,111,114,118]. In one study [114], it was shown such a treatment activates RIP kinase cascade, leading to necroptosis. Genetical knock-down of RIP1 or RIP3 or pharmaceutical treatment with necroptosis inhibitor, necrostatin, can protect against the co-treatment mediated cell death [114].
8. Arginine Deprivation and Cancer Therapy
The suppressed expression of ASS1, mainly due to promoter methylation [111], is making it the most prevalent metabolic deficiency of cancer and rendering cancer cells “addicted” to external arginine [122]. Given the importance of arginine in cellular processes and being the most consumed amino acids in the inner mass of tumors, it is counter-intuitive that ASS1 is epigenetically suppressed in cancer cells. As described above, one possible explanation is that ASS1 diverts aspartate to arginine synthesis from pyrimidine/purine synthesis which is much needed for tumor cells [81]. In addition, recent reports that ASS1 has other tumor suppressor functions such as inhibiting AKT activity [15,16,123]. Several studies showed that overexpression of ASS1 in tumor cells suppresses cell growth and ASS1 behaves as a tumor suppressor [17,124]. At any rate, ASS1 deficiency is a selective trait of tumor cells, making arginine deprivation a highly promising therapeutic strategy. Currently, there are two arginine deprivation therapies under clinical trials: ADI-PEG20 (pegylated arginine deiminase) and PEG-BCT-100 (pegylated recombinant human arginase1). A comparison of these two reagents is shown in Table 2.
They both are effective in depleting serum arginine, yet they are not completely innocuous. As discussed above, ADI-PEG20 generates citrulline and ammonia, and citrulline is able to affect the surrounding immune cells to generate arginine. PEG-BCT-100 produces ornithine and urea, and ornithine can fuel tumor cells to generate polyamines. The concentrations of ammonia and urea also need to be monitored during treatment. Nevertheless, these reagents have been shown to be effective and safe. The promising prospect of using arginine deprivation to starve cancer cells to death has in recent years, fueled the preclinical studies, in vitro (cell lines) and in vivo (xenografts) on a variety of ASS1-low tumors.
8.1. Preclinical Studies
Table 3 is an updated summary of publications on the application of ADI to different cancer cell lines. While the detailed cell killing mechanisms may vary among different cell types (see next section), ADI effectively inhibits the growth and survival of ASS1-low tumor cells, and in those cases tested, the susceptibility to ADI is inversely correlated with the level of ASS1. Resistance to ADI is related to recovery of ASS1 expression. These studies provide strong evidence that ASS1 expression level is a predictor for arginine-deprivation therapy. The table also listed combination treatments of chemotherapeutic agents with ADI on different cell lines. ADI synergizes with cisplatin, oxaliplatin, docetaxel, gemcitabine, TMZ to enhance the killing effects and in some cases, help overcome resistance to these drugs.
8.2. Clinical Trials
Indeed, among the AA (amino acid) deprivation therapies, arginine deprivation is the most advanced (Table 4). The first-generation arginine-deprivation therapy was based on arginase which suffers from the low enzymatic activity and sub-optimal pH [7,153]. The second-generation arginine-deprivation therapy, include human-recombinant arginase I (PEG-BCT-100) and pegylated-ADI (ADI-PEG20) showed significant promises [126,154,155]. Considering the adverse effects of arginase I on immunity described above, the arginase inhibitor (CB-1158) is being evaluated in vivo [97] and clinical trials (Table 4). We will update the clinical trials of both ADI-PEG20 and PEG-BCT-100 in Table 4. There are more than 20 clinical trials of ADI-PEG20 on more than 12 types of cancer, which have been completed or are ongoing. In most cases, phase I/II were completed with an excellent safety profile (Table 4). Co-targeting with other anti-cancer therapy is being developed. Trials on hepatocellular carcinoma and mesotheliomas are the most advanced and have reached phase III. The phase III mesothelioma trial involving pemetrexed, cisplatin and ADI-PEG20 is ongoing and actively recruiting. The phase III hepatocellular carcinoma trial with ADI-PEG20 as monotherapy did not reach the intended goal [156]. One of possibilities is the treatment induces anti-ADI-PEG20 antibody, which neutralized the ADI-PEG20 [157]. Another possibility is due to the re-expression of ASS1 with prior treatment, such as sorafenib, which may reduce the efficacy of ADI-PEG20 [156]. The possible mechanism of drug resistance is discussed in the following section. Arginase clinical trials are mostly in phase I and II. In Table 4, we also included clinical trials for arginase inhibitors, which are intended to enhance the T-cell functions and anti-tumor activities.
9. Arginine Deprivation and Therapy Resistance
One of the challenges in ADI-based therapy is the development of intrinsic resistance [170]. A common mechanism is the restoration or upregulation of ASS1 expression to provide much-needed arginine in starved cancer cells (Figure 6). For tumors (e.g., lymphomas, bladder, mesothelioma, prostate cancer) where ASS1 was silenced by methylation, demethylation of ASS1 promoter is the solution [87,111], although what triggers demethylation is not clear. For tumors of which the ASS1 promoter is not methylated (e.g., melanoma and sarcoma), the activation of c-Myc oncogene which drives the expression of ASS1 appears to be the underlying cause [22,23]. In the latter case, Gas6/Axl activation by arginine deprivation-induced ROS triggers PI3K/Akt/GSK3B activation, leading to the upregulation of c-Myc oncogene [152]. The studies by the lab of MT Kuo and his collaborators further demonstrated how c-Myc could replace negative regulator HIF-1a, which competitively occupied the same E-box of the ASS1 promoter to restore ASS1 expression [133]. On the other hand, c-Myc upregulation is not universally seen in all ADI-resistant cancer cells (e.g., prostate and breast cancer) and c-Myc overexpression does not always confer ADI resistance [18]. Interestingly, in lung cancer, Myc overexpression actually confers sensitivity, rather than resistance, to ADI treatment [139]. These studies suggest that while restoration of ASS1 expression seems to be necessary to provide the much-needed arginine during therapy, the contributing mechanisms vary and there are other context-dependent cellular factors associated with resistance. As mentioned above, ASS1 has been shown to repress Akt activities [123] and behaves as a tumor suppressor [15,124]. Other pathways are needed to offset its tumor suppressive properties. In melanoma, ADI-resistant cells, enhanced AXL and EPH2 tyrosine kinase activities and the activation of down-stream PI3K/AKT signaling are observed [152]. In breast and prostate cancer ADI-resistant cells, Chu et al., [18] using unbiased CRISPR/cas9 loss of function screening uncovered aberrant activation of TREM1/CCL2 is necessary to confer full resistance. TREM1, a receptor normally expressed in myeloid cells, activates tyrosine kinase Syk which turns on PI3K/Akt and STAT3/NFKB, leading to the activation of CCL2. CCL2, a chemokine known to confer drug resistance to a variety of cancer cells, further enhances the activation PI3K/Akt and STAT3/NFkB pathways [18]. As direct targeting ASS1 may affect the metabolism of normal cells, these resistant factors and their associated pathways offer additional opportunities as targets to overcome ADI resistance.
Khadeir et al. [170] further considered the factors which may compromise therapy efficacies in vivo, and suggests that in addition to ASS1 upregulation, protective autophagy which regenerates arginine, development of antibodies against pegylated ADI and the uptake of arginine released from stromal cells. Measures to counteract these resistant factors are being actively pursued to improve arginine-deprivation therapy.
10. Conclusions
As summarized in this article, arginine, in addition to being a building block of protein, can be a signaling metabolite, a transcriptome reprogrammer and a therapeutic target. Recent insights that arginine is a mitochondria and genome protector are exciting, with clinical implications. As a semi-essential amino acid, arginine deprivation based on biologicals which metabolize arginine has been a staple of starvation therapies for years. While the safety profiles for both arginine depletion remedies are generally excellent, as a monotherapy agent, it has not reached the intended potency. Combinations with other targeted or chemotherapies as well as immune checkpoint inhibitors are being actively pursued. Since the cell-killing mechanisms associated with starvation therapies (e.g., autophagic death, caspase-independent apoptosis, necroptosis, etc.) could be quite different from those associated with genotoxic agents (e.g., caspase-dependent apoptosis, mitotic catastrophe), arginine-deprivation should synergize well with other cancer therapies and potentially overcome the resistance of the latter. In addition, an arginine restriction diet also deserves serious attention as a complementary tool for cancer therapy. Unlike using metabolizing enzymes as depletion agents, dietary restriction does not have the complication of metabolic products. Dietary restriction has been widely applied in various metabolic diseases such as obesity, diabetes, rheumatoid arthritis, hypercholesterolemia, etc., and it has gained significant momentum for cancer therapies. Recent work showed that arginine-free diet effectively retards the growth of prostate and breast cancer xenografts in the mouse models. Previous studies of arginine and precursor (glutamate, proline and aspartate)-free diet for 4 weeks showed no side effect in healthy adults, which sheds a light on arginine-restriction diet for cancer therapy. While it may be impractical to follow a strict arginine-free diet, arginine-light diet (such as vegetables, fruits and milk products) could be an alternative choice. It is hoped that an arginine-restricted diet could significantly slow down the growth of ASS1-low tumors and offers benefit in lowering the toxicity of combined chemo- or radio-therapies. Finally yet importantly, as we discussed in the beginning of this review, different tumors utilize different strategies to cope with the nutrient demand or stress, exploring the metabolic heterogeneity of various cancer types definitely is required for personalized medicine [171].
Author Contributions
C.-L.C., S.-C.H., D.K.A., Y.Y. and H.-J.K. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants MOST 105-2314-B-400-019-MY3, MOST 107-2320-B-038-055-MY3, MOST 105-2320-B-038-071-MY3, MOST 108-2320-B-038-011-MY3 (to H.-J.K), MOST 106-2321-B-400-012-MY3 (to C.-L.C.), as well as “TMU Research Center of Cancer Translational Medicine” from The Featured Areas Research Center Program within the framework of the Higher Education Sprout Project by the Ministry of Education (MOE) in Taiwan to Y.Y. and H.-J.K.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boroughs, L.K.; DeBerardinis, R.J. Metabolic pathways promoting cancer cell survival and growth. Nat. Cell Biol. 2015, 17, 351–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Bermudez, J.; Williams, R.T.; Guarecuco, R.; Birsoy, K. Targeting extracellular nutrient dependencies of cancer cells. Mol. Metab. 2020, 33, 67–82. [Google Scholar] [CrossRef]
- Zou, S.; Wang, X.; Liu, P.; Ke, C.; Xu, S. Arginine metabolism and deprivation in cancer therapy. Biomed. Pharmacother. 2019, 118, 109210. [Google Scholar] [CrossRef] [PubMed]
- Riess, C.; Shokraie, F.; Classen, C.F.; Kreikemeyer, B.; Fiedler, T.; Junghanss, C.; Maletzki, C. Arginine-depleting enzymes–An increasingly recognized treatment strategy for therapy-refractory malignancies. Cell. Physiol. Biochem. 2018, 51, 854–870. [Google Scholar] [CrossRef]
- Rogers, L.C.; Van Tine, B.A. Innate and adaptive resistance mechanisms to arginine deprivation therapies in sarcoma and other cancers. Cancer Drug Resist. 2019, 2, 516–526. [Google Scholar] [CrossRef] [Green Version]
- Delage, B.; Fennell, D.A.; Nicholson, L.; McNeish, I.; Lemoine, N.R.; Crook, T.; Szlosarek, P.W. Arginine deprivation and argininosuccinate synthetase expression in the treatment of cancer. Int. J. Cancer 2010, 126, 2762–2772. [Google Scholar] [CrossRef] [PubMed]
