
Gold Nanoparticles: Multifaceted Roles in the Management of Autoimmune Disorders


 , and
, and
Abstract
:1. Introduction
2. Pathogenesis of Autoimmune Diseases
3. The Role of Inflammation and Inflammatory Cytokines in ADs
4. Clinical Management of Autoimmune Disorders
5. Nanotechnology and Autoimmune Diseases
6. Gold Nanoparticles, Characterization, and Immune Stimulation
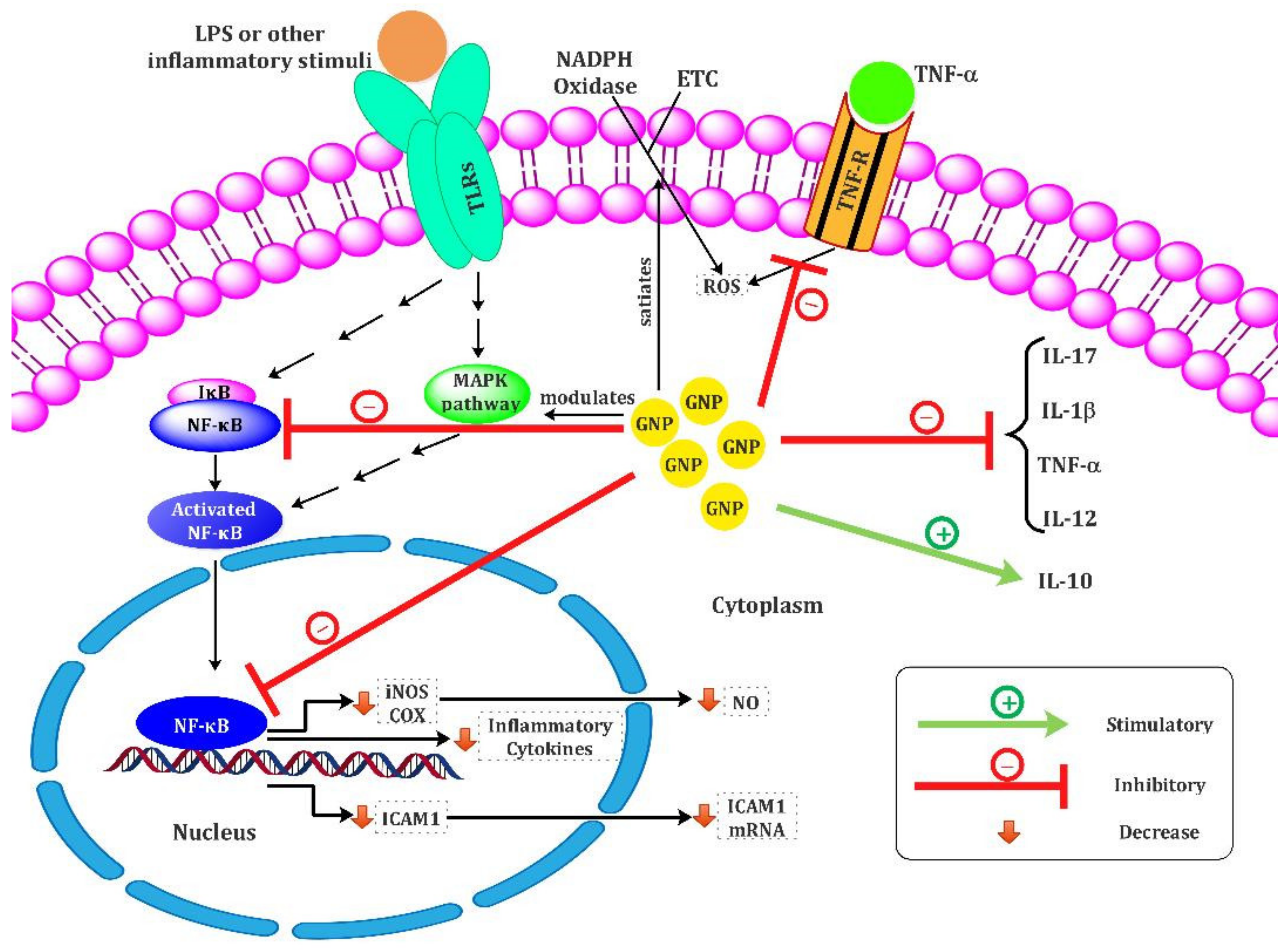
7. Gold Nanoparticles for Immune Stimulation; Focus on Anti-Inflammatory and Antioxidant Properties
8. Impacts of Surface Modification of GNPs on Immune Response
9. GNPs Engineering for Therapeutic Application and Drug Delivery in Biomedical Use
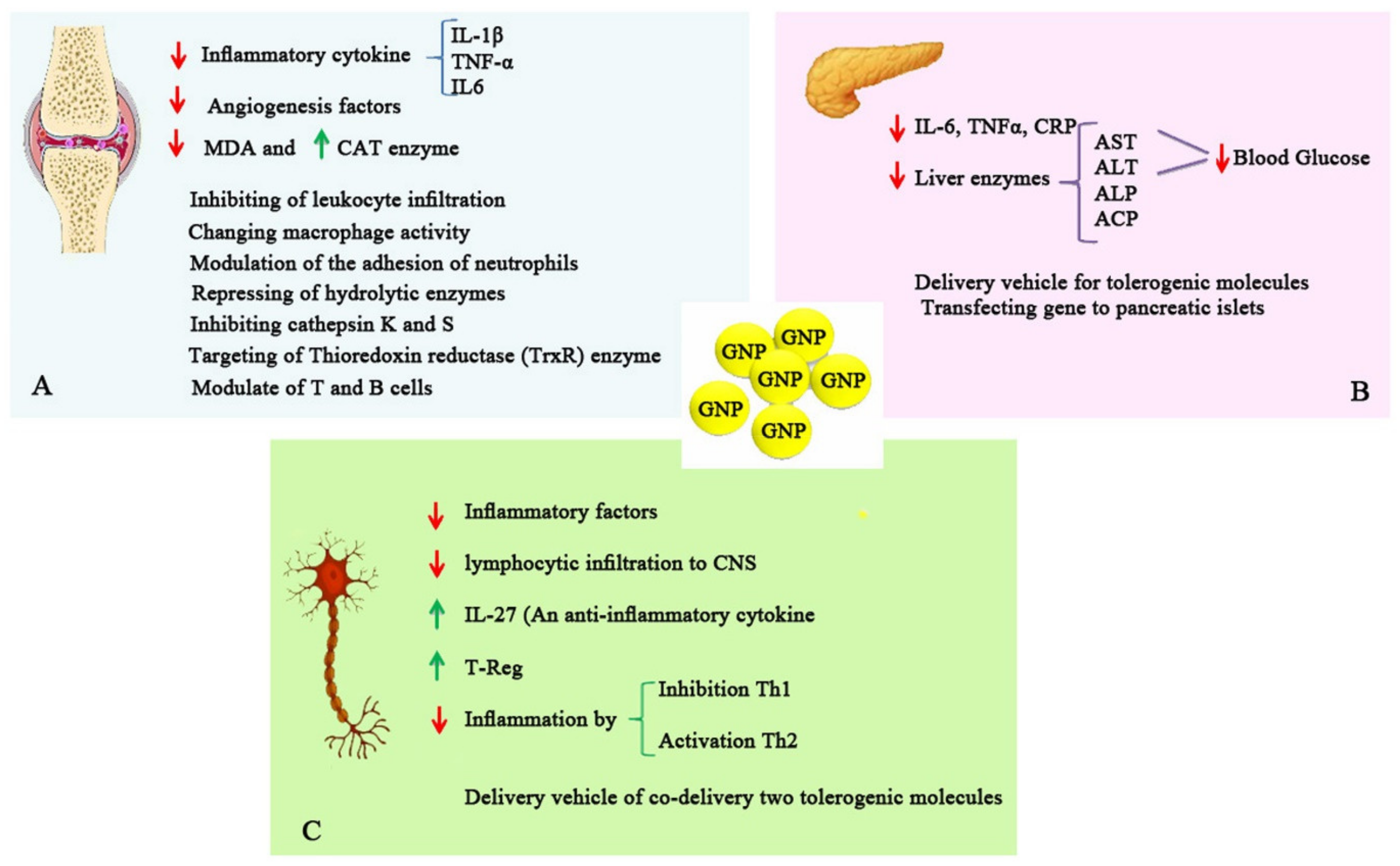
10. Application of GNPs in the Management of Autoimmune Disorders
Application of GNPs for Treatment and Drug Delivery in Rheumatoid Arthritis
11. Application of GNPs in Diabetes and T1DM
12. Application of GNPs in Multiple Sclerosis
13. Application of GNPs in Other Autoimmune Diseases
14. Gold Nanoparticles as Potential Diagnostic Devices for Autoimmune Diseases
15. Conclusions
Funding
Conflicts of Interest
References
- Davidson, A.; Diamond, B. Autoimmune diseases. N. Engl. J. Med. 2001, 345, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Hayter, S.M.; Cook, M.C. Updated assessment of the prevalence, spectrum and case definition of autoimmune disease. Autoimmun. Rev. 2012, 11, 754–765. [Google Scholar] [CrossRef]
- Rook, G.A. Hygiene hypothesis and autoimmune diseases. Clin. Rev. Allergy Immunol. 2012, 42, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.; Diamond, B. General features of autoimmune disease. In The Autoimmune Diseases; Elsevier: Amsterdam, The Netherlands, 2020; pp. 17–44. [Google Scholar]
- Lerner, A.; Jeremias, P.; Matthias, T. The world incidence and prevalence of autoimmune diseases is increasing. Int. J. Celiac Dis. 2015, 3, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zambrano, A.; Lin, Z.-T.; Xing, Y.; Rippy, J.; Wu, T. Immunosensors for biomarker detection in autoimmune diseases. Arch. Immunol. Ther. Exp. 2017, 65, 111–121. [Google Scholar] [CrossRef]
- Cooper, G.S.; Bynum, M.L.; Somers, E.C. Recent insights in the epidemiology of autoimmune diseases: Improved prevalence estimates and understanding of clustering of diseases. J. Autoimmun. 2009, 33, 197–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrie, R.A.; Reider, N.; Cohen, J.; Stuve, O.; Sorensen, P.S.; Cutter, G.; Reingold, S.C.; Trojano, M. A systematic review of the incidence and prevalence of autoimmune disease in multiple sclerosis. Mult. Scler. J. 2015, 21, 282–293. [Google Scholar] [CrossRef] [Green Version]
- National Institute of Allergy and Infectious Diseases, Autoimmune Diseases. Available online: https://www.niaid.nih.gov/diseases-conditions/autoimmune-diseases (accessed on 7 December 2018).
- AARDA. The Cost Burden of Autoimmune Disease: The Latest Front in the War on Healthcare Spending. 2011. Available online: http://www.aarda.org/pdf/cbad.pdf (accessed on 28 March 2011).
