Comparative Physiological Analysis of Methyl Jasmonate in the Delay of Postharvest Physiological Deterioration and Cell Oxidative Damage in Cassava

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Visual PPD Evaluation
2.3. Assays of ROS Accumulation and Antioxidant Enzyme Activities
2.4. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
2.5. Determination of Endogenous Melatonin and GA
2.6. Quantification of Starch, Soluble Sugar, Ascorbic Acid, and Carotenoid
2.7. Statistical Analysis
3. Results
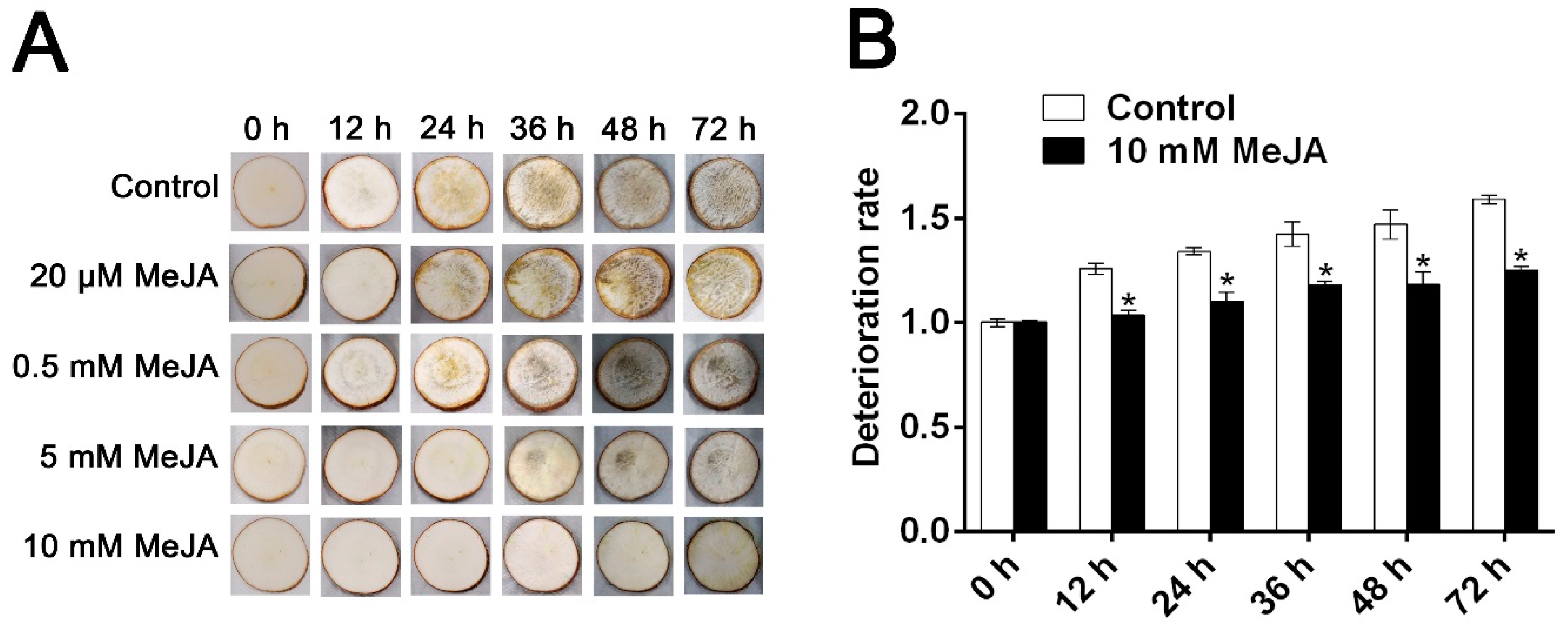
3.1. The Effect of MeJA Treatment on PPD
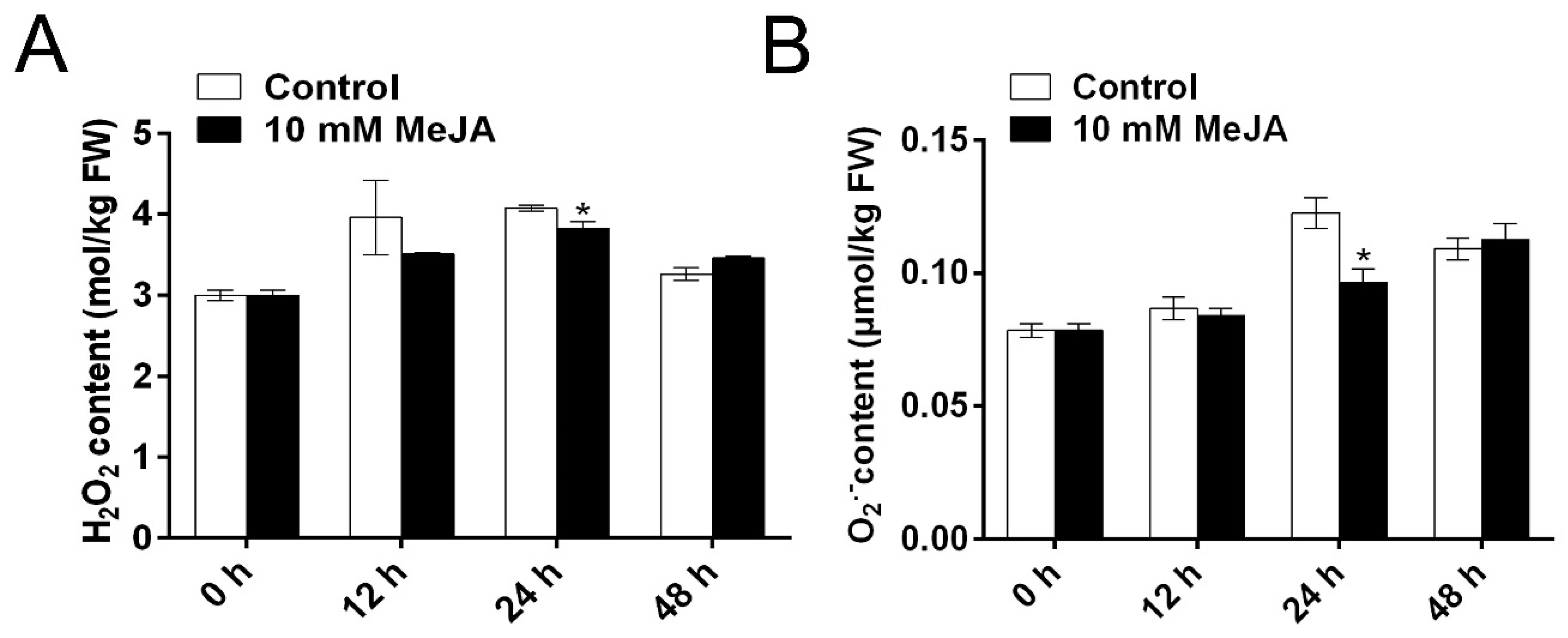
3.2. MeJA Alleviates Cell Oxidative Damage through Modulation of ROS and Underlying Antioxidant Enzymes
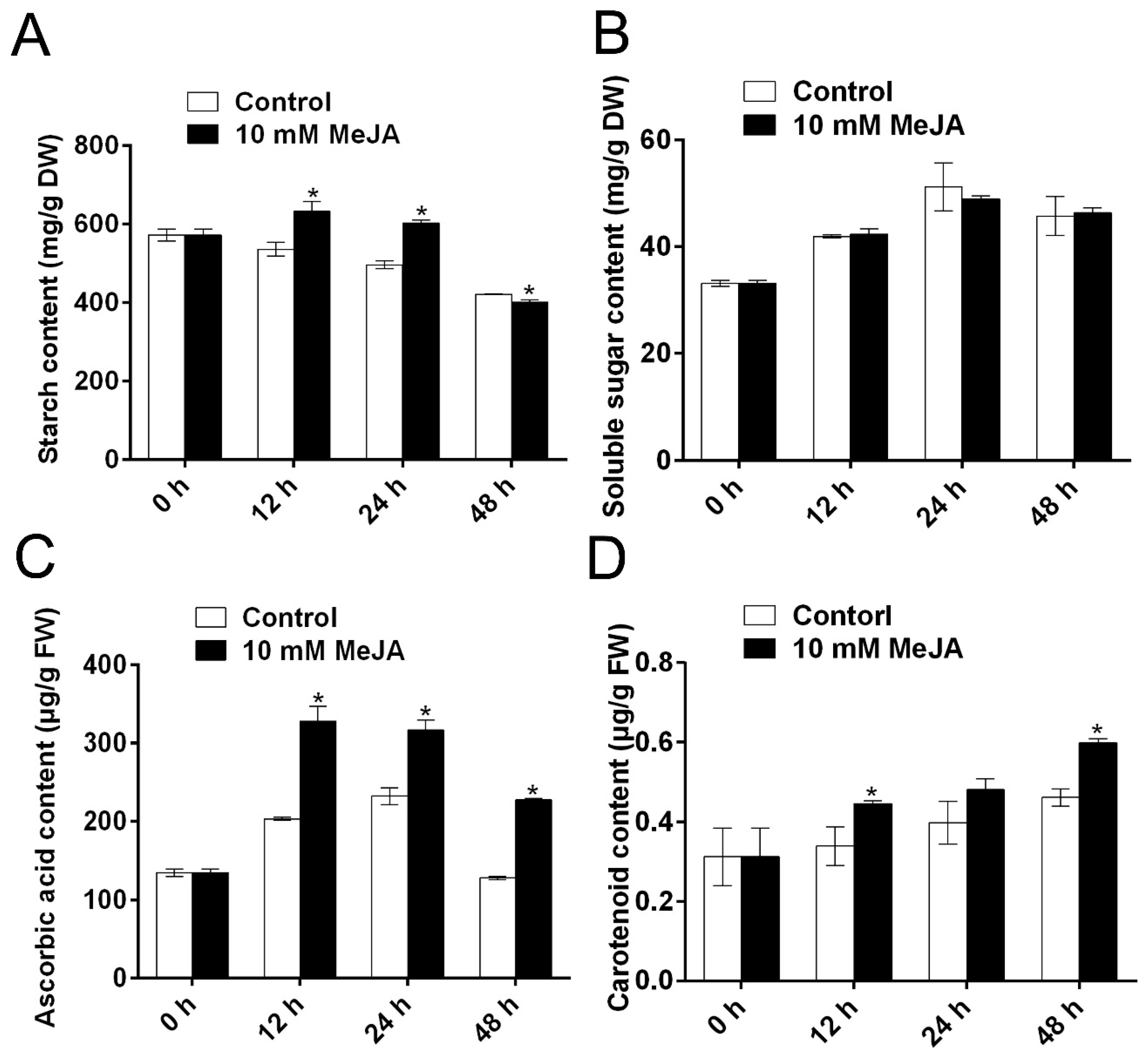
3.3. MeJA Positively Modulates the Quality of Cassava Root Slices
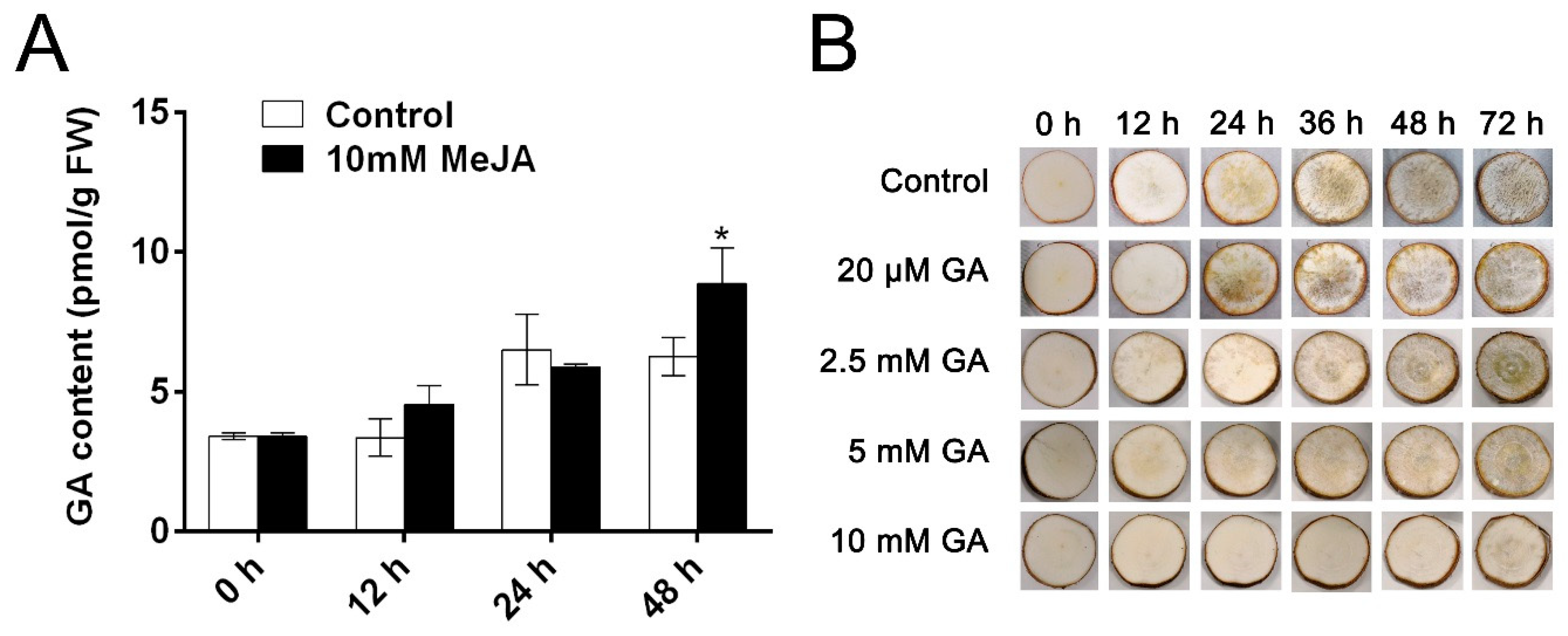
3.4. MeJA Treatment Affects the Endogenous GA Content
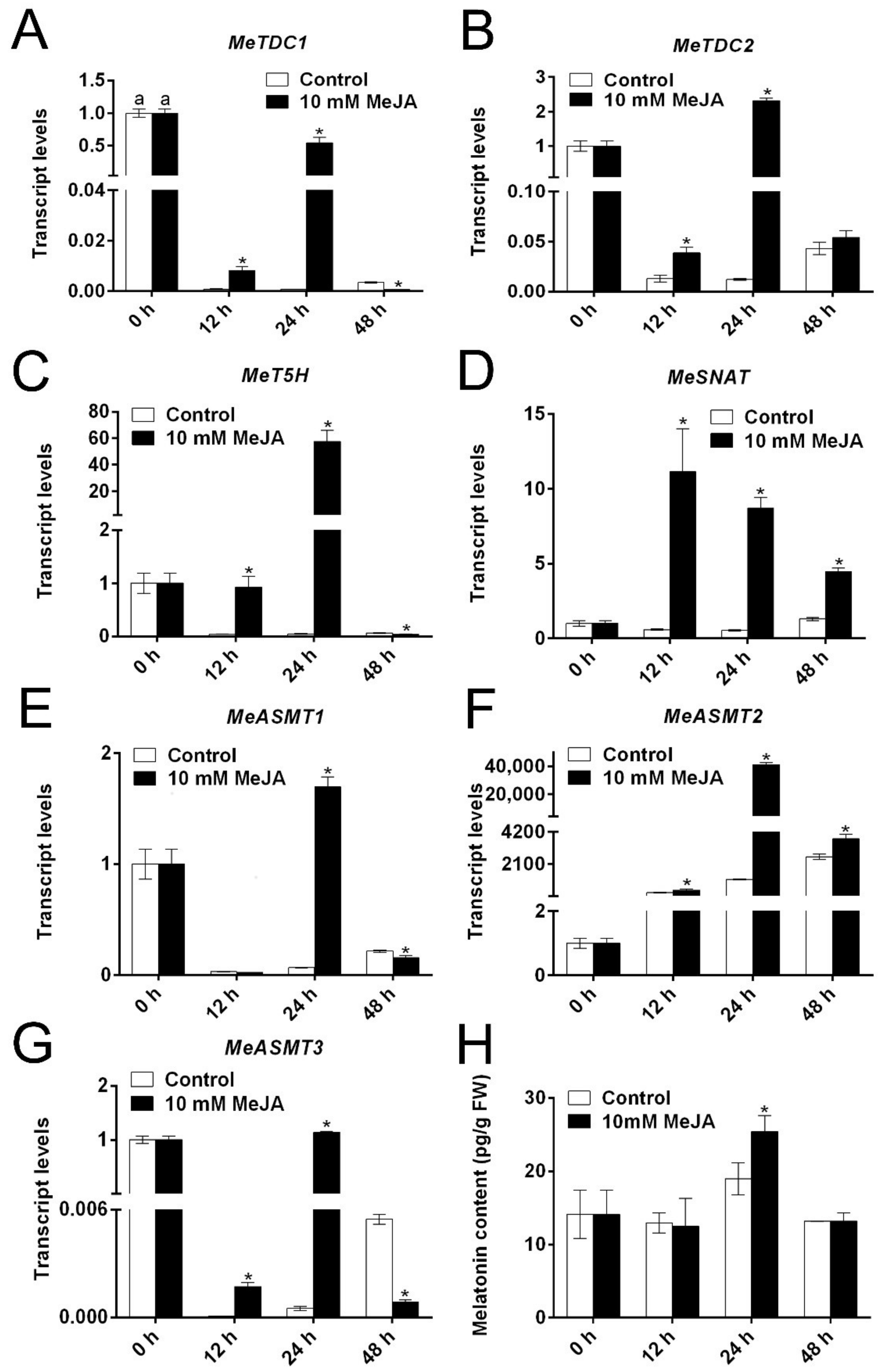
3.5. The Effect of MeJA on the Expression of Melatonin Biosynthesis Genes and Melatonin Level
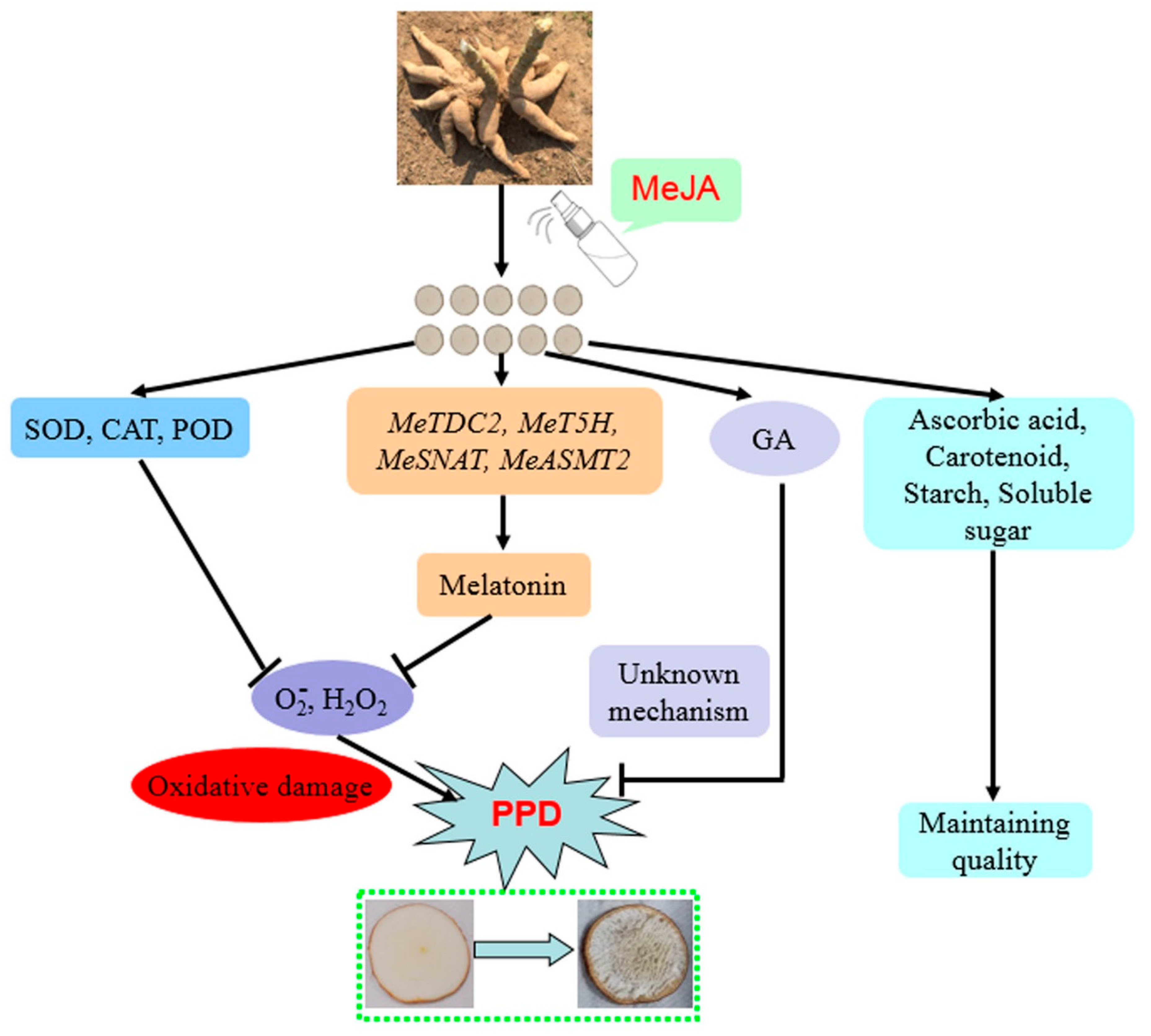
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Morante, N.; Sánchez, T.; Ceballos, H.; Calle, F.; Pérez, J.C.; Egesi, C.; Cuambe, C.E.; Escobar, A.; Ortiz, D.; Chávez, A.L.; et al. Tolerance to postharvest physiological deterioration in cassava roots. Crop Sci. 2010, 50, 1333–1338. [Google Scholar] [CrossRef]
- Iyer, S.; Mattinson, D.S.; Fellman, J.K. Study of the early events leading to cassavaroot postharvest deterioration. Trop. Plant Biol. 2010, 3, 151–165. [Google Scholar] [CrossRef]