- Fultang, L.; Vardon, A.; De Santo, C.; Mussai, F. Molecular basis and current strategies of therapeutic arginine depletion for cancer. Int. J. Cancer 2016, 139, 501–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, M.D.; Bhaumik, J.; Babykutty, S.; Banerjee, U.C.; Fukumura, D. Arginine dependence of tumor cells: Targeting a chink in cancer’s armor. Oncogene 2016, 35, 4957–4972. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Shaibe, E.; Metzer, E.; Halpern, Y.S. Control of utilization of L-arginine, L-ornithine, agmatine, and putrescine as nitrogen sources in Escherichia coli K-12. J. Bacteriol. 1985, 163, 938–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husson, A.; Brasse-Lagnel, C.; Fairand, A.; Renouf, S.; Lavoinne, A. Argininosuccinate synthetase from the urea cycle to the citrulline-NO cycle. Eur. J. Biochem. 2003, 270, 1887–1899. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.P.; Grisolia, S. The intermediate role of carbamyl-L-glutamic acid in citrulline synthesis. J. Biol. Chem. 1948, 174, 389. [Google Scholar] [CrossRef]
- Hamilton, P.B. Proline: Synthesis from ornithine, citrulline, or arginine. J. Biol. Chem. 1952, 198, 587–597. [Google Scholar] [CrossRef]
- Levenberg, B. Role of L-glutamine as donor of carbamyl nitrogen for the enzymatic synthesis of citruline in Agaricus bisporus. J. Biol. Chem. 1962, 237, 2590–2598. [Google Scholar] [CrossRef]
- Qiu, F.; Chen, Y.R.; Liu, X.; Chu, C.Y.; Shen, L.J.; Xu, J.; Gaur, S.; Forman, H.J.; Zhang, H.; Zheng, S.; et al. Arginine starvation impairs mitochondrial respiratory function in ASS1-deficient breast cancer cells. Sci. Signal. 2014, 7, ra31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Changou, C.A.; Chen, Y.R.; Xing, L.; Yen, Y.; Chuang, F.Y.; Cheng, R.H.; Bold, R.J.; Ann, D.K.; Kung, H.J. Arginine starvation-associated atypical cellular death involves mitochondrial dysfunction, nuclear DNA leakage, and chromatin autophagy. Proc. Natl. Acad. Sci. USA 2014, 111, 14147–14152. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.T.; Qi, Y.; Wang, Y.C.; Chi, K.K.; Chung, Y.; Ouyang, C.; Chen, Y.R.; Oh, M.E.; Sheng, X.; Tang, Y.; et al. Arginine starvation kills tumor cells through aspartate exhaustion and mitochondrial dysfunction. Commun. Biol. 2018, 1, 178. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.Y.; Lee, Y.C.; Hsieh, C.H.; Yeh, C.T.; Chao, T.Y.; Chen, P.H.; Lin, I.H.; Hsieh, T.H.; Shih, J.W.; Cheng, C.H.; et al. Genome-wide CRISPR/Cas9 knockout screening uncovers a novel inflammatory pathway critical for resistance to arginine-deprivation therapy. Theranostics 2021, 11, 3624–3641. [Google Scholar] [CrossRef]
- Chen, C.L.; Hsu, S.C.; Chung, T.Y.; Chu, C.Y.; Wang, H.J.; Hsiao, P.W.; Yeh, S.D.; Ann, D.K.; Yen, Y.; Kung, H.J. Arginine is an epigenetic regulator targeting TEAD4 to modulate OXPHOS in prostate cancer cells. Nat. Commun. 2021, 12, 2398. [Google Scholar] [CrossRef]
- Hsu, S.-C.; Chen, C.-L.; Cheng, M.-L.; Chu, C.-Y.; Changou, C.A.; Yu, Y.-L.; Yeh, S.-D.; Kuo, T.-C.; Kuo, C.-C.; Chuu, C.-P.; et al. Arginine starvation elicits chromatin leakage and cGAS-STING activation via epigenetic silencing of metabolic and DNA-repair genes. Theranostics 2021, 11, 7527–7545. [Google Scholar] [CrossRef]
- Cheng, C.T.; Kuo, C.Y.; Ouyang, C.; Li, C.F.; Chung, Y.; Chan, D.C.; Kung, H.J.; Ann, D.K. Metabolic Stress-Induced Phosphorylation of KAP1 Ser473 Blocks Mitochondrial Fusion in Breast Cancer Cells. Cancer Res. 2016, 76, 5006–5018. [Google Scholar] [CrossRef] [Green Version]
- Kremer, J.C.; Prudner, B.C.; Lange, S.E.S.; Bean, G.R.; Schultze, M.B.; Brashears, C.B.; Radyk, M.D.; Redlich, N.; Tzeng, S.C.; Kami, K.; et al. Arginine Deprivation Inhibits the Warburg Effect and Upregulates Glutamine Anaplerosis and Serine Biosynthesis in ASS1-Deficient Cancers. Cell Rep. 2017, 18, 991–1004. [Google Scholar] [CrossRef] [Green Version]
- Long, Y.; Tsai, W.B.; Wangpaichitr, M.; Tsukamoto, T.; Savaraj, N.; Feun, L.G.; Kuo, M.T. Arginine deiminase resistance in melanoma cells is associated with metabolic reprogramming, glucose dependence, and glutamine addiction. Mol. Cancer Ther. 2013, 12, 2581–2590. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.B.; Aiba, I.; Long, Y.; Lin, H.K.; Feun, L.; Savaraj, N.; Kuo, M.T. Activation of Ras/PI3K/ERK pathway induces c-Myc stabilization to upregulate argininosuccinate synthetase, leading to arginine deiminase resistance in melanoma cells. Cancer Res. 2012, 72, 2622–2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brashears, C.B.; Barlin, M.; Ehrhardt, W.R.; Rathore, R.; Schultze, M.; Tzeng, S.C.; Van Tine, B.A.; Held, J.M. Systems level profiling of arginine starvation reveals MYC and ERK adaptive metabolic reprogramming. Cell Death Dis. 2020, 11, 662. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Tchernyshyov, I.; Chang, T.C.; Lee, Y.S.; Kita, K.; Ochi, T.; Zeller, K.I.; De Marzo, A.M.; Van Eyk, J.E.; Mendell, J.T.; et al. c-Myc suppression of miR-23a/b enhances mitochondrial glutaminase expression and glutamine metabolism. Nature 2009, 458, 762–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banjarnahor, S.; Rodionov, R.N.; Konig, J.; Maas, R. Transport of L-Arginine Related Cardiovascular Risk Markers. J. Clin. Med. 2020, 9, 3975. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zou, W. Amino Acids and Their Transporters in T Cell Immunity and Cancer Therapy. Mol. Cell 2020, 80, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Tsun, Z.Y.; Wolfson, R.L.; Shen, K.; Wyant, G.A.; Plovanich, M.E.; Yuan, E.D.; Jones, T.D.; Chantranupong, L.; Comb, W.; et al. Metabolism. Lysosomal amino acid transporter SLC38A9 signals arginine sufficiency to mTORC1. Science 2015, 347, 188–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.C.; Chen, Y.R.; Kensicki, E.; Li, A.Y.; Kong, M.; Li, Y.; Mohney, R.P.; Shen, H.M.; Stiles, B.; Mizushima, N.; et al. Autophagy: Resetting glutamine-dependent metabolism and oxygen consumption. Autophagy 2012, 8, 1477–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishton, R.J.; Sukumar, M.; Restifo, N.P. Arginine Arms T Cells to Thrive and Survive. Cell Metab. 2016, 24, 647–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, P.C.; Quiceno, D.G.; Ochoa, A.C. L-arginine availability regulates T-lymphocyte cell-cycle progression. Blood 2007, 109, 1568–1573. [Google Scholar] [CrossRef] [Green Version]
- Dai, R.; Peng, F.; Xiao, X.; Gong, X.; Jiang, Y.; Zhang, M.; Tian, Y.; Xu, Y.; Ma, J.; Li, M.; et al. Hepatitis B virus X protein-induced upregulation of CAT-1 stimulates proliferation and inhibits apoptosis in hepatocellular carcinoma cells. Oncotarget 2017, 8, 60962–60974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Wang, W.; Wang, J.; Yang, C.; Mao, H.; Fu, X.; Wu, Y.; Cai, J.; Han, J.; Xu, Z.; et al. Overexpression of arginine transporter CAT-1 is associated with accumulation of L-arginine and cell growth in human colorectal cancer tissue. PLoS ONE 2013, 8, e73866. [Google Scholar] [CrossRef] [PubMed]
- Abdelmagid, S.A.; Rickard, J.A.; McDonald, W.J.; Thomas, L.N.; Too, C.K. CAT-1-mediated arginine uptake and regulation of nitric oxide synthases for the survival of human breast cancer cell lines. J. Cell Biochem. 2011, 112, 1084–1092. [Google Scholar] [CrossRef]
- Werner, A.; Pieh, D.; Echchannaoui, H.; Rupp, J.; Rajalingam, K.; Theobald, M.; Closs, E.I.; Munder, M. Cationic Amino Acid Transporter-1-Mediated Arginine Uptake Is Essential for Chronic Lymphocytic Leukemia Cell Proliferation and Viability. Front. Oncol. 2019, 9, 1268. [Google Scholar] [CrossRef] [PubMed]
- Werner, A.; Amann, E.; Schnitzius, V.; Habermeier, A.; Luckner-Minden, C.; Leuchtner, N.; Rupp, J.; Closs, E.I.; Munder, M. Induced arginine transport via cationic amino acid transporter-1 is necessary for human T-cell proliferation. Eur. J. Immunol. 2016, 46, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Miyauchi, S.; Martindale, R.G.; Herdman, A.V.; Podolsky, R.; Miyake, K.; Mager, S.; Prasad, P.D.; Ganapathy, M.E.; Ganapathy, V. Upregulation of the amino acid transporter ATB0,+ (SLC6A14) in colorectal cancer and metastasis in humans. Biochim. Biophys. Acta 2005, 1741, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Prasad, P.D.; Ghamande, S.; Moore-Martin, P.; Herdman, A.V.; Martindale, R.G.; Podolsky, R.; Mager, S.; Ganapathy, M.E.; Ganapathy, V. Up-regulation of the amino acid transporter ATB(0,+) (SLC6A14) in carcinoma of the cervix. Gynecol. Oncol. 2006, 100, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.L.; Corchete, L.; Teodosio, C.; Sarasquete, M.E.; del Mar Abad, M.; Iglesias, M.; Esteban, C.; Sayagues, J.M.; Orfao, A.; Munoz-Bellvis, L. Identification and characterization of the gene expression profiles for protein coding and non-coding RNAs of pancreatic ductal adenocarcinomas. Oncotarget 2015, 6, 19070–19086. [Google Scholar] [CrossRef] [Green Version]
- Babu, E.; Bhutia, Y.D.; Ramachandran, S.; Gnanaprakasam, J.P.; Prasad, P.D.; Thangaraju, M.; Ganapathy, V. Deletion of the amino acid transporter Slc6a14 suppresses tumour growth in spontaneous mouse models of breast cancer. Biochem. J. 2015, 469, 17–23. [Google Scholar] [CrossRef]
- Mossner, J.; Hammermann, R.; Racke, K. Concomitant down-regulation of L-arginine transport and nitric oxide (NO) synthesis in rat alveolar macrophages by the polyamine spermine. Pulm. Pharmacol. Ther. 2001, 14, 297–305. [Google Scholar] [CrossRef]
- Lowman, X.H.; Hanse, E.A.; Yang, Y.; Ishak Gabra, M.B.; Tran, T.Q.; Li, H.; Kong, M. p53 Promotes Cancer Cell Adaptation to Glutamine Deprivation by Upregulating Slc7a3 to Increase Arginine Uptake. Cell Rep. 2019, 26, 3051–3060.e4. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Cao, Y.; Wang, Y.; Li, W.; Liu, X.; Lv, Y.; Li, X.; Mi, J. Cysteine transporter SLC3A1 promotes breast cancer tumorigenesis. Theranostics 2017, 7, 1036–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Han, T.; Bian, Y.; Tong, H.; Wen, X.; Li, Y.; Wan, X. Knockdown of SLCO4C1 inhibits cell proliferation and metastasis in endometrial cancer through inactivating the PI3K/Akt signaling pathway. Oncol. Rep. 2020, 43, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.C.; Qian, Y.; Li, X.; Xu, J.; Kang, W.; Tong, J.H.; To, K.F.; Jin, Y.; Li, W.; Chen, H.; et al. SLC25A22 Promotes Proliferation and Survival of Colorectal Cancer Cells with KRAS Mutations and Xenograft Tumor Progression in Mice via Intracellular Synthesis of Aspartate. Gastroenterology 2016, 151, 945–960.e6. [Google Scholar] [CrossRef] [Green Version]
- Ban, H.; Shigemitsu, K.; Yamatsuji, T.; Haisa, M.; Nakajo, T.; Takaoka, M.; Nobuhisa, T.; Gunduz, M.; Tanaka, N.; Naomoto, Y. Arginine and Leucine regulate p70 S6 kinase and 4E-BP1 in intestinal epithelial cells. Int. J. Mol. Med. 2004, 13, 537–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jewell, J.L.; Guan, K.L. Nutrient signaling to mTOR and cell growth. Trends Biochem. Sci. 2013, 38, 233–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jewell, J.L.; Russell, R.C.; Guan, K.L. Amino acid signalling upstream of mTOR. Nat. Rev. Mol. Cell Biol. 2013, 14, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahbazian, D.; Roux, P.P.; Mieulet, V.; Cohen, M.S.; Raught, B.; Taunton, J.; Hershey, J.W.; Blenis, J.; Pende, M.; Sonenberg, N. The mTOR/PI3K and MAPK pathways converge on eIF4B to control its phosphorylation and activity. EMBO J. 2006, 25, 2781–2791. [Google Scholar] [CrossRef]
- Carroll, B.; Maetzel, D.; Maddocks, O.D.; Otten, G.; Ratcliff, M.; Smith, G.R.; Dunlop, E.A.; Passos, J.F.; Davies, O.R.; Jaenisch, R.; et al. Control of TSC2-Rheb signaling axis by arginine regulates mTORC1 activity. eLife 2016, 5, e11058. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Hall, M.N. Nutrient sensing and TOR signaling in yeast and mammals. EMBO J. 2017, 36, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Tan, B.; Yin, Y.; Gao, H.; Li, X.; Jaeger, L.A.; Bazer, F.W.; Wu, G. L-Arginine stimulates the mTOR signaling pathway and protein synthesis in porcine trophectoderm cells. J. Nutr. Biochem. 2012, 23, 1178–1183. [Google Scholar] [CrossRef] [PubMed]
- Chantranupong, L.; Scaria, S.M.; Saxton, R.A.; Gygi, M.P.; Shen, K.; Wyant, G.A.; Wang, T.; Harper, J.W.; Gygi, S.P.; Sabatini, D.M. The CASTOR Proteins Are Arginine Sensors for the mTORC1 Pathway. Cell 2016, 165, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, B.; Wellendorph, P.; Brauner-Osborne, H. Known regulators of nitric oxide synthase and arginase are agonists at the human G-protein-coupled receptor GPRC6A. Br. J. Pharm. 2006, 147, 855–863. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, T.; Kanazawa, S.; Ichibori, R.; Tanigawa, T.; Magome, T.; Shingaki, K.; Miyata, S.; Tohyama, M.; Hosokawa, K. L-arginine stimulates fibroblast proliferation through the GPRC6A-ERK1/2 and PI3K/Akt pathway. PLoS ONE 2014, 9, e92168. [Google Scholar]
