Novel Molecular Characterization of Colorectal Primary Tumors Based on miRNAs

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
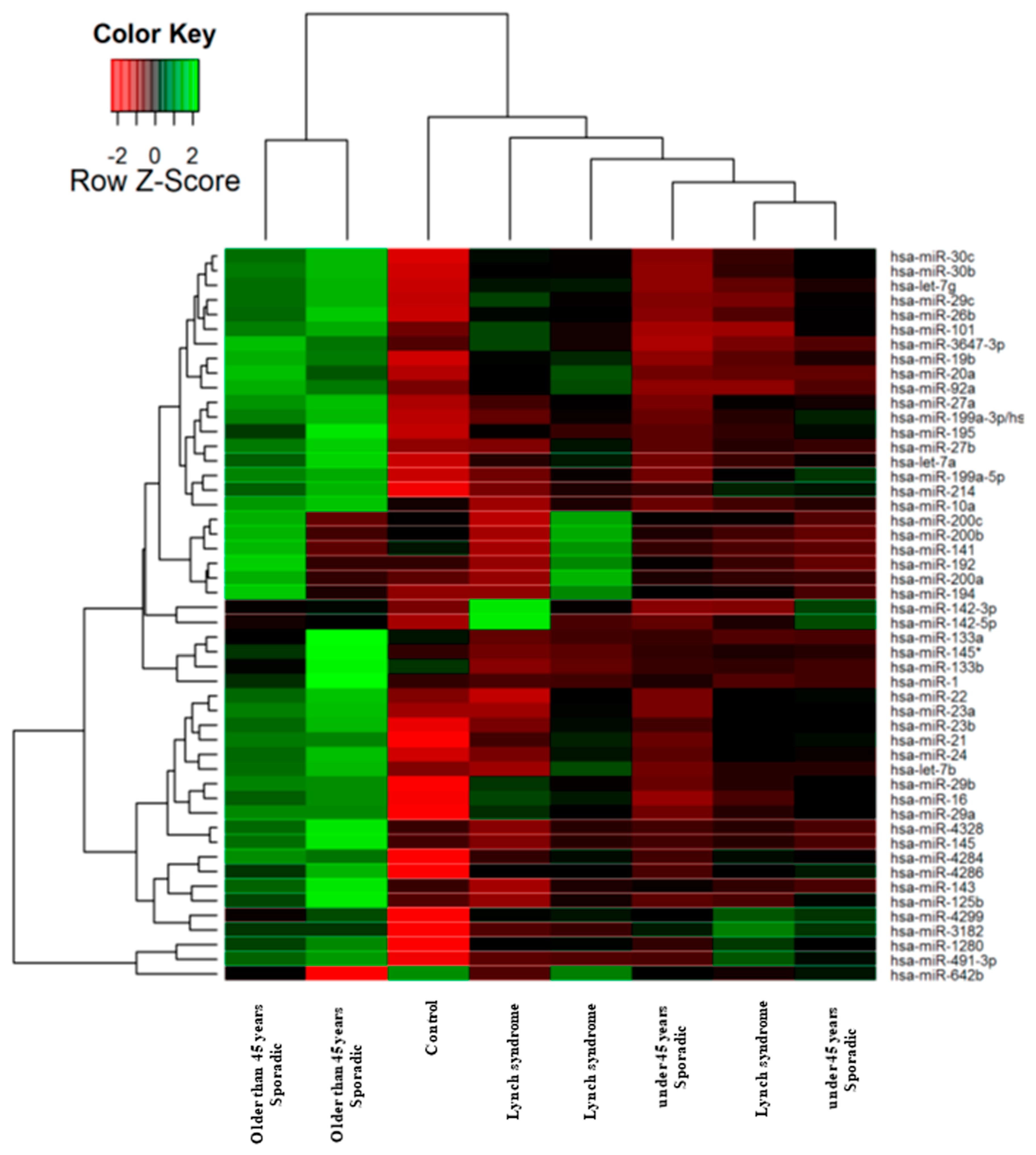
2.1. Identification of Tissue miRNAs Differentially Expressed in Colorectal Tumours Based on Age and MMR Defect
- (1)
- One non-tumour colonic mucosae as a control.
- (2)
- Sporadic CRC-diagnosed patients less than 45 years of age and without Lynch syndrome.
- (3)
- CRC patients with Lynch syndrome.
- (4)
- Sporadic CRC-diagnosed patients over 45 years of age and without Lynch syndrome.
2.2. Selection of miRNAs for qRT-PCR Validation in a Larger Patient Cohort
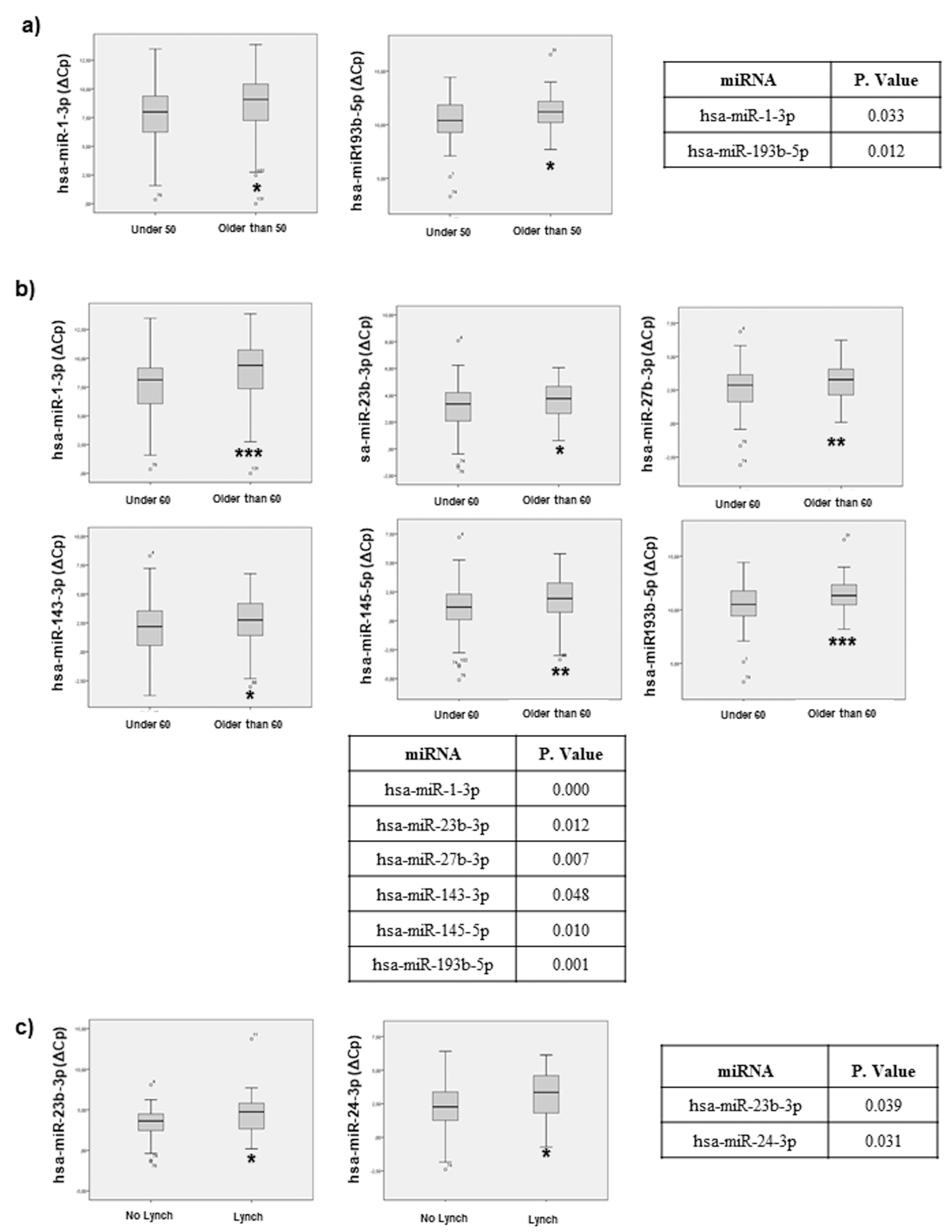
2.3. miRNAs Associated with Sporadic CRC Patient Age at Diagnosis and Discriminate Sporadic CRC vs. Lynch Syndrome Patients
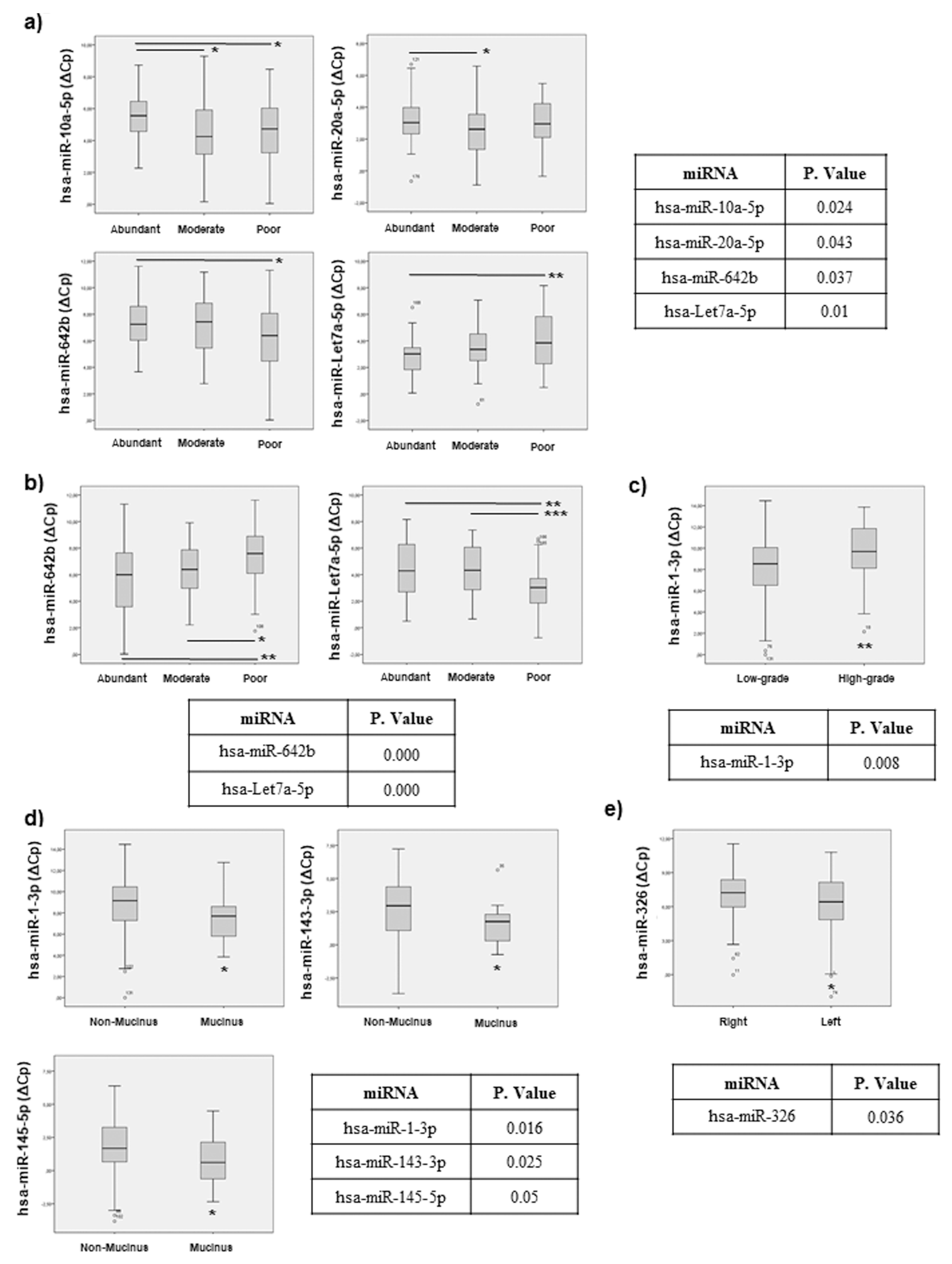
2.4. miRNAs Can Be Associated with Histopathological Features in CRC Patients
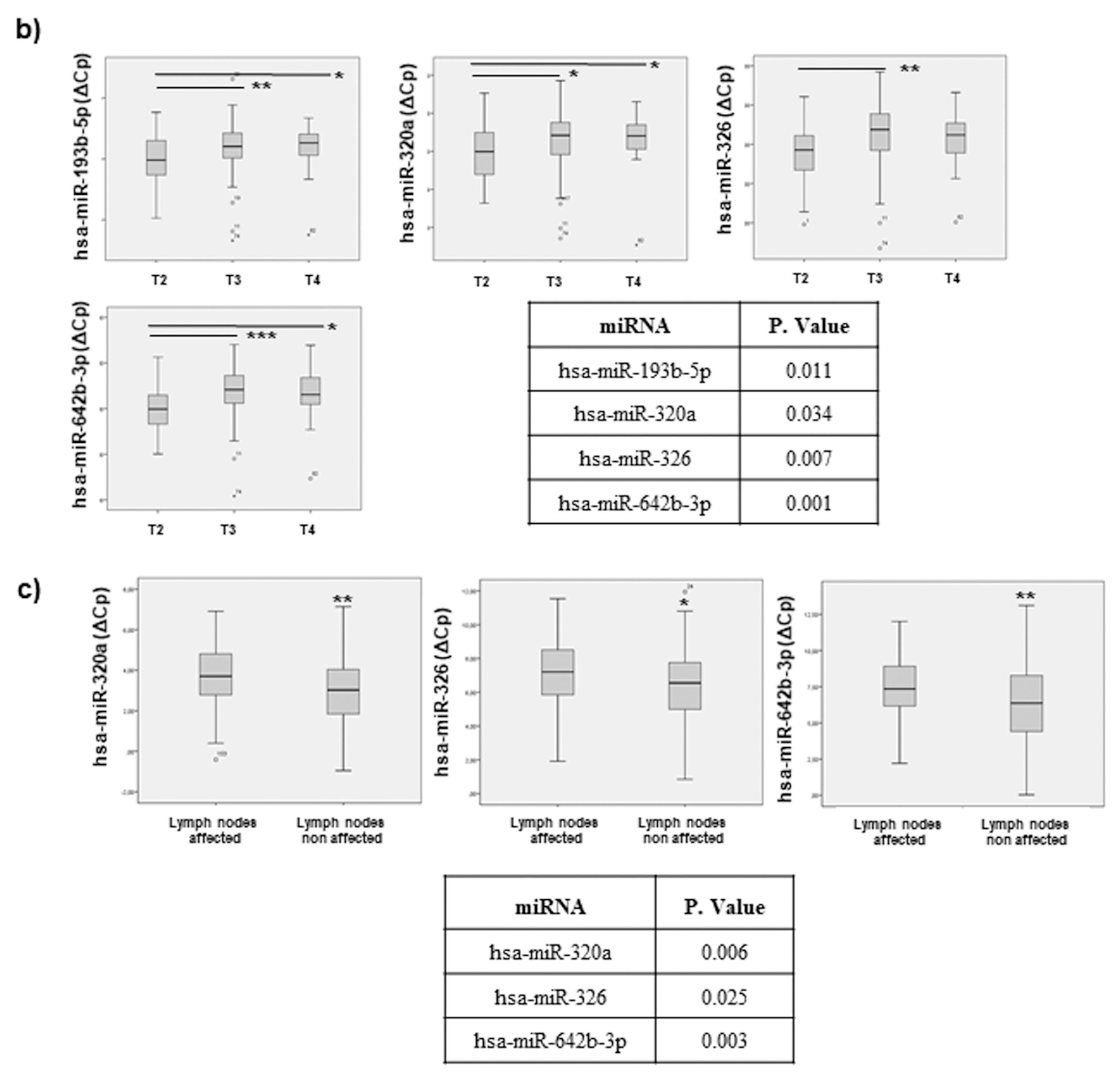
2.5. miRNAs Can Indicate CRC Progression: Association to Tumour Staging and Invasion
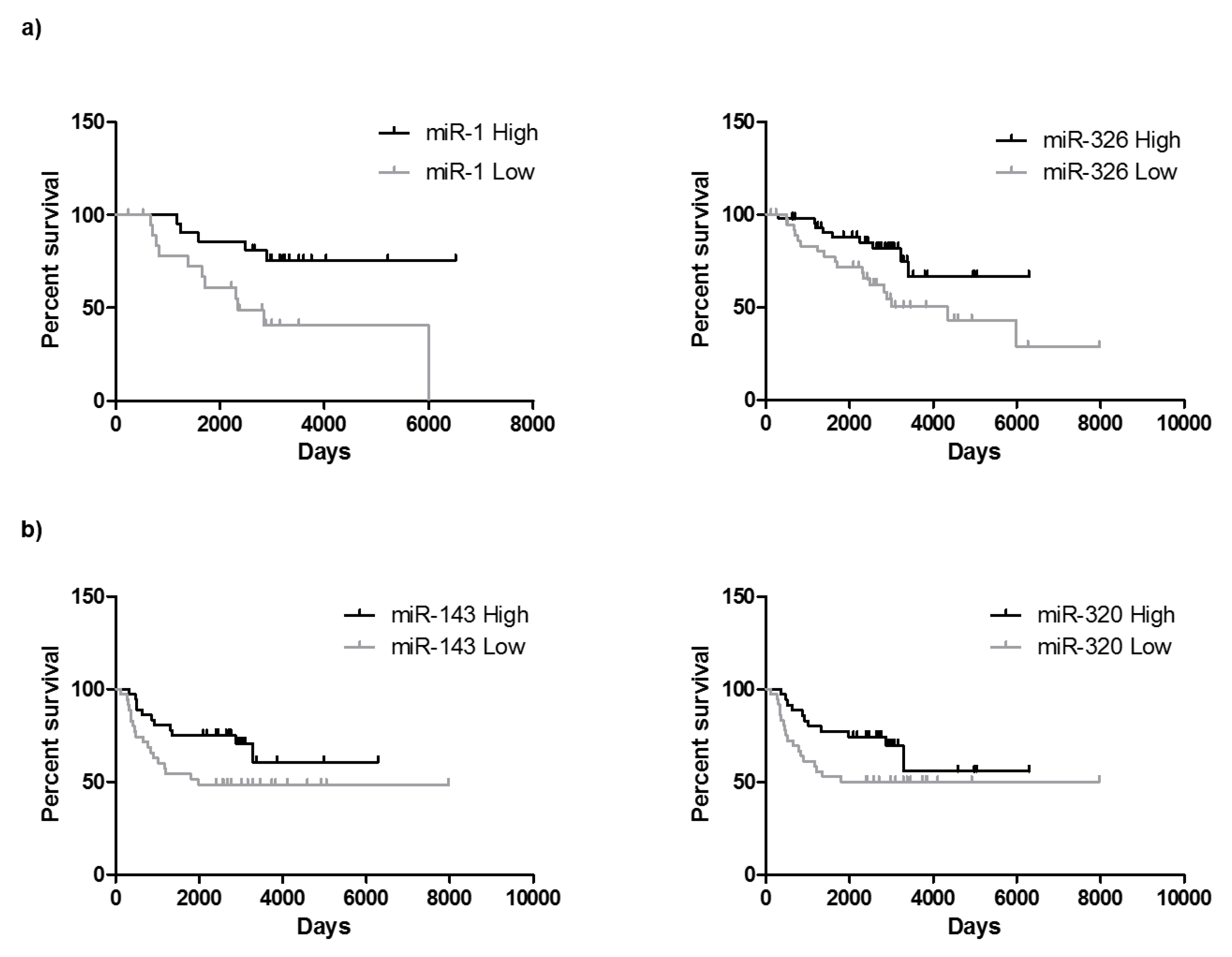
2.6. miRNAs Can Be Useful for Prognosis in CRC: Association with Overall Survival and Progression Free Survival
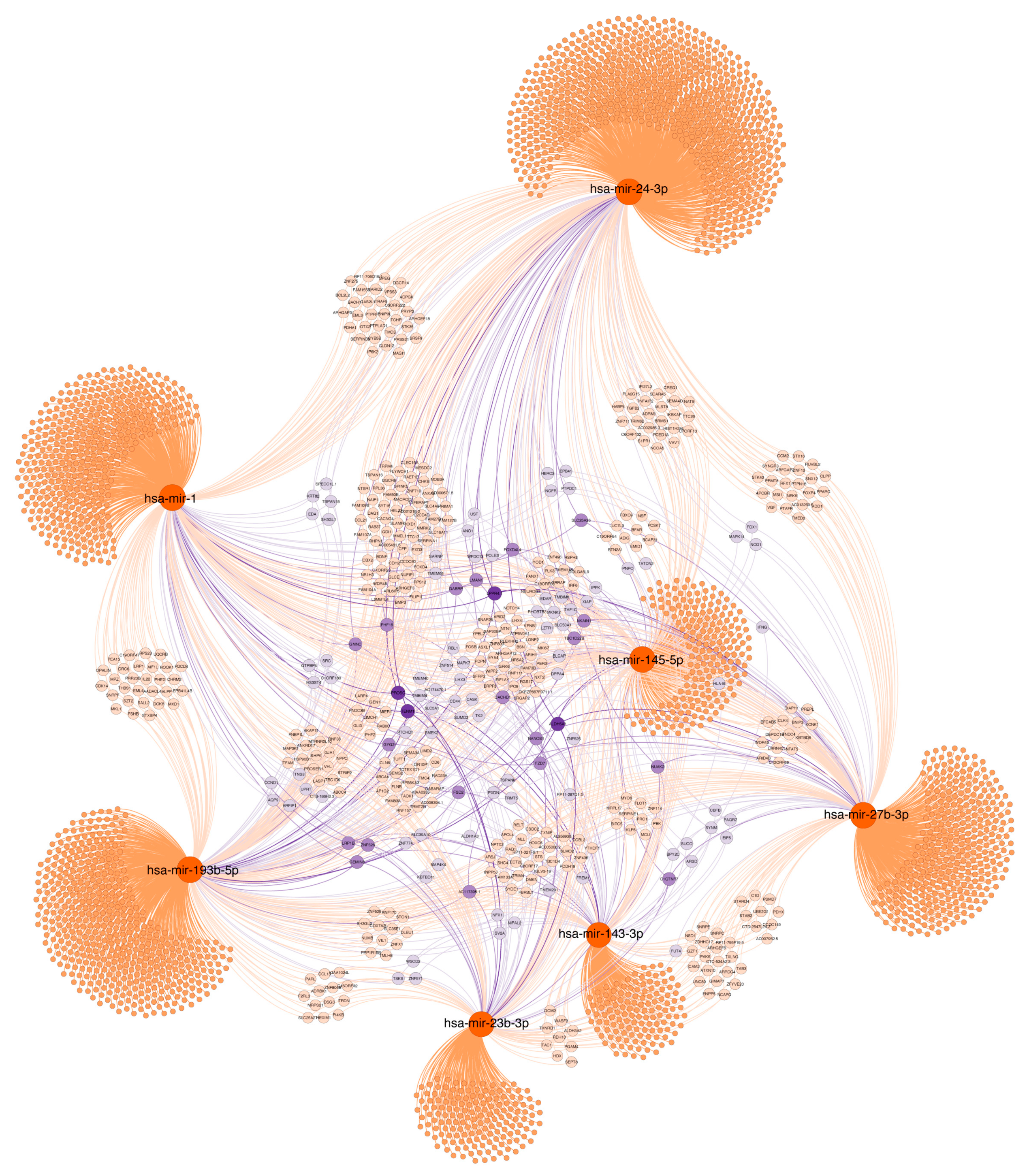
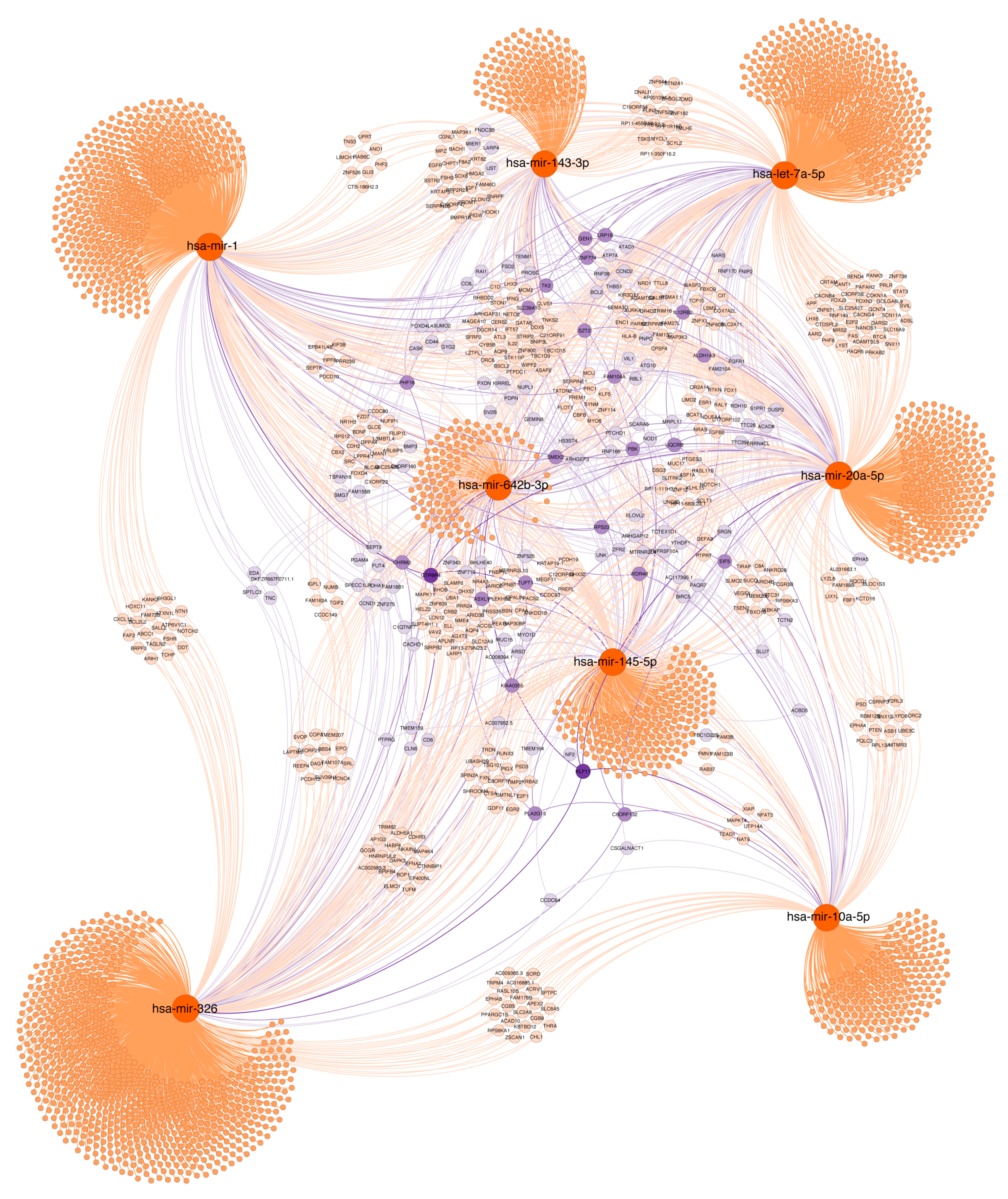
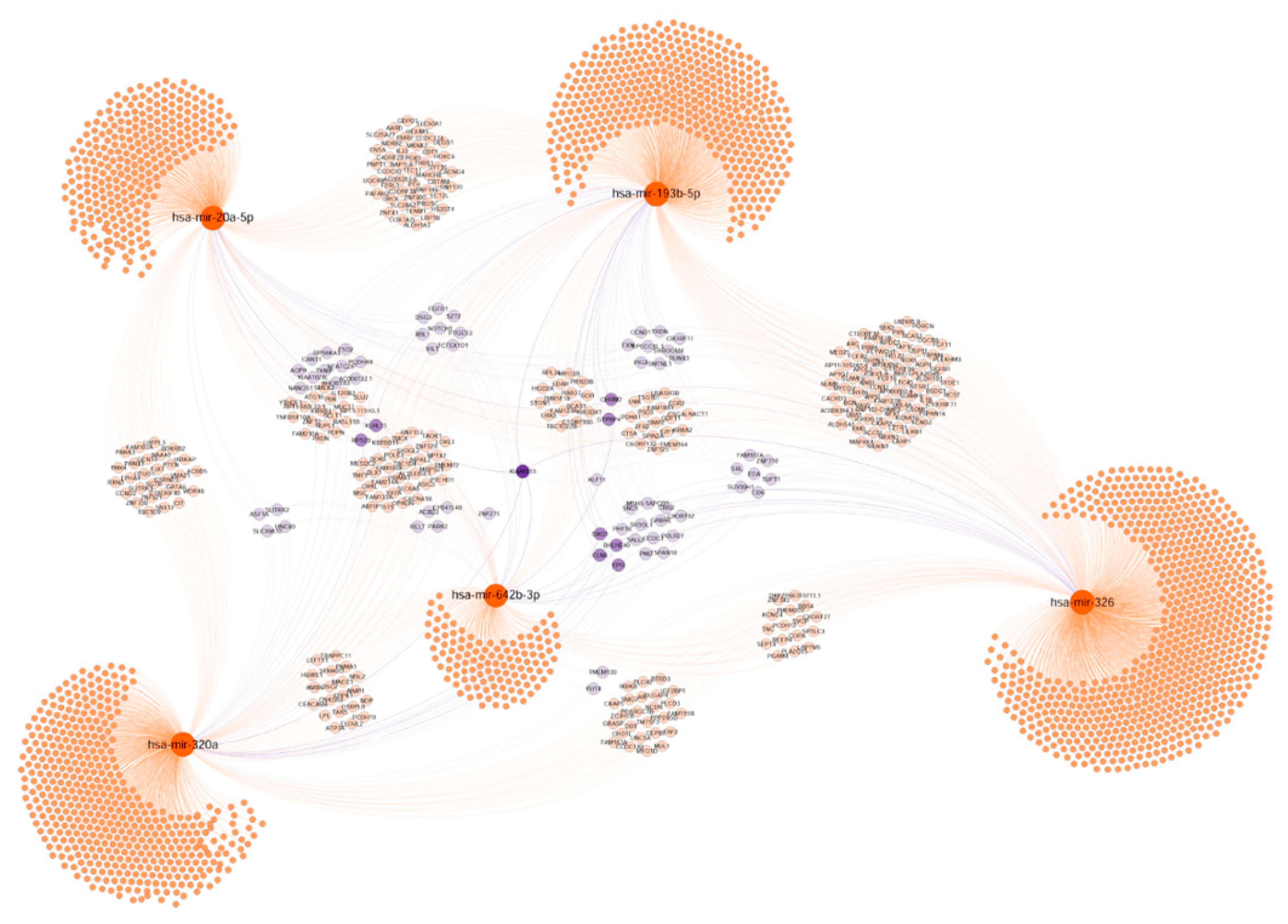
2.7. Functional Significance of miRNAs
2.7.1. Age and Lynch Syndrome
2.7.2. Histopathological Features
2.7.3. Tumour Progression
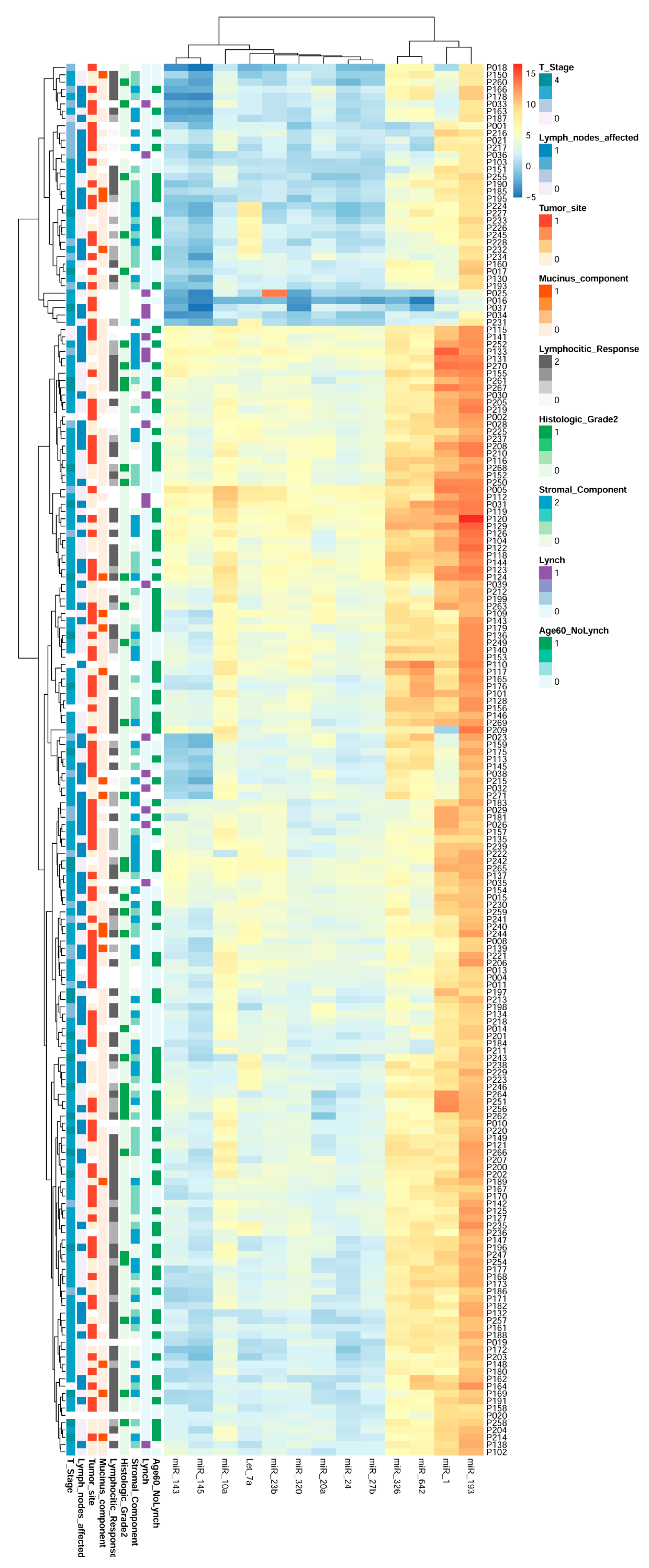
2.8. miRNAs Expression Could Regroup CRC Patients
3. Discussion
4. Material and Methods
4.1. Colorectal Cancer Biopsies Collection: Ethics Statement
4.2. microRNA Array Profiling
4.3. Validation of Selected miRNAs: Total RNA Extraction and RT-qPCR
4.4. Bioinformatics Studies
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Transcript Profiling
Abbreviations
| CRC | Colorectal cancer |
| MiRNAs | microRNAs |
| CIN | Chromosomal instability |
| MSI | Microsatellite instability |
| CIMP | CpG island methylator phenotype |
| MMR | DNA mismatch repair |
| CMS | Consensus molecular subtype |
| TNM | Tumour-node-metastasis |
| FFPE | Formalin-Fixed Paraffin-Embedded |
| LNA | Locked Nucleic Acid |
| Cq | quantitation cycle |
| GO | Gene ontology |
| NSCLC | Non-small-cell lung carcinoma |
| EMT | Epithelial to mesenchymal transition |
| BP | Biological process |
| CC | Cellular component |
| MF | Molecular function |
References