- Buschmann, H.; Rodriguez, M.X.; Tohme, J.; Beeching, J.R. Accumulation of hydroxycoumarins during post-harvest deterioration of tuberous roots of cassava (Manihot esculenta crantz). Ann. Bot-London 2000, 86, 1153–1160. [Google Scholar] [CrossRef]
- Salcedo, A.; Siritunga, D. Insights into the physiological, biochemical and molecular basis of postharvest deterioration in cassava (Manihot esculenta) roots. Am. J. Exp. Agric. 2011, 1, 414–431. [Google Scholar] [CrossRef]
- Saravanan, R.; Ravi, V.; Stephen, R.; Thajudhin, S.; George, J. Post-harvest physiological deterioration of cassava (Manihot esculenta) - A review. Indian J. Agr. Sci. 2016, 86, 1383–1390. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Ann. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Hodges, D.M.; Lester, G.E.; Munro, K.D.; Toivonen, P.M.A. Oxidative stress: Importance for postharvest quality. HortScience 2004, 39, 924–929. [Google Scholar] [CrossRef]
- Acedo, J.; Acedo, A. Controlling postharvest physiological deterioration and surface browning in cassava (Manihot esculenta Crantz) roots with hot water treatment. Acta Hortic. 2013, 989, 357–362. [Google Scholar] [CrossRef]
- Sánchez, T.; Chávez, A.L.; Ceballos, H.; Rodriguez-Amaya, D.B.; Nestel, P.; Ishitani, M. Reduction or delay of post-harvest physiological deterioration in cassava roots with higher carotenoid content. J. Sci. Food Agr. 2006, 86, 634–639. [Google Scholar] [CrossRef]
- Ma, Q.; Zhang, T.; Zhang, P.; Wang, Z. Melatonin attenuates postharvest physiological deterioration of cassava storage roots. J. Pineal Res. 2016, 60, 424–434. [Google Scholar] [CrossRef]