- Mackern-Oberti, J.P.; Llanos, C.; Vega, F.; Salazar-Onfray, F.; Riedel, C.A.; Bueno, S.M.; Kalergis, A.M. Role of dendritic cells in the initiation, progress and modulation of systemic autoimmune diseases. Autoimmun. Rev. 2015, 14, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Probst, H.C.; Muth, S.; Schild, H. Regulation of the tolerogenic function of steady-state DCs. Eur. J. Immunol. 2014, 44, 927–933. [Google Scholar] [CrossRef]
- Hawiger, D.; Inaba, K.; Dorsett, Y.; Guo, M.; Mahnke, K.; Rivera, M.; Ravetch, J.V.; Steinman, R.M.; Nussenzweig, M.C. Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo. J. Exp. Med. 2001, 194, 769–780. [Google Scholar] [CrossRef] [Green Version]
- Steinman, R.M.; Hawiger, D.; Nussenzweig, M.C. Tolerogenic dendritic cells. Annu. Rev. Immunol. 2003, 21, 685–711. [Google Scholar] [CrossRef] [Green Version]
- Wofsy, D.; Ledbetter, J.A.; Hendler, P.L.; Seaman, W.E. Treatment of murine lupus with monoclonal anti-T cell antibody. J. Immunol. 1985, 134, 852–857. [Google Scholar] [PubMed]
- Hang, L.; Theofilopoulos, A.N.; Balderas, R.S.; Francis, S.J.; Dixon, F.J. The effect of thymectomy on lupus-prone mice. J. Immunol. 1984, 132, 1809–1813. [Google Scholar]
- Peng, S.L.; Madaio, M.P.; Hayday, A.C.; Craft, J. Propagation and regulation of systemic autoimmunity by gammadelta T cells. J. Immunol. 1996, 157, 5689–5698. [Google Scholar] [PubMed]
- Chan, O.; Shlomchik, M.J. A new role for B cells in systemic autoimmunity: B cells promote spontaneous T cell activation in MRL-lpr/lpr mice. J. Immunol. 1998, 160, 51–59. [Google Scholar]
- Liu, Y.; Wu, J.; Zhu, Y.; Han, J. Therapeutic application of mesenchymal stem cells in bone and joint diseases. Clin. Exp. Med. 2014, 14, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Katsarou, A.; Gudbjörnsdottir, S.; Rawshani, A.; Dabelea, D.; Bonifacio, E.; Anderson, B.J.; Jacobsen, L.M.; Schatz, D.A.; Lernmark, Å. Type 1 diabetes mellitus. Nat. Rev. Dis. Primers 2017, 3, 1–17. [Google Scholar] [CrossRef]
- Asadzadeh-Aghdaei, H.; Mashayekhi, K.; Koushki, K.; Azimzadeh, P.; Rostami-Nejad, M.; Amani, D.; Chaleshi, V.; Haftcheshmeh, S.M.; Sahebkar, A.; Zali, M.R. V617F-independent upregulation of JAK2 gene expression in patients with inflammatory bowel disease. J. Cell. Biochem. 2019, 120, 15746–15755. [Google Scholar] [CrossRef] [PubMed]
- Bolon, B. Cellular and molecular mechanisms of autoimmune disease. Toxicol. Pathol. 2012, 40, 216–229. [Google Scholar] [CrossRef] [Green Version]
- Chwastiak, L.; Ehde, D.M.; Gibbons, L.E.; Sullivan, M.; Bowen, J.D.; Kraft, G.H. Depressive symptoms and severity of illness in multiple sclerosis: Epidemiologic study of a large community sample. Am. J. Psychiatry 2002, 159, 1862–1868. [Google Scholar] [CrossRef]
- Chen, Z.; Bozec, A.; Ramming, A.; Schett, G. Anti-inflammatory and immune-regulatory cytokines in rheumatoid arthritis. Nat. Rev. Rheumatol. 2019, 15, 9–17. [Google Scholar] [CrossRef]
- Nikoopour, E.; Schwartz, J.A.; Singh, B. Therapeutic benefits of regulating inflammation in autoimmunity. Inflamm. Allergy-Drug Targets (Former. Curr. Drug Targets-Inflamm. Allergy) 2008, 7, 203–210. [Google Scholar] [CrossRef]
- Shen, H.-H.; Yang, Y.-X.; Meng, X.; Luo, X.-Y.; Li, X.-M.; Shuai, Z.-W.; Ye, D.-Q.; Pan, H.-F. NLRP3: A promising therapeutic target for autoimmune diseases. Autoimmun. Rev. 2018, 17, 694–702. [Google Scholar] [CrossRef]
- Shlomchik, M.J.; Craft, J.E.; Mamula, M.J. From T to B and back again: Positive feedback in systemic autoimmune disease. Nat. Rev. Immunol. 2001, 1, 147–153. [Google Scholar]
- Mosmann, T.R.; Schumacher, J.H.; Street, N.F.; Budd, R.; O’garra, A.; Fong, T.A.T.; Bind, M.W.; Moore, K.W.M.; Sher, A.; Fiorentino, D.F. Diversity of cytokine synthesis and function of mouse CD4+ T cells. Immunol. Rev. 1991, 123, 209–229. [Google Scholar] [CrossRef]
- Smith, D.A.; Germolec, D.R. Introduction to immunology and autoimmunity. Environ. Health Perspect. 1999, 107, 661–665. [Google Scholar]
- Fairweather, D.; Cihakova, D. Alternatively activated macrophages in infection and autoimmunity. J. Autoimmun. 2009, 33, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Wilde, B.; Thewissen, M.; Damoiseaux, J.; van Paassen, P.; Witzke, O.; Tervaert, J.W.C. T cells in ANCA-associated vasculitis: What can we learn from lesional versus circulating T cells? Arthritis Res. Ther. 2010, 12, 204. [Google Scholar] [CrossRef] [Green Version]
- Dardalhon, V.; Korn, T.; Kuchroo, V.K.; Anderson, A.C. Role of Th1 and Th17 cells in organ-specific autoimmunity. J. Autoimmun. 2008, 31, 252–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Orozco, N.; Chung, Y.; Chang, S.H.; Wang, Y.-H.; Dong, C. Th17 cells promote pancreatic inflammation but only induce diabetes efficiently in lymphopenic hosts after conversion into Th1 cells. Eur. J. Immunol. 2009, 39, 216–224. [Google Scholar] [PubMed]
- Haak, S.; Gyülveszi, G.; Becher, B. Th17 cells in autoimmune disease: Changing the verdict. Immunotherapy 2009, 1, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Viau, M.; Zouali, M. B-lymphocytes, innate immunity, and autoimmunity. Clin. Immunol. 2005, 114, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, K.; Clauder, A.-K.; Manz, R.A. Targeting B cells and plasma cells in autoimmune diseases. Front. Immunol. 2018, 9, 835. [Google Scholar] [CrossRef]
- Berthelot, J.-M.; Jamin, C.; Amrouche, K.; Le Goff, B.; Maugars, Y.; Youinou, P. Regulatory B cells play a key role in immune system balance. Jt. Bone Spine 2013, 80, 18–22. [Google Scholar] [CrossRef]
- Li, H.; Sun, S.; Lei, Q.; Lei, P.; Cai, X. M1-Polarized Macrophages Promote Self-Renewing Phenotype of Hepatic Progenitor Cells with Jagged1-Notch Signalling Involved: Relevance in Primary Sclerosing Cholangitis. J. Immunol. Res. 2018, 2018, 4807145. [Google Scholar] [CrossRef]
- Koushki, K.; Shahbaz, S.K.; Mashayekhi, K.; Sadeghi, M.; Zayeri, Z.D.; Taba, M.Y.; Banach, M.; Al-Rasadi, K.; Johnston, T.P.; Sahebkar, A. Anti-inflammatory Action of Statins in Cardiovascular Disease: The Role of Inflammasome and Toll-Like Receptor Pathways. Clin. Rev. Allergy Immunol. 2021, 60, 175–199. [Google Scholar] [CrossRef]
- Waldner, H. The role of innate immune responses in autoimmune disease development. Autoimmun. Rev. 2009, 8, 400–404. [Google Scholar] [CrossRef]
- Shu, J.; He, X.; Li, H.; Liu, X.; Qiu, X.; Zhou, T.; Wang, P.; Huang, X. The beneficial effect of human amnion mesenchymal cells in inhibition of inflammation and induction of neuronal repair in EAE mice. J. Immunol. Res. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Mavragani, C.P.; Moutsopoulos, H.M. Sjögren syndrome. CMAJ 2014, 186, E579–E586. [Google Scholar] [CrossRef] [Green Version]
- Loma, I.; Heyman, R. Multiple sclerosis: Pathogenesis and treatment. Curr. Neuropharmacol. 2011, 9, 409–416. [Google Scholar] [CrossRef]
- Pickup, J.C. Insulin-pump therapy for type 1 diabetes mellitus. N. Engl. J. Med. 2012, 366, 1616–1624. [Google Scholar] [CrossRef]
- Aletaha, D.; Smolen, J. The rheumatoid arthritis patient in the clinic: Comparing more than 1300 consecutive DMARD courses. Rheumatology 2002, 41, 1367–1374. [Google Scholar] [CrossRef] [Green Version]
- Sands, B.E.; Kaplan, G.G. The role of TNFα in ulcerative colitis. J. Clin. Pharmacol. 2007, 47, 930–941. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Fazio, V.W.; Feagan, B.G.; Hanauer, S.B. AGA technical review on perianal Crohn’s disease. Gastroenterology 2003, 125, 1508–1530. [Google Scholar] [CrossRef]
- Hegen, H.; Auer, M.; Deisenhammer, F. Pharmacokinetic considerations in the treatment of multiple sclerosis with interferon-β. Expert Opin. drug Metab. Toxicol. 2015, 11, 1803–1819. [Google Scholar] [CrossRef]
- Targan, S.R. Current limitations of IBD treatment: Where do we go from here? Ann. N. Y. Acad. Sci. 2006, 1072, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Shahbaz, S.K.; Varasteh, A.-R.; Koushki, K.; Ayati, S.H.; Mashayekhi, K.; Sadeghi, M.; Moghadam, M.; Sankian, M. Sublingual dendritic cells targeting by aptamer: Possible approach for improvement of sublingual immunotherapy efficacy. Int. Immunopharmacol. 2020, 85, 106603. [Google Scholar] [CrossRef]
- Keshavarz Shahbaz, S.; Foroughi, F.; Soltaninezhad, E.; Jamialahmadi, T.; Penson, P.E.; Sahebkar, A. Application of PLGA nano/microparticle delivery systems for immunomodulation and prevention of allotransplant rejection. Expert Opin. Drug Deliv. 2020, 17, 767–780. [Google Scholar] [CrossRef] [PubMed]
- Boraschi, D.; Costantino, L.; Italiani, P. Interaction of nanoparticles with immunocompetent cells: Nanosafety considerations. Nanomedicine 2012, 7, 121–131. [Google Scholar] [CrossRef]
- Moyano, D.F.; Goldsmith, M.; Solfiell, D.J.; Landesman-Milo, D.; Miranda, O.R.; Peer, D.; Rotello, V.M. Nanoparticle hydrophobicity dictates immune response. J. Am. Chem. Soc. 2012, 134, 3965–3967. [Google Scholar] [CrossRef] [Green Version]
- Zolnik, B.S.; González-Fernández, Á.; Sadrieh, N.; Dobrovolskaia, M.A. Minireview: Nanoparticles and the immune system. Endocrinology 2010, 151, 458–465. [Google Scholar] [CrossRef]
- Schrand, A.M.; Rahman, M.F.; Hussain, S.M.; Schlager, J.J.; Smith, D.A.; Syed, A.F. Metal-based nanoparticles and their toxicity assessment. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2010, 2, 544–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eifler, A.C.; Thaxton, C.S. Nanoparticle therapeutics: FDA approval, clinical trials, regulatory pathways, and case study. In Biomedical Nanotechnology; Springer: Amsterdam, The Netherlands, 2011; pp. 325–338. [Google Scholar]
- Clemente-Casares, X.; Santamaria, P. Nanomedicine in autoimmunity. Immunol. Lett. 2014, 158, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Serra, P.; Santamaria, P. Nanoparticle-based autoimmune disease therapy. Clin. Immunol. 2015, 160, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Neef, T.; Miller, S.D. Tolerogenic nanoparticles to treat islet autoimmunity. Curr. Diabetes Rep. 2017, 17, 84. [Google Scholar] [CrossRef] [PubMed]