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Worthley, D.L.; Leggett, B.A. Colorectal cancer: Molecular features and clinical opportunities. Clin. Biochem. Rev. 2010, 31, 31–38. [Google Scholar] [PubMed]
- Arvelo, F.; Sojo, F.; Cotte, C. Biology of colorectal cancer. Ecancermedicalscience 2015, 9, 520. [Google Scholar] [CrossRef] [PubMed]
- Perez-Villamil, B.; Romera-Lopez, A.; Hernandez-Prieto, S.; Lopez-Campos, G.; Calles, A.; Lopez-Asenjo, J.A.; Sanz-Ortega, J.; Fernandez-Perez, C.; Sastre, J.; Alfonso, R.; et al. Colon cancer molecular subtypes identified by expression profiling and associated to stroma, mucinous type and different clinical behavior. BMC Cancer 2012, 12, 260. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.; Wong, H.L.; Templeton, A.; Tripathy, S.; Whiti Rogers, T.; Croxford, M.; Jones, I.; Sinnathamby, M.; Desai, J.; Tie, J.; et al. Analysis of local chronic inflammatory cell infiltrate combined with systemic inflammation improves prognostication in stage II colon cancer independent of standard clinicopathologic criteria. Int. J. Cancer. 2016, 138, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reynies, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The consensus molecular subtypes of colorectal cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Schellerer, V.S.; Merkel, S.; Schumann, S.C.; Schlabrakowski, A.; Fortsch, T.; Schildberg, C.; Hohenberger, W.; Croner, R.S. Despite aggressive histopathology survival is not impaired in young patients with colorectal cancer: CRC in patients under 50 years of age. Int. J. Colorectal Dis. 2012, 27, 71–79. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, J.B.; Maggard, M.A.; Livingston, E.H.; Yo, C.K. Colorectal cancer in the young. Am. J. Surg. 2004, 187, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.Y.; Fletcher, W.S.; Sullivan, E.S.; Vetto, J.T. Colorectal cancer in young patients: Characteristics and outcome. Am. Surg. 1994, 60, 607–612. [Google Scholar] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell. Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Pasquinelli, A.E. MicroRNAs and their targets: Recognition, regulation and an emerging reciprocal relationship. Nat. Rev. Genet. 2012, 13, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. microRNAs as oncogenes and tumor suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Welch, C.; Chen, Y.; Stallings, R.L. MicroRNA-34a functions as a potential tumor suppressor by inducing apoptosis in neuroblastoma cells. Oncogene 2007, 26, 5017–5022. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Lu, Z.; Liu, C.; Meng, Y.; Ma, Y.; Zhao, W.; Liu, J.; Yu, J.; Chen, J. miRNA-96 suppresses KRAS and functions as a tumor suppressor gene in pancreatic cancer. Cancer Res. 2010, 70, 6015–6025. [Google Scholar] [CrossRef] [PubMed]
- Jahid, S.; Sun, J.; Edwards, R.A.; Dizon, D.; Panarelli, N.C.; Milsom, J.W.; Sikandar, S.S.; Gumus, Z.H.; Lipkin, S.M. miR-23a promotes the transition from indolent to invasive colorectal cancer. Cancer Discov. 2012, 2, 540–553. [Google Scholar] [CrossRef] [PubMed]
- Slaby, O.; Svoboda, M.; Michalek, J.; Vyzula, R. MicroRNAs in colorectal cancer: Translation of molecular biology into clinical application. Mol. Cancer 2009, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Ju, J. Implications of microRNAs in colorectal cancer development, diagnosis, prognosis, and therapeutics. Front Genet. 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Slattery, M.L.; Wolff, E.; Hoffman, M.D.; Pellatt, D.F.; Milash, B.; Wolff, R.K. MicroRNAs and colon and rectal cancer: Differential expression by tumor location and subtype. Genes Chromosomes Cancer 2011, 50, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Balaguer, F.; Moreira, L.; Lozano, J.J.; Link, A.; Ramirez, G.; Shen, Y.; Cuatrecasas, M.; Arnold, M.; Meltzer, S.J.; Syngal, S.; et al. Colorectal cancers with microsatellite instability display unique miRNA profiles. Clin. Cancer Res. 2011, 17, 6239–6249. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Lotsari, J.E.; Al-Sohaily, S.; Warusavitarne, J.; Kohonen-Corish, M.R.; Peltomaki, P. Identification of subgroup-specific miRNA patterns by epigenetic profiling of sporadic and Lynch syndrome-associated colorectal and endometrial carcinoma. Clin. Epigenetics 2015, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Kerimis, D.; Kontos, C.K.; Christodoulou, S.; Papadopoulos, I.N.; Scorilas, A. Elevated expression of miR-24-3p is a potentially adverse prognostic factor in colorectal adenocarcinoma. Clin. Biochem. 2017, 50, 285–292. [Google Scholar] [CrossRef]
- Yin, Y.; Zhong, J.; Li, S.W.; Li, J.Z.; Zhou, M.; Chen, Y.; Sang, Y.; Liu, L. TRIM11, a direct target of miR-24-3p, promotes cell proliferation and inhibits apoptosis in colon cancer. Oncotarget 2016, 7, 86755–86765. [Google Scholar] [CrossRef]