- Hu, W.; Kong, H.; Guo, Y.; Zhang, Y.; Ding, Z.; Tie, W.; Yan, Y.; Huang, Q.; Peng, M.; Shi, H.; et al. Comparative physiological and transcriptomic analyses reveal the actions of melatonin in the delay of postharvest physiological deterioration of cassava. Front. Plant Sci. 2016, 7, e0136993. [Google Scholar] [CrossRef]
- Hu, W.; Tie, W.; Ou, W.; Yan, Y.; Kong, H.; Zuo, J.; Ding, X.; Ding, Z.; Liu, Y.; Wu, C.; et al. Crosstalk between calcium and melatonin affects postharvest physiological deterioration and quality loss in cassava. Postharvest Biol. Tec. 2018, 140, 42–49. [Google Scholar] [CrossRef]
- Cortés, D.F.; Reilly, K.; Okogbenin, E.; Beeching, J.R.; Iglesias, C.; Tohme, J. Mapping wound-response genes involved in post-harvest physiological deterioration (PPD) of cassava (Manihot esculenta Crantz). Euphytica 2002, 128, 47–53. [Google Scholar] [CrossRef]
- Tumuhimbise, R.; Melis, R.; Shanahan, P. Genetic variation in cassava for postharvest physiological deterioration. Arch. Agron. Soil Sci. 2015, 61, 1333–1342. [Google Scholar] [CrossRef]
- Fung, R.W.; Wang, C.Y.; Smith, D.L.; Gross, K.C.; Tao, Y.; Tian, M. Characterization of alternative oxidase (AOX) gene expression in response to methyl salicylate and methyl jasmonate pre-treatment and low temperature in tomatoes. J. Plant Physiol. 2006, 163, 1049–1060. [Google Scholar] [CrossRef]