- Prosperi, D.; Colombo, M.; Zanoni, I.; Granucci, F. Drug nanocarriers to treat autoimmunity and chronic inflammatory diseases. Semin. Immunol. 2017, 34, 61–67. [Google Scholar] [CrossRef]
- Guo, R.; Song, Y.; Wang, G.; Murray, R.W. Does core size matter in the kinetics of ligand exchanges of monolayer-protected Au clusters? J. Am. Chem. Soc. 2005, 127, 2752–2757. [Google Scholar] [CrossRef]
- Prades, R.; Guerrero, S.; Araya, E.; Molina, C.; Salas, E.; Zurita, E.; Selva, J.; Egea, G.; López-Iglesias, C.; Teixidó, M.; et al. Delivery of gold nanoparticles to the brain by conjugation with a peptide that recognizes the transferrin receptor. Biomaterials 2012, 33, 7194–7205. [Google Scholar] [CrossRef]
- Hainfeld, J.F.; Slatkin, D.N.; Smilowitz, H.M. The use of gold nanoparticles to enhance radiotherapy in mice. Phys. Med. Biol. 2004, 49, N309. [Google Scholar] [CrossRef]
- Craig, G.E.; Brown, S.D.; Lamprou, D.A.; Graham, D.; Wheate, N.J. Cisplatin-tethered gold nanoparticles that exhibit enhanced reproducibility, drug loading, and stability: A step closer to pharmaceutical approval? Inorg. Chem. 2012, 51, 3490–3497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalmodia, S.; Vandhana, S.; Rama, B.R.T.; Jayashree, B.; Seethalakshmi, T.S.; Umashankar, V.; Yang, W.; Barrow, C.J.; Krishnakumar, S.; Elchuri, S.V. Bio-conjugation of antioxidant peptide on surface-modified gold nanoparticles: A novel approach to enhance the radical scavenging property in cancer cell. Cancer Nanotechnol. 2016, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tom, R.T.; Suryanarayanan, V.; Reddy, P.G.; Baskaran, S.; Pradeep, T. Ciprofloxacin-protected gold nanoparticles. Langmuir 2004, 20, 1909–1914. [Google Scholar] [CrossRef]
- Koushki, K.; Varasteh, A.-R.; Shahbaz, S.K.; Sadeghi, M.; Mashayekhi, K.; Ayati, S.H.; Moghadam, M.; Sankian, M. Dc-specific aptamer decorated gold nanoparticles: A new attractive insight into the nanocarriers for allergy epicutaneous immunotherapy. Int. J. Pharm. 2020, 584, 119403. [Google Scholar] [CrossRef]
- Sadeghi, M.; Koushki, K.; Mashayekhi, K.; Ayati, S.H.; Shahbaz, S.K.; Moghadam, M.; Sankian, M. DC-targeted gold nanoparticles as an efficient and biocompatible carrier for modulating allergic responses in sublingual immunotherapy. Int. Immunopharmacol. 2020, 86, 106690. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Ho, V.H.; Chen, C.; Yang, Z.; Liu, D.; Chen, R.; Zhou, D. Efficient, pH-triggered drug delivery using a pH-responsive DNA-conjugated gold nanoparticle. Adv. Healthc. Mater. 2013, 2, 275–280. [Google Scholar] [CrossRef]
- Song, K.; Xu, P.; Meng, Y.; Geng, F.; Li, J.; Li, Z.; Xing, J.; Chen, J.; Kong, B. Smart gold nanoparticles enhance killing effect on cancer cells. Int. J. Oncol. 2013, 42, 597–608. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Kaushal, A.; Kumar, A.; Kumar, D. Ultrasensitive transglutaminase based nanosensor for early detection of celiac disease in human. Int. J. Biol. Macromol. 2017, 105, 905–911. [Google Scholar] [CrossRef]
- Jeong, E.H.; Jung, G.; Am Hong, C.; Lee, H. Gold nanoparticle (AuNP)-based drug delivery and molecular imaging for biomedical applications. Arch. Pharmacal Res. 2014, 37, 53–59. [Google Scholar] [CrossRef]
- Real-Fernández, F.; Rossi, G.; Lolli, F.; Papini, A.M.; Rovero, P. Label-free method for anti-glucopeptide antibody detection in Multiple Sclerosis. MethodsX 2015, 2, 141–144. [Google Scholar] [CrossRef]
- Hashimoto, M.; Toshima, H.; Yonezawa, T.; Kawai, K.; Narushima, T.; Kaga, M.; Endo, K. Responses of RAW264. 7 macrophages to water-dispersible gold and silver nanoparticles stabilized by metal–carbon σ-bonds. J. Biomed. Mater. Res. Part A 2014, 102, 1838–1849. [Google Scholar] [CrossRef]
- Connor, E.E.; Mwamuka, J.; Gole, A.; Murphy, C.J.; Wyatt, M.D. Gold nanoparticles are taken up by human cells but do not cause acute cytotoxicity. Small 2005, 1, 325–327. [Google Scholar] [CrossRef]
- Paciotti, G.F.; Myer, L.; Weinreich, D.; Goia, D.; Pavel, N.; McLaughlin, R.E.; Tamarkin, L. Colloidal gold: A novel nanoparticle vector for tumor directed drug delivery. Drug Deliv. 2004, 11, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Lasagna-Reeves, C.; Gonzalez-Romero, D.; Barria, M.A.; Olmedo, I.; Clos, A.; Ramanujam, V.M.S.; Urayama, A.; Vergara, L.; Kogan, M.J.; Soto, C. Bioaccumulation and toxicity of gold nanoparticles after repeated administration in mice. Biochem. Biophys. Res. Commun. 2010, 393, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Neuss, S.; Leifert, A.; Fischler, M.; Wen, F.; Simon, U.; Schmid, G.; Brandau, W.; Jahnen-Dechent, W. Size-dependent cytotoxicity of gold nanoparticles. Small 2007, 3, 1941–1949. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-S.; Hung, Y.-C.; Liau, I.; Huang, G.S. Assessment of the in vivo toxicity of gold nanoparticles. Nanoscale Res. Lett. 2009, 4, 858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, H.; Abdelhalim, M.; Alhomida, A.; Al Ayed, M. Transient increase in IL-1β, IL-6 and TNF-α gene expression in rat liver exposed to gold nanoparticles. Genet. Mol. Res. 2013, 12, 5851–5857. [Google Scholar] [CrossRef] [PubMed]
- Schaeublin, N.M.; Braydich-Stolle, L.K.; Maurer, E.I.; Park, K.; MacCuspie, R.I.; Afrooz, A.R.M.N.; Vaia, R.A.; Saleh, N.B.; Hussain, S.M. Does shape matter? Bioeffects of gold nanomaterials in a human skin cell model. Langmuir 2012, 28, 3248–3258. [Google Scholar] [CrossRef]
- Hirn, S.; Semmler-Behnke, M.; Schleh, C.; Wenk, A.; Lipka, J.; Schäffler, M.; Takenaka, S.; Möller, W.; Schmid, G.; Simon, U.; et al. Particle size-dependent and surface charge-dependent biodistribution of gold nanoparticles after intravenous administration. Eur. J. Pharm. Biopharm. 2011, 77, 407–416. [Google Scholar] [CrossRef] [Green Version]
- De Jong, W.H.; Hagens, W.I.; Krystek, P.; Burger, M.C.; Sips, A.J.; Geertsma, R.E. Particle size-dependent organ distribution of gold nanoparticles after intravenous administration. Biomaterials 2008, 29, 1912–1919. [Google Scholar] [CrossRef]
- Huo, S.; Ma, H.; Huang, K.; Liu, J.; Wei, T.; Jin, S.; Zhang, J.; He, S.; Liang, X.-J. Superior penetration and retention behavior of 50 nm gold nanoparticles in tumors. Cancer Res. 2013, 73, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-D.; Di Wu, X.S.; Liu, P.-X.; Yang, N.; Zhao, B.; Zhang, H.; Sun, Y.-M.; Zhang, L.-A.; Fan, F.-Y. Size-dependent in vivo toxicity of PEG-coated gold nanoparticles. Int. J. Nanomed. 2011, 6, 2071. [Google Scholar] [CrossRef] [Green Version]
- Kalishwaralal, K.; Deepak, V.; Pandian, S.R.K.; Gurunathan, S. Biological synthesis of gold nanocubes from Bacillus licheniformis. Bioresour. Technol. 2009, 100, 5356–5358. [Google Scholar] [CrossRef]
- Jeon, K.-I.; Byun, M.-S.; Jue, D.-M. Gold compound auranofin inhibits IκB kinase (IKK) by modifying Cys-179 of IKKβ subunit. Exp. Mol. Med. 2003, 35, 61–66. [Google Scholar] [CrossRef]
- Rizwan, H.; Mohanta, J.; Si, S.; Pal, A. Gold nanoparticles reduce high glucose-induced oxidative-nitrosative stress regulated inflammation and apoptosis via tuberin-mTOR/NF-κB pathways in macrophages. Int. J. Nanomed. 2017, 12, 5841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Khan, M.J. Nano-gold displayed anti-inflammatory property via NF-kB pathways by suppressing COX-2 activity. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1149–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dohnert, M.B.; Ferreira, G.K.; Silveira, P.C.L.; Zanoni, E.T.; Dohnert, L.H.; de Souza, C.T.; Paula, M.M.S. Inflammatory cytokines content in Achilles tendinopathy after phonophoresis treatment combined with gold nanoparticles and diclophenac diethylammonium in rats. Inflammation 2015, 38, 1044–1049. [Google Scholar] [CrossRef]
- Edrees, H.M.; Elbehiry, A.; Elmosaad, Y.M. Hypoglycemic and anti-inflammatory effect of gold nanoparticles in streptozotocin-induced type 1 diabetes in experimental rats. Nanotechnology 2017, 3, 4. [Google Scholar]
- Lai, T.-H.; Chung, C.-H.; Chen, B.-H.; Hung, C.-F.; Inbaraj, B.S.; Ma, M.-C.; Chen, H.-M.; Tsou, C.-J.; Wu, P.-H.; Wu, W.-B. Gold nanoparticles compromise TNF-α-induced endothelial cell adhesion molecule expression through NF-κB and protein degradation pathways and reduce neointima formation in a rat carotid balloon injury model. J. Biomed. Nanotechnol. 2016, 12, 2185–2201. [Google Scholar] [CrossRef]
- Chen, Y.-P.; Dai, Z.-H.; Liu, P.-C.; Chuu, J.-J.; Lee, K.-Y.; Lee, S.-L.; Chen, Y.-J. Effects of nanogold on the alleviation of carbon tetrachloride-induced hepatic injury in rats. Chin. J. Physiol. 2012, 55, 1–5. [Google Scholar]
- Kingston, M.; Pfau, J.C.; Gilmer, J.; Brey, R. Selective inhibitory effects of 50-nm gold nanoparticles on mouse macrophage and spleen cells. J. Immunotoxicol. 2016, 13, 198–208. [Google Scholar] [CrossRef] [Green Version]
- Sumbayev, V.V.; Yasinska, I.M.; Garcia, C.P.; Gilliland, D.; Lall, G.S.; Gibbs, B.F.; Bonsall, D.R.; Varani, L.; Rossi, F.; Calzolai, L. Gold nanoparticles downregulate interleukin-1β-induced pro-inflammatory responses. Small 2013, 9, 472–477. [Google Scholar] [CrossRef]
- Kajita, M.; Hikosaka, K.; Iitsuka, M.; Kanayama, A.; Toshima, N.; Miyamoto, Y. Platinum nanoparticle is a useful scavenger of superoxide anion and hydrogen peroxide. Free Radic. Res. 2007, 41, 615–626. [Google Scholar] [CrossRef]
- De Carvalho, T.G.; Garcia, V.B.; de Araújo, A.A.; da Silva Gasparotto, L.H.; Silva, H.; Guerra, G.C.B.; de Castro Miguel, E.; de Carvalho Leitão, R.F.; da Silva Costa, D.V.; Cruz, L.J.; et al. Spherical neutral gold nanoparticles improve anti-inflammatory response, oxidative stress and fibrosis in alcohol-methamphetamine-induced liver injury in rats. Int. J. Pharm. 2018, 548, 1–14. [Google Scholar] [CrossRef] [PubMed]
- De Araújo Júnior, R.F.; de Araújo, A.A.; Pessoa, J.B.; Neto, F.P.F.; da Silva, G.R.; Oliveira, A.L.C.S.L.; de Carvalho, T.G.; Silva, H.F.O.; Eugênio, M.; Sant’Anna, C.; et al. Anti-inflammatory, analgesic and anti-tumor properties of gold nanoparticles. Pharmacol. Rep. 2017, 69, 119–129. [Google Scholar] [CrossRef]
