Natural Bis-Benzylisoquinoline Alkaloids-Tetrandrine, Fangchinoline, and Cepharanthine, Inhibit Human Coronavirus OC43 Infection of MRC-5 Human Lung Cells

 and
and
Abstract
:1. Introduction
2. Materials and Methods
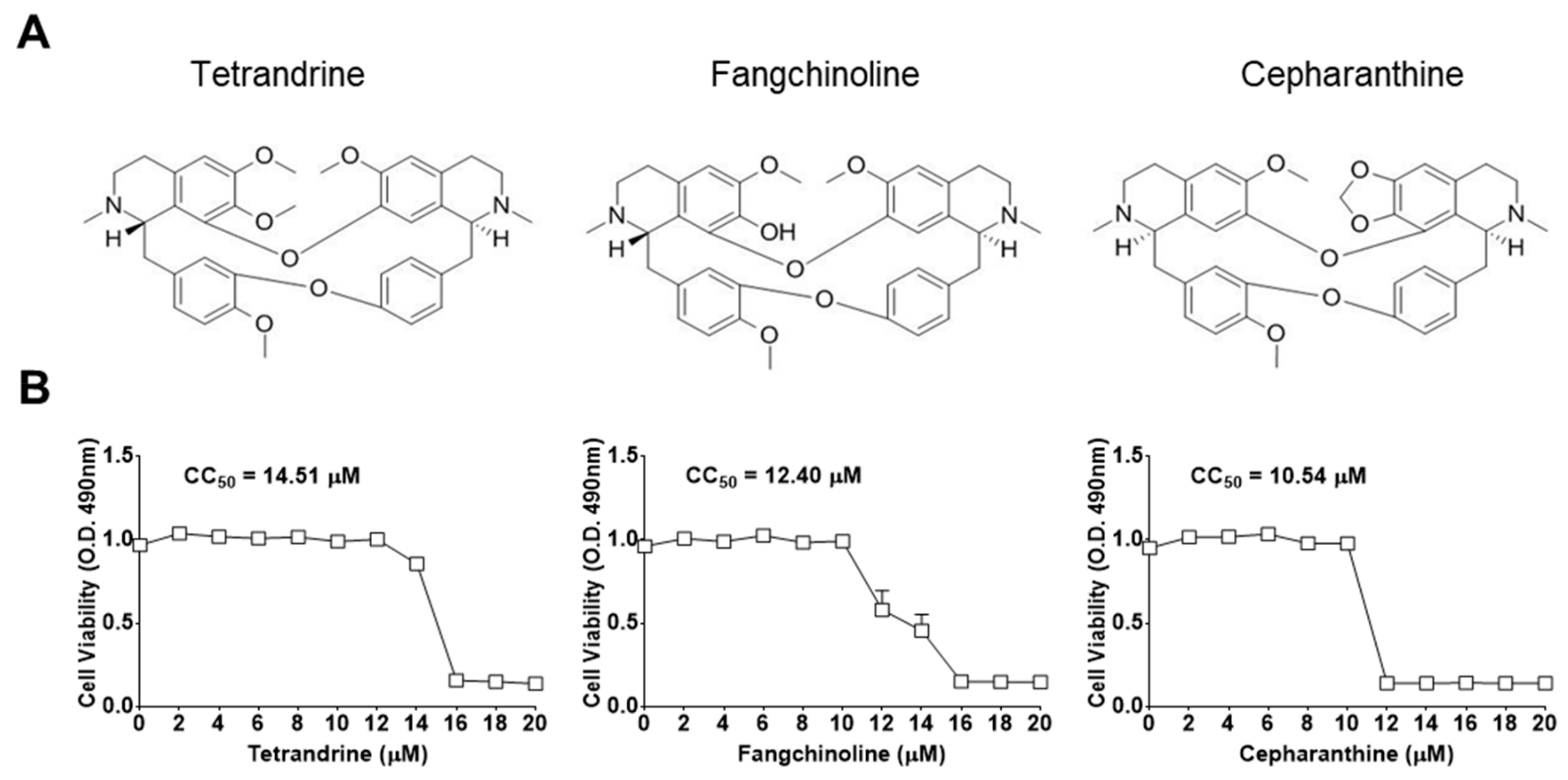
2.1. Preparation of Compounds
2.2. Human Cell Line Culture
2.3. HCoV-OC43 Propagation
2.4. Virus Titration
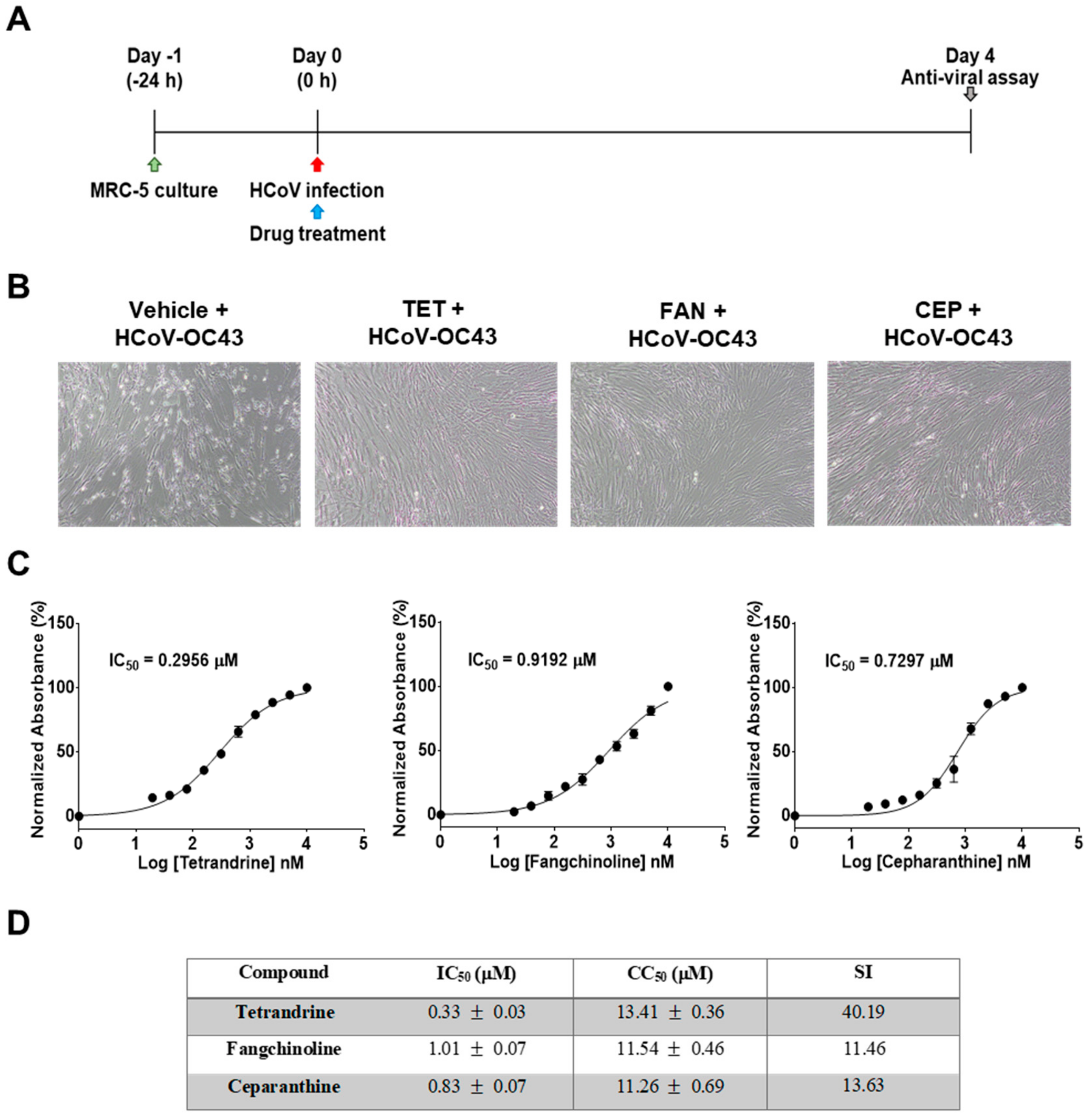
2.5. Determination of the Half Maximal Inhibitory Concentration (IC50) of TET, FAN, and CEP
2.6. Time-of-Addition Assay
2.7. MTS Assay of Cell Viability
2.8. Quantification of the Copy Number of HCoV-OC43 RNA
2.9. Cytokine mRNA Quantification by qRT-PCR
2.10. Western Blot Analysis
2.11. Cytometric Bead Array (CBA) of Cytokines
2.12. Immunohistochemical Analysis
2.13. Statistical Analysis
3. Results
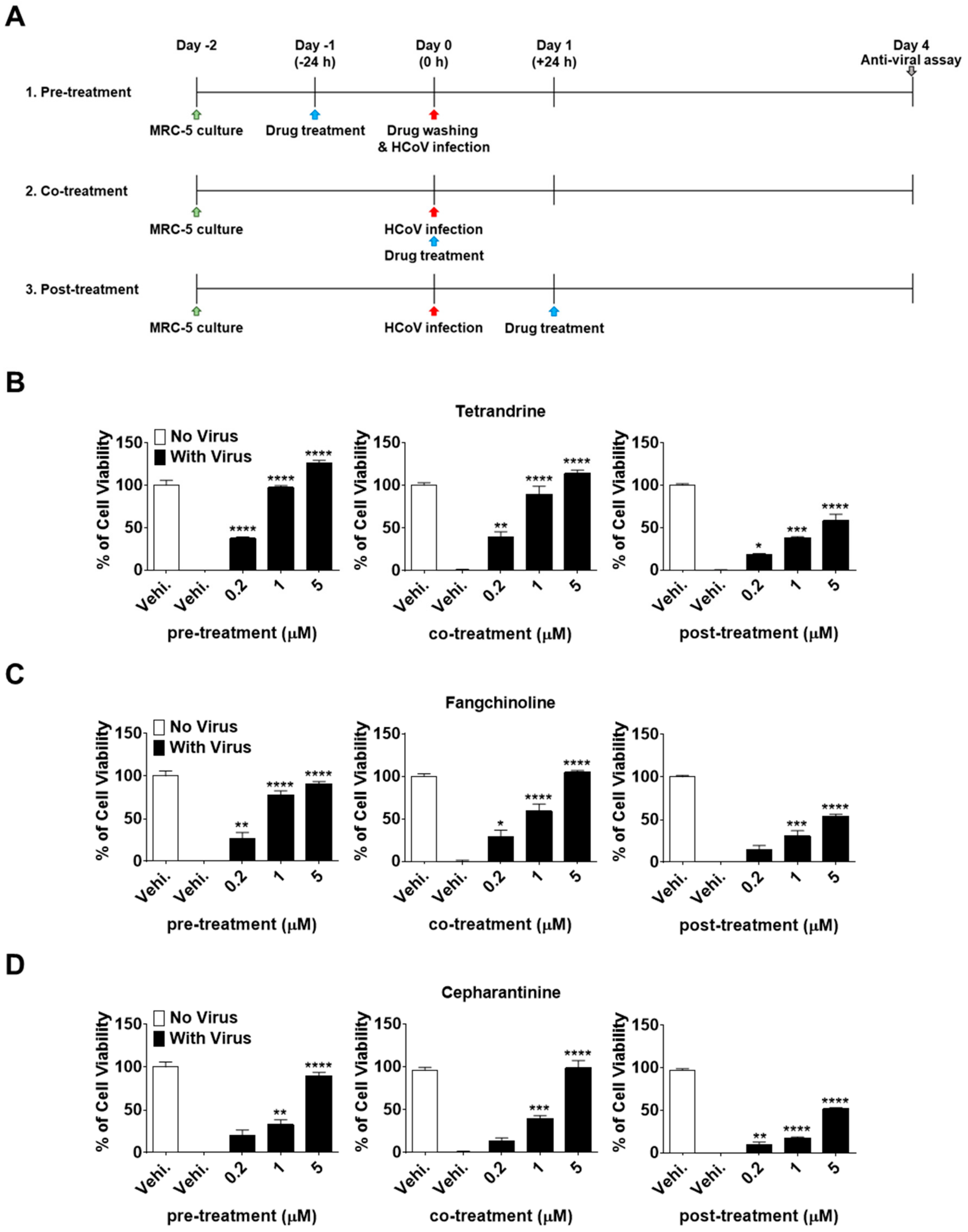
