Myxovirus Resistance Protein 1 (MX1), a Novel HO-1 Interactor, Tilts the Balance of Endoplasmic Reticulum Stress towards Pro-Death Events in Prostate Cancer


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Treatments, Reagents, and Antibodies
2.2. Plasmids and Transient Transfections
2.3. RNA Isolation, c-DNA Synthesis, and Quantitative Real-Time Polymerase Chain Reaction
2.4. RNA Sequencing
2.5. Immunofluorescence Assay
2.6. Protein Extraction, Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis and Western Blot
2.7. Apoptosis Analysis by Flow Cytometry
2.8. siRNA Transfection
2.9. Statistical Analysis
2.10. Bioinformatics Analysis
2.10.1. Oncomine Meta-Analysis in Prostate Cancer Patients
2.10.2. RNA-Seq Gene Expression and Correlation Analysis in Prostate Cancer Patients
2.10.3. Gene Expression Microarray and Relapse-Free Survival in Prostate Cancer Patients
2.10.4. cBioPortal Exome Analysis in Prostate Cancer Patients
2.10.5. Statistical Analysis
3. Results
3.1. Clinical Relevance of the HO-1 Interactome in Prostate Cancer
3.2. Transcriptomic Analysis of HO-1 Interactors and Risk of Relapse in Prostate Cancer Patients
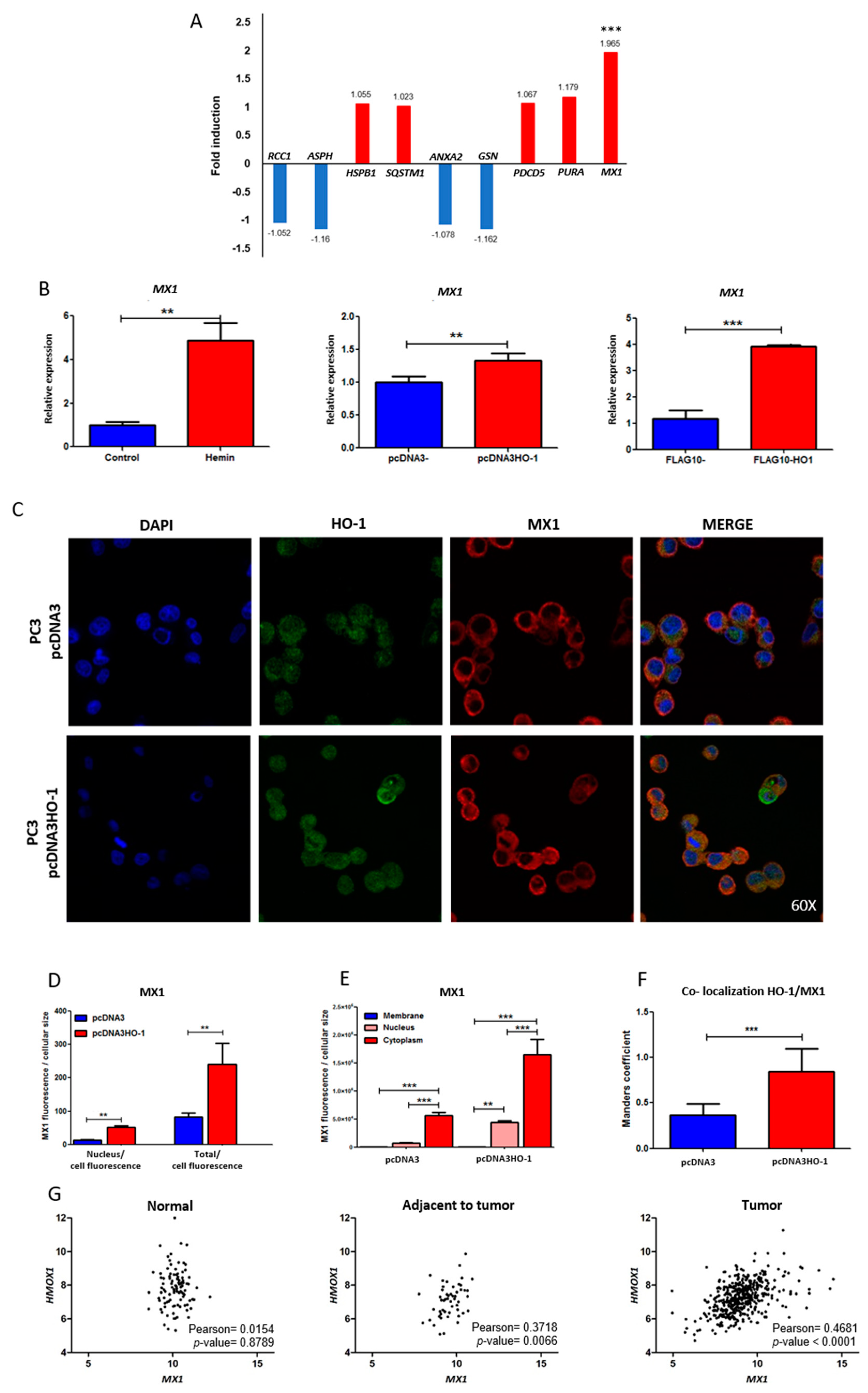
3.3. mRNA Expression Profiles of HO-1 Interactors under Genetic Induction of HO-1
3.4. Effect of HO-1 Modulation on the Expression and Localization of MX1