- Pi, M.; Nishimoto, S.K.; Quarles, L.D. GPRC6A: Jack of all metabolism (or master of none). Mol. Metab. 2017, 6, 185–193. [Google Scholar] [CrossRef]
- Pi, M.; Quarles, L.D. GPRC6A regulates prostate cancer progression. Prostate 2012, 72, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Palmer, R.M.; Ashton, D.S.; Moncada, S. Vascular endothelial cells synthesize nitric oxide from L-arginine. Nature 1988, 333, 664–666. [Google Scholar] [CrossRef]
- Keshet, R.; Erez, A. Arginine and the metabolic regulation of nitric oxide synthesis in cancer. Dis. Model. Mech. 2018, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Wink, D.A.; Miranda, K.M.; Espey, M.G.; Pluta, R.M.; Hewett, S.J.; Colton, C.; Vitek, M.; Feelisch, M.; Grisham, M.B. Mechanisms of the antioxidant effects of nitric oxide. Antioxid. Redox Signal. 2001, 3, 203–213. [Google Scholar] [CrossRef]
- Lala, P.K.; Chakraborty, C. Role of nitric oxide in carcinogenesis and tumour progression. Lancet Oncol. 2001, 2, 149–156. [Google Scholar] [CrossRef]
- Isenberg, J.S.; Martin-Manso, G.; Maxhimer, J.B.; Roberts, D.D. Regulation of nitric oxide signalling by thrombospondin 1: Implications for anti-angiogenic therapies. Nat. Rev. Cancer 2009, 9, 182–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukumura, D.; Kashiwagi, S.; Jain, R.K. The role of nitric oxide in tumour progression. Nat. Rev. Cancer 2006, 6, 521–534. [Google Scholar] [CrossRef]
- Ambs, S.; Ogunfusika, M.O.; Merriam, W.G.; Bennett, W.P.; Billiar, T.R.; Harris, C.C. Up-regulation of inducible nitric oxide synthase expression in cancer-prone p53 knockout mice. Proc. Natl. Acad. Sci. USA 1998, 95, 8823–8828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radisavljevic, Z. Inactivated tumor suppressor Rb by nitric oxide promotes mitosis in human breast cancer cells. J. Cell Biochem. 2004, 92, 1–5. [Google Scholar] [CrossRef]
- Chen, C.N.; Hsieh, F.J.; Cheng, Y.M.; Chang, K.J.; Lee, P.H. Expression of inducible nitric oxide synthase and cyclooxygenase-2 in angiogenesis and clinical outcome of human gastric cancer. J. Surg. Oncol. 2006, 94, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Lee, S.G.; Song, S.H.; Heo, D.S.; Park, B.J.; Lee, D.W.; Kim, K.H.; Sung, M.W. The effect of nitric oxide on cyclooxygenase-2 (COX-2) overexpression in head and neck cancer cell lines. Int. J. Cancer 2003, 107, 729–738. [Google Scholar] [CrossRef]
- Rahman, M.A.; Dhar, D.K.; Yamaguchi, E.; Maruyama, S.; Sato, T.; Hayashi, H.; Ono, T.; Yamanoi, A.; Kohno, H.; Nagasue, N. Coexpression of inducible nitric oxide synthase and COX-2 in hepatocellular carcinoma and surrounding liver: Possible involvement of COX-2 in the angiogenesis of hepatitis C virus-positive cases. Clin. Cancer Res. 2001, 7, 1325–1332. [Google Scholar] [PubMed]
- Garrido, P.; Shalaby, A.; Walsh, E.M.; Keane, N.; Webber, M.; Keane, M.M.; Sullivan, F.J.; Kerin, M.J.; Callagy, G.; Ryan, A.E.; et al. Impact of inducible nitric oxide synthase (iNOS) expression on triple negative breast cancer outcome and activation of EGFR and ERK signaling pathways. Oncotarget 2017, 8, 80568–80588. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Rivera, E.; Jayaraman, P.; Parikh, F.; Davies, M.A.; Ekmekcioglu, S.; Izadmehr, S.; Milton, D.R.; Chipuk, J.E.; Grimm, E.A.; Estrada, Y.; et al. Inducible nitric oxide synthase drives mTOR pathway activation and proliferation of human melanoma by reversible nitrosylation of TSC2. Cancer Res. 2014, 74, 1067–1078. [Google Scholar] [CrossRef] [Green Version]
- Poillet-Perez, L.; Xie, X.; Zhan, L.; Yang, Y.; Sharp, D.W.; Hu, Z.S.; Su, X.; Maganti, A.; Jiang, C.; Lu, W.; et al. Autophagy maintains tumour growth through circulating arginine. Nature 2018, 563, 569–573. [Google Scholar] [CrossRef]
- Wang, H.; Li, Q.F.; Chow, H.Y.; Choi, S.C.; Leung, Y.C. Arginine deprivation inhibits pancreatic cancer cell migration, invasion and EMT via the down regulation of Snail, Slug, Twist, and MMP1/9. J. Physiol. Biochem. 2020, 76, 73–83. [Google Scholar] [CrossRef]
- Kim, R.H.; Coates, J.M.; Bowles, T.L.; McNerney, G.P.; Sutcliffe, J.; Jung, J.U.; Gandour-Edwards, R.; Chuang, F.Y.; Bold, R.J.; Kung, H.J. Arginine deiminase as a novel therapy for prostate cancer induces autophagy and caspase-independent apoptosis. Cancer Res. 2009, 69, 700–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, P.P.; Kang, S.A.; Rameseder, J.; Zhang, Y.; Ottina, K.A.; Lim, D.; Peterson, T.R.; Choi, Y.; Gray, N.S.; Yaffe, M.B.; et al. The mTOR-regulated phosphoproteome reveals a mechanism of mTORC1-mediated inhibition of growth factor signaling. Science 2011, 332, 1317–1322. [Google Scholar] [CrossRef] [Green Version]
- Porstmann, T.; Santos, C.R.; Griffiths, B.; Cully, M.; Wu, M.; Leevers, S.; Griffiths, J.R.; Chung, Y.L.; Schulze, A. SREBP activity is regulated by mTORC1 and contributes to Akt-dependent cell growth. Cell Metab. 2008, 8, 224–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Chen, X.; Zang, A.; Li, T.; Hu, Y.; Ma, S.; Lu, M.; Yin, H.; Wang, H.; Zhang, X.; et al. TSC1/mTOR-controlled metabolic-epigenetic cross talk underpins DC control of CD8+ T-cell homeostasis. PLoS Biol. 2019, 17, e3000420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Sahra, I.; Howell, J.J.; Asara, J.M.; Manning, B.D. Stimulation of de novo pyrimidine synthesis by growth signaling through mTOR and S6K1. Science 2013, 339, 1323–1328. [Google Scholar] [CrossRef] [Green Version]
- Robitaille, A.M.; Christen, S.; Shimobayashi, M.; Cornu, M.; Fava, L.L.; Moes, S.; Prescianotto-Baschong, C.; Sauer, U.; Jenoe, P.; Hall, M.N. Quantitative phosphoproteomics reveal mTORC1 activates de novo pyrimidine synthesis. Science 2013, 339, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, S.; Adler, L.; Yizhak, K.; Sarver, A.; Silberman, A.; Agron, S.; Stettner, N.; Sun, Q.; Brandis, A.; Helbling, D.; et al. Diversion of aspartate in ASS1-deficient tumours fosters de novo pyrimidine synthesis. Nature 2015, 527, 379–383. [Google Scholar] [CrossRef]
- Kim, S.S.; Xu, S.; Cui, J.; Poddar, S.; Le, T.M.; Hayrapetyan, H.; Li, L.; Wu, N.; Moore, A.M.; Zhou, L.; et al. Histone deacetylase inhibition is synthetically lethal with arginine deprivation in pancreatic cancers with low argininosuccinate synthetase 1 expression. Theranostics 2020, 10, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.Y.; Su, G.C.; Huang, W.Y.; Ko, M.Y.; Yeh, H.Y.; Chang, G.D.; Lin, S.J.; Chi, P. Promotion of homology-directed DNA repair by polyamines. Nat. Commun. 2019, 10, 65. [Google Scholar] [CrossRef]
- Snyder, R.D.; Lachmann, P.J. Hyperthermia, polyamine depletion, and inhibition of X-ray-induced DNA strand break repair. Radiat. Res. 1989, 120, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Snyder, R.D.; Sunkara, P.S. Effect of polyamine depletion on DNA damage and repair following UV irradiation of HeLa cells. Photochem. Photobiol. 1990, 52, 525–532. [Google Scholar] [CrossRef]
- Locke, M.; Ghazaly, E.; Freitas, M.O.; Mitsinga, M.; Lattanzio, L.; Lo Nigro, C.; Nagano, A.; Wang, J.; Chelala, C.; Szlosarek, P.; et al. Inhibition of the Polyamine Synthesis Pathway Is Synthetically Lethal with Loss of Argininosuccinate Synthase 1. Cell Rep. 2016, 16, 1604–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scully, R.; Panday, A.; Elango, R.; Willis, N.A. DNA double-strand break repair-pathway choice in somatic mammalian cells. Nat. Rev. Mol. Cell Biol. 2019, 20, 698–714. [Google Scholar] [CrossRef]
- Bronte, V.; Zanovello, P. Regulation of immune responses by L-arginine metabolism. Nat. Rev. Immunol. 2005, 5, 641–654. [Google Scholar] [CrossRef]
- Morris, S.M., Jr. Arginine: Master and commander in innate immune responses. Sci. Signal. 2010, 3, pe27. [Google Scholar] [CrossRef]
- Rodriguez, P.C.; Zea, A.H.; Culotta, K.S.; Zabaleta, J.; Ochoa, J.B.; Ochoa, A.C. Regulation of T cell receptor CD3zeta chain expression by L-arginine. J. Biol. Chem. 2002, 277, 21123–21129. [Google Scholar] [CrossRef] [Green Version]
- Rath, M.; Muller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via Arginase or Nitric Oxide Synthase: Two Competing Arginine Pathways in Macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grzywa, T.M.; Sosnowska, A.; Matryba, P.; Rydzynska, Z.; Jasinski, M.; Nowis, D.; Golab, J. Myeloid Cell-Derived Arginase in Cancer Immune Response. Front. Immunol. 2020, 11, 938. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.C.; Zea, A.H.; DeSalvo, J.; Culotta, K.S.; Zabaleta, J.; Quiceno, D.G.; Ochoa, J.B.; Ochoa, A.C. L-arginine consumption by macrophages modulates the expression of CD3 zeta chain in T lymphocytes. J. Immunol. 2003, 171, 1232–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serafini, P.; Mgebroff, S.; Noonan, K.; Borrello, I. Myeloid-derived suppressor cells promote cross-tolerance in B-cell lymphoma by expanding regulatory T cells. Cancer Res. 2008, 68, 5439–5449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiger, R.; Rieckmann, J.C.; Wolf, T.; Basso, C.; Feng, Y.; Fuhrer, T.; Kogadeeva, M.; Picotti, P.; Meissner, F.; Mann, M.; et al. L-Arginine Modulates T Cell Metabolism and Enhances Survival and Anti-tumor Activity. Cell 2016, 167, 829–842.e13. [Google Scholar] [CrossRef] [Green Version]
- Steggerda, S.M.; Bennett, M.K.; Chen, J.; Emberley, E.; Huang, T.; Janes, J.R.; Li, W.; MacKinnon, A.L.; Makkouk, A.; Marguier, G.; et al. Inhibition of arginase by CB-1158 blocks myeloid cell-mediated immune suppression in the tumor microenvironment. J. Immunother. Cancer 2017, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Espey, M.G.; Miranda, K.M.; Thomas, D.D.; Xavier, S.; Citrin, D.; Vitek, M.P.; Wink, D.A. A chemical perspective on the interplay between NO, reactive oxygen species, and reactive nitrogen oxide species. Ann. N. Y. Acad. Sci. 2002, 962, 195–206. [Google Scholar] [CrossRef]
- Wink, D.A.; Mitchell, J.B. Chemical biology of nitric oxide: Insights into regulatory, cytotoxic, and cytoprotective mechanisms of nitric oxide. Free Radic. Biol. Med. 1998, 25, 434–456. [Google Scholar] [CrossRef]
- Palmieri, E.M.; Gonzalez-Cotto, M.; Baseler, W.A.; Davies, L.C.; Ghesquiere, B.; Maio, N.; Rice, C.M.; Rouault, T.A.; Cassel, T.; Higashi, R.M.; et al. Nitric oxide orchestrates metabolic rewiring in M1 macrophages by targeting aconitase 2 and pyruvate dehydrogenase. Nat. Commun. 2020, 11, 698. [Google Scholar] [CrossRef]
- Van den Bossche, J.; Baardman, J.; Otto, N.A.; van der Velden, S.; Neele, A.E.; van den Berg, S.M.; Luque-Martin, R.; Chen, H.J.; Boshuizen, M.C.; Ahmed, M.; et al. Mitochondrial Dysfunction Prevents Repolarization of Inflammatory Macrophages. Cell Rep. 2016, 17, 684–696. [Google Scholar] [CrossRef] [Green Version]
- Qualls, J.E.; Neale, G.; Smith, A.M.; Koo, M.S.; DeFreitas, A.A.; Zhang, H.; Kaplan, G.; Watowich, S.S.; Murray, P.J. Arginine usage in mycobacteria-infected macrophages depends on autocrine-paracrine cytokine signaling. Sci. Signal. 2010, 3, ra62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, N.D.; Asim, M.; Barry, D.P.; de Sablet, T.; Singh, K.; Piazuelo, M.B.; Gobert, A.P.; Chaturvedi, R.; Wilson, K.T. Immune evasion by Helicobacter pylori is mediated by induction of macrophage arginase II. J. Immunol. 2011, 186, 3632–3641. [Google Scholar] [CrossRef] [Green Version]
- Schleicher, U.; Paduch, K.; Debus, A.; Obermeyer, S.; Konig, T.; Kling, J.C.; Ribechini, E.; Dudziak, D.; Mougiakakos, D.; Murray, P.J.; et al. TNF-Mediated Restriction of Arginase 1 Expression in Myeloid Cells Triggers Type 2 NO Synthase Activity at the Site of Infection. Cell Rep. 2016, 15, 1062–1075. [Google Scholar] [CrossRef] [Green Version]
- Brin, E.; Wu, K.; Lu, H.T.; He, Y.; Dai, Z.; He, W. PEGylated arginine deiminase can modulate tumor immune microenvironment by affecting immune checkpoint expression, decreasing regulatory T cell accumulation and inducing tumor T cell infiltration. Oncotarget 2017, 8, 58948–58963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Hu, H. Arginine Deiminase Induces Immunogenic Cell Death and Is Enhanced by N-acetylcysteine in Murine MC38 Colorectal Cancer Cells and MDA-MB-231 Human Breast Cancer Cells In Vitro. Molecules 2021, 26, 511. [Google Scholar] [CrossRef]
- Werner, A.; Koschke, M.; Leuchtner, N.; Luckner-Minden, C.; Habermeier, A.; Rupp, J.; Heinrich, C.; Conradi, R.; Closs, E.I.; Munder, M. Reconstitution of T Cell Proliferation under Arginine Limitation: Activated Human T Cells Take Up Citrulline via L-Type Amino Acid Transporter 1 and Use It to Regenerate Arginine after Induction of Argininosuccinate Synthase Expression. Front. Immunol. 2017, 8, 864. [Google Scholar] [CrossRef] [Green Version]