- Karaman, S.; Ozturk, B.; Genc, N.; Celik, S.M. Effect of preharvest application of methyl jasmonate on fruit quality of plum (Prunus salicina Lindell cv. “Fortune”) at harvest and during cold storage. J. Food Proces. Pres. 2013, 37, 1049–1059. [Google Scholar] [CrossRef]
- Wei, J.X.; Wen, X.C.; Tang, L. Effect of methyl jasmonic acid on peach fruit ripening progress. Sci. Hortic-Amsterdam 2017, 220, 206–213. [Google Scholar] [CrossRef]
- Fan, L.L.; Shi, J.Y.; Zuo, J.H.; Gao, L.P.; Lv, J.Y.; Wang, Q. Methyl jasmonate delays postharvest ripening and senescence in the non-climacteric eggplant (Solanum melongena L.) fruit. Postharvest Biol. Tec. 2016, 120, 76–83. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, Y.; Cho, E.; Kwak, S.; Kwon, S.; Bae, J.; Lee, B.; Meen, B.; Huh, G.H. Alterations in intracellular and extracellular activities of antioxidant enzymes during suspension culture of sweetpotato. Phytochemistry 2004, 65, 2471–2484. [Google Scholar] [CrossRef]
- Wang, C.Y.; Buta, J.G. Maintaining quality of fresh-cut kiwifruit with volatile compounds. Postharvest Biol. Tec. 2003, 28, 181–186. [Google Scholar] [CrossRef]
- González-Aguilar, G.A.; Fortiz, J.; Cruz, R.; Baez, R.; Wang, C.Y. Methyl jasmonate reduces chilling injury and maintains postharvest quality of mango fruit. J. Agr. Food Chem. 2000, 48, 515–519. [Google Scholar] [CrossRef]
- Meng, D.M.; Song, T.Z.; Shen, L.; Zhang, X.H.; Sheng, J.P. Postharvest application of methyl jasmonate for improving quality retention of Agaricus bisporus fruit bodies. J. Agr. Food Chem. 2012, 60, 6056–6062. [Google Scholar] [CrossRef]