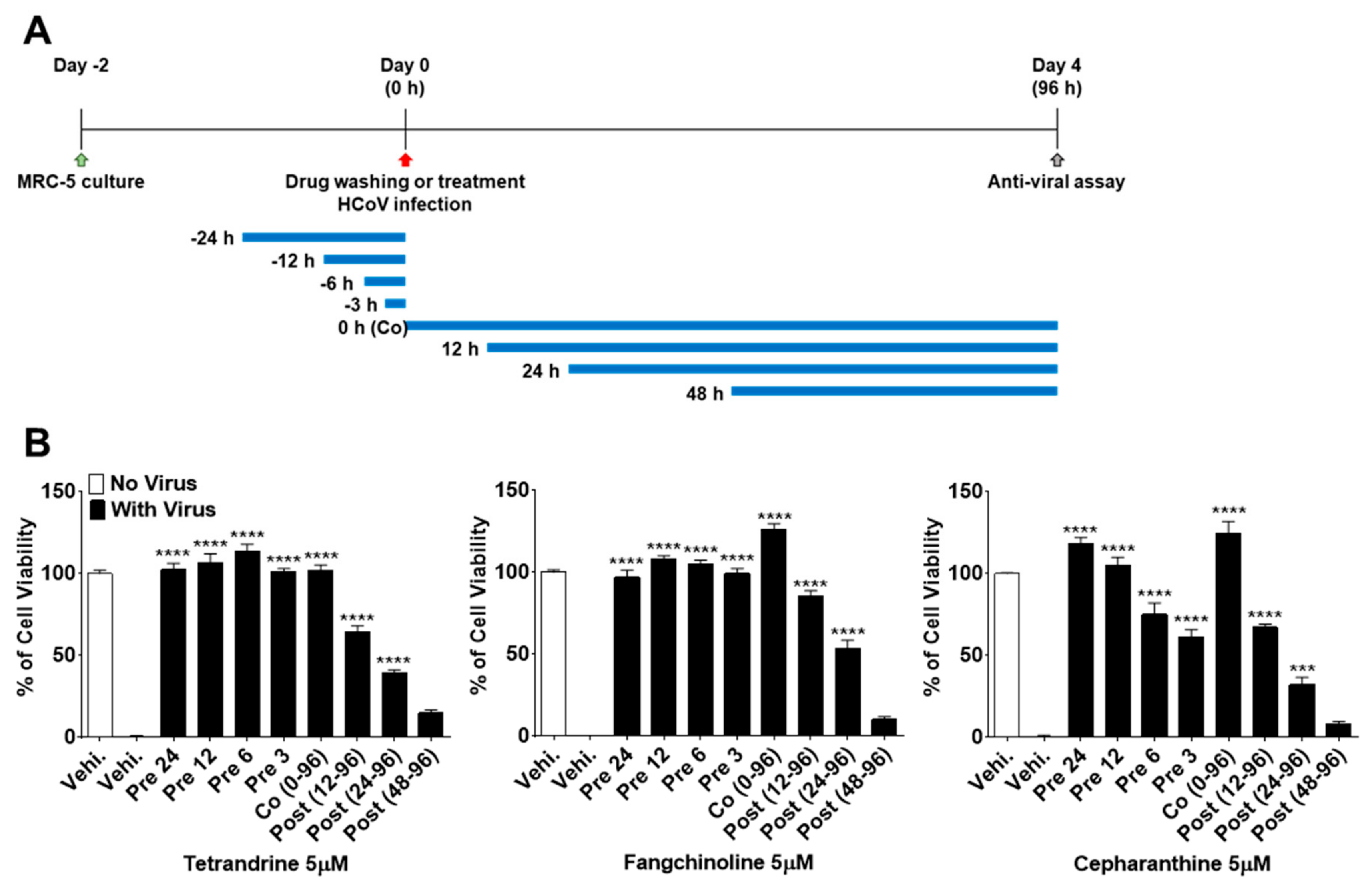
3.1. TET, FAN, and CEP Protected MRC-5 Cells from the Cytopathic Effect of HCoV-OC43
3.2. TEN, FAN, and CEP Have Antiviral Activities at the Early Stage of HCoV-OC43 Infection
3.3. TET, FAN, and CEP Inhibited HCoV-OC43 Replication and N Protein Expression in MRC-5 Cells
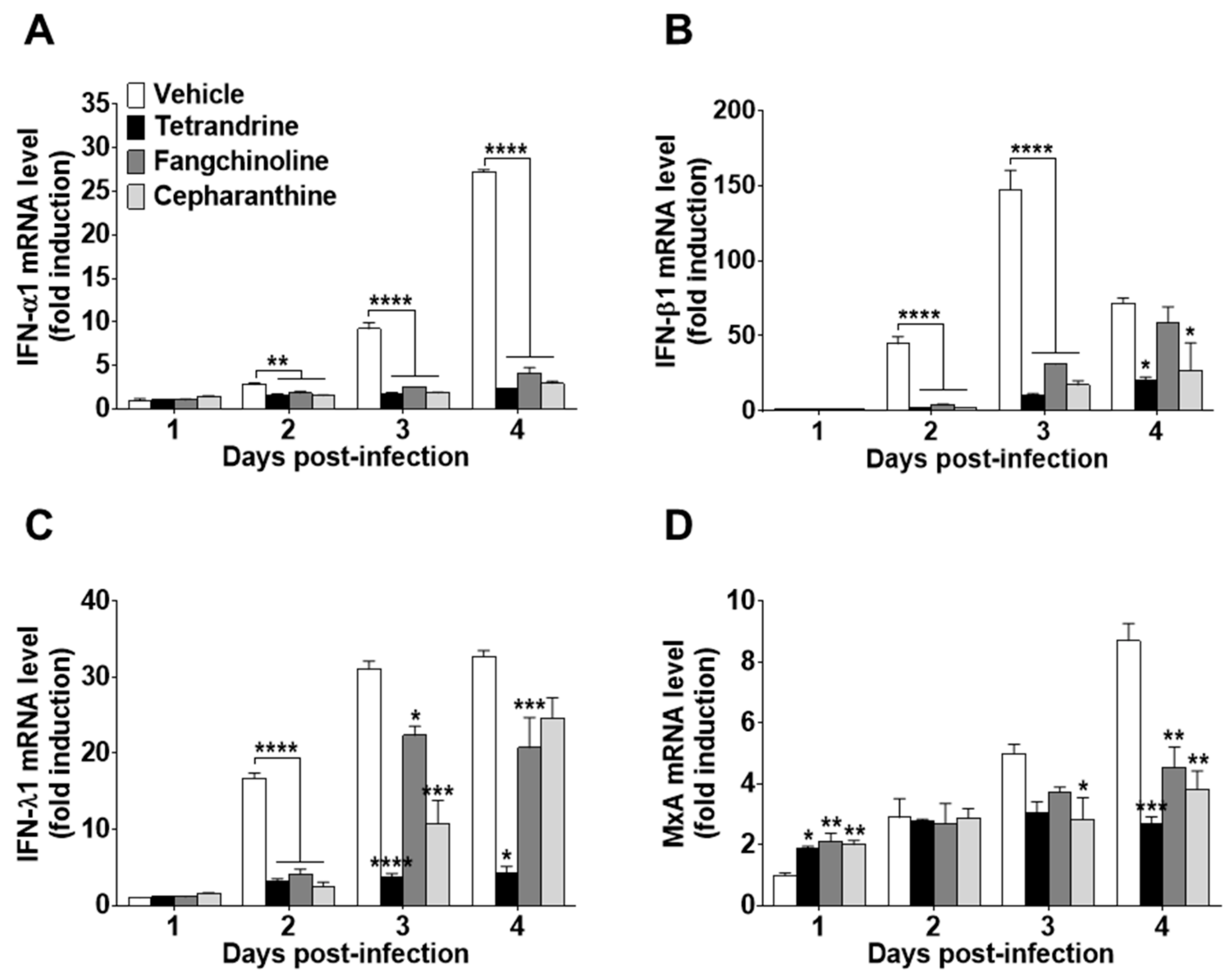
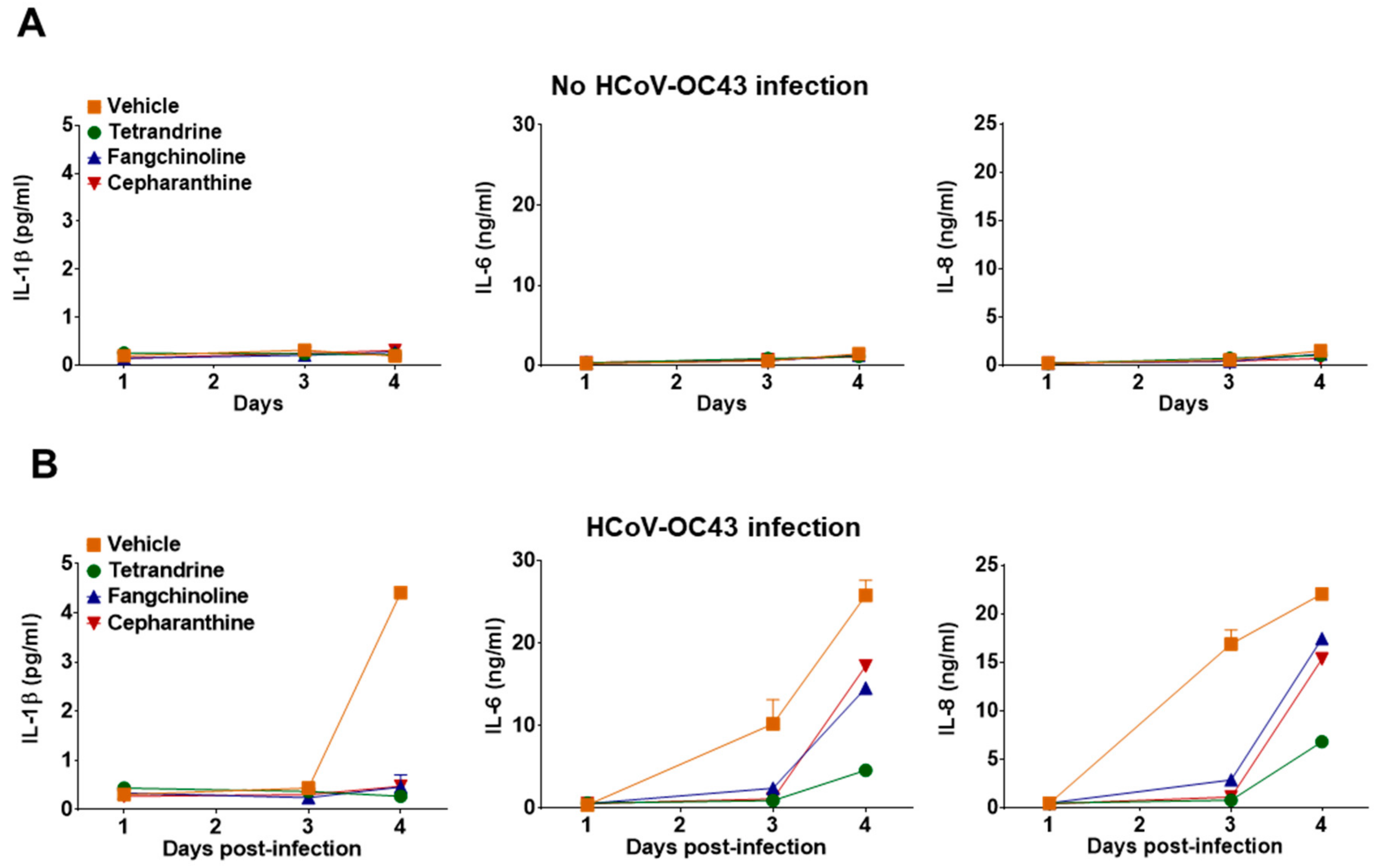
3.4. Antiviral Gene Expression and Inflammatory Cytokine Production Induced by HCoV-OC43 Infection were Reduced in Compound-Treated MRC-5 Cells
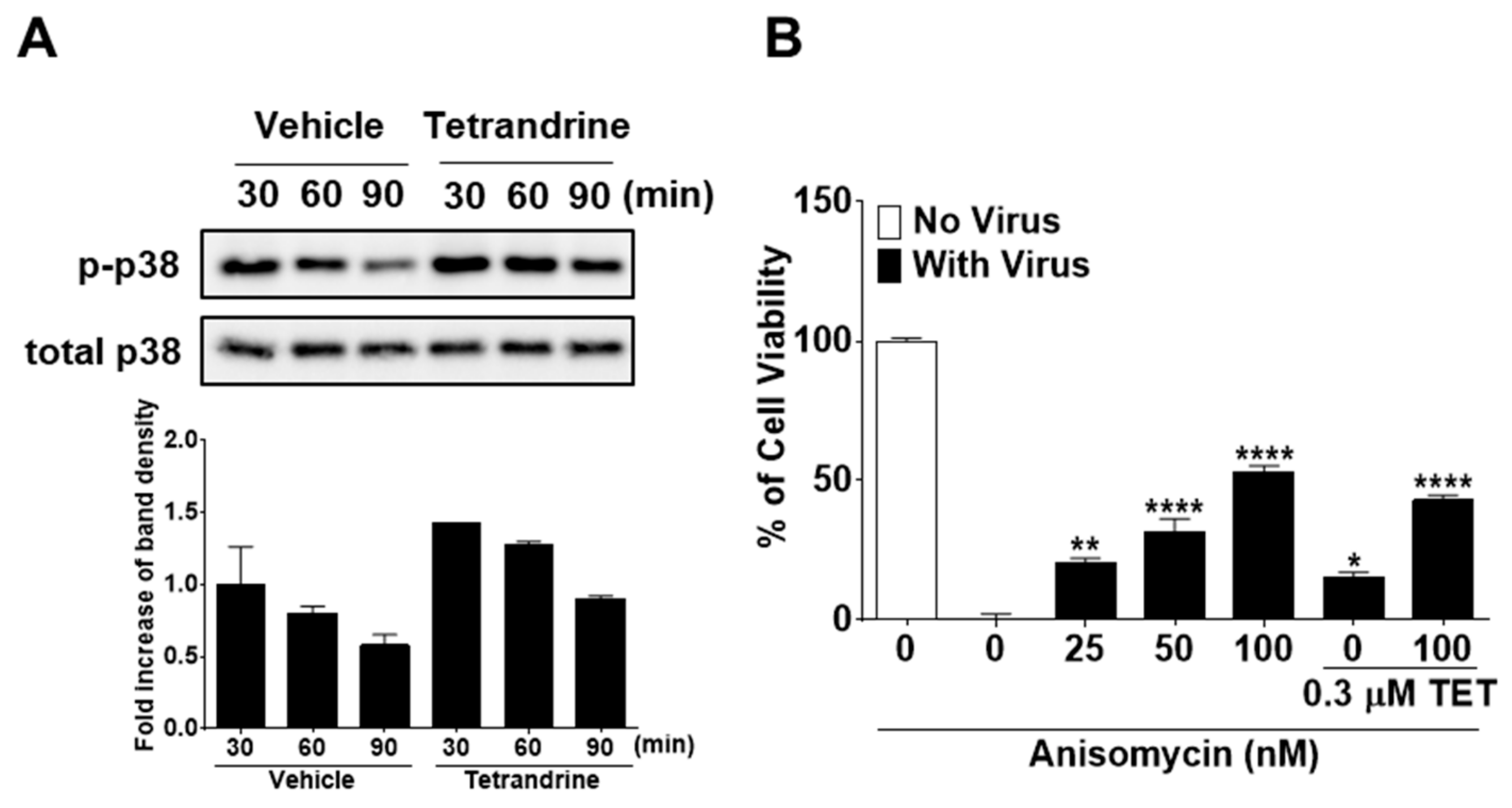
3.5. TET Promoted the Phosphorylation of P38 Mitogen-Activated Protein Kinase (MAPK) in HCoV-OC43-Infected MRC-5 Cells, but Enhanced P38 Phosphorylation by ANM Had no Antiviral Synergistic Effect with TET
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CEP | Cepharanthine |