- Moynihan, K.D.; Irvine, D.J. Roles for Innate Immunity in Combination Immunotherapies. Cancer Res. 2017, 77, 5215–5221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Wang, G.; Huang, D.; Sui, M.; Xu, Y. Cancer Immunotherapy Based on Natural Killer Cells: Current Progress and New Opportunities. Front. Immunol. 2019, 10, 1205. [Google Scholar] [CrossRef]
- Szlosarek, P.W.; Klabatsa, A.; Pallaska, A.; Sheaff, M.; Smith, P.; Crook, T.; Grimshaw, M.J.; Steele, J.P.; Rudd, R.M.; Balkwill, F.R.; et al. In vivo loss of expression of argininosuccinate synthetase in malignant pleural mesothelioma is a biomarker for susceptibility to arginine depletion. Clin. Cancer Res. 2006, 12, 7126–7131. [Google Scholar] [CrossRef] [Green Version]
- Delage, B.; Luong, P.; Maharaj, L.; O'Riain, C.; Syed, N.; Crook, T.; Hatzimichael, E.; Papoudou-Bai, A.; Mitchell, T.J.; Whittaker, S.J.; et al. Promoter methylation of argininosuccinate synthetase-1 sensitises lymphomas to arginine deiminase treatment, autophagy and caspase-dependent apoptosis. Cell Death Dis. 2012, 3, e342. [Google Scholar] [CrossRef]
- Bowles, T.L.; Kim, R.; Galante, J.; Parsons, C.M.; Virudachalam, S.; Kung, H.J.; Bold, R.J. Pancreatic cancer cell lines deficient in argininosuccinate synthetase are sensitive to arginine deprivation by arginine deiminase. Int. J. Cancer 2008, 123, 1950–1955. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.X.; Cochrane, D.R.; Tessier-Cloutier, B.; Chen, S.Y.; Ho, G.; Pathak, K.V.; Alcazar, I.N.; Farnell, D.; Leung, S.; Cheng, A.; et al. Arginine Depletion Therapy with ADI-PEG20 Limits Tumor Growth in Argininosuccinate Synthase-Deficient Ovarian Cancer, Including Small-Cell Carcinoma of the Ovary, Hypercalcemic Type. Clin. Cancer Res. 2020, 26, 4402–4413. [Google Scholar] [CrossRef]
- Bean, G.R.; Kremer, J.C.; Prudner, B.C.; Schenone, A.D.; Yao, J.C.; Schultze, M.B.; Chen, D.Y.; Tanas, M.R.; Adkins, D.R.; Bomalaski, J.; et al. A metabolic synthetic lethal strategy with arginine deprivation and chloroquine leads to cell death in ASS1-deficient sarcomas. Cell Death Dis. 2016, 7, e2406. [Google Scholar] [CrossRef]
- Noh, E.J.; Kang, S.W.; Shin, Y.J.; Choi, S.H.; Kim, C.G.; Park, I.S.; Wheatley, D.N.; Min, B.H. Arginine deiminase enhances dexamethasone-induced cytotoxicity in human T-lymphoblastic leukemia CCRF-CEM cells. Int. J. Cancer 2004, 112, 502–508. [Google Scholar] [CrossRef]
- Jiang, H.; Guo, S.; Xiao, D.; Bian, X.; Wang, J.; Wang, Y.; Zhou, H.; Cai, J.; Zheng, Z. Arginine deiminase expressed in vivo, driven by human telomerase reverse transcriptase promoter, displays high hepatoma targeting and oncolytic efficiency. Oncotarget 2017, 8, 37694–37704. [Google Scholar] [CrossRef] [Green Version]
- Savaraj, N.; Wu, C.; Kuo, M.T.; You, M.; Wangpaichitr, M.; Robles, C.; Spector, S.; Feun, L. The relationship of arginine deprivation, argininosuccinate synthetase and cell death in melanoma. Drug Target. Insights 2007, 2, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Syed, N.; Langer, J.; Janczar, K.; Singh, P.; Lo Nigro, C.; Lattanzio, L.; Coley, H.M.; Hatzimichael, E.; Bomalaski, J.; Szlosarek, P.; et al. Epigenetic status of argininosuccinate synthetase and argininosuccinate lyase modulates autophagy and cell death in glioblastoma. Cell Death Dis. 2013, 4, e458. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.P.; Jungbluth, A.A.; Wu, B.W.; Bomalaski, J.; Old, L.J.; Ritter, G. Arginine deiminase PEG20 inhibits growth of small cell lung cancers lacking expression of argininosuccinate synthetase. Br. J. Cancer 2012, 106, 324–332. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Bian, X.; Liu, X.; Wang, Y.; Zhou, H.; Ma, X.; Quan, C.; Yao, Y.; Zheng, Z. Intracellular expression of arginine deiminase activates the mitochondrial apoptosis pathway by inhibiting cytosolic ferritin and inducing chromatin autophagy. BMC Cancer 2020, 20, 665. [Google Scholar] [CrossRef]
- Khalil, N.; Abi-Habib, R.J. [HuArgI (co)-PEG5000]-induced arginine deprivation leads to autophagy dependent cell death in pancreatic cancer cells. Investig. New Drugs 2020, 38, 1236–1246. [Google Scholar] [CrossRef]
- Gaude, E.; Frezza, C. Tissue-specific and convergent metabolic transformation of cancer correlates with metastatic potential and patient survival. Nat. Commun. 2016, 7, 13041. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; Lo, P.H.Y.; Saichi, N.; Ueda, K.; Hirata, M.; Tanikawa, C.; Matsuda, K. Argininosuccinate synthase 1 is an intrinsic Akt repressor transactivated by p53. Sci. Adv. 2017, 3, e1603204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.Y.; Wu, W.R.; Wang, Y.H.; Wang, J.W.; Fang, F.M.; Tsai, J.W.; Li, S.H.; Hung, H.C.; Yu, S.C.; Lan, J.; et al. ASS1 as a novel tumor suppressor gene in myxofibrosarcomas: Aberrant loss via epigenetic DNA methylation confers aggressive phenotypes, negative prognostic impact, and therapeutic relevance. Clin. Cancer Res. 2013, 19, 2861–2872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yau, T.; Cheng, P.N.; Chan, P.; Chan, W.; Chen, L.; Yuen, J.; Pang, R.; Fan, S.T.; Poon, R.T. A phase 1 dose-escalating study of pegylated recombinant human arginase 1 (Peg-rhArg1) in patients with advanced hepatocellular carcinoma. Investig. New Drugs 2013, 31, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Ensor, C.M.; Holtsberg, F.W.; Bomalaski, J.S.; Clark, M.A. Pegylated arginine deiminase (ADI-SS PEG20,000 mw) inhibits human melanomas and hepatocellular carcinomas in vitro and in vivo. Cancer Res. 2002, 62, 5443–5450. [Google Scholar] [PubMed]
- Ott, P.A.; Carvajal, R.D.; Pandit-Taskar, N.; Jungbluth, A.A.; Hoffman, E.W.; Wu, B.W.; Bomalaski, J.S.; Venhaus, R.; Pan, L.; Old, L.J.; et al. Phase I/II study of pegylated arginine deiminase (ADI-PEG 20) in patients with advanced melanoma. Investig. New Drugs 2013, 31, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keshet, R.; Szlosarek, P.; Carracedo, A.; Erez, A. Rewiring urea cycle metabolism in cancer to support anabolism. Nat Rev. Cancer 2018, 18, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Sahu, D.; Gupta, S.; Hau, A.M.; Nakashima, K.; Leivo, M.Z.; Searles, S.C.; Elson, P.; Bomalaski, J.S.; Casteel, D.E.; Boss, G.R.; et al. Argininosuccinate Synthetase 1 Loss in Invasive Bladder Cancer Regulates Survival through General Control Nonderepressible 2 Kinase-Mediated Eukaryotic Initiation Factor 2alpha Activity and Is Targetable by Pegylated Arginine Deiminase. Am. J. Pathol. 2017, 187, 200–213. [Google Scholar] [CrossRef] [Green Version]
- Roeksomtawin, S.; Navasumrit, P.; Waraprasit, S.; Parnlob, V.; Sricharunrat, T.; Bhudhisawasdi, V.; Savaraj, N.; Ruchirawat, M. Decreased argininosuccinate synthetase expression in Thai patients with cholangiocarcinoma and the effects of ADI-PEG20 treatment in CCA cell lines. Oncol. Lett. 2018, 16, 1529–1538. [Google Scholar] [CrossRef] [Green Version]
- Burrows, N.; Cane, G.; Robson, M.; Gaude, E.; Howat, W.J.; Szlosarek, P.W.; Pedley, R.B.; Frezza, C.; Ashcroft, M.; Maxwell, P.H. Hypoxia-induced nitric oxide production and tumour perfusion is inhibited by pegylated arginine deiminase (ADI-PEG20). Sci. Rep. 2016, 6, 22950. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Tsai, S.T.; Kuo, C.C.; Chang, J.S.; Jin, Y.T.; Chang, J.Y.; Hsiao, J.R. Arginine deprivation as a new treatment strategy for head and neck cancer. Oral. Oncol. 2012, 48, 1227–1235. [Google Scholar] [CrossRef]
- Tsai, W.B.; Aiba, I.; Lee, S.Y.; Feun, L.; Savaraj, N.; Kuo, M.T. Resistance to arginine deiminase treatment in melanoma cells is associated with induced argininosuccinate synthetase expression involving c-Myc/HIF-1alpha/Sp4. Mol. Cancer 2009, 8, 3223–3233. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.B.; Long, Y.; Park, J.R.; Chang, J.T.; Liu, H.; Rodriguez-Canales, J.; Savaraj, N.; Feun, L.G.; Davies, M.A.; Wistuba, I.I.; et al. Gas6/Axl is the sensor of arginine-auxotrophic response in targeted chemotherapy with arginine-depleting agents. Oncogene 2016, 35, 1632–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, Y.; Tsai, W.B.; Wang, D.; Hawke, D.H.; Savaraj, N.; Feun, L.G.; Hung, M.C.; Chen, H.H.; Kuo, M.T. Argininosuccinate synthetase 1 (ASS1) is a common metabolic marker of chemosensitivity for targeted arginine- and glutamine-starvation therapy. Cancer Lett. 2017, 388, 54–63. [Google Scholar] [CrossRef]
- Cheon, D.J.; Walts, A.E.; Beach, J.A.; Lester, J.; Bomalaski, J.S.; Walsh, C.S.; Ruprecht Wiedemeyer, W.; Karlan, B.Y.; Orsulic, S. Differential expression of argininosuccinate synthetase in serous and non-serous ovarian carcinomas. J. Pathol. Clin. Res. 2015, 1, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.K.; Deorukhkar, A.A.; Venkatesulu, B.P.; Li, X.; Tailor, R.; Bomalaski, J.S.; Krishnan, S. Exploiting Arginine Auxotrophy with Pegylated Arginine Deiminase (ADI-PEG20) to Sensitize Pancreatic Cancer to Radiotherapy via Metabolic Dysregulation. Mol. Cancer Ther. 2019, 18, 2381–2393. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; MacKenzie, E.D.; Karim, S.A.; Hedley, A.; Blyth, K.; Kalna, G.; Watson, D.G.; Szlosarek, P.; Frezza, C.; Gottlieb, E. Reversed argininosuccinate lyase activity in fumarate hydratase-deficient cancer cells. Cancer Metab. 2013, 1, 12. [Google Scholar] [CrossRef] [Green Version]
- Chalishazar, M.D.; Wait, S.J.; Huang, F.; Ireland, A.S.; Mukhopadhyay, A.; Lee, Y.; Schuman, S.S.; Guthrie, M.R.; Berrett, K.C.; Vahrenkamp, J.M.; et al. MYC-Driven Small-Cell Lung Cancer is Metabolically Distinct and Vulnerable to Arginine Depletion. Clin. Cancer Res. 2019, 25, 5107–5121. [Google Scholar] [CrossRef] [Green Version]
- Yeon, A.; You, S.; Kim, M.; Gupta, A.; Park, M.H.; Weisenberger, D.J.; Liang, G.; Kim, J. Rewiring of cisplatin-resistant bladder cancer cells through epigenetic regulation of genes involved in amino acid metabolism. Theranostics 2018, 8, 4520–4534. [Google Scholar] [CrossRef] [PubMed]
- McAlpine, J.A.; Lu, H.T.; Wu, K.C.; Knowles, S.K.; Thomson, J.A. Down-regulation of argininosuccinate synthetase is associated with cisplatin resistance in hepatocellular carcinoma cell lines: Implications for PEGylated arginine deiminase combination therapy. BMC Cancer 2014, 14, 621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savaraj, N.; Wu, C.; Li, Y.Y.; Wangpaichitr, M.; You, M.; Bomalaski, J.; He, W.; Kuo, M.T.; Feun, L.G. Targeting argininosuccinate synthetase negative melanomas using combination of arginine degrading enzyme and cisplatin. Oncotarget 2015, 6, 6295–6309. [Google Scholar] [CrossRef] [Green Version]
- Long, Y.; Tsai, W.B.; Chang, J.T.; Estecio, M.; Wangpaichitr, M.; Savaraj, N.; Feun, L.G.; Chen, H.H.; Kuo, M.T. Cisplatin-induced synthetic lethality to arginine-starvation therapy by transcriptional suppression of ASS1 is regulated by DEC1, HIF-1alpha, and c-Myc transcription network and is independent of ASS1 promoter DNA methylation. Oncotarget 2016, 7, 82658–82670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prudner, B.C.; Rathore, R.; Robinson, A.M.; Godec, A.; Chang, S.F.; Hawkins, W.G.; Hirbe, A.C.; Van Tine, B.A. Arginine Starvation and Docetaxel Induce c-Myc-Driven hENT1 Surface Expression to Overcome Gemcitabine Resistance in ASS1-Negative Tumors. Clin. Cancer Res. 2019, 25, 5122–5134. [Google Scholar] [CrossRef] [Green Version]
- Thongkum, A.; Wu, C.; Li, Y.Y.; Wangpaichitr, M.; Navasumrit, P.; Parnlob, V.; Sricharunrat, T.; Bhudhisawasdi, V.; Ruchirawat, M.; Savaraj, N. The Combination of Arginine Deprivation and 5-Fluorouracil Improves Therapeutic Efficacy in Argininosuccinate Synthetase Negative Hepatocellular Carcinoma. Int. J. Mol. Sci. 2017, 18, 1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daylami, R.; Muilenburg, D.J.; Virudachalam, S.; Bold, R.J. Pegylated arginine deiminase synergistically increases the cytotoxicity of gemcitabine in human pancreatic cancer. J. Exp. Clin. Cancer Res. 2014, 33, 102. [Google Scholar] [CrossRef]
- Alexandrou, C.; Al-Aqbi, S.S.; Higgins, J.A.; Boyle, W.; Karmokar, A.; Andreadi, C.; Luo, J.L.; Moore, D.A.; Viskaduraki, M.; Blades, M.; et al. Sensitivity of Colorectal Cancer to Arginine Deprivation Therapy is Shaped by Differential Expression of Urea Cycle Enzymes. Sci. Rep. 2018, 8, 12096. [Google Scholar] [CrossRef]
- Przystal, J.M.; Hajji, N.; Khozoie, C.; Renziehausen, A.; Zeng, Q.; Abaitua, F.; Hajitou, A.; Suwan, K.; Want, E.; Bomalaski, J.; et al. Efficacy of arginine depletion by ADI-PEG20 in an intracranial model of GBM. Cell Death Dis. 2018, 9, 1192. [Google Scholar] [CrossRef] [PubMed]
- Wangpaichitr, M.; Wu, C.; Bigford, G.; Theodoropoulos, G.; You, M.; Li, Y.Y.; Verona-Santos, J.; Feun, L.G.; Nguyen, D.M.; Savaraj, N. Combination of arginine deprivation with TRAIL treatment as a targeted-therapy for mesothelioma. Anticancer Res. 2014, 34, 6991–6999. [Google Scholar] [PubMed]