- Wang, S.Y.; Bowman, L.; Ding, M. Methyl jasmonate enhances antioxidant activity and flavonoid content in blackberries Rubus sp. and promotes antiproliferation of human cancer cells. Food Chem. 2008, 107, 1261–1269. [Google Scholar] [CrossRef]
- Chanjirakul, K.; Wang, S.Y.; Wang, C.H.; Siriphanich, J. Effect of natural volatile compounds on antioxidant capacity and antioxidant enzymes in raspberries. Postharvest Biol. Tec. 2006, 40, 106–115. [Google Scholar] [CrossRef]
- Kondo, S.; Mattheis, J. Aroma volatile biosynthesis in apples at harvest or after harvest affected by jasmonates. Acta Hort. 2006, 712, 381–388. [Google Scholar] [CrossRef]
- Shafiq, M.; Singh, Z.; Khan, A.S. Time of methyl jasmonate application influences the development of ‘Cripps Pink’ apple fruit colour. J. Sci. Food Agr. 2013, 93, 611–618. [Google Scholar] [CrossRef]
- Wang, S.Y.; Zheng, W. Preharvest application of methyl jasmonate increases fruit quality and antioxidant capacity in raspberries. Int. J. Food Sci. Tech. 2005, 40, 187–195. [Google Scholar] [CrossRef]
- Kucuker, E.; Ozturk, B.; Celik, S.M.; Aksit, H. Pre-harvest spray application of methyl jasmonate plays an important role in fruit ripening, fruit quality and bioactive compounds of Japanese plums. Sci. Hortic-Amsterdam 2014, 176, 162–169. [Google Scholar] [CrossRef]
- Wei, Y.; Chang, Y.; Zeng, H.; Liu, G.; He, C.; Shi, H. RAV transcription factors are essential for disease resistance against cassava bacterial blight via activation of melatonin biosynthesis genes. J. Pineal Res. 2018, 64, e12454. [Google Scholar] [CrossRef]