3.5. Correlation of Expression between MX1 and HMOX1 Genes in Normal Prostate and Prostate Cancer Samples
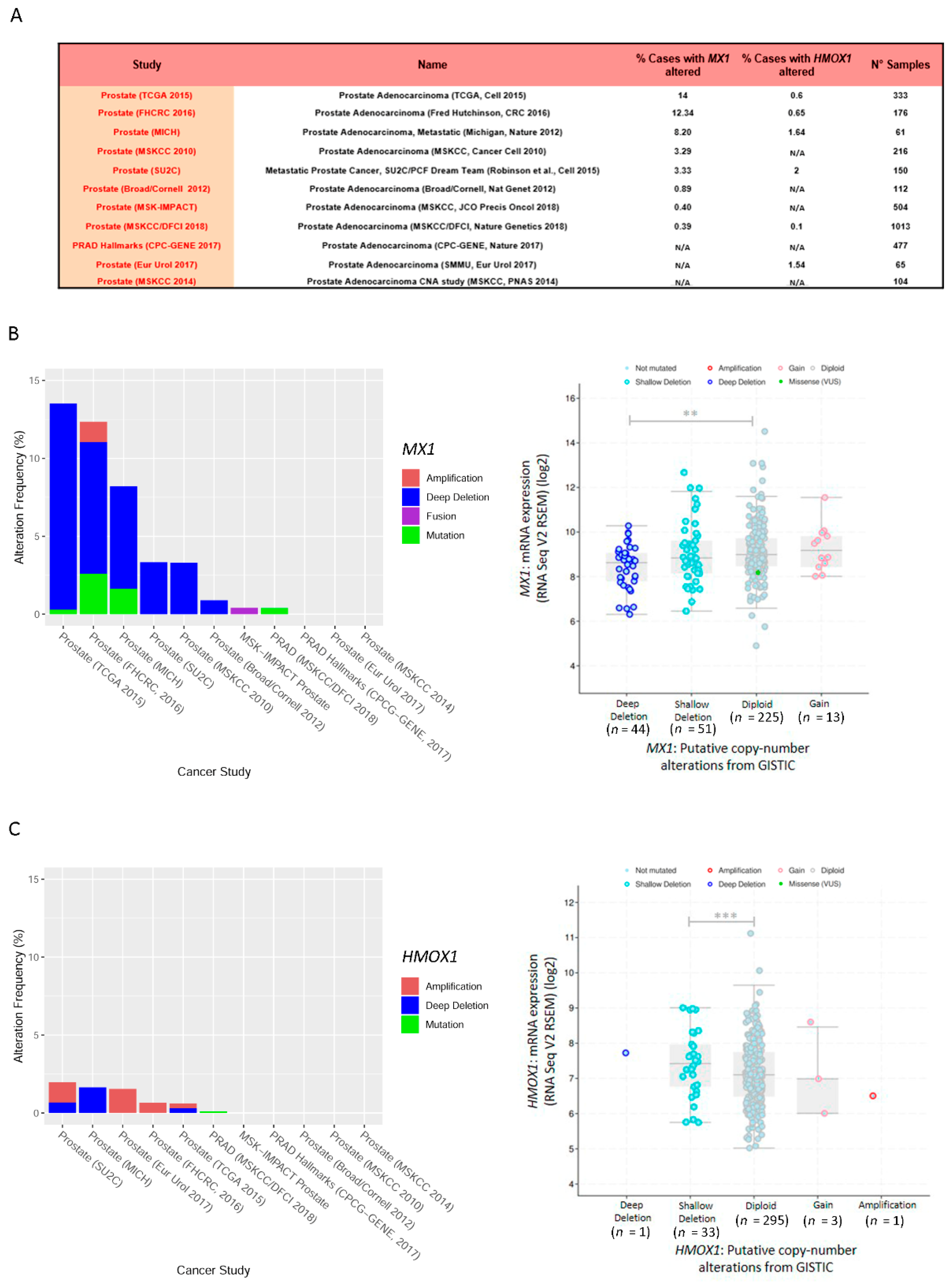
3.6. Exome and RNA-Seq Correlation Analyses of MX1 and HMOX1 in Prostate Cancer
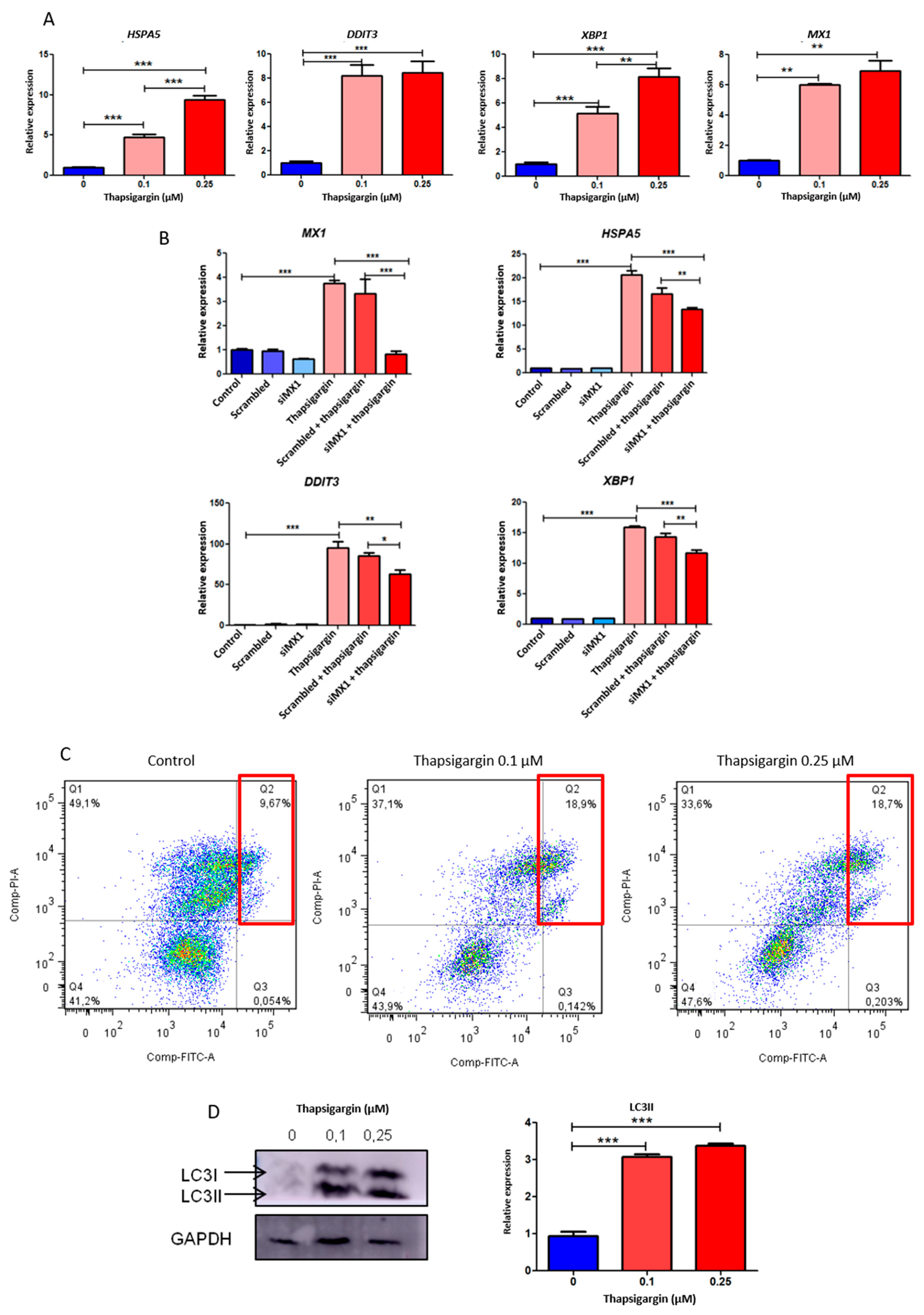
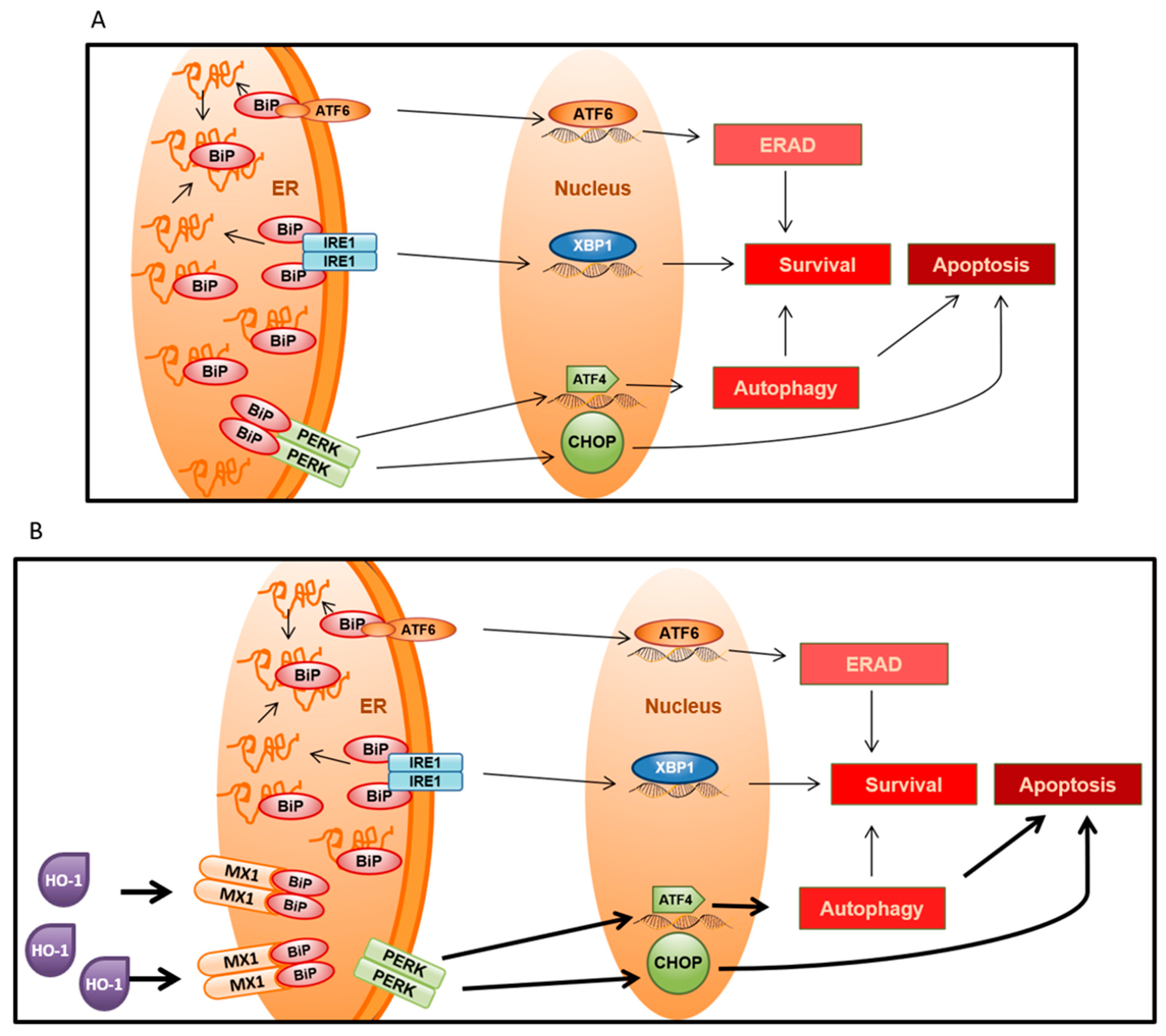
3.7. MX1 and Endoplasmic Reticulum Stress in Prostate Cancer
3.8. Effect of Thapsigargin-Induced Endoplasmic Reticulum Stress in Apoptosis and Autophagy in Prostate Cancer
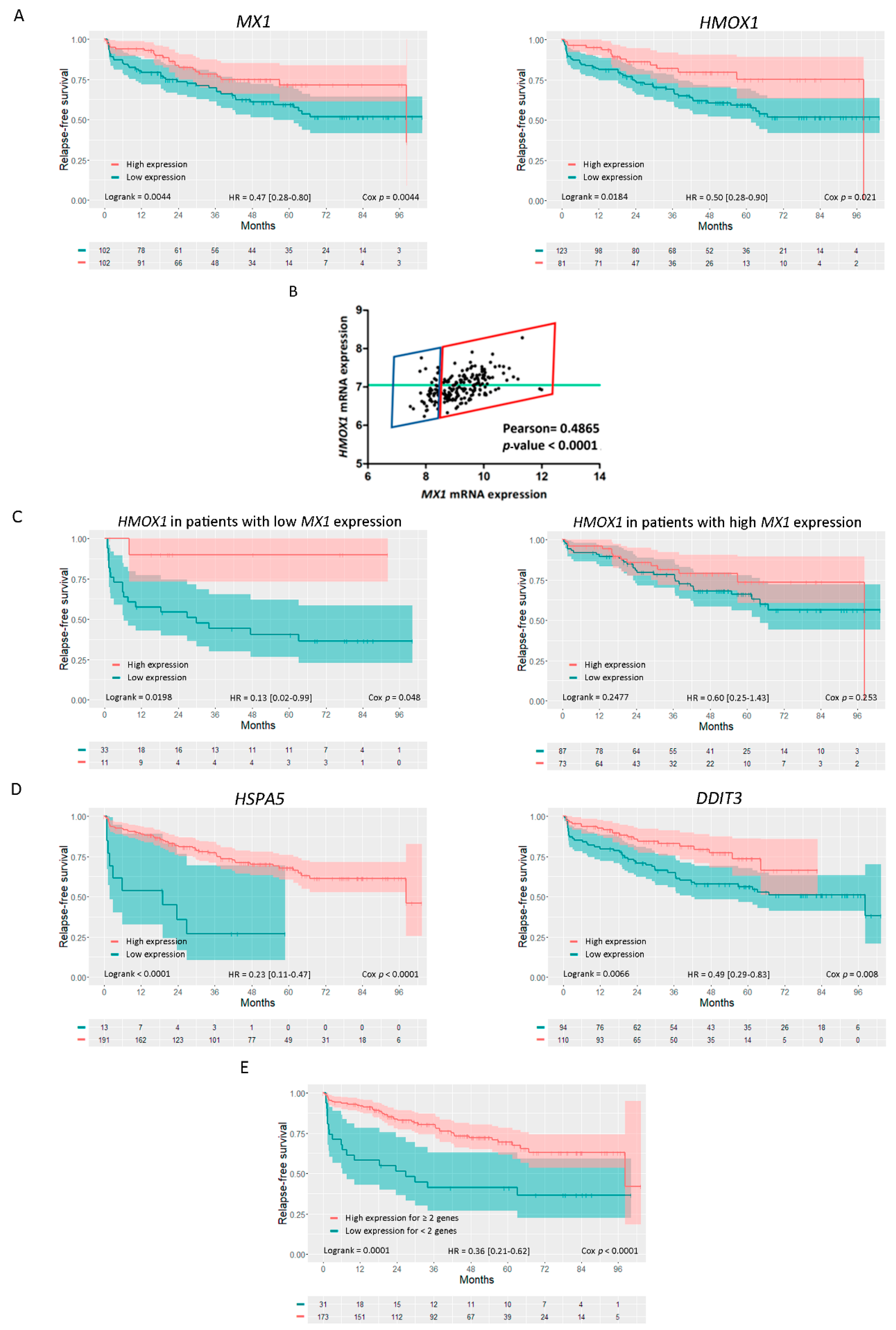
3.9. Analysis of HMOX1, MX1, and Endoplasmic Reticulum Stress Genes as Risk Predictors of Clinical Outcome in Prostate Cancer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dulak, J.; Jozkowicz, A. Novel Faces of Heme Oxygenase-1: Mechanisms and Therapeutic Potentials. Antioxidants Redox Signal. 2014, 20, 1673–1676. [Google Scholar] [CrossRef] [Green Version]
- Chau, L.-Y. Heme oxygenase-1: Emerging target of cancer therapy. J. Biomed. Sci. 2015, 22, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitti, M.; Piras, S.; Marinari, U.M.; Moretta, L.; Pronzato, M.A.; Furfaro, A.L. HO-1 Induction in Cancer Progression: A Matter of Cell Adaptation. Antioxidants 2017, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Gueron, G.; De Siervi, A.; Ferrando, M.; Salierno, M.; De