| Dpi | Days post-infection |
| FAN | Fangchinoline |
| HCoV-OC43 | Human coronavirus OC43 |
| IFN | Interferon |
| IL | Interleukin |
| TET | Tetrandrine |
| TNF | Tumor necrosis factor |
References
- Lin, L.-T.; Hsu, W.-C.; Lin, C.-C. Antiviral Natural Products and Herbal Medicines. J. Tradit. Complement. Med. 2014, 4, 24–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, K.W.; Ernst, E. Antiviral agents from plants and herbs: a systematic review. Antivir. Ther. 2003, 8, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Jassim, S.; Naji, M. Novel antiviral agents: a medicinal plant perspective. J. Appl. Microbiol. 2003, 95, 412–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akram, M.; Tahir, I.M.; Shah, S.M.A.; Mahmood, Z.; Altaf, A.; Ahmad, K.; Munir, N.; Daniyal, M.; Nasir, S.; Mehboob, H. Antiviral potential of medicinal plants against HIV, HSV, influenza, hepatitis, and coxsackievirus: A systematic review. Phytother. Res. 2018, 32, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Bhagya, N.; Chandrashekar, K. Tetrandrine – A molecule of wide bioactivity. Phytochemistry 2016, 125, 5–13. [Google Scholar] [CrossRef]
- Weber, C.; Opatz, T. Bisbenzylisoquinoline Alkaloids. Alkaloids Chem. Biol. 2019, 81, 1–114. [Google Scholar]
- Hu, S.; Dutt, J.; Zhao, T.; Foster, C.S. Tetrandrine potently inhibits herpes simplex virus type-1-induced keratitis in BALB/c mice. Ocul. Immunol. Inflamm. 1997, 5, 173–180. [Google Scholar] [CrossRef]
- Liou, J.-T.; Chen, Z.-Y.; Ho, L.-J.; Yang, S.-P.; Chang, D.-M.; Liang, C.-C.; Lai, J.-H. Differential effects of triptolide and tetrandrine on activation of COX-2, NF-κB, and AP-1 and virus production in dengue virus-infected human lung cells. Eur. J. Pharmacol. 2008, 589, 288–298. [Google Scholar] [CrossRef]