- Tsai, C.-Y.; Lu, S.-L.; Hu, C.-W.; Yeh, C.-S.; Lee, G.-B.; Lei, H.-Y. Size-dependent attenuation of TLR9 signaling by gold nanoparticles in macrophages. J. Immunol. 2012, 188, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Dohnert, M.B.; Venâncio, M.; Possato, J.C.; Zeferino, R.C.; Dohnert, L.H.; Zugno, A.I.; De Souza, C.T.; Paula, M.M.S.; Luciano, T.F. Gold nanoparticles and diclofenac diethylammonium administered by iontophoresis reduce inflammatory cytokines expression in Achilles tendinitis. Int. J. Nanomed. 2012, 7, 1651. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.-T.; He, W.; Wamer, W.G.; Hu, X.; Wu, X.; Lo, Y.M.; Yin, J.-J. Enzyme-mimetic effects of gold@ platinum nanorods on the antioxidant activity of ascorbic acid. Nanoscale 2013, 5, 1583–1591. [Google Scholar] [CrossRef]
- Yakimovich, T.; Lemaire, E.D.; Kofman, J. Engineering design review of stance-control knee-ankle-foot orthoses. J. Rehabil. Res. Dev. 2009, 46, 257–267. [Google Scholar] [CrossRef]
- Martín, R.; Menchon, C.; Apostolova, N.; Victor, V.M.; Alvaro, M.; Herance, J.R.; García, H. Nano-jewels in biology. Gold and platinum on diamond nanoparticles as antioxidant systems against cellular oxidative stress. ACS Nano 2010, 4, 6957–6965. [Google Scholar] [CrossRef]
- Muller, A.P.; Ferreira, G.K.; Pires, A.J.; de Bem Silveira, G.; de Souza, D.L.; de Abreu Brandolfi, J.; de Souza, C.T.; Paula, M.M.; Silveira, P.C.L. Gold nanoparticles prevent cognitive deficits, oxidative stress and inflammation in a rat model of sporadic dementia of Alzheimer’s type. Mater. Sci. Eng. C 2017, 77, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.S.; Kim, W.J.; Kim, J.J.; Kim, T.J.; Ye, S.K.; Song, M.D.; Kang, H.; Kim, D.W.; Moon, W.K.; Lee, K.H. Gold nanoparticles attenuate LPS-induced NO production through the inhibition of NF-κB and IFN-β/STAT1 pathways in RAW264.7 cells. Nitric Oxide 2010, 23, 214–219. [Google Scholar] [CrossRef]
- Sul, O.-J.; Kim, J.-C.; Kyung, T.-W.; Kim, H.-J.; Kim, Y.-Y.; Kim, S.-H.; Kim, J.-S.; Choi, H.-S. Gold nanoparticles inhibited the receptor activator of nuclear factor-κb ligand (RANKL)-induced osteoclast formation by acting as an antioxidant. Biosci. Biotechnol. Biochem. 2010, 74, 2209–2213. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Sasaki, J.; Yamaguchi, S.; Kawai, K.; Kawakami, H.; Iwasaki, Y.; Imazato, S. Gold nanoparticles inhibit matrix metalloproteases without cytotoxicity. J. Dent. Res. 2015, 94, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- BarathManiKanth, S.; Kalishwaralal, K.; Sriram, M.; Pandian, S.R.K.; Youn, H.-s.; Eom, S.; Gurunathan, S. Anti-oxidant effect of gold nanoparticles restrains hyperglycemic conditions in diabetic mice. J. Nanobiotechnology 2010, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haupenthal, D.P.d.S.; Possato, J.C.; Zaccaron, R.P.; Mendes, C.; Rodrigues, M.S.; Nesi, R.T.; Pinho, R.A.; Feuser, P.E.; Machado-de-Ávila, R.A.; Comim, C.M. Effects of chronic treatment with gold nanoparticles on inflammatory responses and oxidative stress in Mdx mice. J. Drug Target. 2020, 28, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, T.; Ishigaki, S.; Yamamoto, M.; Liang, Y.; Niwa, j.-I.; Takeuchi, H.; Doyu, M.; Sobue, G. Differential expression of inflammation-and apoptosis-related genes in spinal cords of a mutant SOD1 transgenic mouse model of familial amyotrophic lateral sclerosis. J. Neurochem. 2002, 80, 158–167. [Google Scholar] [CrossRef]
- Yen, H.J.; Hsu, S.h.; Tsai, C.L. Cytotoxicity and immunological response of gold and silver nanoparticles of different sizes. Small 2009, 5, 1553–1561. [Google Scholar] [CrossRef]
- Pan, Y.; Leifert, A.; Ruau, D.; Neuss, S.; Bornemann, J.; Schmid, G.; Brandau, W.; Simon, U.; Jahnen-Dechent, W. Gold nanoparticles of diameter 1.4 nm trigger necrosis by oxidative stress and mitochondrial damage. Small 2009, 5, 2067–2076. [Google Scholar] [CrossRef]
- Andreazza, A.C.; Kauer-Sant’Anna, M.; Frey, B.N.; Bond, D.J.; Kapczinski, F.; Young, L.T.; Yatham, L.N. Oxidative stress markers in bipolar disorder: A meta-analysis. J. Affect. Disord. 2008, 111, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Małaczewska, J. The splenocyte proliferative response and cytokine secretion in mice after oral administration of commercial gold nanocolloid. Pol. J. Vet. Sci. 2015, 18, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishanth, R.P.; Jyotsna, R.G.; Schlager, J.J.; Hussain, S.M.; Reddanna, P. Inflammatory responses of RAW 264.7 macrophages upon exposure to nanoparticles: Role of ROS-NFκB signaling pathway. Nanotoxicology 2011, 5, 502–516. [Google Scholar] [CrossRef] [PubMed]
- Gosens, I.; Post, J.A.; de la Fonteyne, L.J.J.; Jansen, E.H.J.M.; Geus, J.W.; Cassee, F.R.; de Jong, W.H. Impact of agglomeration state of nano-and submicron sized gold particles on pulmonary inflammation. Part. Fibre Toxicol. 2010, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, K.E.; Bakhiet, A.O.; Awadalla, M.E.; Khan, H.A. A priming dose protects against gold nanoparticles-induced proinflammatory cytokines mRNA expression in mice. Nanomedicine 2018, 13, 313–323. [Google Scholar] [CrossRef]
- Khan, H.A.; Abdelhalim, M.A.K.; Alhomida, A.S.; Al-Ayed, M.S. Effects of naked gold nanoparticles on proinflammatory cytokines mRNA expression in rat liver and kidney. BioMed Res. Int. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chithrani, B.D.; Ghazani, A.A.; Chan, W.C. Determining the size and shape dependence of gold nanoparticle uptake into mammalian cells. Nano Lett. 2006, 6, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Patra, H.K.; Banerjee, S.; Chaudhuri, U.; Lahiri, P.; Dasgupta, A.K. Cell selective response to gold nanoparticles. Nanomed. Nanotechnol. Biol. Med. 2007, 3, 111–119. [Google Scholar] [CrossRef]
- Saleh, H.M.; Soliman, O.A.; Elshazly, M.O.; Raafat, A.; Gohar, A.K.; Salaheldin, T.A. Acute hematologic, hepatologic, and nephrologic changes after intraperitoneal injections of 18 nm gold nanoparticles in hamsters. Int. J. Nanomed. 2016, 11, 2505. [Google Scholar] [CrossRef] [Green Version]
- Barreto, E.; Serra, M.F.; dos Santos, R.V.; dos Santos, C.E.A.; Hickmann, J.; Cotias, A.C.; Pão, C.R.R.; Trindade, S.G.; Schimidt, V.; Giacomelli, C.; et al. Local administration of gold nanoparticles prevents pivotal pathological changes in murine models of atopic asthma. J. Biomed. Nanotechnol. 2015, 11, 1038–1050. [Google Scholar] [CrossRef]
- Mulens-Arias, V.; Balfourier, A.; Nicolás-Boluda, A.; Carn, F.; Gazeau, F. Disturbance of adhesomes by gold nanoparticles reveals a size-and cell type-bias. Biomater. Sci. 2019, 7, 389–408. [Google Scholar] [CrossRef]
- Malaczewska, J. The in vitro effect of commercially available noble metal nanocolloids on the splenocyte proliferative response and cytokine production in mice. Pol. J. Vet. Sci. 2014, 17, 37–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pivodová, V.; Franková, J.; Galandáková, A.; Ulrichová, J. In vitro AuNPs’ cytotoxicity and their effect on wound healing. Nanobiomedicine 2015, 2, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.-H.; Lee, M.-Y.; Park, S.-J.; Choi, J.-S.; Oh, M.-K.; Kim, I.-S. Auranofin blocks interleukin-6 signalling by inhibiting phosphorylation of JAK1 and STAT3. Immunology 2007, 122, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Parnsamut, C.; Brimson, S. Effects of silver nanoparticles and gold nanoparticles on IL-2, IL-6, and TNF-α production via MAPK pathway in leukemic cell lines. Genet. Mol. Res. 2015, 14, 3650. [Google Scholar] [CrossRef] [PubMed]
- Zortéa, D.; Silveira, P.C.; Souza, P.S.; Fidelis, G.S.; Paganini, C.S.; Pozzi, B.G.; Tuon, T.; De Souza, C.T.; Paula, M.M.; Pinho, R.A. Effects of phonophoresis and gold nanoparticles in experimental model of muscle overuse: Role of oxidative stress. Ultrasound Med. Biol. 2015, 41, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Hussein, R.M.; Saleh, H. Promising therapeutic effect of gold nanoparticles against dinitrobenzene sulfonic acid-induced colitis in rats. Nanomedicine 2018, 13, 1657–1679. [Google Scholar] [CrossRef]
- Kesarkar, R.; Sangar, V.; Oza, G.; Sawant, T.; Kothari, S.; Sharon, M.; Chowdhary, A. Synthesis, Characterization and Hepatoprotective Activity of Neem Gold Nanoparticles for Improved Efficacy and Sustained Drug Release Profile of Azidothymidine. Int. J. Pharm. Sci. Rev. Res. 2014, 26, 117–122. [Google Scholar]
- Goodman, C.M.; McCusker, C.D.; Yilmaz, T.; Rotello, V.M. Toxicity of gold nanoparticles functionalized with cationic and anionic side chains. Bioconjugate Chem. 2004, 15, 897–900. [Google Scholar] [CrossRef]
- Bastús, N.G.; Sánchez-Tilló, E.; Pujals, S.; Farrera, C.; Kogan, M.J.; Giralt, E.; Celada, A.; Lloberas, J.; Puntes, V. Peptides conjugated to gold nanoparticles induce macrophage activation. Mol. Immunol. 2009, 46, 743–748. [Google Scholar] [CrossRef]
- Bastús, N.G.; Sánchez-Tilló, E.; Pujals, S.; Farrera, C.; López, C.; Giralt, E.; Celada, A.; Lloberas, J.; Puntes, V. Homogeneous conjugation of peptides onto gold nanoparticles enhances macrophage response. ACS Nano 2009, 3, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Dul, M.; Nikolic, T.; Stefanidou, M.; McAteer, M.A.; Williams, P.; Mous, J.; Roep, B.O.; Kochba, E.; Levin, Y.; Peakman, M.; et al. Conjugation of a peptide autoantigen to gold nanoparticles for intradermally administered antigen specific immunotherapy. Int. J. Pharm. 2019, 562, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Gao, C. Influences of size and surface coating of gold nanoparticles on inflammatory activation of macrophages. Colloids Surf. B Biointerfaces 2017, 160, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Mocan, T.; Matea, C.; Tabaran, F.; Iancu, C.; Orasan, R.; Mocan, L. In Vitro Administration of Gold Nanoparticles Functionalized with MUC-1 Protein Fragment Generates Anticancer Vaccine Response via Macrophage Activation and Polarization Mechanism. J. Cancer 2015, 6, 583–592. [Google Scholar] [CrossRef]