Luca, P.; Elguero, B.; Meiss, R.; Navone, N.; Vazquez, E. Critical Role of Endogenous Heme Oxygenase 1 as a Tuner of the Invasive Potential of Prostate Cancer Cells. Mol. Cancer Res. 2009, 7, 1745–1755. [Google Scholar] [CrossRef] [Green Version]
- Ferrando, M.; Gueron, G.; Elguero, B.; Giudice, J.; Salles, A.; Leskow, F.C.; Jares-Erijman, E.A.; Colombo, L.; Meiss, R.; Navone, N.; et al. Heme oxygenase 1 (HO-1) challenges the angiogenic switch in prostate cancer. Angiogenesis 2011, 14, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Gueron, G.; Giudice, J.; Valacco, P.; Paez, A.; Elguero, B.; Toscani, M.; Jaworski, F.; Leskow, F.C.; Cotignola, J.; Martí, M.A.; et al. Heme-oxygenase-1 implications in cell morphology and the adhesive behavior of prostate cancer cells. Oncotarget 2014, 5, 4087–4102. [Google Scholar] [CrossRef] [Green Version]
- Paez, A.; Vazquez, E.; Gueron, G. Heme oxygenase 1 governs the cytoskeleton at filopodia: Pulling the brakes on the migratory capacity of prostate tumoral cells. Cell Death Discov. 2017, 3, 17020. [Google Scholar] [CrossRef] [Green Version]
- Paez, A.V.; Pallavicini, C.; Schuster, F.; Valacco, M.P.; Giudice, J.; Ortiz, E.G.; Anselmino, N.; Labanca, E.; Binaghi, M.; Salierno, M.; et al. Heme oxygenase-1 in the forefront of a multi-molecular network that governs cell-cell contacts and filopodia-induced zippering in prostate cancer. Cell Death Dis. 2016, 7, e2570. [Google Scholar] [CrossRef]
- Łoboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [Green Version]
- Schönthal, A.H. Endoplasmic Reticulum Stress: Its Role in Disease and Novel Prospects for Therapy. Scientifica 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic Reticulum Stress and Oxidative Stress: A Vicious Cycle or a Double-Edged Sword? Antioxidants Redox Signal. 2007, 9, 2277–2294. [Google Scholar] [CrossRef] [Green Version]
- McCullough, K.D.; Martindale, J.L.; Klotz, L.-O.; Aw, T.-Y.; Holbrook, N.J. Gadd153 Sensitizes Cells to Endoplasmic Reticulum Stress by Down-Regulating Bcl2 and Perturbing the Cellular Redox State. Mol. Cell. Boil. 2001, 21, 1249–1259. [Google Scholar] [CrossRef] [Green Version]
- Rutkowski, D.T.; Arnold, S.M.; Miller, C.N.; Wu, J.; Li, J.; Gunnison, K.M.; Mori, K.; Akha, A.A.S.; Raden, D.; Kaufman, R.J. Adaptation to ER Stress Is Mediated by Differential Stabilities of Pro-Survival and Pro-Apoptotic mRNAs and Proteins. PLoS Boil. 2006, 4, e374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlovic, J.; Zürcher, T.; Haller, O.; Staeheli, P. Resistance to influenza virus and vesicular stomatitis virus conferred by expression of human MxA protein. J. Virol. 