- You, M.; Savaraj, N.; Kuo, M.T.; Wangpaichitr, M.; Varona-Santos, J.; Wu, C.; Nguyen, D.M.; Feun, L. TRAIL induces autophagic protein cleavage through caspase activation in melanoma cell lines under arginine deprivation. Mol. Cell Biochem. 2013, 374, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.B.; Long, Y.; Chang, J.T.; Savaraj, N.; Feun, L.G.; Jung, M.; Chen, H.H.W.; Kuo, M.T. Chromatin remodeling system p300-HDAC2-Sin3A is involved in Arginine Starvation-Induced HIF-1alpha Degradation at the ASS1 promoter for ASS1 Derepression. Sci. Rep. 2017, 7, 10814. [Google Scholar] [CrossRef] [Green Version]
- Kuo, M.T.; Long, Y.; Tsai, W.B.; Li, Y.Y.; Chen, H.H.W.; Feun, L.G.; Savaraj, N. Collaboration Between RSK-EphA2 and Gas6-Axl RTK Signaling in Arginine Starvation Response That Confers Resistance to EGFR Inhibitors. Transl. Oncol. 2020, 13, 355–364. [Google Scholar] [CrossRef]
- Dillon, B.J.; Holtsberg, F.W.; Ensor, C.M.; Bomalaski, J.S.; Clark, M.A. Biochemical characterization of the arginine degrading enzymes arginase and arginine deiminase and their effect on nitric oxide production. Med. Sci Monit. 2002, 8, BR248–BR253. [Google Scholar]
- Agrawal, V.; Woo, J.H.; Mauldin, J.P.; Jo, C.; Stone, E.M.; Georgiou, G.; Frankel, A.E. Cytotoxicity of human recombinant arginase I (Co)-PEG5000 in the presence of supplemental L-citrulline is dependent on decreased argininosuccinate synthetase expression in human cells. Anticancer Drugs 2012, 23, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Basu, A.; Palm, T.; Hua, J.; Youngster, S.; Hwang, L.; Liu, H.C.; Li, X.; Peng, P.; Zhang, Y.; et al. Engineering an arginine catabolizing bioconjugate: Biochemical and pharmacological characterization of PEGylated derivatives of arginine deiminase from Mycoplasma arthritidis. Bioconjug. Chem. 2006, 17, 1447–1459. [Google Scholar] [CrossRef] [PubMed]
- Abou-Alfa, G.K.; Qin, S.; Ryoo, B.Y.; Lu, S.N.; Yen, C.J.; Feng, Y.H.; Lim, H.Y.; Izzo, F.; Colombo, M.; Sarker, D.; et al. Phase III randomized study of second line ADI-PEG 20 plus best supportive care versus placebo plus best supportive care in patients with advanced hepatocellular carcinoma. Ann. Oncol. 2018, 29, 1402–1408. [Google Scholar] [CrossRef]
- Hall, P.E.; Ready, N.; Johnston, A.; Bomalaski, J.S.; Venhaus, R.R.; Sheaff, M.; Krug, L.; Szlosarek, P.W. Phase II Study of Arginine Deprivation Therapy With Pegargiminase in Patients With Relapsed Sensitive or Refractory Small-cell Lung Cancer. Clin. Lung Cancer 2020, 21, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.J.; Hsiao, H.H.; Hsu, Y.T.; Liu, Y.C.; Kao, H.W.; Liu, T.C.; Cho, S.F.; Feng, X.; Johnston, A.; Bomalaski, J.S.; et al. Phase I study of ADI-PEG20 plus low-dose cytarabine for the treatment of acute myeloid leukemia. Cancer Med. 2021, 10, 2946–2955. [Google Scholar] [CrossRef]
- Tsai, H.J.; Jiang, S.S.; Hung, W.C.; Borthakur, G.; Lin, S.F.; Pemmaraju, N.; Jabbour, E.; Bomalaski, J.S.; Chen, Y.P.; Hsiao, H.H.; et al. A Phase II Study of Arginine Deiminase (ADI-PEG20) in Relapsed/Refractory or Poor-Risk Acute Myeloid Leukemia Patients. Sci. Rep. 2017, 7, 11253. [Google Scholar] [CrossRef] [Green Version]
- Lowery, M.A.; Yu, K.H.; Kelsen, D.P.; Harding, J.J.; Bomalaski, J.S.; Glassman, D.C.; Covington, C.M.; Brenner, R.; Hollywood, E.; Barba, A.; et al. A phase 1/1B trial of ADI-PEG 20 plus nab-paclitaxel and gemcitabine in patients with advanced pancreatic adenocarcinoma. Cancer 2017, 123, 4556–4565. [Google Scholar] [CrossRef]
- Harding, J.J.; Do, R.K.; Dika, I.E.; Hollywood, E.; Uhlitskykh, K.; Valentino, E.; Wan, P.; Hamilton, C.; Feng, X.; Johnston, A.; et al. A phase 1 study of ADI-PEG 20 and modified FOLFOX6 in patients with advanced hepatocellular carcinoma and other gastrointestinal malignancies. Cancer Chemother. Pharmacol. 2018, 82, 429–440. [Google Scholar] [CrossRef]
- Hall, P.E.; Lewis, R.; Syed, N.; Shaffer, R.; Evanson, J.; Ellis, S.; Williams, M.; Feng, X.; Johnston, A.; Thomson, J.A.; et al. A Phase I Study of Pegylated Arginine Deiminase (Pegargiminase), Cisplatin, and Pemetrexed in Argininosuccinate Synthetase 1-Deficient Recurrent High-grade Glioma. Clin. Cancer Res. 2019, 25, 2708–2716. [Google Scholar] [CrossRef] [Green Version]
- Beddowes, E.; Spicer, J.; Chan, P.Y.; Khadeir, R.; Corbacho, J.G.; Repana, D.; Steele, J.P.; Schmid, P.; Szyszko, T.; Cook, G.; et al. Phase 1 Dose-Escalation Study of Pegylated Arginine Deiminase, Cisplatin, and Pemetrexed in Patients With Argininosuccinate Synthetase 1-Deficient Thoracic Cancers. J. Clin. Oncol. 2017, 35, 1778–1785. [Google Scholar] [CrossRef] [Green Version]
- Yao, S.; Janku, F.; Subbiah, V.; Stewart, J.; Patel, S.P.; Kaseb, A.; Westin, S.N.; Naing, A.; Tsimberidou, A.M.; Hong, D.; et al. Phase 1 trial of ADI-PEG20 plus cisplatin in patients with pretreated metastatic melanoma or other advanced solid malignancies. Br. J. Cancer 2021, 124, 1533–1539. [Google Scholar] [CrossRef]
- Tomlinson, B.K.; Thomson, J.A.; Bomalaski, J.S.; Diaz, M.; Akande, T.; Mahaffey, N.; Li, T.; Dutia, M.P.; Kelly, K.; Gong, I.Y.; et al. Phase I Trial of Arginine Deprivation Therapy with ADI-PEG 20 Plus Docetaxel in Patients with Advanced Malignant Solid Tumors. Clin. Cancer Res. 2015, 21, 2480–2486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szlosarek, P.W.; Steele, J.P.; Nolan, L.; Gilligan, D.; Taylor, P.; Spicer, J.; Lind, M.; Mitra, S.; Shamash, J.; Phillips, M.M.; et al. Arginine Deprivation With Pegylated Arginine Deiminase in Patients With Argininosuccinate Synthetase 1-Deficient Malignant Pleural Mesothelioma: A Randomized Clinical Trial. JAMA Oncol. 2017, 3, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Feun, L.G.; Marini, A.; Walker, G.; Elgart, G.; Moffat, F.; Rodgers, S.E.; Wu, C.J.; You, M.; Wangpaichitr, M.; Kuo, M.T.; et al. Negative argininosuccinate synthetase expression in melanoma tumours may predict clinical benefit from arginine-depleting therapy with pegylated arginine deiminase. Br. J. Cancer 2012, 106, 1481–1485. [Google Scholar] [CrossRef] [PubMed]
- De Santo, C.; Cheng, P.; Beggs, A.; Egan, S.; Bessudo, A.; Mussai, F. Metabolic therapy with PEG-arginase induces a sustained complete remission in immunotherapy-resistant melanoma. J. Hematol. Oncol. 2018, 11, 68. [Google Scholar] [CrossRef]
- Yau, T.; Cheng, P.N.; Chan, P.; Chen, L.; Yuen, J.; Pang, R.; Fan, S.T.; Wheatley, D.N.; Poon, R.T. Preliminary efficacy, safety, pharmacokinetics, pharmacodynamics and quality of life study of pegylated recombinant human arginase 1 in patients with advanced hepatocellular carcinoma. Investig. New Drugs 2015, 33, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Khadeir, R.; Szyszko, T.; Szlosarek, P.W. Optimizing arginine deprivation for hard-to-treat cancers. Oncotarget 2017, 8, 96468–96469. [Google Scholar] [CrossRef]
- Jalili, M.; Scharm, M.; Wolkenhauer, O.; Damaghi, M.; Salehzadeh-Yazdi, A. Exploring the Metabolic Heterogeneity of Cancers: A Benchmark Study of Context-Specific Models. J. Pers. Med 2021, 11, 496. [Google Scholar]
Figure 1.
Arginine-related metabolic pathways. In normal cells, arginine can be directly transported into the cell or synthesized from citrulline and aspartate in the urea cycle. The resources of citrulline include glutamine, glutamate and proline. Arginine can be converted into nitric oxide (NO), urea and agmatine. Agmatine and ornithine are important resource for putrescine, which is a key precursor for polyamines. ASS: argininosuccinate synthetase, ASL: argininosuccinate lyase, ADC: arginine decarboxylase, AGM: agmatinase, ODC: ornithine decarboxylase, ARG: arginase, NOS: nitric oxide synthase, OCT: ornithine carbamoyl transferase, OAT: ornithine aminotransferase, PDG: phosphate-dependent glutaminase, P5CS: pyrroline-5-carboxylate synthase, ProDH: proline dehydrogenase.
Figure 1.
Arginine-related metabolic pathways. In normal cells, arginine can be directly transported into the cell or synthesized from citrulline and aspartate in the urea cycle. The resources of citrulline include glutamine, glutamate and proline. Arginine can be converted into nitric oxide (NO), urea and agmatine. Agmatine and ornithine are important resource for putrescine, which is a key precursor for polyamines. ASS: argininosuccinate synthetase, ASL: argininosuccinate lyase, ADC: arginine decarboxylase, AGM: agmatinase, ODC: ornithine decarboxylase, ARG: arginase, NOS: nitric oxide synthase, OCT: ornithine carbamoyl transferase, OAT: ornithine aminotransferase, PDG: phosphate-dependent glutaminase, P5CS: pyrroline-5-carboxylate synthase, ProDH: proline dehydrogenase.

Figure 2.
Arginine-related signaling pathways. Arginine can be transported by solute carriers (SLCs), including lysosomal SLC38A9, to activate the down-stream mTORC1 signaling pathway. In addition, arginine also can bind G-protein coupled receptor, GPRCA6, to activate down-stream RAS/ERK or PI3K pathway to reprogram the general.
Figure 2.
Arginine-related signaling pathways. Arginine can be transported by solute carriers (SLCs), including lysosomal SLC38A9, to activate the down-stream mTORC1 signaling pathway. In addition, arginine also can bind G-protein coupled receptor, GPRCA6, to activate down-stream RAS/ERK or PI3K pathway to reprogram the general.

Figure 3.
Arginine deprivation signaling pathways. In contrast to arginine stimulation, arginine deprivation suppresses mTOR pathway via activation of AMPK. Additionally, arginine deprivation induces p38-signaling pathway, which impairs mitochondria functions. Arginine deprivation also can induce ER stress via GCN2, resulting in aspartate exhaustion and decreased DNA synthesis. In general, arginine deprivation causes cancer cell death. EMT: epithelial-mesenchymal transition.
Figure 3.
Arginine deprivation signaling pathways. In contrast to arginine stimulation, arginine deprivation suppresses mTOR pathway via activation of AMPK. Additionally, arginine deprivation induces p38-signaling pathway, which impairs mitochondria functions. Arginine deprivation also can induce ER stress via GCN2, resulting in aspartate exhaustion and decreased DNA synthesis. In general, arginine deprivation causes cancer cell death. EMT: epithelial-mesenchymal transition.

Figure 4.
Arginine acts as an epigenetic regulator. In the presence of arginine, mTOR induces the ACLY and ACSS2 to increase the level of acetyl-CoA, which is the main resource of histone acetyl-transferases (HATs). Increased histone acetylation induces the chromatin-remodeling and gene activation. Conversely, arginine deprivation causes metabolites depletion, including alpha-ketoglutarate (α-KG), which down-regulates lysine-demethylases (KDMs) and induces globe repressive histone methylations.
Figure 4.
Arginine acts as an epigenetic regulator. In the presence of arginine, mTOR induces the ACLY and ACSS2 to increase the level of acetyl-CoA, which is the main resource of histone acetyl-transferases (HATs). Increased histone acetylation induces the chromatin-remodeling and gene activation. Conversely, arginine deprivation causes metabolites depletion, including alpha-ketoglutarate (α-KG), which down-regulates lysine-demethylases (KDMs) and induces globe repressive histone methylations.

Figure 5.
Arginine deprivation-induced types of cell death. The general cell-killing mechanisms by arginine deprivation include caspase-dependent apoptosis, caspases-independent apoptosis, caspase-independent autophagic cell death and necorptosis.
Figure 5.
Arginine deprivation-induced types of cell death. The general cell-killing mechanisms by arginine deprivation include caspase-dependent apoptosis, caspases-independent apoptosis, caspase-independent autophagic cell death and necorptosis.

Figure 6.
Therapy-resistance machinery. Upon arginine-depletion, TAM receptor, Axl, is activated by the growth arrest-specific protein 6 (Gas6), resulting in the activation of downstream signaling cascades. The activation of signaling pathways leads to MYC-induced ASS1 restoration and cell survival. In addition, the TREM1 induces CCL2 expression via ERK/AKT/mTOR/ STAT3 pathway, which leads to cell survival in ASS1-restoration cells, which also leads to the therapy-resistance.
Figure 6.
Therapy-resistance machinery. Upon arginine-depletion, TAM receptor, Axl, is activated by the growth arrest-specific protein 6 (Gas6), resulting in the activation of downstream signaling cascades. The activation of signaling pathways leads to MYC-induced ASS1 restoration and cell survival. In addition, the TREM1 induces CCL2 expression via ERK/AKT/mTOR/ STAT3 pathway, which leads to cell survival in ASS1-restoration cells, which also leads to the therapy-resistance.