- Sakurai, Y.; Kolokoltsov, A.A.; Chen, C.-C.; Tidwell, M.W.; Bauta, W.E.; Klugbauer, N.; Grimm, C.; Wahl-Schott, C.; Biel, M.; Davey, R.A. Ebola virus. Two-pore channels control Ebola virus host cell entry and are drug targets for disease treatment. Science 2015, 347, 995–998. [Google Scholar] [CrossRef]
- Wan, Z.; Lu, Y.; Liao, Q.; Wu, Y.; Chen, X. Fangchinoline Inhibits Human Immunodeficiency Virus Type 1 Replication by Interfering with gp160 Proteolytic Processing. PLoS ONE 2012, 7, e39225. [Google Scholar] [CrossRef]
- Baba, M.; Okamoto, M.; Kashiwaba, N.; Ono, M. Anti-HIV-1 activity and structure-activity relationship of cepharanoline derivatives in chronically infected cells. Antivir. Chem. Chemother. 2001, 12, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Y.; Zhang, M.; Li, G.; Cen, Y. Study on the inhibitory effect of cepharanthine on herpes simplex type-1 virus (HSV-1) in vitro. Zhong Yao Cai 2004, 27, 107–110. [Google Scholar] [PubMed]
- Lim, Y.X.; Ng, Y.L.; Tam, J.P.; Liu, D.X. Human Coronaviruses: A Review of Virus-Host Interactions. Diseases 2016, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Zumla, A.; Chan, J.F.W.; Azhar, E.I.; Hui, D.S.C.; Yuen, K.-Y.; Chan, J.F.-W. Coronaviruses—drug discovery and therapeutic options. Nat. Rev. Drug Discov. 2016, 15, 327–347. [Google Scholar] [CrossRef]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [Green Version]
- 2018 Annual review of diseases prioritized under the Research and Development Blueprint. Available online: https://www.who.int/emergencies/diseases/2018prioritization-report.pdf?ua=1 (accessed on 21 October 2019).