- Chen, S.; Zong, Z.H.; Wu, D.D.; Sun, K.X.; Liu, B.L.; Zhao, Y. The role of metastasis-associated in colon cancer 1 (MACC1) in endometrial carcinoma tumorigenesis and progression. Mol. Carcinog. 2017, 56, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Boopathi, E.; Addya, S.; Phillips, B.; Rigoutsos, I.; Penn, R.B.; Rattan, S. Aging-associated changes in microRNA expression profile of internal anal sphincter smooth muscle: Role of microRNA-133a. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G964–G973. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.D.; Hong, Y.F.; Yusufuaji, Y.; Tang, B.P.; Zhou, X.H.; Xu, G.J.; Li, J.X.; Sun, L.; Zhang, J.H.; Xin, Q.; et al. Altered expression of hyperpolarization-activated cyclic nucleotide-gated channels and microRNA-1 and -133 in patients with age-associated atrial fibrillation. Mol. Med. Rep. 2015, 12, 3243–3248. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Hvid, T.; Kelly, M.; Lindegaard, B.; Dethlefsen, C.; Winding, K.; Mathur, N.; Scheele, C.; Pedersen, B.K.; Laye, M.J. Muscle specific miRNAs are induced by testosterone and independently upregulated by age. Front. Physiol. 2013, 4, 394. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, T.; Yoshino, H.; Enokida, H.; Hidaka, H.; Chiyomaru, T.; Nohata, N.; Kinoshita, T.; Fuse, M.; Seki, N.; Nakagawa, M. Novel molecular targets regulated by tumor suppressors microRNA-1 and microRNA-133a in bladder cancer. Int. J. Oncol. 2012, 40, 1821–1830. [Google Scholar] [CrossRef]
- Yu, Q.Q.; Wu, H.; Huang, X.; Shen, H.; Shu, Y.Q.; Zhang, B.; Xiang, C.C.; Yu, S.M.; Guo, R.H.; Chen, L. MiR-1 targets PIK3CA and inhibits tumorigenic properties of A549 cells. Biomed. Pharm. 2014, 68, 155–161. [Google Scholar] [CrossRef]
- Ukai, T.; Sato, M.; Akutsu, H.; Umezawa, A.; Mochida, J. MicroRNA-199a-3p, microRNA-193b, and microRNA-320c are correlated to aging and regulate human cartilage metabolism. J. Orthop. Res. 2012, 30, 1915–1922. [Google Scholar] [CrossRef]
- Kara, M.; Yumrutas, O.; Ozcan, O.; Celik, O.I.; Bozgeyik, E.; Bozgeyik, I.; Tasdemir, S. Differential expressions of cancer-associated genes and their regulatory miRNAs in colorectal carcinoma. Gene 2015, 567, 81–86. [Google Scholar] [CrossRef]
- Wu, K.; Zhao, Z.; Ma, J.; Chen, J.; Peng, J.; Yang, S.; He, Y. Deregulation of miR-193b affects the growth of colon cancer cells via transforming growth factor-beta and regulation of the SMAD3 pathway. Oncol. Lett. 2017, 13, 2557–2562. [Google Scholar] [CrossRef] [PubMed]