- MacParland, S.A.; Tsoi, K.M.; Ouyang, B.; Ma, X.-Z.; Manuel, J.; Fawaz, A.; Ostrowski, M.A.; Alman, B.A.; Zilman, A.; Chan, W.C.W.; et al. Phenotype Determines Nanoparticle Uptake by Human Macrophages from Liver and Blood. ACS Nano 2017, 11, 2428–2443. [Google Scholar] [CrossRef] [PubMed]
- Bartneck, M.; Keul, H.A.; Wambach, M.; Bornemann, J.; Gbureck, U.; Chatain, N.; Neuss, S.; Tacke, F.; Groll, J.; Zwadlo-Klarwasser, G. Effects of nanoparticle surface-coupled peptides, functional endgroups, and charge on intracellular distribution and functionality of human primary reticuloendothelial cells. Nanomed. Nanotechnol. Biol. Med. 2012, 8, 1282–1292. [Google Scholar] [CrossRef]
- Rosi, N.L.; Mirkin, C.A. Nanostructures in Biodiagnostics. Chem. Rev. 2005, 105, 1547–1562. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.C.; Giljohann, D.A.; Daniel, W.L.; Zheng, D.; Prigodich, A.E.; Mirkin, C.A. Scavenger receptors mediate cellular uptake of polyvalent oligonucleotide-functionalized gold nanoparticles. Bioconjugate Chem. 2010, 21, 2250–2256. [Google Scholar]
- Massich, M.D.; Giljohann, D.A.; Schmucker, A.L.; Patel, P.C.; Mirkin, C.A. Cellular response of polyvalent oligonucleotide—Gold nanoparticle conjugates. ACS Nano 2010, 4, 5641–5646. [Google Scholar] [CrossRef]
- Jensen, S.A.; Day, E.S.; Ko, C.H.; Hurley, L.A.; Luciano, J.P.; Kouri, F.M.; Merkel, T.J.; Luthi, A.J.; Patel, P.C.; Cutler, J.I.; et al. Spherical Nucleic Acid Nanoparticle Conjugates as an RNAi-Based Therapy for Glioblastoma. Sci. Transl. Med. 2013, 5, 209ra152. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-H.; Lei, Q.; Luo, G.-F.; Jia, H.-Z.; Hong, S.; Liu, Y.-X.; Cheng, Y.-J.; Zhang, X.-Z. Rational Design of Multifunctional Gold Nanoparticles via Host–Guest Interaction for Cancer-Targeted Therapy. ACS Appl. Mater. Interfaces 2015, 7, 17171–17180. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.-Y.; Chen, H.; Zhou, Y.-Y.; Wang, L.-T.; Hou, Y.; Xia, X.-H.; Ding, Y. Intraorgan Targeting of Gold Conjugates for Precise Liver Cancer Treatment. ACS Appl. Mater. Interfaces 2017, 9, 31458–31468. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Liang, J.-J.; Geng, D.-D.; Wu, D.; Dong, L.; Shen, W.-B.; Xia, X.-H.; Zhang, C. Development of a Liver-Targeting Gold–PEG–Galactose Nanoparticle Platform and a Structure–Function Study. Part. Part. Syst. Charact. 2014, 31, 347–356. [Google Scholar] [CrossRef]
- Lima, S.A.C.; Reis, S. Temperature-responsive polymeric nanospheres containing methotrexate and gold nanoparticles: A multi-drug system for theranostic in rheumatoid arthritis. Colloids Surf. B Biointerfaces 2015, 133, 378–387. [Google Scholar] [CrossRef]
- Farooq, M.U.; Novosad, V.; Rozhkova, E.A.; Wali, H.; Ali, A.; Fateh, A.A.; Neogi, P.B.; Neogi, A.; Wang, Z. Gold Nanoparticles-enabled Efficient Dual Delivery of Anticancer Therapeutics to HeLa Cells. Sci. Rep. 2018, 8, 2907. [Google Scholar] [CrossRef] [Green Version]
- Nosratabadi, R.; Rastin, M.; Sankian, M.; Haghmorad, D.; Mahmoudi, M. Hyperforin-loaded gold nanoparticle alleviates experimental autoimmune encephalomyelitis by suppressing Th1 and Th17 cells and upregulating regulatory T cells. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 1961–1971. [Google Scholar] [CrossRef]
- Rovais, M.R.A.; Alirezapour, B.; Moassesi, M.E.; Amiri, M.; Novin, F.B.; Maadi, E. Internalization capabilities of gold-198 nanoparticles: Comparative evaluation of effects of chitosan agent on cellular uptake into MCF-7. Appl. Radiat. Isot. 2018, 142, 85–91. [Google Scholar] [CrossRef]
- Paciotti, G.F.; Kingston, D.G.; Tamarkin, L. Colloidal gold nanoparticles: A novel nanoparticle platform for developing multifunctional tumor-targeted drug delivery vectors. Drug Dev. Res. 2006, 67, 47–54. [Google Scholar] [CrossRef]
- Libutti, S.K.; Paciotti, G.F.; Byrnes, A.A.; Alexander, H.R.; Gannon, W.E.; Walker, M.; Seidel, G.D.; Yuldasheva, N.; Tamarkin, L. Phase I and pharmacokinetic studies of CYT-6091, a novel PEGylated colloidal gold-rhTNF nanomedicine. Clin. Cancer Res. 2010, 16, 6139–6149. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.D.; Nativo, P.; Smith, J.-A.; Stirling, D.; Edwards, P.R.; Venugopal, B.; Flint, D.J.; Plumb, J.A.; Graham, D.; Wheate, N.J. Gold nanoparticles for the improved anticancer drug delivery of the active component of oxaliplatin. J. Am. Chem. Soc. 2010, 132, 4678–4684. [Google Scholar] [CrossRef]
- Dreaden, E.C.; Mwakwari, S.C.; Sodji, Q.H.; Oyelere, A.K.; El-Sayed, M.A. Tamoxifen− poly (ethylene glycol)− thiol gold nanoparticle conjugates: Enhanced potency and selective delivery for breast cancer treatment. Bioconjugate Chem. 2009, 20, 2247–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McInnes, I.B.; Schett, G. The pathogenesis of rheumatoid arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibofsky, A.; Winchester, R.J.; Patarroyo, M.; Fotino, M.; Kunkel, H.G. Disease associations of the Ia-like human alloantigens. Contrasting patterns in rheumatoid arthritis and systemic lupus erythematosus. J. Exp. Med. 1978, 148, 1728–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chabib, L.; Ikawati, Z.; Martien, R.; Ismail, H.; Wahyudi, M.D.P.; Arimurni, D.A.; Muhtadi, W.K.; Hidayat, A. Rheumatoid arthritis and the challenge of using nanoparticles for its treatment. MATEC Web Conf. 2018, 154, 04005. [Google Scholar] [CrossRef]
- Schnabel, A.; Gross, W.L. Low-dose methotrexate in rheumatic diseases—Efficacy, side effects, and risk factors for side effects. Semin. Arthritis Rheum. 1994, 23, 310–327. [Google Scholar] [CrossRef]
- Smolen, J.S.; Aletaha, D.; Koeller, M.; Weisman, M.H.; Emery, P. New therapies for treatment of rheumatoid arthritis. Lancet 2007, 370, 1861–1874. [Google Scholar] [CrossRef]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef]
- Taysi, S.; Polat, F.; Gul, M.; Sari, R.A.; Bakan, E. Lipid peroxidation, some extracellular antioxidants, and antioxidant enzymes in serum of patients with rheumatoid arthritis. Rheumatol. Int. 2002, 21, 200–204. [Google Scholar] [CrossRef]
- Brown, C.L.; Bushell, G.; Whitehouse, M.W.; Agrawal, D.; Tupe, S.; Paknikar, K.; Tiekink, E.R. Nanogoldpharmaceutics. Gold Bull. 2007, 40, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Jain, P.K.; Lee, K.S.; El-Sayed, I.H.; El-Sayed, M.A. Calculated absorption and scattering properties of gold nanoparticles of different size, shape, and composition: Applications in biological imaging and biomedicine. J. Phys. Chem. B 2006, 110, 7238–7248. [Google Scholar] [CrossRef] [Green Version]
- Chamberland, D.L.; Agarwal, A.; Kotov, N.; Fowlkes, J.B.; Carson, P.L.; Wang, X. Photoacoustic tomography of joints aided by an Etanercept-conjugated gold nanoparticle contrast agent—An ex vivo preliminary rat study. Nanotechnology 2008, 19, 095101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forestier, J. Rheumatoid arthritis and its treatment by gold salts. Lancet 1934, 224, 646–648. [Google Scholar] [CrossRef]
- Felson, D.T.; Anderson, J.J.; Meenan, R.F. The comparative efficacy and toxicity of second-line drugs in rheumatoid arthritis results of two metaanalyses. Arthritis Rheum. Off. J. Am. Coll. Rheumatol. 1990, 33, 1449–1461. [Google Scholar] [CrossRef] [PubMed]
- Shaw, C.F. Gold-based therapeutic agents. Chem. Rev. 1999, 99, 2589–2600. [Google Scholar] [CrossRef]
- Kean, W.F.; Kean, I. Clinical pharmacology of gold. Inflammopharmacology 2008, 16, 112–125. [Google Scholar] [CrossRef]
- Barrera, P.; Agnes, M.T.; van de Putte, L.B.; van der Meer, J.W. Effects of antirheumatic agents on cytokines. Semin. Arthritis Rheum. 1996, 25, 234–253. [Google Scholar] [CrossRef] [Green Version]
- Bondeson, J. The mechanisms of action of disease-modifying antirheumatic drugs: A review with emphasis on macrophage signal transduction and the induction of proinflammatory cytokines. Gen. Pharmacol. Vasc. Syst. 1997, 29, 127–150. [Google Scholar] [CrossRef]
- Lampa, J.; Klareskog, L.; Rönnelid, J. Effects of gold on cytokine production in vitro; increase of monocyte dependent interleukin 10 production and decrease of interferon-gamma levels. J. Rheumatol. 2002, 29, 21–28. [Google Scholar]
- Stern, I.; Wataha, J.; Lewis, J.; Messer, R.L.; Lockwood, P.; Tseng, W. Anti-rheumatic gold compounds as sublethal modulators of monocytic LPS-induced cytokine secretion. Toxicol. Vitr. 2005, 19, 365–371. [Google Scholar] [CrossRef]
- Wataha, J.C.; Lewis, J.B.; Volkmann, K.R.; Lockwood, P.E.; Messer, R.L.W.; Bouillaguet, S. Sublethal concentrations of Au (III), Pd (II), and Ni (II) differentially alter inflammatory cytokine secretion from activated monocytes. J. Biomed. Mater. Res. Part B Appl. Biomater. An. Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2004, 69, 11–17. [Google Scholar] [CrossRef]
- Weidauer, E.; Yasuda, Y.; Biswal, B.K.; Cherny, M.; James, M.N.; Brömme, D. Effects of disease-modifying anti-rheumatic drugs (DMARDs) on the activities of rheumatoid arthritis-associated cathepsins K and S. Biol. Chem. 2007, 388, 331–336. [Google Scholar] [CrossRef]
- Mustacich, D.; Powis, G. Thioredoxin reductase. Biochem. J. 2000, 346, 1–8. [Google Scholar] [CrossRef] [PubMed]
- James, L.R.; Xu, Z.-Q.; Sluyter, R.; Hawksworth, E.L.; Kelso, C.; Lai, B.; Paterson, D.J.; de Jonge, M.D.; Dixon, N.E.; Beck, J.L.; et al. An investigation into the interactions of gold nanoparticles and anti-arthritic drugs with macrophages, and their reactivity towards thioredoxin reductase. J. Inorg. Biochem. 2015, 142, 28–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, H.; Yamaguchi, M.; Shibasaki, T.; Ishii, T.; Bannai, S. Induction of stress proteins in mouse peritoneal macrophages by the antirheumatic agents gold sodium thiomalate and auranofin. Biochem. Pharmacol. 1995, 49, 1453–1457. [Google Scholar] [CrossRef]
- Heimbürger, M.; Lerner, R.; Palmblad, J. Effects of antirheumatic drugs on adhesiveness of endothelial cells and neutrophils. Biochem. Pharmacol. 1998, 56, 1661–1669. [Google Scholar] [CrossRef]