- Monto, A.S.; Lim, S.K. The Tecumseh Study of Respiratory Illness. VI. Frequency of and Relationship between Outbreaks of Coronavims Infection. J. Infect. Dis. 1974, 129, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Macnaughton, M.R. Occurrence and frequency of coronavirus infections in humans as determined by enzyme-linked immunosorbent assay. Infect. Immun. 1982, 38, 419–423. [Google Scholar] [Green Version]
- Zhang, R.; Wang, K.; Ping, X.; Yu, W.; Qian, Z.; Xiong, S.; Sun, B. The ns12.9 Accessory Protein of Human Coronavirus OC43 Is a Viroporin Involved in Virion Morphogenesis and Pathogenesis. J. Virol. 2015, 89, 11383–11395. [Google Scholar] [CrossRef] [Green Version]
- Myint, S.H. Human coronavirus infections. In The Coronaviridae; Springer: Boston, MA, USA, 1995; pp. 389–401. [Google Scholar]
- Marra, M.A.; Jones, S.J.M.; Astell, C.R.; Holt, R.A.; Brooks-Wilson, A.; Butterfield, Y.S.N.; Khattra, J.; Asano, J.K.; Barber, S.A.; Chan, S.Y.; et al. The Genome Sequence of the SARS-Associated Coronavirus. Science 2003, 300, 1399–1404. [Google Scholar] [CrossRef] [Green Version]
- St-Jean, J.R.; Jacomy, H.; Desforges, M.; Vabret, A.; Freymuth, F.; Talbot, P.J. Human Respiratory Coronavirus OC43: Genetic Stability and Neuroinvasion. J. Virol. 2004, 78, 8824–8834. [Google Scholar] [CrossRef] [Green Version]
- Beidas, M.; Chehadeh, W. PCR array profiling of antiviral genes in human embryonic kidney cells expressing human coronavirus OC43 structural and accessory proteins. Arch. Virol. 2018, 163, 2065–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Dong, W.; Milewska, A.; Golda, A.; Qi, Y.; Zhu, Q.K.; Marasco, W.A.; Baric, R.S.; Sims, A.C.; Pyrc, K.; et al. Human Coronavirus HKU1 Spike Protein Uses O-Acetylated Sialic Acid as an Attachment Receptor Determinant and Employs Hemagglutinin-Esterase Protein as a Receptor-Destroying Enzyme. J. Virol. 2015, 89, 7202–7213. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.-D.; Zhang, R.-X.; Wu, N.; Yuan, W.; Luo, S.-H.; Chen, H.; Liu, Y.; Wang, Y.; He, B.-C.; Deng, Z.-L. Tetrandrine inhibits the proliferation of human osteosarcoma cells by upregulating the PTEN pathway. Oncol. Rep. 2017, 37, 2795–2802. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Wang, X.; Tu, Y.; Masaki, H.; Tanaka, S.; Onda, K.; Sugiyama, K.; Yamada, H.; Hirano, T. Tetrandrine and cepharanthine induce apoptosis through caspase cascade regulation, cell cycle arrest, MAPK activation and PI3K/Akt/mTOR signal modification in glucocorticoid resistant human leukemia Jurkat T cells. Chem. Interact. 2019, 310, 108726. [Google Scholar] [CrossRef]
- Gaunt, E.R.; Hardie, A.; Claas, E.C.J.; Simmonds, P.; Templeton, K.E. Epidemiology and Clinical Presentations of the Four Human Coronaviruses 229E, HKU1, NL63, and OC43 Detected over 3 Years Using a Novel Multiplex Real-Time PCR Method. J. Clin. Microbiol. 2010, 48, 2940–2947. [Google Scholar] [CrossRef] [Green Version]
- Herman, P.; Verlinden, Y.; Breyer, D.; Van Cleemput, E.; Brochier, B.; Sneyers, M.; Snacken, R.; Hermans, P.; Kerkhofs, P.; Liesnard, C.; et al. Biosafety Risk Assessment of the Severe Acute Respiratory Syndrome (SARS) Coronavirus and Containment Measures for the Diagnostic and Research Laboratories. Available online: https://www.sciensano.be/fr/biblio/biosafety-risk-assessment-severe-acute-respiratory-syndrome-sars-coronavirus-and-containment (accessed on 4 November 2019).