- Senovilla, L.; Vacchelli, E.; Galon, J.; Adjemian, S.; Eggermont, A.; Fridman, W.H.; Sautes-Fridman, C.; Ma, Y.; Tartour, E.; Zitvogel, L.; et al. Trial watch: Prognostic and predictive value of the immune infiltrate in cancer. Oncoimmunology 2012, 1, 1323–1343. [Google Scholar] [CrossRef] [PubMed]
- Epis, M.R.; Giles, K.M.; Kalinowski, F.C.; Barker, A.; Cohen, R.J.; Leedman, P.J. Regulation of expression of deoxyhypusine hydroxylase (DOHH), the enzyme that catalyzes the activation of eIF5A, by miR-331-3p and miR-642-5p in prostate cancer cells. J. Biol. Chem. 2012, 287, 35251–35259. [Google Scholar] [CrossRef] [PubMed]
- Nordentoft, I.; Birkenkamp-Demtroder, K.; Agerbaek, M.; Theodorescu, D.; Ostenfeld, M.S.; Hartmann, A.; Borre, M.; Orntoft, T.F.; Dyrskjot, L. miRNAs associated with chemo-sensitivity in cell lines and in advanced bladder cancer. BMC Med. Genom. 2012, 5, 40. [Google Scholar] [CrossRef] [PubMed]
- Thorns, C.; Schurmann, C.; Gebauer, N.; Wallaschofski, H.; Kumpers, C.; Bernard, V.; Feller, A.C.; Keck, T.; Habermann, J.K.; Begum, N.; et al. Global microRNA profiling of pancreatic neuroendocrine neoplasias. Anticancer Res. 2014, 34, 2249–2254. [Google Scholar] [PubMed]
- Sayagues, J.M.; Corchete, L.A.; Gutierrez, M.L.; Sarasquete, M.E.; Del Mar Abad, M.; Bengoechea, O.; Ferminan, E.; Anduaga, M.F.; Del Carmen, S.; Iglesias, M.; et al. Genomic characterization of liver metastases from colorectal cancer patients. Oncotarget 2016, 7, 72908–72922. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, Y.; Wang, H.; Zhang, G.; Ding, Y.; Zhao, L. Tumor suppressor miR-1 restrains epithelial-mesenchymal transition and metastasis of colorectal carcinoma via the MAPK and PI3K/AKT pathway. J. Transl. Med. 2014, 12, 244. [Google Scholar] [CrossRef]
- Xu, X.; Wu, X.; Jiang, Q.; Sun, Y.; Liu, H.; Chen, R.; Wu, S. Downregulation of microRNA-1 and microRNA-145 contributes synergistically to the development of colon cancer. Int. J. Mol. Med. 2015, 36, 1630–1638. [Google Scholar] [CrossRef]
- Zhang, G.J.; Li, Y.; Zhou, H.; Xiao, H.X.; Zhou, T. miR20a is an independent prognostic factor in colorectal cancer and is involved in cell metastasis. Mol. Med. Rep. 2014, 10, 283–291. [Google Scholar] [CrossRef]