- Wei, Y.; Hu, W.; Wang, Q.; Liu, W.; Wu, C.; Zeng, H.; Yan, Y.; Li, X.; He, C.; Shi, H. Comprehensive transcriptional and functional analyses of melatonin synthesis genes in cassava reveal their novel role in hypersensitive-like cell death. Sci. Rep. 2016, 6, 35029. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.F. Experimental Guidance Plant Physiology; Higher Education Press: Beijing, China, 2007; pp. 145–225. [Google Scholar]
- Cao, J.K.; Jiang, W.B.; Zhao, Y.M. Experiment Guidance of Postharvest Fruits and Vegetables Physiology and Biochemistry; Light Industry Press: Beijing, China, 2007. [Google Scholar]
- Duan, J.J.; Li, J.; Guo, S.; Kang, Y. Exogenous spermidine affects polyamine metabolism in salinity-stressed Cucumis sativus roots and enhances short-term salinity. J. Plant Physiol. 2008, 165, 1620–1635. [Google Scholar] [CrossRef]
- Rider, J.E.; Hacker, A.; Mackintosh, C.A.; Pegg, A.E.; Woster, P.M.; Casero, J. Spermine and spermidine mediate protection against oxidative damage caused by hydrogen peroxide. Amino Acids 2007, 33, 231–240. [Google Scholar] [CrossRef]
- Anjum, S.A.; Wang, L.; Farooq, M.; Khan, I.; Xue, L. Methyl jasmonate-induced alteration in lipid peroxidation, antioxidative defence system and yield in soybean under drought. J. Agron. Crop Sci. 2011, 197, 296–301. [Google Scholar] [CrossRef]
- Chavez, A.L.; Bedoya, J.M.; Sánchez, T.; Iglesias, C.; Ceballos, H.; Roca, W. Iron, carotene, and ascorbic acid in cassava roots and leaves. Food Nutr. Bull. 2000, 21, 410–413. [Google Scholar] [CrossRef]
- Fischer, C.; Holl, W. Food Reserves in Scots Pine (Pinus sylvestris L.). 1: Seasonal Changes in the Carbohydrate and Fat Reserves of Pine Needles. Trees 1991, 6, 147–155. [Google Scholar] [CrossRef]