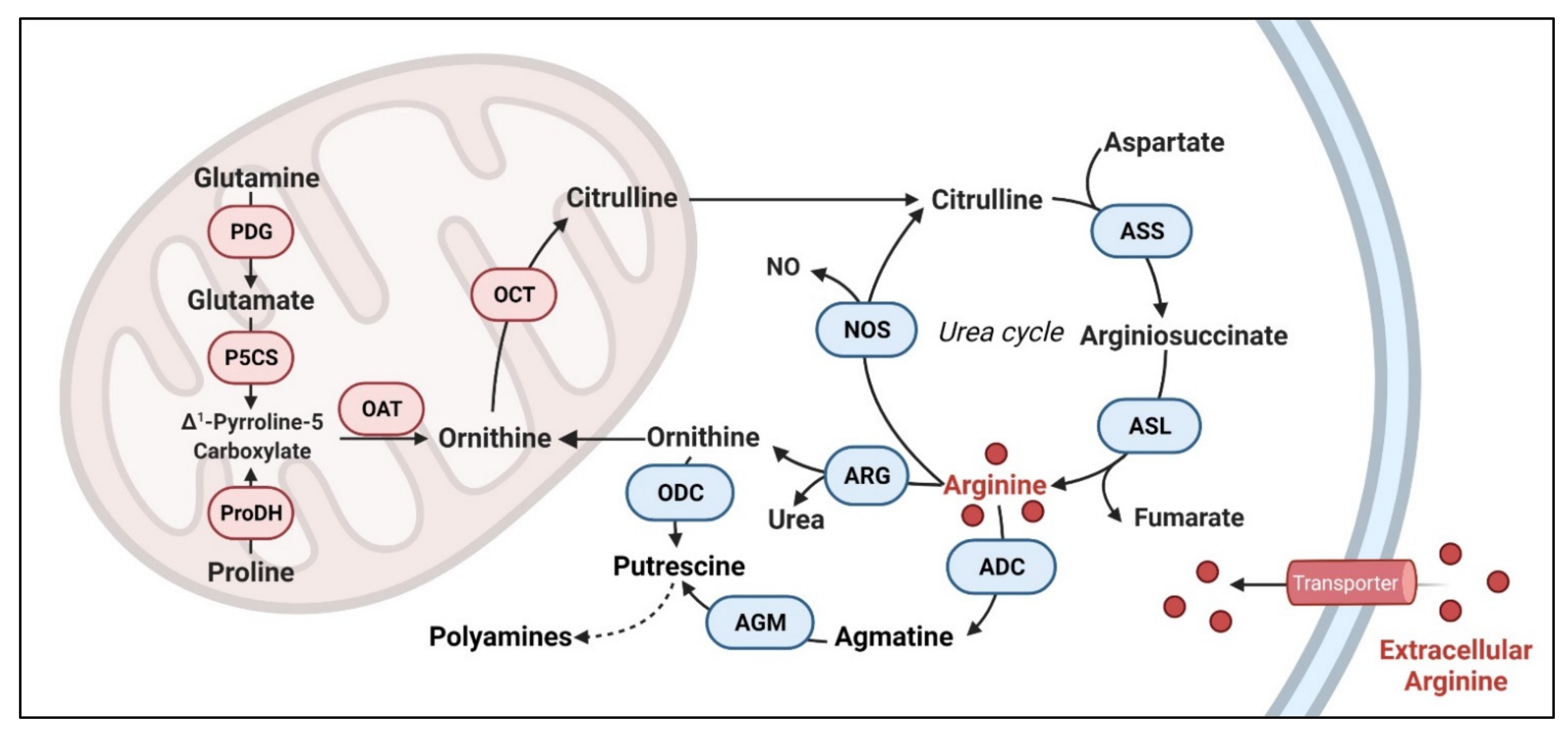{kind=link}
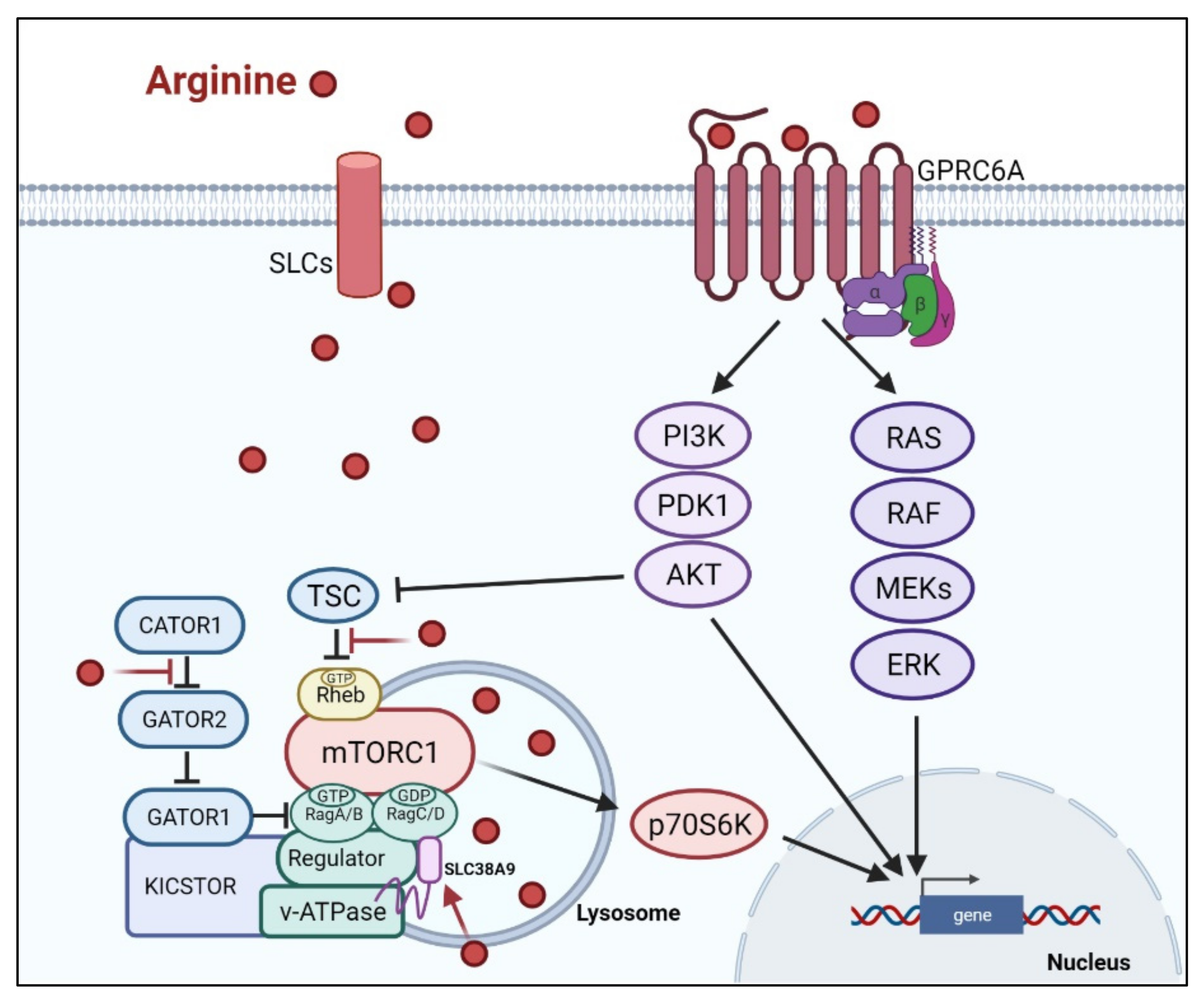{kind=link}
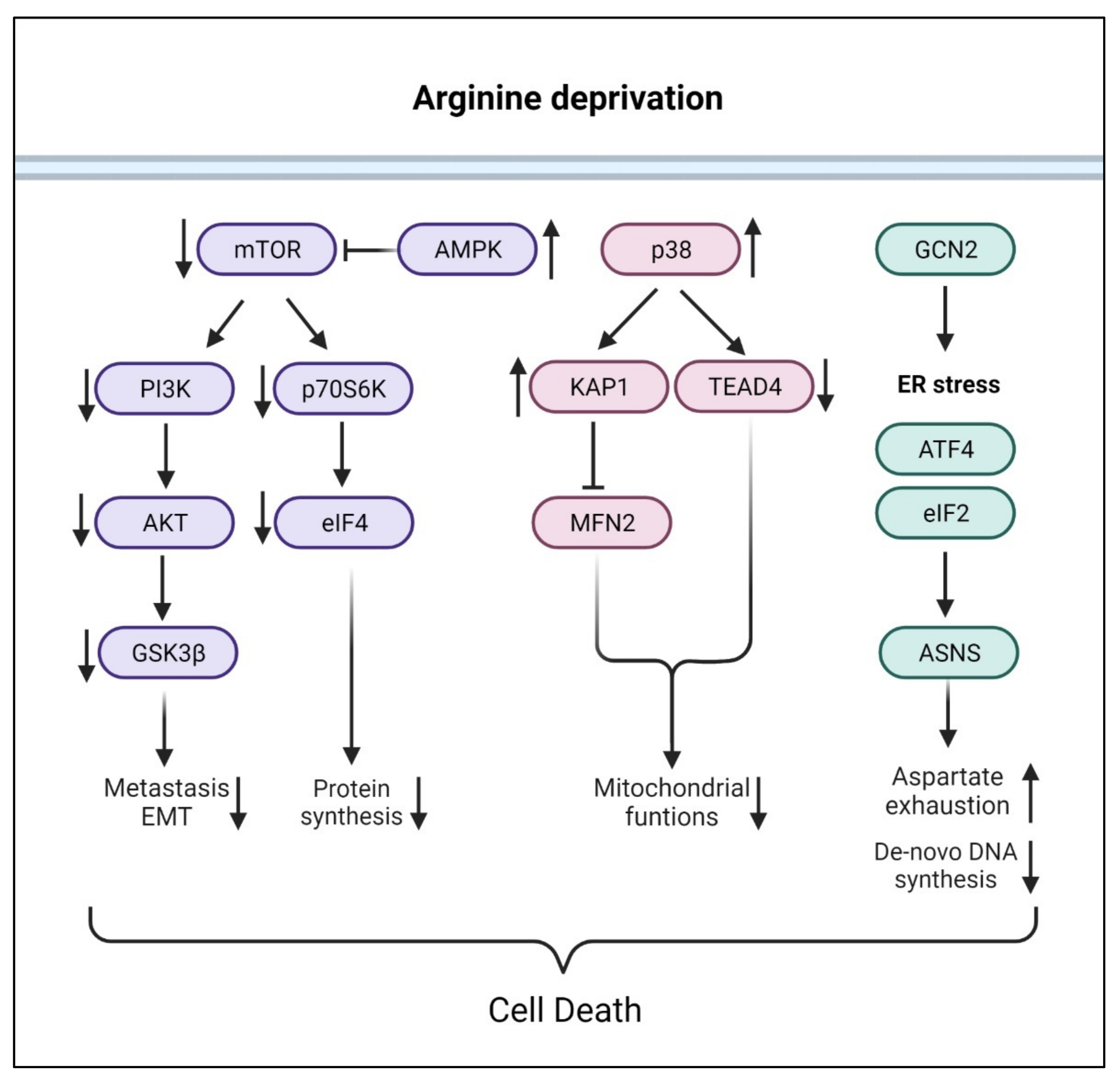{kind=link}
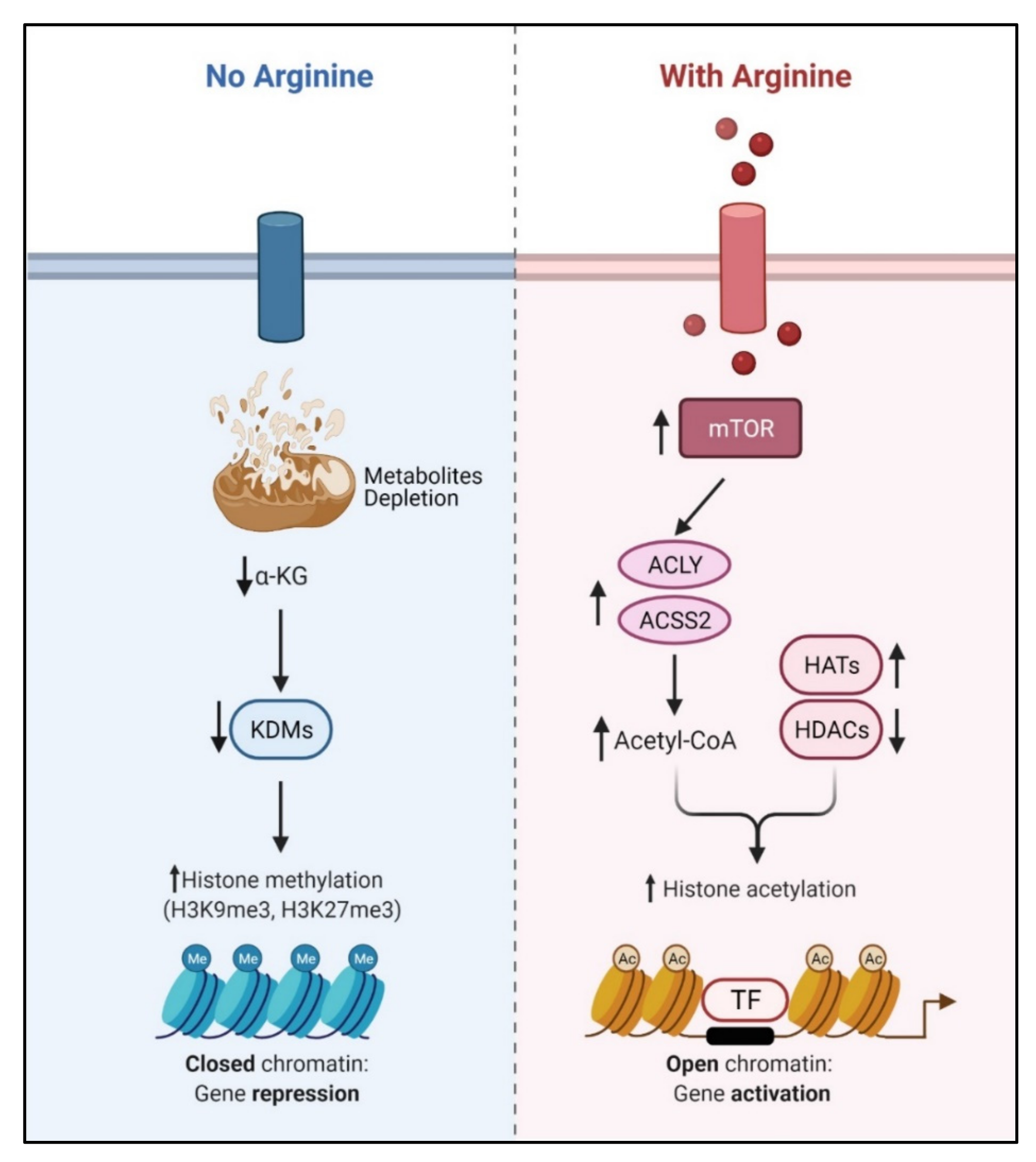{kind=link}
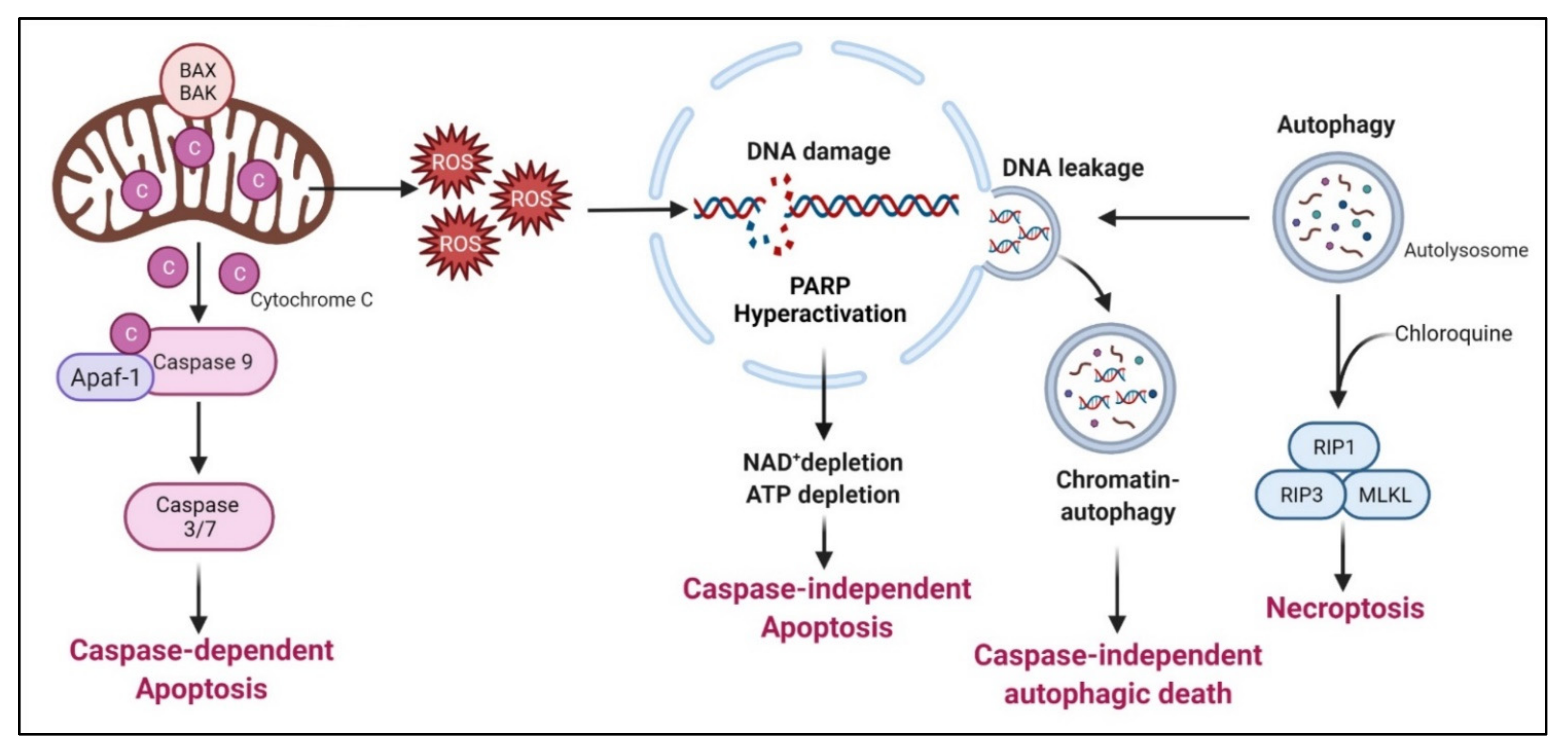{kind=link}
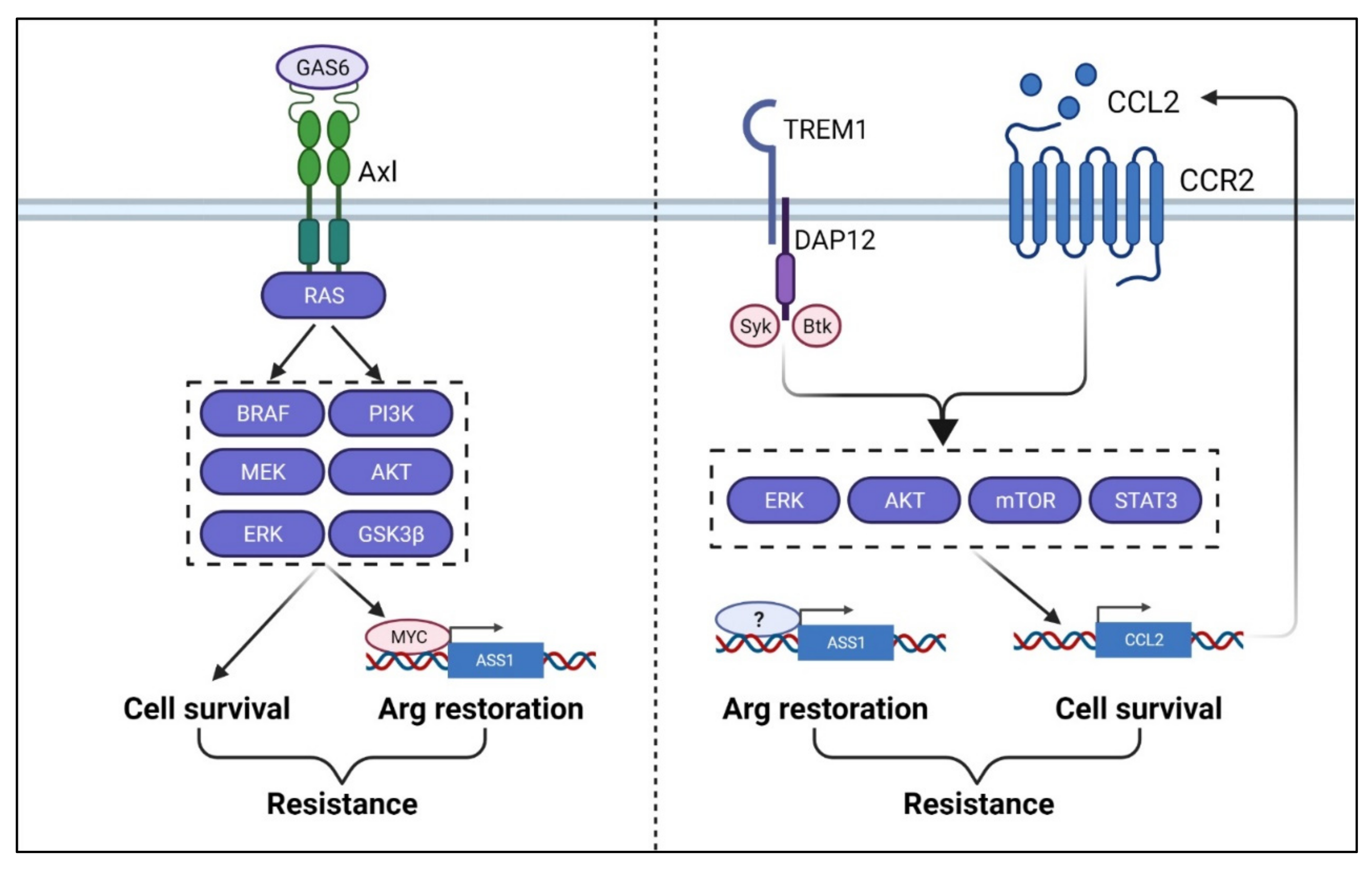{kind=link}
Table 1.
Summary of arginine transporters (solute carriers) in cancer vs. immune cells.
| Cancer | Immune Cells | ||
|---|---|---|---|
| SLC | Type | SLC | Type |
| SLC7A1 | Hepatocellular carcinoma [33], colorectal cancer [34], breast cancer [35], leukemia [36] | SLC7A1 | memory CD4(+) T cells and CD8(+) T cells [37] |
| SLC6A14 | Colorectal cancer [38], cervical cancer [39], pancreatic ductal adenocarcinomas [40], breast cancer [41] | SLC7A2 | Macrophage [42] |
| SLC7A3 | Osteosarcoma [43] | - | - |
| SLC7A9/SCL3A1 | Breast cancer [44] | - | - |
| SLC214C1 | Endometrial cancer [45] | - | - |
| SLC25A2 | Colorectal cancer [46] | - | - |
Table 2.
Comparison of arginine-depleting enzymes.
| Comparison of the Enzymatic Properties | Pegylated Arginine Deiminase (ADI-PEG20) | Pegylated Recombinant Human Arginase I (PEG-BCT-100) |
|---|---|---|
| Reaction products | L-citrulline + ammonia | L-ornithine + urea |
| Arginine affinity | High (Km~0.1–1 mM) | Low (Km~2.9mM) [125] |
| Half-life | 7 days [126] | 2~3 days [125] |
| Time requires to maximal arginine depletion in plasma | 4 days [127] | 4 h [125] |
| Origin of enzyme [4] | Mycoplasma | Human |
| Immunogenicity [128] | Antigenic (requires pegylation) | No |
| Clinical trials (Clinicaltrials.gov) | 25 trials Phase I, II, III | 10 trails Phase I, II |
Table 3.
ADI preclinical studies.
| ADI as a Single Agent | ||||
|---|---|---|---|---|
| Type of Cancer | Cell Line | Remarks | Ref | |
| Bladder cancer | T24, J82, UM-UC-3, 5637, RT112, and RT4 | ADI-PEG20 reduces the colony formation and cell viability by caspase-independent apoptotic cell death in ASS1-deficient cell lines. | [129] | |
| Breast | MDA-MD-231, ZR-75, T47D, MCF-7, SK-BR-3, MCF-10A | ADI-PEG20 induces the autophagy-dependent cell death, leading to mitochondrial dysfunction and growth inhibition. | [15] | |
| Cholangiocarcinoma | HuCCA, RmCCA-1, BJ-1 | ADI-PEG20 treatment reduces cholangiocarcinoma cell viability and proliferation. | [130] | |
| Colon carcinoma, Bladder carcinoma | HCT116, UMUC3 | ADI-PEG20 reduces hypoxia-induced NO pathway and vascular perfusion. | [131] | |
| Head and neck cancer | FaDu, HONE-1, KB, OEC-M1, UMSCC-1, SCC-4, SCC-15, SCC-25 | ADI-PEG20 inhibits the proliferation of head and neck cancer cells. | [132] | |
| Lymphomas | NcNc, Karpas-422, MyLa | ADI-PEG20 induces the caspase-dependent apoptosis in ASS1-methylated lymphoma cell lines. | [111] | |
| Melanoma | A2058, SK-Mel-2, A375 | ADI resistant cell lines are preferentially sensitive to glycolytic inhibitors and glutaminase inhibitors | [23] | |
| Melanoma | A375, A2058, SK-MEL-2 | ADI-resistance is due to the induction of ASS1 expression via c-Myc/HIF-1α/Sp4 pathway | [133] | |
| Melanoma, Breast cancer | UCSD354L, UACC62, UACC257, MEL1220, A20558, A375, SK-MEL-2, SK-MEL-5, SK-MEL-624, WM35, WM46, WM1799, WM2664, WM3248, SB-2, MDA-MB-231, SKOV3 | ADI-resistance is due to the induction of ASS1 expression via Gas6/Axl/Shp2 signal. | [134] | |
| Melanoma, Breast cancer | A2058, A375, BJ-1, WM2664, BT20, BT549, Hs578T, MDA-MB-157, MDA-MB-231, MDA-MB-436, MDA-MB-453, MDA-MB-468, HCC70, HCC38, HCC1806 | Knockdown of GLS increase the sensitivity to ADI-PEG20 | [135] | |
| Myxofibrosarcoma | OH931, NMFH-1, and NMFH-2 | ADI-PEG20 attenuates the cell viability in ASS1-deficient myxofibrosarcoma cells | [124] | |
| Ovarian cancer | OVCAR3, CAOV3, OVCAR4, IGROV1, TOV112D, OVCAR8, OV90, ES2, TOV21G | The ASS1 expression levels in ovarian cancer cell lines are inversely correlated with the susceptibility to ADI-PEG20 | [136] | |
| Pancreatic cancer | MiaPaCa-2, AsPc-1, BxPc-3, Capan1, HPAC, SW1990, L3.6pl, Panc-1 | ADI-PEG20 enhances the radio-sensitization by triggering the ER stress pathway, resulting in apoptosis in pancreatic tumor cells. | [137] | |
| Pancreatic cancer | BxPC-3, Capan-I, HPAC, HFAF-II, L3.3, MIA-PaCa-2, Panc-1 | ADI-PEG20 inhibits the pancreatic cancer cell growth via induction of apoptosis | [112] | |
| Prostate cancer | CWR22 | ADI-PEG20 induces the mitochondrial dysfunction, nuclear DNA leakage, and chromatin autophagy | [16] | |
| Renal cell carcinoma | UOK262 | ADI-PEG 20 inhibits the cellular proliferation in fumarate hydratase-deficient cells | [138] | |
| Small-cell lung cancer | GLC1, GLC8, NCI-H1092, NCI-H2141, SBC4, NCI-H82, NCI-H524, NCI-H446, NCI-H889, NCI-H69, NCI-H1963, H1048, DMS53 | MYC-driven human SCLC is preferentially sensitive to ADI-PEG20 in vivo | [139] | |
| Small-cell lung cancer | SW1222, SK-LC-13, SE1271, NCI-H82, NCI-H146, NCI-H209, SHP-77, NCI-H740, NCI-H889, NCI-H526, NCI-H69 | ADI-PEG20 induces apoptosis and autophagy in ASS1-negative SCLC cell lines | [119] | |
| Combination treatment with ADI-PEG20 | ||||
| Co-targeting reagent(s) | Type of cancer | Cell line | Remarks | Ref |
| Chloroquine | Glioblastoma | DBTRG, GAMG, SNB19, U87, U118, CCF, LN229, 8MG, T87G, MO59J, MO59K, 42MG | Combination of chloroquine inhibits autophagy and accelerates ADI-PEG20 induced cell death | [118] |