- Rotelli, M.T.; Di Lena, M.; Cavallini, A.; Lippolis, C.; Bonfrate, L.; Chetta, N.; Portincasa, P.; Altomare, D.F. Fecal microRNA profile in patients with colorectal carcinoma before and after curative surgery. Int. J. Colorectal Dis. 2015, 30, 891–898. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.; Wu, H.; Li, Y.; Zhang, Y.; Liu, M.; Li, X.; Tang, H. miR-10a suppresses colorectal cancer metastasis by modulating the epithelial-to-mesenchymal transition and anoikis. Cell Death Dis. 2017, 8, e2739. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Hui, H.; Wang, L.J.; Wang, H.; Liu, Q.F.; Han, S.X. MicroRNA-326 functions as a tumor suppressor in colorectal cancer by targeting the nin one binding protein. Oncol. Rep. 2015, 33, 2309–2318. [Google Scholar] [CrossRef] [PubMed]
- Omrane, I.; Kourda, N.; Stambouli, N.; Privat, M.; Medimegh, I.; Arfaoui, A.; Uhrhammer, N.; Bougatef, K.; Baroudi, O.; Bouzaienne, H.; et al. MicroRNAs 146a and 147b biomarkers for colorectal tumor’s localization. Biomed. Res. Int. 2014, 2014, 584852. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Luo, Y.; Mu, Y.F.; Qin, S.L.; Qi, Y.; Qiu, Y.E.; Zhong, M. miR-193b directly targets STMN1 and inhibits the malignant phenotype in colorectal cancer. Am. J. Cancer. Res. 2016, 6, 2463–2475. [Google Scholar] [PubMed]
- Hur, K.; Toiyama, Y.; Schetter, A.J.; Okugawa, Y.; Harris, C.C.; Boland, C.R.; Goel, A. Identification of a metastasis-specific MicroRNA signature in human colorectal cancer. J. Natl. Cancer. Inst. 2015, 107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, X.; Liu, Y.; Ye, Y.; Zhang, H.; He, P.; Zhang, Q.; Dong, L.; Liu, Y.; Dong, J. microRNA-320a inhibits tumor invasion by targeting neuropilin 1 and is associated with liver metastasis in colorectal cancer. Oncol. Rep. 2012, 27, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Dong, T.; Zhou, H.; Wang, L.; Huang, A.; Feng, B.; Quan, Y.; Jin, R.; Zhang, W.; Sun, J.; et al. miR-320a suppresses colorectal cancer progression by targeting Rac1. Carcinogenesis 2014, 35, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.K.; Jung, S.H.; Yim, S.H.; Jung, J.H.; Choi, H.J.; Kang, W.K.; Park, S.W.; Oh, S.T.; Kim, J.G.; Lee, S.H.; et al. Predictive microRNAs for lymph node metastasis in endoscopically resectable submucosal colorectal cancer. Oncotarget 2016, 7, 32902–32915. [Google Scholar] [CrossRef] [PubMed]
- Zaravinos, A. The regulatory role of MicroRNAs in EMT and cancer. J. Oncol. 2015, 2015, 865816. [Google Scholar] [CrossRef]