1990, 64, 3370–3375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochs, G.; Janzen, C.; Hohenberg, H.; Haller, O. Antivirally active MxA protein sequesters La Crosse virus nucleocapsid protein into perinuclear complexes. Proc. Natl. Acad. Sci. USA 2002, 99, 3153–3158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haller, O.; Staeheli, P.; Schwemmle, M.; Kochs, G. Mx GTPases: Dynamin-like antiviral machines of innate immunity. Trends Microbiol. 2015, 23, 154–163. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Anselmino, N.; Bizzotto, J.; Sanchis, P.; Lage-Vickers, S.; Ortiz, E.; Valacco, P.; Paez, A.; Labanca, E.; Meiss, R.; Navone, N.; et al. HO-1 Interactors Involved in the Colonization of the Bone Niche: Role of ANXA2 in Prostate Cancer Progression. Biomolecules 2020, 10, 467. [Google Scholar] [CrossRef] [Green Version]
- Ross-Adams, H.; Lamb, A.D.; Dunning, M.; Halim, S.; Lindberg, J.; Massie, C.E.; Egevad, L.; Russell, R.; Ramos-Montoya, A.; Vowler, S.; et al. Integration of copy number and transcriptomics provides risk stratification in prostate cancer: A discovery and validation cohort study. EBioMedicine 2015, 2, 1133–1144. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Kosinski, M.; Biecek, P. survminer: Drawing Survival Curves using “ggplot2” 2019. Computer program.