- Griem, P.; Gleichmann, E. [Gold antirheumatic drug: Desired and adverse effects of Au (I) and Au (III) [corrected] on the immune system. Z. Fur Rheumatol. 1996, 55, 348–358. [Google Scholar]
- Harth, M.; Cousin, K.; McCain, G.A. Sodium aurothiomalate inhibits T cell responses to interleukin-2. Immunopharmacol. Immunotoxicol. 1988, 10, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Wolf, R.E.; Hall, V.C. Inhibition of in vitro proliferative response of cultured t lymphocytes to interleukin-2 by gold sodium thiomalate. Arthritis Rheum. Off. J. Am. Coll. Rheumatol. 1988, 31, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Kropshofer, H.; Vogt, A.B.; Gleichmann, E.; Griem, P. Drug-induced inhibition of insulin recognition by T-cells: The antirheumatic drug aurothiomalate inhibits MHC binding of insulin peptide. Mol. Immunol. 1998, 35, 1081–1087. [Google Scholar] [CrossRef]
- Hirohata, S.; Nakanishi, K.; Yanagida, T.; Kawai, M.; Kikuchi, H.; Isshi, K. Synergistic inhibition of human B cell activation by gold sodium thiomalate and auranofin. Clin. Immunol. 1999, 91, 226–233. [Google Scholar] [CrossRef]
- Kiely, P.; Helbert, M.R.; Miles, J.; Oliveira, D. Immunosuppressant effect of gold on IgG subclasses and IgE; evidence for sparing of Th2 responses. Clin. Exp. Immunol. 2000, 120, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Lockie, L.M.; Smith, D.M. Forty-seven years experience with gold therapy in 1,019 rheumatoid arthritis patients. Semin. Arthritis Rheum. 1985, 14, 238–246. [Google Scholar] [CrossRef]
- Greinacher, A.; Eichler, P.; Lubenow, N.; Kiefel, V. Drug-inducedand Drug-dependent Immune Thrombocytopenias. Rev. Clin. Exp. Hematol. 2001, 5, 166–200. [Google Scholar] [CrossRef]
- Sakkas, L.; Chikanza, I.; Vaughan, R.; Welsh, K.; Panayi, G. Gold induced nephropathy in rheumatoid arthritis and HLA class II genes. Ann. Rheum. Dis. 1993, 52, 300–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Perez, M.; Gonzalez-Dominguez, J.; Mataran, L.; Garcia-Perez, S.; Salvatierra, D. Association of HLA-DR5 with mucocutaneous lesions in patients with rheumatoid arthritis receiving gold sodium thiomalate. J. Rheumatol. 1994, 21, 41–43. [Google Scholar] [PubMed]
- Evans, D.T.; Knapp, L.A.; Jing, P.; Mitchen, J.L.; Dykhuizen, M.; Montefiori, D.C.; Pauza, C.D.; Watkins, D.I. Rapid and slow progressors differ by a single MHC class I haplotype in a family of MHC-defined rhesus macaques infected with SIV. Immunol. Lett. 1999, 66, 53–59. [Google Scholar] [CrossRef]
- Havarinasab, S.; Johansson, U.; Pollard, K.M.; Hultman, P. Gold causes genetically determined autoimmune and immunostimulatory responses in mice. Clin. Exp. Immunol. 2007, 150, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Danscher, G. In vivo liberation of gold ions from gold implants. Autometallographic tracing of gold in cells adjacent to metallic gold. Histochem. Cell Biol. 2002, 117, 447–452. [Google Scholar] [CrossRef]
- Brown, C.L.; Whitehouse, M.W.; Tiekink, E.R.T.; Bushell, G.R. Colloidal metallic gold is not bio-inert. Inflammopharmacology 2008, 16, 133–137. [Google Scholar] [CrossRef]
- Zou, J. Gold (III)-induced oxidation of glycine: Relevance to the toxic sideeffects of gold drugs. J. Inorg. Biochem. 1999, 74, 352. [Google Scholar]
- Eisler, R. Chrysotherapy: A synoptic review. Inflamm. Res. 2003, 52, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Abraham, G.E.; Himmel, P.B. Management of rheumatoid arthritis: Rationale for the use of colloidal metallic gold. J. Nutr. Environ. Med. 1997, 7, 295–305. [Google Scholar] [CrossRef]
- Shukla, R.; Bansal, V.; Chaudhary, M.; Basu, A.; Bhonde, R.R.; Sastry, M. Biocompatibility of gold nanoparticles and their endocytotic fate inside the cellular compartment: A microscopic overview. Langmuir 2005, 21, 10644–10654. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-Y.; Shiau, A.-L.; Chen, S.-Y.; Chen, Y.-H.; Cheng, P.-C.; Chang, M.-Y.; Chen, D.-H.; Chou, C.-H.; Wang, C.-R.; Wu, C.-L. Amelioration of collagen-induced arthritis in rats by nanogold. Arthritis Rheum. Off. J. Am. Coll. Rheumatol. 2007, 56, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Leonavičienė, L.; Kirdaitė, G.; Bradūnaitė, R.; Vaitkienė, D.; Vasiliauskas, A.; Zabulytė, D.; Ramanavičienė, A.; Ramanavičius, A.; Ašmenavičius, T.; Mackiewicz, Z. Effect of gold nanoparticles in the treatment of established collagen arthritis in rats. Medicina 2012, 48, 16. [Google Scholar] [CrossRef]
- Kirdaite, G.; Leonaviciene, L.; Bradunaite, R.; Vasiliauskas, A.; Rudys, R.; Ramanaviciene, A.; Mackiewicz, Z. Antioxidant effects of gold nanoparticles on early stage of collagen-induced arthritis in rats. Res. Vet. Sci. 2019, 124, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Szekanecz, Z.; Besenyei, T.; Szentpétery, Á.; Koch, A.E. Angiogenesis and vasculogenesis in rheumatoid arthritis. Curr. Opin. Rheumatol. 2010, 22, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Shih, T.; Lindley, C. Bevacizumab: An angiogenesis inhibitor for the treatment of solid malignancies. Clin. Ther. 2006, 28, 1779–1802. [Google Scholar] [CrossRef] [PubMed]
- Gotink, K.J.; Verheul, H.M.W. Anti-angiogenic tyrosine kinase inhibitors: What is their mechanism of action? Angiogenesis 2010, 13, 1–14. [Google Scholar] [CrossRef] [Green Version]
- El-Ansary, M.; Eldin, T.; Ali, O.; Elsayed, A.; Elshoky, H. Functionalized gold nanoparticles for inhibition of vascular endothelial growth factor in arthritic patients. J. Nanomed. Res. 2015, 2, 00036. [Google Scholar]
- Yamashita, A.; Yonemitsu, Y.; Okano, S.; Nakagawa, K.; Nakashima, Y.; Irisa, T.; Iwamoto, Y.; Nagai, Y.; Hasegawa, M.; Sueishi, K. Fibroblast growth factor-2 determines severity of joint disease in adjuvant-induced arthritis in rats. J. Immunol. 2002, 168, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.-A.; Bae, D.-G.; Ryoo, J.-W.; Kim, H.-R.; Park, G.-S.; Cho, C.-S.; Chae, C.-B.; Kim, W.-U. Arginine-rich anti-vascular endothelial growth factor (anti-VEGF) hexapeptide inhibits collagen-induced arthritis and VEGF-stimulated productions of TNF-α and IL-6 by human monocytes. J. Immunol. 2005, 174, 5846–5855. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, R.; Mukherjee, P.; Xiong, Z.; Atala, A.; Soker, S.; Mukhopadhyay, D. Gold Nanoparticles Inhibit VEGF165-Induced Proliferation of HUVEC Cells. Nano Lett. 2004, 4, 2479–2481. [Google Scholar] [CrossRef]
- Maciejewski, M.; Sands, C.; Nair, N.; Ling, S.; Verstappen, S.; Hyrich, K.; Barton, A.; Ziemek, D.; Lewis, M.R.; Plant, D. Prediction of response of methotrexate in patients with rheumatoid arthritis using serum lipidomics. Sci. Rep. 2021, 11, 7266. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F.; Finotto, S. IL-6 signaling in autoimmunity, chronic inflammation and inflammation-associated cancer. Cytokine Growth Factor Rev. 2011, 22, 83–89. [Google Scholar] [CrossRef]
- Lee, M.-Y.; Yang, J.-A.; Jung, H.S.; Beack, S.; Choi, J.E.; Hur, W.; Koo, H.; Kim, K.; Yoon, S.K.; Kwang Hahn, S.K. Hyaluronic acid–gold nanoparticle/interferon α complex for targeted treatment of hepatitis C virus infection. ACS Nano 2012, 6, 9522–9531. [Google Scholar] [CrossRef]
- Lee, H.; Lee, M.-Y.; Bhang, S.H.; Kim, B.-S.; Kim, Y.S.; Ju, J.H.; Kim, K.S.; Hahn, S.K. Hyaluronate–gold nanoparticle/tocilizumab complex for the treatment of rheumatoid arthritis. Acs Nano 2014, 8, 4790–4798. [Google Scholar] [CrossRef]
- James, L.R.A.; Sluyter, R.; Dillon, C.T.; Ralph, S.F. Effects of gold nanoparticles and gold anti-arthritic compounds on inflammation marker expression in macrophages. Aust. J. Chem. 2017, 70, 1057–1068. [Google Scholar] [CrossRef]
- Krol, S.; Ellis-Behnke, R.; Marchetti, P. Nanomedicine for treatment of diabetes in an aging population: State-of-the-art and future developments. Maturitas 2012, 73, 61–67. [Google Scholar] [CrossRef]
- Zhi, Z.-L.; Khan, F.; Pickup, J.C. Multilayer nanoencapsulation: A nanomedicine technology for diabetes research and management. Diabetes Res. Clin. Pract. 2013, 100, 162–169. [Google Scholar] [CrossRef] [Green Version]
- Pickup, J.C.; Zhi, Z.-L.; Khan, F.; Saxl, T.; Birch, D.J. Nanomedicine and its potential in diabetes research and practice. Diabetes/Metab. Res. Rev. 2008, 24, 604–610. [Google Scholar] [CrossRef]
- Bhumkar, D.R.; Joshi, H.M.; Sastry, M.; Pokharkar, V.B. Chitosan reduced gold nanoparticles as novel carriers for transmucosal delivery of insulin. Pharm. Res. 2007, 24, 1415–1426. [Google Scholar] [CrossRef] [Green Version]
- Joshi, H.M.; Bhumkar, D.R.; Joshi, K.; Pokharkar, V.; Sastry, M. Gold nanoparticles as carriers for efficient transmucosal insulin delivery. Langmuir 2006, 22, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Aronson, D. Hyperglycemia and the pathobiology of diabetic complications. In Cardiovascular Diabetology: Clinical, Metabolic and Inflammatory Facets; Karger Publishers: Basel, Switzerland, 2008; Volume 45, pp. 1–16. [Google Scholar]
- Harrison, D.; Griendling, K.K.; Landmesser, U.; Hornig, B.; Drexler, H. Role of oxidative stress in atherosclerosis. Am. J. Cardiol. 2003, 91, 7–11. [Google Scholar] [CrossRef]
- Giugliano, D.; Ceriello, A.; Paolisso, G. Oxidative stress and diabetic vascular complications. Diabetes Care 1996, 19, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.M.; Russell, J.W.; Low, P.; Feldman, E.L. Oxidative stress in the pathogenesis of diabetic neuropathy. Endocr. Rev. 2004, 25, 612–628. [Google Scholar] [CrossRef]
- Liu, C.-L.; Wu, H.-T.; Hsiao, Y.-H.; Lai, C.-W.; Shih, C.-W.; Peng, Y.-K.; Tang, K.-C.; Chang, H.-W.; Chien, Y.-C.; Hsiao, J.-K.; et al. Insulin-directed synthesis of fluorescent gold nanoclusters: Preservation of insulin bioactivity and versatility in cell imaging. Angew. Chem. Int. Ed. 2011, 50, 7056–7060. [Google Scholar] [CrossRef] [PubMed]
- Shilo, M.; Berenstein, P.; Dreifuss, T.; Nash, Y.; Goldsmith, G.; Kazimirsky, G.; Motiei, M.; Frenkel, D.; Brodie, C.; Popovtzer, R. Insulin-coated gold nanoparticles as a new concept for personalized and adjustable glucose regulation. Nanoscale 2015, 7, 20489–20496. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-C.; Chen, W.-J.; Chen, Y.-C. Using Dextran-encapsulated gold nanoparticles as insulin carriers to prolong insulin activity. Nanomedicine 2017, 12, 1823–1834. [Google Scholar] [CrossRef]