- Ketabchi, S.; Shahrtash, M. Effects of methyl jasmonate and cytokinin on biochemical responses of maize seedlings infected by Fusarium moniliforme. Asian J. Exp. Biol. Sci. 2011, 2, 299–305. [Google Scholar]
- Mellidou, I.; Keulemans, J.; Davey, M.W.; Kanellis, A. Ascorbic acid accumulation, biosynthesis and recycling during tomato fruit ripening. Acta Hortic. 2012, 939, 255–261. [Google Scholar] [CrossRef]
- Dagar, A.; Weksler, A.; Friedman, H.; Lurie, S. Gibberellic acid (GA3) application at the end of pit ripening: Effect on ripening and storage of two harvests of ‘september snow’ peach. Sci. Hortic-Amsterdam 2012, 140, 125–130. [Google Scholar] [CrossRef]
- Panigrahi, J.; Gheewala, B.; Patel, M.; Patel, N.; Gantait, S. Gibberellic acid coating: A novel approach to expand the shelf-life in green chilli (Capsicum annuum L.). Sci. Hortic-Amsterdam 2017, 225, 581–588. [Google Scholar] [CrossRef]
- Huang, H.; Jing, G.X.; Wang, H.; Duan, X.W.; Qu, H.X.; Jiang, Y.M. The combined effects of phenylurea and gibberellins on quality maintenance and shelf life extension of banana fruit during storage. Sci. Hortic-Amsterdam 2014, 167, 36–42. [Google Scholar] [CrossRef]
- Zhang, Y.; Huber, D.; Hu, M.; Jiang, G.; Gao, Z.; Xu, X.; Jiang, Y.; Zhang, Z. Melatonin delay of postharvest browning in litchi fruit by enhancing anti-oxidative processes and oxidation repair. J. Agr. Food Chem. 2018. [Google Scholar] [CrossRef]