- Shirafkan, N.; Mansoori, B.; Mohammadi, A.; Shomali, N.; Ghasbi, M.; Baradaran, B. MicroRNAs as novel biomarkers for colorectal cancer: New outlooks. Biomed. Pharm. 2018, 97, 1319–1330. [Google Scholar] [CrossRef]
- Masuda, T.; Hayashi, N.; Kuroda, Y.; Ito, S.; Eguchi, H.; Mimori, K. MicroRNAs as biomarkers in colorectal cancer. Cancers 2017, 9, 124. [Google Scholar] [CrossRef] [PubMed]
- To, K.K.; Tong, C.W.; Wu, M.; Cho, W.C. MicroRNAs in the prognosis and therapy of colorectal cancer: From bench to bedside. World J. Gastroenterol. 2018, 24, 2949–2973. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Silver, J.; Oshlack, A.; Holmes, M.; Diyagama, D.; Holloway, A.; Smyth, G.K. A comparison of background correction methods for two-colour microarrays. Bioinformatics 2007, 23, 2700–2707. [Google Scholar] [CrossRef] [PubMed]
- Falcon, S.; Gentleman, R. Using GOstats to test gene lists for GO term association. Bioinformatics 2007, 23, 257–258. [Google Scholar] [CrossRef] [PubMed]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. In Proceedings of the Third International AAAI Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Group | Age | Staging | Tumour Site | Histologic Grade |
|---|---|---|---|---|---|
| 1 | Control | 51 | NA | NA | NA |
| 2 | <45 Non Lynch | 27 | IIA | Left | - |
| 3 | <45 Non Lynch | 35 | IIA | Right | Well differentiated |
| 4 | Lynch | 46 | - | - | Moderately differentiated |
| 5 | Lynch | 47 | IIIB | Transverse | Moderately differentiated |
| 6 | Lynch | 41 | IIA | Left | Well differentiated |
| 7 | >45 Non Lynch | 70 | IVA | Left | Well differentiated |
| 8 | >45 Non Lynch | 64 | IIIB | Left | Well differentiated |
| miRNA ID | Accession Number | <45 Non Lynch | Lynch | >45 Non Lynch |
|---|---|---|---|---|
| logFC | logFC | logFC | ||
| hsa-miR-1-3p | MIMAT0000416 | 0.02 | −0.22 | 3.13 |
| hsa-miR-10a-5p | MIMAT0000253 | −0.43 | −0.50 | 2.55 |
| hsa-miR-16-5p | MIMAT0000069 | 1.97 | 2.99 | 4.47 |
| hsa-miR-20a-5p | MIMAT0000075 | 0.71 | 1.83 | 3.66 |
| hsa-miR-23b-3p | MIMAT0000418 | 2.83 | 2.96 | 6.05 |
| hsa-miR-24-3p | MIMAT0000080 | 1.82 | 2.31 | 5.15 |
| hsa-miR-27b-3p | MIMAT0000419 | 0.68 | 1.07 | 3.93 |
| hsa-miR-29c-3p | MIMAT0000681 | 1.29 | 1.92 | 4.16 |
| hsa-miR-99a-5p | MIMAT0000097 | −0.54 | −1.46 | 0.38 |
| hsa-miR-143-3p | MIMAT0000435 | 0.17 | −0.35 | 4.59 |
| hsa-miR-145-5p | MIMAT0000437 | −0.03 | −0.02 | 4.45 |
| hsa-miR-193b-5p | MIMAT0004767 | −0.23 | −0.59 | 0.36 |
| hsa-miR-320a | MIMAT0000510 | −1.68 | −1.60 | 0.05 |