- Kumari, Y.; Singh, S.K.; Kumar, R.; Kumar, B.; Kaur, G.; Gulati, M.; Tewari, D.; Gowthamarajan, K.; Karri, V.N.R.; Ayinkamiye, C.; et al. Modified apple polysaccharide capped gold nanoparticles for oral delivery of insulin. Int. J. Biol. Macromol. 2020, 149, 976–988. [Google Scholar] [CrossRef]
- Christen, U.; Wolfe, T.; Möhrle, U.; Hughes, A.C.; Rodrigo, E.; Green, E.A.; Flavell, R.A.; von Herrath, M.G. A dual role for TNF-α in type 1 diabetes: Islet-specific expression abrogates the ongoing autoimmune process when induced late but not early during pathogenesis. J. Immunol. 2001, 166, 7023–7032. [Google Scholar] [CrossRef] [PubMed]
- Zorena, K.; Mysliwska, J.; Mysliwiec, M.; Balcerska, A.; Lipowski, P.; Raczynska, K. Relationship between serum levels of tumor necrosis factor-alpha and interleukin-6 in diabetes mellitus type 1 children. Cent. Eur. J. Immunol. 2007, 32, 124. [Google Scholar]
- Lechleitner, M.; Koch, T.; Herold, M.; Dzien, A.; Hoppichler, F. Tumour necrosis factor-alpha plasma level in patients with type 1 diabetes mellitus and its association with glycaemic control and cardiovascular risk factors. J. Intern. Med. 2000, 248, 67–76. [Google Scholar] [CrossRef]
- Karthick, V.; Kumar, V.G.; Dhas, T.S.; Singaravelu, G.; Sadiq, A.M.; Govindaraju, K. Effect of biologically synthesized gold nanoparticles on alloxan-induced diabetic rats—An in vivo approach. Colloids Surf. B Biointerfaces 2014, 122, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Selim, M.E.; Hendi, A.A.; Alfallaj, E. The possible counteractive effect of gold nanoparticles against streptozotocin-induced type 1 diabetes in young male albino rats. Pak. J. Pharm. Sci. 2016, 29, 29. [Google Scholar]
- Bluestone, J.A.; Herold, K.; Eisenbarth, G. Genetics, pathogenesis and clinical interventions in type 1 diabetes. Nature 2010, 464, 1293–1300. [Google Scholar] [CrossRef] [Green Version]
- Okubo, Y.; Torrey, H.; Butterworth, J.; Zheng, H.; Faustman, D.L. Treg activation defect in type 1 diabetes: Correction with TNFR2 agonism. Clin. Transl. Immunol. 2016, 5, e56. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, M.; Roncarolo, M.-G. Immune intervention with T regulatory cells: Past lessons and future perspectives for type 1 diabetes. Semin. Immunol. 2011, 23, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Creusot, R.J.; Giannoukakis, N.; Trucco, M.; Clare-Salzler, M.J.; Fathman, C.G. It’s time to bring dendritic cell therapy to type 1 diabetes. Diabetes 2014, 63, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Yeste, A.; Takenaka, M.C.; Mascanfroni, I.D.; Nadeau, M.; Kenison, J.E.; Patel, B.; Tukpah, A.-M.; Babon, J.A.B.; DeNicola, M.; Kent, S.C.; et al. Tolerogenic nanoparticles inhibit T cell–mediated autoimmunity through SOCS2. Sci. Signal. 2016, 9, ra61. [Google Scholar] [CrossRef] [Green Version]
- Rink, J.S.; McMahon, K.M.; Chen, X.; Mirkin, C.A.; Thaxton, C.S.; Kaufman, D.B. Transfection of pancreatic islets using polyvalent DNA-functionalized gold nanoparticles. Surgery 2010, 148, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Jin, R.; Wu, G.; Li, Z.; Mirkin, A.C.A.; Schatz, G.C. What controls the melting properties of DNA-linked gold nanoparticle assemblies? J. Am. Chem. Soc. 2003, 125, 1643–1654. [Google Scholar] [CrossRef] [PubMed]
- Rosi, N.L.; Giljohann, D.A.; Thaxton, C.S.; Lytton-Jean, A.K.R.; Han, M.S.; Mirkin, C.A. Oligonucleotide-modified gold nanoparticles for intracellular gene regulation. Science 2006, 312, 1027–1030. [Google Scholar] [CrossRef] [PubMed]
- Tullman, M.J. Overview of the epidemiology, diagnosis, and disease progression associated with multiple sclerosis. Am. J. Manag. Care 2013, 19, S15–S20. [Google Scholar] [PubMed]
- Rostami, A.; Ciric, B. Role of Th17 cells in the pathogenesis of CNS inflammatory demyelination. J. Neurol. Sci. 2013, 333, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Lovett-Racke, A.E.; Yang, Y.; Racke, M.K. Th1 versus Th17: Are T cell cytokines relevant in multiple sclerosis? Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2011, 1812, 246–251. [Google Scholar] [CrossRef] [Green Version]
- Luchtman, D.W.; Ellwardt, E.; Larochelle, C.; Zipp, F. IL-17 and related cytokines involved in the pathology and immunotherapy of multiple sclerosis: Current and future developments. Cytokine Growth Factor Rev. 2014, 25, 403–413. [Google Scholar] [CrossRef]
- Link, J. Interferon-gamma, interleukin-4 and transforming growth factor-beta mRNA expression in multiple sclerosis and myasthenia gravis. Acta Neurol. Scandinavica. Suppl. 1994, 158, 1. [Google Scholar]
- Raphael, I.; Nalawade, S.; Eagar, T.N.; Forsthuber, T.G. T cell subsets and their signature cytokines in autoimmune and inflammatory diseases. Cytokine 2015, 74, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Venkatesha, S.H.; Dudics, S.; Acharya, B.; Moudgil, K.D. Cytokine-modulating strategies and newer cytokine targets for arthritis therapy. Int. J. Mol. Sci. 2015, 16, 887–906. [Google Scholar] [CrossRef] [Green Version]
- Mastorodemos, V.; Ioannou, M.; Verginis, P. Cell-based modulation of autoimmune responses in multiple sclerosis and experimental autoimmmune encephalomyelitis: Therapeutic implications. Neuroimmunomodulation 2015, 22, 181–195. [Google Scholar] [CrossRef] [Green Version]
- Astier, A.L.; Hafler, D.A. Abnormal Tr1 differentiation in multiple sclerosis. J. Neuroimmunol. 2007, 191, 70–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polak, P.; Shefi, O. Nanometric agents in the service of neuroscience: Manipulation of neuronal growth and activity using nanoparticles. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 1467–1479. [Google Scholar] [CrossRef]
- Liu, Z.; Shen, Y.; Wu, Y.; Yang, Y.; Wu, J.; Zhou, P.; Lu, X.; Guo, Z. An intrinsic therapy of gold nanoparticles in focal cerebral ischemia-reperfusion injury in rats. J. Biomed. Nanotechnol. 2013, 9, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Papastefanaki, F.; Jakovcevski, I.; Poulia, N.; Djogo, N.; Schulz, F.; Martinovic, T.; Ciric, D.; Loers, G.; Vossmeyer, T.; Weller, H.; et al. Intraspinal Delivery of Polyethylene Glycol-coated Gold Nanoparticles Promotes Functional Recovery After Spinal Cord Injury. Mol. Ther. 2015, 23, 993–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghaie, T.; Jazayeri, M.H.; Avan, A.; Anissian, A.; Salari, A.-A. Gold nanoparticles and polyethylene glycol alleviate clinical symptoms and alter cytokine secretion in a mouse model of experimental autoimmune encephalomyelitis. IUBMB Life 2019, 71, 1313–1321. [Google Scholar] [CrossRef]
- Yeste, A.; Nadeau, M.; Burns, E.J.; Weiner, H.L.; Quintana, F.J. Nanoparticle-mediated codelivery of myelin antigen and a tolerogenic small molecule suppresses experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA 2012, 109, 11270–11275. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Peters, T.; Sindrilaru, A.; Scharffetter-Kochanek, K. Key role of macrophages in the pathogenesis of CD18 hypomorphic murine model of psoriasis. J. Investig. Dermatol. 2009, 129, 1100–1114. [Google Scholar] [CrossRef] [Green Version]
- Crisan, D.; Scharffetter-Kochanek, K.; Crisan, M.; Schatz, S.; Hainzl, A.; Olenic, L.; Filip, A.; Schneider, L.A.; Sindrilaru, A. Topical silver and gold nanoparticles complexed with Cornus mas suppress inflammation in human psoriasis plaques by inhibiting NF-κB activity. Exp. Dermatol. 2018, 27, 1166–1169. [Google Scholar] [CrossRef] [Green Version]
- Szentpétery, Á.; Horváth, Á.; Gulyás, K.; Pethö, Z.; Bhattoa, H.P.; Szántó, S.; Szücs, G.; FitzGerald, O.; Schett, G.; Szekanecz, Z. Effects of targeted therapies on the bone in arthritides. Autoimmun. Rev. 2017, 16, 313–320. [Google Scholar] [CrossRef]
- Florea, A.; Melinte, G.; Simon, I.; Cristea, C. Electrochemical biosensors as potential diagnostic devices for autoimmune diseases. Biosensors 2019, 9, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y. Developing Electrochemical Biosensors for Point-of-care Diagnostics of Cardiovascular Biomarkers. Master’s Thesis, The University of Western Ontario, London, ON, Canada, 2013. [Google Scholar]
- Tuteja, S.K.; Sabherwal, P.; Deep, A.; Rastogi, R.; Paul, A.K.; Suri, C.R. Biofunctionalized rebar graphene (f-RG) for label-free detection of cardiac marker troponin I. ACS Appl. Mater. Interfaces 2014, 6, 14767–14771. [Google Scholar] [CrossRef]
- Hosu, O.; Selvolini, G.; Cristea, C.; Marrazza, G. Electrochemical immunosensors for disease detection and diagnosis. Curr. Med. Chem. 2018, 25, 4119–4137. [Google Scholar] [CrossRef]
- Lim, S.A.; Yoshikawa, H.; Tamiya, E.; Yasin, H.M.; Ahmed, P.M.U. A highly sensitive gold nanoparticle bioprobe based electrochemical immunosensor using screen printed graphene biochip. RSC Adv. 2014, 4, 58460–58466. [Google Scholar] [CrossRef]
- Giannetto, M.; Bianchi, V.; Gentili, S.; Fortunati, S.; De Munari, I.; Careri, M. An integrated IoT-Wi-Fi board for remote data acquisition and sharing from innovative immunosensors. Case of study: Diagnosis of celiac disease. Sens. Actuators B Chem. 2018, 273, 1395–1403. [Google Scholar] [CrossRef]