| Chloroquine | Sarcoma | Osteosarcoma (U-2 OS, MNNG/HOS, MG-63, NOS-1, HuO 9N2), Leiomyosarcoma (SK-LMS-1, SK-UT-1, SK-UT-1B), Synovial sarcoma (SYO-1, Fuji), Chondrosarcoma (HCH-1), Ewing’s sarcoma (LUPI, RD-ES, SK-ES), Alveolar soft part sarcoma (ASPS-1) | The combination of chloroquine with ADI-PEG20 causes synthetic lethality via necroptosis in sarcoma cell lines | [114] |
| Cisplatin | Bladder cancer | T24, J82, RT4 | Ass1 is down-regulated in cisplatin-resistant bladder cancer cells. The combination with ADI-PEG20 increases the susceptibility and induces apoptosis in cisplatin-resistant cancer cells. | [140] |
| Cisplatin | Hepatocellular carcinoma | Sk-Hep1, Huh7, Tong, HCC36, Hep3B, Malhavu, PLC5, Huh6, HepG2, SNU398 and SNU182 | The combination of cisplatin with ADI-PEG20 suppresses ASS1 expression in HCC cell lines | [141] |
| Cisplatin | Melanomas | A375, Sk-Mel2, A2058, Mel1220 | The combination of cisplatin with ADI-PEG20 enhances the cell death via apoptosis in melanoma cells | [142] |
| Cisplatin | Small-cell lung cancer, Ovarian cancer, Ovarian adenocarcinoma, Glioblastoma, Melanoma | SCLC, S, H465, SR2, A2780, A2008, A172, A2058 | The combination of cisplatin with ADI-PEG20 induces synergistical lethality | [143] |
| Docetaxel | Sarcoma, pancreatic cancer, and melanoma | SK-LMS-1, SK-UT-1, HTB-93, HT-1080, SK-MEL-2, AS-Pc-1, MiaPaCa-2, MNNG, RDES, and RD HPAC, SYO-1 and FUJI, LUPI, RH28 | The combination of docetaxel with ADI-PEG20 overcomes the gemcitabine resistance | [144] |
| 5-Flurouracie | Hepatocellular carcinoma (HCC) | BJ1, A2058, Mel1220, SNU398, SNU387, HepG2, Huh-1 | The combination of ADI-PEG20 with 5-FU improves the anti-tumor effect in ASS1-negative HCC cells | [145] |
| Gemcitabine | Pancreatic cancer | MIA-PaCa2, PANC-1, L3.3 | The combination of gemcitabine synergistically enhances ADI-PEG20 anti-tumor effect | [146] |
| Oxaliplatin | Colorectal cancer | HCT116, SW480, RKO, HT29 | The combination ADI-PEG20 with Oxaliplatin shows the synergistic growth inhibition in the ASS1-negative cell lines CRCs | [147] |
| Paclitaxel | Prostate cancer | CSR22Rv1, PC3, LNCaP | The combination of paclitaxel with ADI-PEG20 retards CWR22Rv1 tumor growth in vivo | [75] |
| Temozolomide | Glioblastoma | LN229 and U87 | The combination of ADI-PEG20 with Temozolomide suppresses the tumor growth irrespective of ASS1 status | [148] |
| TNF-related apoptosis-inducing ligan (TRAL) | Malignant pleural mesothelioma (MPM) | H211, H290, H2052, H2373, GARD REN, BJ-1 | The combination of TNF-related apoptosis-inducing ligan (TRAL) enhances ADI-PEG20 mediated apoptosis in MPM cells | [149] |
| TNF-related apoptosis-inducing ligan (TRAL) | Melanoma | A375, A2058 | The combination of TRAIL with ADI-PEG20 accelerates the cell death in melanoma cell lines | [150] |
| HAT inhibitor(s) | Melanoma | A2058, K-Mel-2, RCC4 | The combination of HAT inhibitors enhances ADI-PEG20 cell killing effect. | [151] |
| HDAC inhibitor(s) | Pancreatic cancer | Panc1, MiaPaca2, Panc02.03, HS766t, HPAF-II, Suit2, Su8686, Panc03.27, Panc10.05 | The combination of HDAC inhibitors with ADI-PEG20 induces the degradation of DNA repair enzyme, C-terminal-binding protein interacting protein (CtIP), resulting in DNA damage and apoptosis. | [82] |
| BET bromodomain-targeting c-Myc inhibitor | Melanoma | A2058 | The combination of ADI-PEG20 with JQ1, a BET bromodomain-targeting c-Myc inhibitor, significantly enhances the killing effect in ADI-resistant cells | [152] |
| Polyamide inhibitor | Mesothelioma | MSTO, Ju77, H28, H226 | The combination of polyamide inhibitor with ADI-PEG20 causes synthetically lethal effect in MPM cells | [87] |
| PHGDH or GLS inhibitor | Leiomyosarcoma, Melanoma | SKLMS1, SKUT1, SKMEL2 | The combination of ADI-PEG20 with PHGDH or GLS inhibitor significantly increases cell death | [22] |
| N-acetylcysteine | Breast | MDA-MD-231 | Combination of N-acetylcysteine with ADI-PEG 20 induces the immunogenic cell death. | [106] |
| PI3K/AKT inhibitor | Melanoma, Breast cancer | A2058, SK-MEL-2, MDA-MB-231, and A375 | The combination of PI3K/AKT inhibitor enhances ADI-PEG20–mediated cell apoptosis. | [24] |
Table 4.
Clinical trials for arginine-depletion enzymes and an arginase inhibitor.
| Start Date | NCT Number | Type of Cancer | Treatment | Phase | Status | Ref |
|---|---|---|---|---|---|---|
| Trials for pegylayed arginine deimnase | ||||||
| Jun-2020 | NCT04587830 | Glioblastoma Multiforme (GBM) | ADI-PEG20|Temozolomide | Phase 1 | Recruiting | |
| Apr-2019 | NCT03922880 | Uveal Melanoma | ADI-PEG20|Nivolumab|Ipilimumab | Phase 1 | Active, not recruiting | |
| Jun-2018 | NCT03498222 | Carcinoma, Non-Small-Cell Lung | ADI-PEG20|Atezolizumab|Pemetrexed|Carboplatin | Phase 1 | Withdrawn | |
| May-2018 | NCT03449901 | Soft Tissue Sarcoma|Osteosarcoma|Ewing’s Sarcoma| Small Cell Lung Cancer | ADI-PEG20|Gemcitabine|Docetaxel | Phase 2 | Active, not recruiting | |
| Aug-2017 | NCT02709512 | Mesothelioma | ADI-PEG20 |Pemetrexed and Cisplatin | Phase 2|Phase 3 | Recruiting | |
| Jul-2017 | NCT03254732 | Advanced Solid Cancers | ADI-PEG20|Pembrolizumab | Phase 1 | Active, not recruiting | |
| Jan-2017 | NCT02875093 | Acute Myeloid Leukemia | ADI-PEG20|Cytarabine | Phase 1 | Terminated | [158] |
| Jan-2015 | NCT01910012 | Acute Myeloid Leukemia | ADI-PEG20 | Phase 2 | Completed | [159] |