| hsa-miR-326 | MIMAT0000756 | −1.38 | −1.89 | −2.89 |
| hsa-miR-642b-3p | MIMAT0018444 | −1.46 | −1.57 | −3.05 |
| hsa-Let7a-5p | MIMAT0000062 | 1.14 | 1.58 | 3.68 |
| SNORD4A | NR_000010.1 | −0.42 | 1.00 | 2.10 |
| Characteristics | Patients |
|---|---|
| Gender (m:f) absolute numbers | 115:77 |
| Lynch syndrome | |
| Lynch | 20 |
| Sporadic | 170 |
| Age, years at diagnosis | |
| Under 50 | 59 |
| Over 50 | 132 |
| Under 60 | 87 |
| Over 60 | 103 |
| Tumour site | |
| Right | 71 |
| Left | 101 |
| Unknown | 20 |
| T stage | |
| Tis | 2 |
| T1 | 1 |
| T2 | 29 |
| T3 | 126 |
| T4 | 27 |
| Tx | 7 |
| N stage | |
| N0 | 80 |
| N1 | 44 |
| N2 | 58 |
| Nx | 10 |
| M stage | |
| M0 | 166 |
| M1 | 16 |
| NS/NC | 10 |
| AJCC staging 7th ed | |
| Stage 0 | 2 |
| Stage I | 25 |
| Stage IIA | 50 |
| Stage IIB | 4 |
| Stage IIC | 2 |
| Stage IIIA | 3 |
| Stage IIIB | 53 |
| Stage IIIC | 29 |
| Stage IVA | 13 |
| Stage IVB | 3 |
| Characteristics | Patients |
|---|---|
| Histologic grade | |
| Well differentiated | 92 |
| Moderately differentiated | 54 |
| Poorly differentiated | 35 |
| Undifferentiated | 0 |
| Unknown | 11 |
| Stroma | |
| Abundant | 33 |
| Moderate | 57 |
| Poor | 59 |
| Unknown | 43 |
| Mucinus component | |
| Mucinus | 17 |
| No mucinus | 142 |
| Unknown | 33 |
| Peritumoral inflammatory infiltrate | |
| Marked | 24 |
| Moderate | 40 |
| Poor | 86 |
| Unknown | 42 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conde, E.; Pascual, A.; Prieto-Cuadra, D.; Laza, V.F.; Molina-Cerrillo, J.; Ramos-Muñoz, M.E.; Rodríguez-Serrano, E.M.; Soto, J.L.; Carrato, A.; García-Bermejo, M.L.; et al. Novel Molecular Characterization of Colorectal Primary Tumors Based on miRNAs. Cancers 2019, 11, 346. https://doi.org/10.3390/cancers11030346
Conde E, Pascual A, Prieto-Cuadra D, Laza VF, Molina-Cerrillo J, Ramos-Muñoz ME, Rodríguez-Serrano EM, Soto JL, Carrato A, García-Bermejo ML, et al. Novel Molecular Characterization of Colorectal Primary Tumors Based on miRNAs. Cancers. 2019; 11(3):346. https://doi.org/10.3390/cancers11030346
Chicago/Turabian StyleConde, Elisa, Alejandro Pascual, Daniel Prieto-Cuadra, Val F. Laza, Javier Molina-Cerrillo, Miren Edurne Ramos-Muñoz, Esperanza Macarena Rodríguez-Serrano, José Luis Soto, Alfredo Carrato, María Laura García-Bermejo, and et al. 2019. "Novel Molecular Characterization of Colorectal Primary Tumors Based on miRNAs" Cancers 11, no. 3: 346. https://doi.org/10.3390/cancers11030346