- Hu, W.; Yang, H.; Tie, W.; Yan, Y.; Ding, Z.; Liu, Y.; Wu, C.; Wang, J.; Reiter, R.; Tan, D.; et al. Natural variation in banana varieties highlights the role of melatonin in postharvest ripening and quality. J. Agr. Food Chem. 2017, 65, 9987–9994. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, Z.; Chai, H.; Cheng, N.; Yang, Y.; Wang, D.; Yang, T.; Cao, W. Melatonin treatment delays postharvest senescence and regulates reactive oxygen species metabolism in peach fruit. Postharvest Biol. Tec. 2016, 118, 103–110. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Fard, J.R. Melatonin treatment attenuates postharvest decay and maintains nutritional quality of strawberry fruits (Fragaria × anannasa cv. Selva) by enhancing GABA shunt activity. Food Chem. 2017, 221, 1650–1657. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Li, B.; Li, X.; Wei, Y.; Liu, D.; Shi, H. Comparative Physiological Analysis of Methyl Jasmonate in the Delay of Postharvest Physiological Deterioration and Cell Oxidative Damage in Cassava. Biomolecules 2019, 9, 451. https://doi.org/10.3390/biom9090451
Liu G, Li B, Li X, Wei Y, Liu D, Shi H. Comparative Physiological Analysis of Methyl Jasmonate in the Delay of Postharvest Physiological Deterioration and Cell Oxidative Damage in Cassava. Biomolecules. 2019; 9(9):451. https://doi.org/10.3390/biom9090451
Chicago/Turabian StyleLiu, Guoyin, Bing Li, Xiuqiong Li, Yunxie Wei, Debing Liu, and Haitao Shi. 2019. "Comparative Physiological Analysis of Methyl Jasmonate in the Delay of Postharvest Physiological Deterioration and Cell Oxidative Damage in Cassava" Biomolecules 9, no. 9: 451. https://doi.org/10.3390/biom9090451