- Shahkarami, M.; Yen, C.; Glaser, C.; Xia, D.; Watt, J.; Wadford, D.A. Laboratory Testing for Middle East Respiratory Syndrome Coronavirus, California, USA, 2013–2014. Emerg. Infect. Dis. 2015, 21, 1664–1666. [Google Scholar] [CrossRef]
- Okamoto, M.; Ono, M.; Baba, M. Potent Inhibition of HIV Type 1 Replication by an Antiinflammatory Alkaloid, Cepharanthine, in Chronically Infected Monocytic Cells. AIDS Res. Hum. Retrovir. 1998, 14, 1239–1245. [Google Scholar] [CrossRef]
- Hulswit, R.; De Haan, C.; Bosch, B.-J. Coronavirus Spike Protein and Tropism Changes. Adv. Virus Res. 2016, 96, 29–57. [Google Scholar]
- Matrosovich, M.; Herrler, G.; Klenk, H.D. Sialic Acid Receptors of Viruses. Top. Curr. Chem. 2015, 367, 1–28. [Google Scholar] [CrossRef]
- Hulswit, R.J.G.; Lang, Y.; Bakkers, M.J.G.; Li, W.; Li, Z.; Schouten, A.; Ophorst, B.; Van Kuppeveld, F.J.M.; Boons, G.-J.; Bosch, B.-J.; et al. Human coronaviruses OC43 and HKU1 bind to 9-O-acetylated sialic acids via a conserved receptor-binding site in spike protein domain A. Proc. Natl. Acad. Sci. 2019, 116, 2681–2690. [Google Scholar] [CrossRef]
- Zeng, Q.; Langereis, M.A.; Van Vliet, A.L.W.; Huizinga, E.G.; De Groot, R.J. Structure of coronavirus hemagglutinin-esterase offers insight into corona and influenza virus evolution. Proc. Natl. Acad. Sci. 2008, 105, 9065–9069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoltz, M.; Klingstrom, J. Alpha/Beta Interferon (IFN-α/β)-Independent Induction of IFN-λ1 (Interleukin-29) in Response to Hantaan Virus Infection. J. Virol. 2010, 84, 9140–9148. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Yoshizumi, M.; Ishii, H.; Oishi, K.; Ryo, A. Cytokine production and signaling pathways in respiratory virus infection. Front. Microbiol. 2013, 4, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, E.; Yoshizumi, M.; Tanaka, R.; Ryo, A.; Ishioka, T.; Tsukagoshi, H.; Kozawa, K.; Okayama, Y.; Okabe-Kado, J.; Goya, T.; et al. Cytokine profiles, signalling pathways and effects of fluticasone propionate in respiratory syncytial virus-infected human foetal lung fibroblasts. Cell Boil. Int. 2013, 37, 326–339. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Human) | Sense (5′→3′) | Antisense (5′→3′) | Product Size (bp) |
|---|---|---|---|
| MxA | CAACCTGTGCAGCCAGTATG | GTCCTGCTCCACACCTAGAG | 85 |
| IFN-α1 | GTGCTCAGCTGCAAGTCAAG | TTATCCAGGCTGTGGGTCTC | 65 |
| IFN-β1 | ACCAACAAGTGTCTCCTCCA | GTAGTGGAGAAGCACAACAGG | 50 |
| IFN-λ1 | GTCACCTTCAACCTCTTCCG | TCAGACACAGGTTCCCATCG | 70 |
| SIAE | TATGAACACCGTCTCCACCC | CAACCACAGTGCCATGACAA | 65 |
| β-Actin | GGAAATCGTGCGTGACATCA | ATCTCCTGCTCGAAGTCCAG | 63 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.E.; Min, J.S.; Jang, M.S.; Lee, J.Y.; Shin, Y.S.; Park, C.M.; Song, J.H.; Kim, H.R.; Kim, S.; Jin, Y.-H.; et al. Natural Bis-Benzylisoquinoline Alkaloids-Tetrandrine, Fangchinoline, and Cepharanthine, Inhibit Human Coronavirus OC43 Infection of MRC-5 Human Lung Cells. Biomolecules 2019, 9, 696. https://doi.org/10.3390/biom9110696
Kim DE, Min JS, Jang MS, Lee JY, Shin YS, Park CM, Song JH, Kim HR, Kim S, Jin Y-H, et al. Natural Bis-Benzylisoquinoline Alkaloids-Tetrandrine, Fangchinoline, and Cepharanthine, Inhibit Human Coronavirus OC43 Infection of MRC-5 Human Lung Cells. Biomolecules. 2019; 9(11):696. https://doi.org/10.3390/biom9110696
Chicago/Turabian StyleKim, Dong Eon, Jung Sun Min, Min Seong Jang, Jun Young Lee, Young Sup Shin, Chul Min Park, Jong Hwan Song, Hyoung Rae Kim, Seungtaek Kim, Young-Hee Jin, and et al. 2019. "Natural Bis-Benzylisoquinoline Alkaloids-Tetrandrine, Fangchinoline, and Cepharanthine, Inhibit Human Coronavirus OC43 Infection of MRC-5 Human Lung Cells" Biomolecules 9, no. 11: 696. https://doi.org/10.3390/biom9110696