| Nov-2014 | NCT02101580 | Advanced Pancreatic Cancer | ADI-PEG20 Plus Nab-Paclitaxel and Gemcitabine | Phase 1 | Completed | [160] |
| Nov-2014 | NCT02101593 | Hepatocellular Carcinoma | ADI-PEG20|Sorafenib | Phase 1 | Completed | |
| Nov-2014 | NCT02102022 | Advanced Gastrointestinal (GI) Malignancies| Hepatocellular Carcinoma|Gastric Cancer|Colorectal Cancer | ADI-PEG20|modified FOLFOX6 | Phase 1|Phase 2 | Terminated | [161] |
| Oct-2014 | NCT02006030 | Unresectable Hepatocellular Carcinoma | ADI-PEG20|Transarterial chemoembolization | Phase 2 | Completed | |
| Apr-2014 | NCT02029690 | Pleural Mesothelioma Malignant Advanced|Peritoneal Mesothelioma Malignant Advanced|Non-squamous Non-small Cell Lung Carcinoma|Uveal Melanoma|Hepatocellular Carcinoma|Glioma|Sarcomatoid Carcinoma | ADI-PEG20 With Pemetrexed and Cisplatin | Phase 1 | Terminated | [162,163] |
| Apr-2014 | NCT01948843 | HER2 Negative Metastatic Breast Cancer | ADI-PEG20|Doxorubicin | Phase 1 | Completed | |
| Dec-2013 | NCT01910025 | Non-Hodgkin’s Lymphoma | ADI-PEG20 | Phase 2 | Completed | |
| Sep-2012 | NCT01665183 | Cutaneous Melanoma, Uveal Melanoma, Ovarian Carcinoma or Other Advanced Solid Tumors | ADI-PEG20|Cisplatin | Phase 1 | Completed | [164] |
| Dec-2011 | NCT01528384 | Arginosuccinate Synthetase Deficient | ADI-PEG20 | Phase 1 | Completed | |
| Sep-2011 | NCT01497925 | Solid Tumors|Prostate Cancer|Non-Small Cell Lung Cancer | ADI-PEG20|Docetaxel | Phase 1 | Completed | [165] |
| Jul-2011 | NCT01287585 | Hepatocellular Carcinoma | ADI-PEG20 | Phase 3 | Completed | [156] |
| Jan-2011 | NCT01266018 | Small Cell Lung Cancer | ADI-PEG20 | Phase 2 | Terminated | |
| Jan-2011 | NCT01279967 | Malignant Pleural Mesothelioma | ADI-PEG20 | Phase 2 | Unknown status | [166] |
| Jul-2007 | NCT00520299 | Metastatic Melanoma|Skin Cancer|Neoplasm | ADI-PEG20 | Phase 1|Phase 2 | Completed | [127] |
| Jun-2004 | NCT00450372 | Melanoma (Skin) | ADI-PEG20 | Phase 2 | Completed | [167] |
| Sep-2002 | NCT00056992 | Carcinoma, Hepatocellular | ADI-PEG20 | Phase 2 | Completed | |
| Sep-2001 | NCT00029900 | Melanoma| Neoplasm Metastasis | ADI-PEG20 | Phase 1 | Completed | |
| Trials for pegylated recombinant human arginase and arginase-1 peptide vaccine | ||||||
| Dec-2018 | NCT03689192 | Non Small Cell Lung Cancer|Urothelial Carcinoma|Malignant Melanoma|Ovarian Cancer|Colorectal Cancer|Breast Cancer|Squamous Cell Carcinoma of the Head and Neck|Metastatic Cancer | Arginase-1 Peptide Vaccine (ARG1-18,19,20) | Phase 1 | Recruiting | |
| Aug-2018 | NCT03455140 | Cancer|Pediatric Solid Tumor|Pediatric AML|Pediatric ALL | Pegylated Recombinant Human Arginase (BCT-100) | Phase 1|Phase 2 | Recruiting | |
| Sep-2016 | NCT02899286 | Relapsed or Refractory Acute Myeloid Leukemia | Pegylated Recombinant Human Arginase (BCT-100) | Phase 2 | Unknown status | |
| Aug-2016 | NCT02732184 | Acute Myeloid Leukemia|Myelodysplastic Syndrome | Co-ArgI-PEG modified human arginase I | Phase 2 | Completed | |
| Nov-2014 | NCT02285101 | Melanoma|Prostate Adenocarcinoma | Pegylated recombinant human arginase (PEG-BCT-100) | Phase 1 | Completed | [168] |
| Apr-2014 | NCT02089763 | Hepatocellular Carcinoma | Pegylated recombinant human arginase | Phase 2 | Terminated | |
| Apr-2014 | NCT02089633 | Hepatocellular Carcinoma | Pegylated recombinant human arginase|Oxaliplain|Capecitabine | Phase 2 | Completed | |
| Apr-2012 | NCT01551628 | Leukemia|Lymphoma | Recombinant human arginase 1 Peg5000 | Phase 1 | Terminated | |
| Mar-2010 | NCT01092091 | Neoplasm| Hepatocellular Carcinoma | Pegylated Recombinant Human Arginase I (BCT-100-002) | Phase 1|Phase 2 | Completed | [125,169] |
| May-2008 | NCT00988195 | Neoplasm| Hepatocellular Carcinoma | Pegylated Recombinant Human Arginase I|Doxorubicin | Phase 1 | Completed | |
| Trials for arginase inhibitor (INCB1158) | ||||||
| Sep-2019 | NCT03837509 | Relapsed or Refractory Multiple Myeloma | INCB001158|Daratumumab SC | Phase 1|Phase 2 | Recruiting | |
| Jul-2019 | NCT03910530 | Advanced Solid Tumors|Metastatic Solid Tumors | Retifanlimab|INCB001158| Retifanlimab + INCB001158 | Phase 1 | Active, not recruiting | |
| Mar-2018 | NCT03361228 | Solid Tumors | INCB001158|Epacadostat|Pembrolizumab | Phase 1|Phase 2 | Terminated | |
| Nov-2017 | NCT03314935 | Biliary Tract Cancer|Colorectal Cancer|Endometrial Cancer|Gastroesophageal Cancer|Ovarian Cancer|Solid Tumors | INCB001158|Oxaliplatin|Leucovorin|5-Fluorouracil|Gemcitabine|Cisplatin|Paclitaxel | Phase 1|Phase 2 | Active, not recruiting | |
| Sep-2016 | NCT02903914 | Metastatic Cancer|Solid Tumors|Colorectal Cancer|Gastric Cancer|Head and Neck Cancer|Lung Cancer|Renal Cell Carcinoma|Bladder Cancer|Urothelial Cancer|Mesothelioma | INCB001158|Pembrolizumab | Phase 1|Phase 2 | Active, not recruiting | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, C.-L.; Hsu, S.-C.; Ann, D.K.; Yen, Y.; Kung, H.-J. Arginine Signaling and Cancer Metabolism. Cancers 2021, 13, 3541. https://doi.org/10.3390/cancers13143541
AMA Style
Chen C-L, Hsu S-C, Ann DK, Yen Y, Kung H-J. Arginine Signaling and Cancer Metabolism. Cancers. 2021; 13(14):3541. https://doi.org/10.3390/cancers13143541
Chicago/Turabian StyleChen, Chia-Lin, Sheng-Chieh Hsu, David K. Ann, Yun Yen, and Hsing-Jien Kung. 2021. "Arginine Signaling and Cancer Metabolism" Cancers 13, no. 14: 3541. https://doi.org/10.3390/cancers13143541
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.