- Budczies, J.; Klauschen, F.; Sinn, B.V.; Győrffy, B.; Schmitt, W.; Darb-Esfahani, S.; Denkert, C. Cutoff Finder: A Comprehensive and Straightforward Web Application Enabling Rapid Biomarker Cutoff Optimization. PLoS ONE 2012, 7, e51862. [Google Scholar] [CrossRef] [Green Version]
- Bussolati, B.; Ahmed, A.; Pemberton, H.; Landis, R.C.; Di Carlo, F.; Haskard, D.; Mason, J.C. Bifunctional role for VEGF-induced heme oxygenase-1 in vivo: Induction of angiogenesis and inhibition of leukocytic infiltration. Blood 2004, 103, 761–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegiel, B.; Chin, B.Y.; Otterbein, L.E. Inhale to survive, cycle or die? Carbon monoxide and cellular proliferation. Cell Cycle 2008, 7, 1379–1384. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, J.N.; The Cancer Genome Atlas Research Network; Li, J.; Mills, G.B.; Shaw, K.R.M.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Wang, M.; Kaufman, R.J. The impact of the endoplasmic reticulum protein-folding environment on cancer development. Nat. Rev. Cancer 2014, 14, 581–597. [Google Scholar] [CrossRef]
- Verfaillie, T.; Garg, A.D.; Agostinis, P. Targeting ER stress induced apoptosis and inflammation in cancer. Cancer Lett. 2013, 332, 249–264. [Google Scholar] [CrossRef]
- Jozkowicz, A.; Was, H.; Dulak, J. Heme Oxygenase-1 in Tumors: Is It a False Friend? Antioxidants Redox Signal. 2007, 9, 2099–2118. [Google Scholar] [CrossRef] [Green Version]
- Saccà, P.; Meiss, R.; Casas, G.; Mazza, O.; Calvo, J.C.; Navone, N.; Vazquez, E. Nuclear translocation of haeme oxygenase-1 is associated to prostate cancer. Br. J. Cancer 2007, 97, 1683–1689. [Google Scholar] [CrossRef] [PubMed]
- Jaworski, F.M.; Gentilini, L.; Gueron, G.; Meiss, R.P.; Ortiz, E.G.; Berguer, P.M.; Ahmed, A.; Navone, N.; Rabinovich, G.A.; Compagno, D.; et al. In Vivo Hemin Conditioning Targets the Vascular and Immunologic Compartments and Restrains Prostate Tumor Development. Clin. Cancer Res. 2017, 23, 5135–5148. [Google Scholar] [CrossRef] [Green Version]
- Elguero, B.; Gueron, G.; Giudice, J.; A Toscani, M.; De Luca, P.; Zalazar, F.; Leskow, F.C.; Meiss, R.; Navone, N.; De Siervi, A.; et al. Unveiling the Association of STAT3 and HO-1 in Prostate Cancer: Role beyond Heme Degradation. Neoplasia 2012, 14, 1043–1056. [Google Scholar] [CrossRef] [Green Version]
- Mushinski, J.F.; Nguyen, P.; Stevens, L.M.; Khanna, C.; Lee, S.; Chung, E.J.; Lee, M.-J.; Kim, Y.S.; Linehan, W.M.; Horisberger, M.A.; et al. Inhibition of Tumor Cell Motility by the Interferon-inducible GTPase MxA. J. Boil. Chem. 2009, 284, 15206–15214. [Google Scholar] [CrossRef] [Green Version]
- A Horisberger, M. Interferon-induced human protein MxA is a GTPase which binds transiently to cellular proteins. J. Virol. 1992, 66, 4705–4709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagano, K.; Masters, J.R.; Akpan, A.; Yang, A.; Corless, S.; Wood, C.; Hastie, C.; Zvelebil, M.; Cramer, R.; Naaby-Hansen, S. Differential protein synthesis and expression levels in normal and neoplastic human prostate cells and their regulation by type I and II interferons. Oncogene 2003, 23, 1693–1703. [Google Scholar] [CrossRef] [Green Version]
- Kotenko, S.V.; Gallagher, G.; Baurin, V.V.; Lewis-Antes, A.; Shen, M.; Shah, N.K.; Langer, J.; Sheikh, F.; Dickensheets, H.; Donnelly, R.P. IFN-λs mediate antiviral protection through a distinct class II cytokine receptor complex. Nat. Immunol. 2002, 4, 69–77. [Google Scholar] [CrossRef]