- Dahlbom, I.; Olsson, M.; Forooz, N.K.; Sjöholm, A.G.; Truedsson, L.; Hansson, T. Immunoglobulin G (IgG) anti-tissue transglutaminase antibodies used as markers for IgA-deficient celiac disease patients. Clin. Diagn. Lab. Immunol. 2005, 12, 254–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Huang, N.; Lu, Q.; Liu, M.; Li, H.; Zhang, Y.; Yao, S. A double signal electrochemical human immunoglobulin G immunosensor based on gold nanoparticles-polydopamine functionalized reduced graphene oxide as a sensor platform and AgNPs/carbon nanocomposite as signal probe and catalytic substrate. Biosens. Bioelectron. 2016, 77, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Neves, M.M.; González-García, M.B.; Santos-Silva, A.; Costa-García, A. Voltammetric immunosensor for the diagnosis of celiac disease based on the quantification of anti-gliadin antibodies. Sens. Actuators B Chem. 2012, 163, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Neves, M.M.; González-García, M.B.; Nouws, H.P.; Costa-García, A. Celiac disease detection using a transglutaminase electrochemical immunosensor fabricated on nanohybrid screen-printed carbon electrodes. Biosens. Bioelectron. 2012, 31, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Kaur, A.; Shimoni, O.; Wallach, M. Novel screening test for celiac disease using peptide functionalised gold nanoparticles. World J. Gastroenterol. 2018, 24, 5379–5390. [Google Scholar] [CrossRef]
- Balkenhohl, T.; Lisdat, F. Screen-printed electrodes as impedimetric immunosensors for the detection of anti-transglutaminase antibodies in human sera. Anal. Chim. Acta 2007, 597, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Briani, C.; Samaroo, D.; Alaedini, A. Celiac disease: From gluten to autoimmunity. Autoimmun. Rev. 2008, 7, 644–650. [Google Scholar] [CrossRef]
- Derkus, B.; Emregul, E.; Yucesan, C.; Emregul, K.C. Myelin basic protein immunosensor for multiple sclerosis detection based upon label-free electrochemical impedance spectroscopy. Biosens. Bioelectron. 2013, 46, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, R.; Li, P.; Yi, W.; Zhang, Z.; Chen, S.; Su, S.; Zhao, L.; Hu, C. Development of a novel method to measure macrophage migration inhibitory factor (MIF) in sera of patients with rheumatoid arthritis by combined electrochemical immunosensor. Int. Immunopharmacol. 2008, 8, 859–865. [Google Scholar] [CrossRef]
- Kou, B.-B.; Zhang, L.; Xie, H.; Wang, D.; Yuan, Y.-L.; Chai, Y.-Q.; Yuan, R. DNA Enzyme-Decorated DNA Nanoladders as Enhancer for Peptide Cleavage-Based Electrochemical Biosensor. ACS Appl. Mater. Interfaces 2016, 8, 22869–22874. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characterization of GNP | Animal Model/Cell Line | Mechanism of Actions | Main Effect | Ref. | ||
|---|---|---|---|---|---|---|
| Inflammatory | Anti-Inflammatory | Anti-Oxidant | ||||
| 5, 15, 20, and 35 nm GNPs | C57BL/6 male mice—THP-1 cells | -Decreased TNF-a | ✔ | [96] | ||
| -Decreased HIF-1 | ||||||
| -Decreased NF-kB | ||||||
| -Down-regulation of IL-1β-induced inflammatory by reducing NF-kB. | ||||||
| -The 5 nm AuNPs completely blocked the inflammatory process, 15 nm AuNPs were less effective, and 35 nm AuNPs did not display a statistically significant effect. | ||||||
| 35 mm | Rat | -Decreased IL-1b | ✔ | [101] | ||
| -Decreased TNF-a | ||||||
| 10 nm | Rat | -Decreased IL-1b | ✔ | [91] | ||
| -Decreased TNF-a | ||||||
| 50 nm | RAW 264.7 macrophages | -Decreased IL-1b | ✔ | ✔ | [95] | |
| -Decreased ROS | ||||||
| -Reduced interleukin (IL)-17 and TNFα triggered by LPS | ||||||
| 15 nm | Mice | -Reduced interleukin (IL)-17a | ✔ | [68] | ||
| -Reduced neutrophil recruitment | ||||||
| -Increased IL-10 levels | ||||||
| 6.3 nm | Mice | -Reduced the levels of IL-1, IL-5, and IL-6 in the BAL | ✔ | ✔ | [124] | |
| -Reduced IL-4, IL-5, IL-6, IL-13, eotaxin-1, and eotaxin-2 in lung tissue | ||||||
| -Inhibited inflammatory infiltration in the airways | ||||||
| -Significant reduction in the levels of malondialdehyde (MDA) | ||||||
| 10 and 50 nm | Rat Liver | -Both sizes significantly transient increase cytokine gene expression include TNF-α, IL-6, and IL-1β | ✔ | [81] | ||
| -The GNPs with 50 nm size induced more severe inflammatory responses compared to smaller GNPs. | ||||||
| 3, 11, 16, 30, and 40 nm | RAW264.7 (ATCC, TIB-71) SV40-transformed endothelial cells SVEC4-10 7 (ATCC, CRL-2181) and the murine mesenchymal stem cell line C3H10T1/2 (ATCC, CCL-226) | -Macrophage production of the monocyte chemoattractant RANTES/CCL5 depended on GNP size, i.e., GNP 11 nm significantly decreased CCL5 secretion while GNP 16 nm had the opposite effect. | ✔ | [125] | ||
| -Enhanced TNFα secretion | ||||||
| -Did not induce IL-10 secretion | ||||||
| 10, 30, 50, and 80 nm | BALB/c mice | -50 nm GNP significantly induced the M1 macrophage phenotype. | ✔ | [123] | ||
| -Increased IL-b, IL-6, and TNF-a in 50 nm nanospheres treatment | ||||||
| -50 nm GNP via activation of the NF-κB signal pathway led to SAA activation | ||||||
| 50–250 nm | Wistar-derived rats | -Increased of in IL-6 and TNF-a 250 nm single GNPs | ✔ | [118] | ||
| -Significant increase in immune cells, especially macrophages | ||||||
| -Increased MCP-1 and MIP-2 | ||||||
| -Increased TNF-α and IL-6 levels after treatment with 250 nm single GNPss | ||||||
| -Increased neutrophils after 24 h along with single 250 nm particles | ||||||
| 10–15 nm | RAW264.7 cells | -Blocked the activation | ✔ | ✔ | [107] | |
| -Inhibitory effects on IFN-b mRNA expression | ||||||
| -Attenuate nitric oxide levels | ||||||
| 20 nm | Rat | -Decreased IL-1β | ✔ | ✔ | [90] | |
| -Downregulated mRNA expression of iNOS, COX-2, TNF-α, and NF-kB | ||||||
| Up to 5 nm | Mice | -All cytokines were unaffected along with intermediate concentrations (2.5–5 ppm) | ✔ | [126] | ||
| TNF-α and IL-1β significantly decreased along with the highest concentration (10 ppm) but stimulated IL-6. | ||||||
| Production of TNF-α and IL-2 was decreased along with low concentrations but stimulated IL-1α | ||||||
| 25–50 nm | NHDF and NHEK | -Decreased TNF-a | ✔ + antiangiogenic activity | [127] | ||
| -Decreased IL-6 | ||||||
| -Decreased of IL-2 levels | ||||||
| -Decreased proteins involved in angiogenesis such as VEGF and bFGF. | ||||||
| 20 nm | RAW264.7 cells | -Decreased gene expression of MMP-2/-9, CX3CL-1, CCL-8, CX3CL-10, ICAM, IL-1α, and TNF-α in a dose-dependent manner | ✔ | ✔ | [89] | |
| -Inhibited of NF-κB pathway via ERK1/2MAPK/Akt/tuber in-mTOR kinases interference, which resulted in reducing oxidative-nitrosative stress | ||||||
| Auranofin | -Blocked IL-6 | ✔ | [128] | |||
| -Blockaded of JAK1/STAT3 signalling. | ||||||
| Au-S = 2.81 Au-M = 5.52 Au-L = 38.05 | cell culture | -GNPs (especially those with a smaller diameter) up-regulate the expressions of pro-inflammatory genes | ✔ | [113] | ||
| -IL-1, IL-6, and TNF-a. | ||||||
| -Expressions of proinflammatory genes decreased with the increased size of AuNPs | ||||||
| 10 and 50 nm | Rat | -Increased cytokines gene expression by both sizes of GNPs (10 and 50 nm) in the liver | [120] | |||
| -The GNPs with 50 nm size induced severe inflammatory response compared with smaller GNP size | ||||||
| -The GNPs do not have any effect on IL-1β in the kidney | ||||||
| -The GNPs with 10 nm size do not have any effect on TNF-𝛼 and IL-6 gene expression | ||||||
| -The GNPs with 50 nm size significantly increase expression of IL-6 and TNF-𝛼 in the kidneys of rats | ||||||
| 10–50 nm | Leukemic cell lines (T-lymphocytic Jurkat and monocytic U937 cells) | -Stimulated TNF-α production | ✔ | [129] | ||
| -Inhibited interleukin-6 | ||||||
| -Inhibited interleukin-2 production | ||||||
| 5.5 nm | Cell culture (HUVECs; VECs) | -Reduced TNF-a | ✔ | ✔ | [93] | |
| -Reduced monocyte adhesion to VECs in vitro and arterial | ||||||
| -Reduced NF-kB | ||||||
| -Reduced ROS | ||||||
| 25 nm | Wistar rats | -Decreased TNF-a | ✔ | ✔ | [130] | |
| -Decreased IL-6 | ||||||
| -Decreased SOD and Catalase (CAT) activity | ||||||
| -Decreased superoxide and Nitrite levels | ||||||
| 16–25 nm | Wistar rats | -Decreased TNF-α levels | ✔ | ✔ | [131] | |
| -Decreased IL-6 levels | ||||||
| -Significant decrease in antioxidant markers such as -GSH, SOD and CAT in the colon. | ||||||
| 30–40 nm | Rat/in vitro | -Downregulation of TNF-a | ✔ | [132] | ||
| -Controlled IL-6 secretion | ||||||
| -Upregulation IL-10 | ||||||
| Disease | Electrode Architecture | Target | Label | Detection Method | Sample | Ref |
|---|---|---|---|---|---|---|
| Celiac Disease | GNPs/SAM-GCE | IgA anti-tTG | AP | CPV | Serum from patients | [262] |
| IgB anti-tTG | ||||||
| CNTs/GNPs-SPE | IgA anti-tTG | AP | CV | Serum from patients | [263] | |
| IgB anti-tTG | ||||||
| Au/SAM-GCE | IgA anti-tTG | HRP | CV | Serum from patients | [264] | |
| IgB anti-tTG | ||||||
| Screen-printed carbon electrodes (SPCE) nanostructure with carbon-nanotubes and GNPs | IgA and IgG type anti-tTG | AP | CV | Serum from patients | [265] | |
| GNP-Peptide-AGA | anti-gliadin antibody | Spiked samples Serum from patients | [266] | |||
| GQD/PAMAM/GNP/MWCNT | IgA anti-tTG | DPV with redox probe | Human serum | [72] | ||
| Poly (sodium-4-styrensulfonic acid)-gold SPE | Anti-tTG | POD | EIS | Serum from patients | [267] | |
| Gold electrodes with carboxylic-ended bipodal alkanethio | AGA | HRP | Chronoamperometry | Serum from patients | [268] | |
| Rheumatoid Arthritis | GNPs-NTiP-Thi-gold electrode | MIF | DPV with redox probe | Serum from patients | [264] | |
| Non-Specific Biomarkers | Electroplating gold onto a disposable printed circuit board electrode | IL-12 | EIS | Spiked serum | [269] | |
| GNPs-PDA-GO | HIgG | AgNPs/carbon nanocomposite/benzoquinon | Spiked serum | [270] | ||
| Multiple Sclerosis | Gold sensor chip | Glucopeptide CSF114(Glc) antigen | SPR detection system | Serum | [74] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koushki, K.; Keshavarz Shahbaz, S.; Keshavarz, M.; Bezsonov, E.E.; Sathyapalan, T.; Sahebkar, A. Gold Nanoparticles: Multifaceted Roles in the Management of Autoimmune Disorders. Biomolecules 2021, 11, 1289. https://doi.org/10.3390/biom11091289
Koushki K, Keshavarz Shahbaz S, Keshavarz M, Bezsonov EE, Sathyapalan T, Sahebkar A. Gold Nanoparticles: Multifaceted Roles in the Management of Autoimmune Disorders. Biomolecules. 2021; 11(9):1289. https://doi.org/10.3390/biom11091289
Chicago/Turabian StyleKoushki, Khadijeh, Sanaz Keshavarz Shahbaz, Mohsen Keshavarz, Evgeny E. Bezsonov, Thozhukat Sathyapalan, and Amirhossein Sahebkar. 2021. "Gold Nanoparticles: Multifaceted Roles in the Management of Autoimmune Disorders" Biomolecules 11, no. 9: 1289. https://doi.org/10.3390/biom11091289