- Jonasch, E.; Haluska, F.G. Interferon in Oncological Practice: Review of Interferon Biology, Clinical Applications, and Toxicities. Oncologist 2001, 6, 34–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shou, J.; Soriano, R.; Hayward, S.W.; Cunha, G.R.; Williams, P.M.; Gao, W.-Q. Expression profiling of a human cell line model of prostatic cancer reveals a direct involvement of interferon signaling in prostate tumor progression. Proc. Natl. Acad. Sci. USA 2002, 99, 2830–2835. [Google Scholar] [CrossRef] [Green Version]
- Schulz, W.A.; Alexa, A.; Jung, V.; Hader, C.; Hoffmann, M.J.; Yamanaka, M.; Fritzsche, S.; Wlazlinski, A.; Müller, M.; Lengauer, T.; et al. Factor interaction analysis for chromosome 8 and DNA methylation alterations highlights innate immune response suppression and cytoskeletal changes in prostate cancer. Mol. Cancer 2007, 6, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomlins, S.; Laxman, B.; Varambally, S.; Cao, X.; Yu, J.; Helgeson, B.E.; Cao, Q.; Prensner, J.R.; Rubin, M.A.; Shah, R.B.; et al. Role of the TMPRSS2-ERG Gene Fusion in Prostate Cancer. Neoplasia 2008, 10, 177-IN9. [Google Scholar] [CrossRef] [Green Version]
- Attard, G.; Clark, J.; Ambroisine, L.; Fisher, G.; Kovacs, G.; Flohr, P.; Berney, D.M.; Foster, C.S.; Fletcher, A.; Gerald, W.L.; et al. Duplication of the fusion of TMPRSS2 to ERG sequences identifies fatal human prostate cancer. Oncogene 2007, 27, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Isler, J.A.; Skalet, A.H.; Alwine, J.C. Human Cytomegalovirus Infection Activates and Regulates the Unfolded Protein Response. J. Virol. 2005, 79, 6890–6899. [Google Scholar] [CrossRef] [Green Version]
- Haruki, A.N.; Naito, T.; Nishie, T.; Saito, S.; Nagata, K. Interferon-Inducible Antiviral Protein MxA Enhances Cell Death Triggered by Endoplasmic Reticulum Stress. J. Interf. Cytokine Res. 2011, 31, 847–856. [Google Scholar] [CrossRef]
- Medigeshi, G.R.; Lancaster, A.M.; Hirsch, A.J.; Briese, T.; Lipkin, W.I.; DeFilippis, V.; Früh, K.; Mason, P.W.; Nikolich-Žugich, J.; Nelson, J.A. West Nile Virus Infection Activates the Unfolded Protein Response, Leading to CHOP Induction and Apoptosis. J. Virol. 2007, 81, 10849–10860. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Dataset | Platform | Measured Genes | N° Samples | GEO Accession |
|---|---|---|---|---|---|
| 1 | Arreduani Prostate | HG U133 Plus 2.0 Array | 19,574 | 21 | GSE55945 |
| 2 | Grasso Prostate | Agilent Human Genome 44K | 19,189 | 122 | GSE35988 |
| 3 | Holzbeierlein Prostate | HG U95A-Av2 Array | 8603 | 54 | N/A |
| 4 | Lapointe Prostate | Undefined Platform | 10,166 | 112 | GSE3933 |
| 5 | LaTulippe Prostate | HG U95A-Av2 Array | 8603 | 35 | GSE68882 |
| 6 | Liu Prostate | HG U133A Array | 12,624 | 57 | N/A |
| 7 | Luo Prostate | Hu35k (A-D) and HG U95A-Av2 array | 15,302 | 30 | GSE68545 |
| 8 | Magee Prostate | HumanGeneFL Array | 5338 | 15 | N/A |
| 9 | Singh Prostate | HG U95A-Av2 Array | 8603 | 102 | GSE68907 |
| 10 | Taylor Prostate | Undefined Platform | 22,238 | 185 | GSE21032 |
| 11 | Tomlins Prostate | Undefined Platform | 10,656 | 101 | GSE6099 |
| 12 | Vanaja Prostate | HG U133(A-B) Array | 17,779 | 40 | N/A |
| 13 | Varambally Prostate | HG U133 Plus 2.0 Array | 19,574 | 19 | GSE3325 |
| 14 | Wallace Prostate | HG U133 2.0 Array | 12,603 | 89 | GSE6956 |
| 15 | Welsh Prostate | HG U95A-Av2 Array | 8603 | 34 | N/A |
| 16 | Yu Prostate | HG U95A-Av2 Array | 8603 | 112 | GSE68555 |
| EXPRESSION | SURVIVAL | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Oncomine | TCGA | Ross Adams | |||||||
| Expression Cluster | Gene | Over- or Under-Expression | Median Rank | p-Value | Over- or Under-Expression | Fold Change | p-Value | Cox p- Value | Hazards Ratio (IC95) |
| A | CBX3 |  | 825 (top 20%) | 4.21 × 10−04 |  | 1.312 | 8.67 × 10−12 | 0.085 | 0.65 (0.4–1.06) |
| RCC1 * |  | 487 (top 15%) | 0.012 |  | 1.74 | 6.48 × 10−31 | 0.034 | 4.85 (1.20–2.09) | |
| TMOD3 |  | 2035 (top 22%) | 0.016 |  | 1.152 | 9.23 × 10−05 | 0.001 | 0.42 (0.26–0.69) | |
| NOA1 |  | 203 (top 3%) | 3.76 × 10−06 |  | 1.762 | 9.53 × 10−40 | … | … | |
| EEF2 |  | 299 (top 24%) | 1.06 × 10−05 |  | 2.43 | 1.31 × 10−73 | 0.0002 | 0.30 (0.15–0.59) | |
| ASPH * |  | 798 (top 9%) | 0.003 |  | 0.613 | 1.87 × 10−13 | 0.001 | 0.34 (0.18–0.66) | |
| HSPB1* |  | 182.5 (top 11%) | 1.06 × 10−04 |  | 0.584 | 9.95 × 10−24 | 0.001 | 0.36 (0.20–0.66) | |
| SQSTM1 * |  | 1581 (top 35%) | 0.003 |  | 0.961 | 9.39× 10−01 | 0.009 | 0.40 (0.2–0.79) | |
| ANXA2 * |  | 133 (top 16%) | 0.003 |  | 0.496 | 4.48 × 10−19 | 0.001 | 0.40 (0.23–0.70) | |
| GSN * |  | 586.5 (top 10%) | 0.005 |  | 0.533 | 3.4 × 10−21 | 0.001 | 0.27 (0.12–0.60) | |
| B | TOP1 |  | 1082 (top 25%) | 0.003 |  | 1.468 | 5.58 × 10−14 | 0.005 | 0.48 (0.29–0.80) |
| PDCD5 * |  | 836.5 (top 22%) | 0.011 |  | 1.164 | 2.41 × 10−05 | 0.0001 | 5.80 (2.72–12.32) | |
| THRAP3 |  | 1898 (top 24%) | 0.002 |  | 1.073 | 5.84 × 10−05 | 0.004 | 0.49 (0.3–0.8) | |
| SLC12A7 |  | 1914 (top 25%) | 5.5 × 10−04 |  | 0.849 | 3.86 × 10−01 | 0.001 | 0.39(0.19–0.65) | |
| PURA * |  | 1280 (top 16%) | 0.031 |  | 0.951 | 1.87 × 10−01 | 0.001 | 0.41 (0.25–0.69) | |
| RPA1 |  | 2093 (top 23%) | 7.71× 10−04 |  | 1.246 | 5.39 × 10−06 | 0.013 | 2 (1.16–3.47) | |
| SFSR3 |  | 793 (top 11%) | 0.01 | N/A | … | … | 0.024 | 1.77 (1.08–2.91) | |
| MX1 * |  | 1542 (top 19%) | 0.031 |  | 0.578 | 1.04 × 10−07 | 0.0044 | 0.47 (0.28–0.80) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortiz, E.; Sanchis, P.; Bizzotto, J.; Lage-Vickers, S.; Labanca, E.; Navone, N.; Cotignola, J.; Vazquez, E.; Gueron, G. Myxovirus Resistance Protein 1 (MX1), a Novel HO-1 Interactor, Tilts the Balance of Endoplasmic Reticulum Stress towards Pro-Death Events in Prostate Cancer. Biomolecules 2020, 10, 1005. https://doi.org/10.3390/biom10071005
Ortiz E, Sanchis P, Bizzotto J, Lage-Vickers S, Labanca E, Navone N, Cotignola J, Vazquez E, Gueron G. Myxovirus Resistance Protein 1 (MX1), a Novel HO-1 Interactor, Tilts the Balance of Endoplasmic Reticulum Stress towards Pro-Death Events in Prostate Cancer. Biomolecules. 2020; 10(7):1005. https://doi.org/10.3390/biom10071005
Chicago/Turabian StyleOrtiz, Emiliano, Pablo Sanchis, Juan Bizzotto, Sofia Lage-Vickers, Estefania Labanca, Nora Navone, Javier Cotignola, Elba Vazquez, and Geraldine Gueron. 2020. "Myxovirus Resistance Protein 1 (MX1), a Novel HO-1 Interactor, Tilts the Balance of Endoplasmic Reticulum Stress towards Pro-Death Events in Prostate Cancer" Biomolecules 10, no. 7: 1005. https://doi.org/10.3390/biom10071005