Wheat Yellow Mosaic Virus NIb Interacting with Host Light Induced Protein (LIP) Facilitates Its Infection through Perturbing the Abscisic Acid Pathway in Wheat

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Plasmids
2.2. Phylogenetic Analysis and Promoter Cis-Acting Element Prediction Analysis
2.3. Yeast Two-Hybrid Assay
2.4. Sub-Cellular Localization, Bimolecular Fluorescence Complementation and Co-Immunoprecipitation (Co-IP) Assays
2.5. Plant RNA Isolation and Quantitative Real Time PCR Analysis
2.6. Virus-Induced Gene Silencing
2.7. Mechanical Friction Inoculation of Barley stripe mosaic virus and Wheat yellow mosaic virus
3. Results
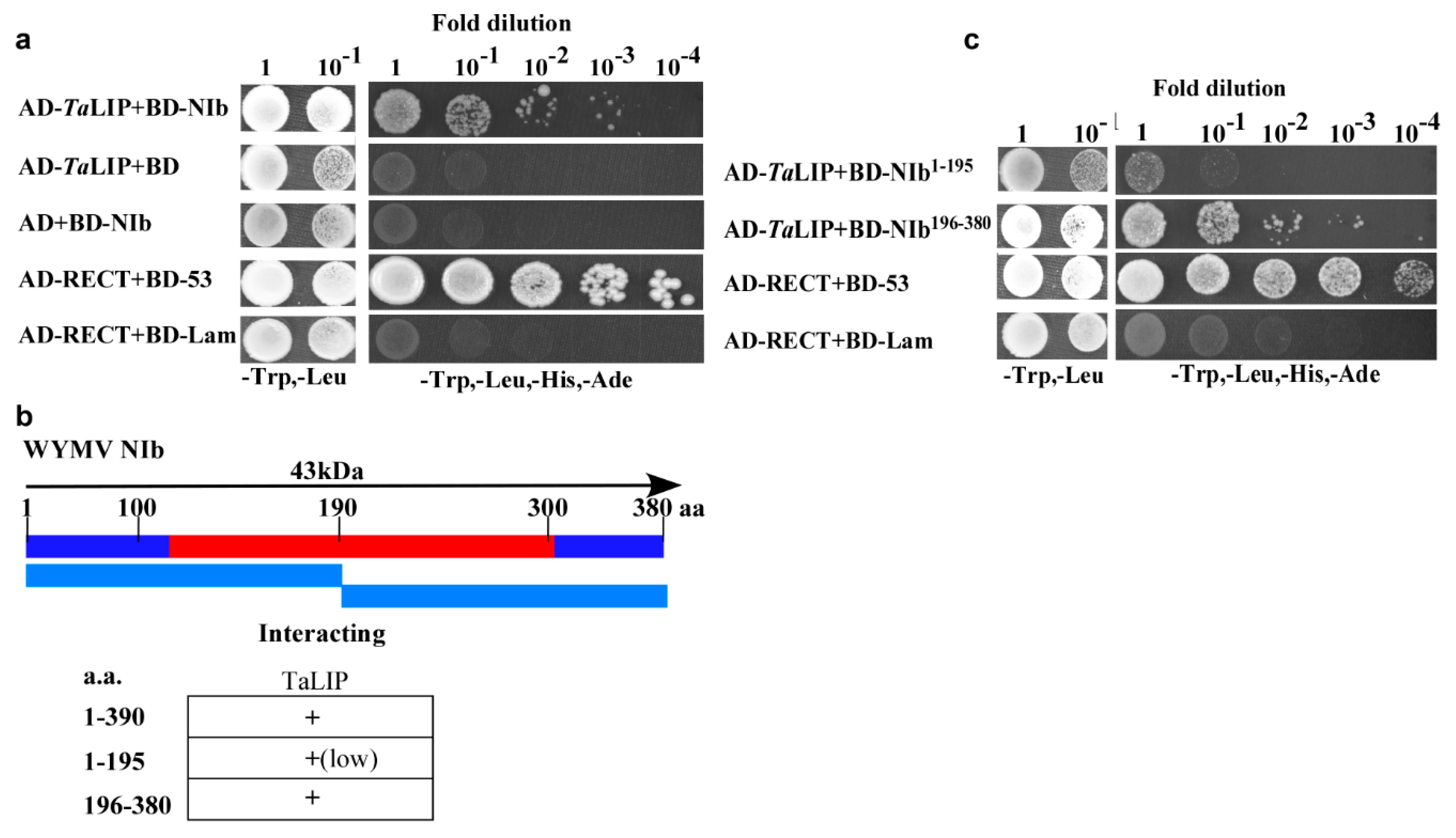
3.1. TaLIP Interacts with Wheat Yellow Mosaic Virus NIb and C-Terminus of NIb196–380aa Is the Major Region for This Interaction
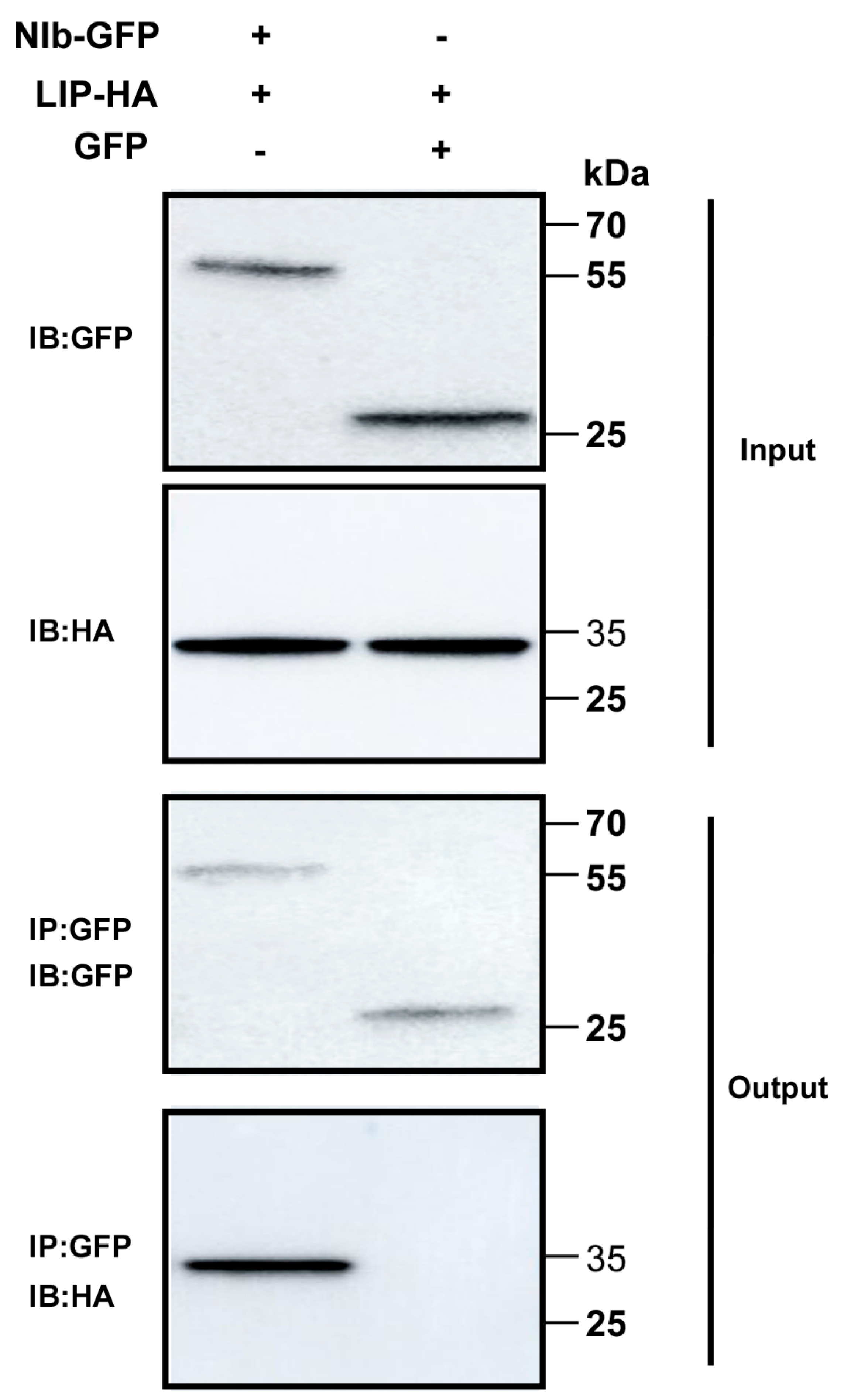
3.2. TaLIP Interacts with WYMV NIb In Vivo
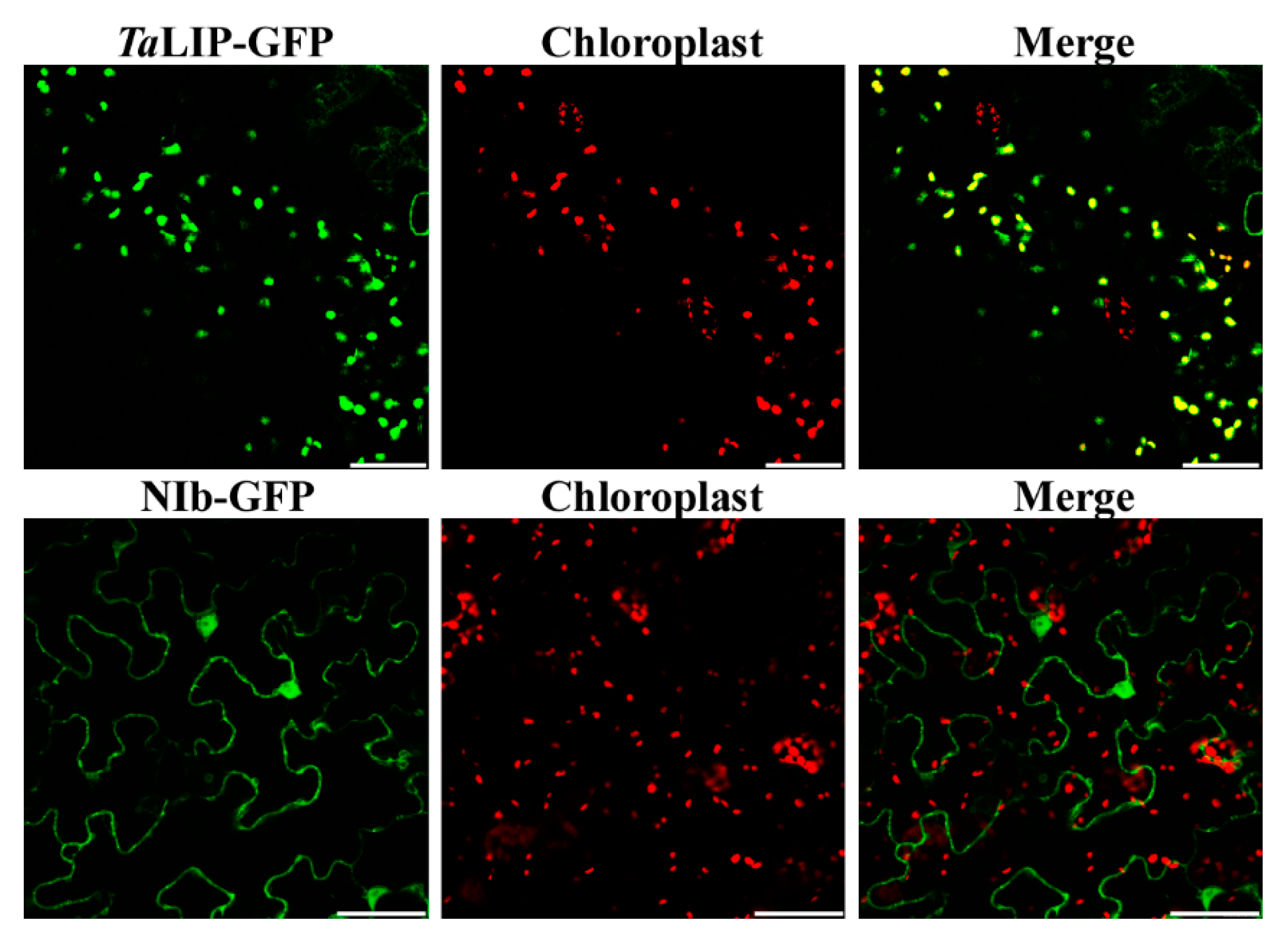
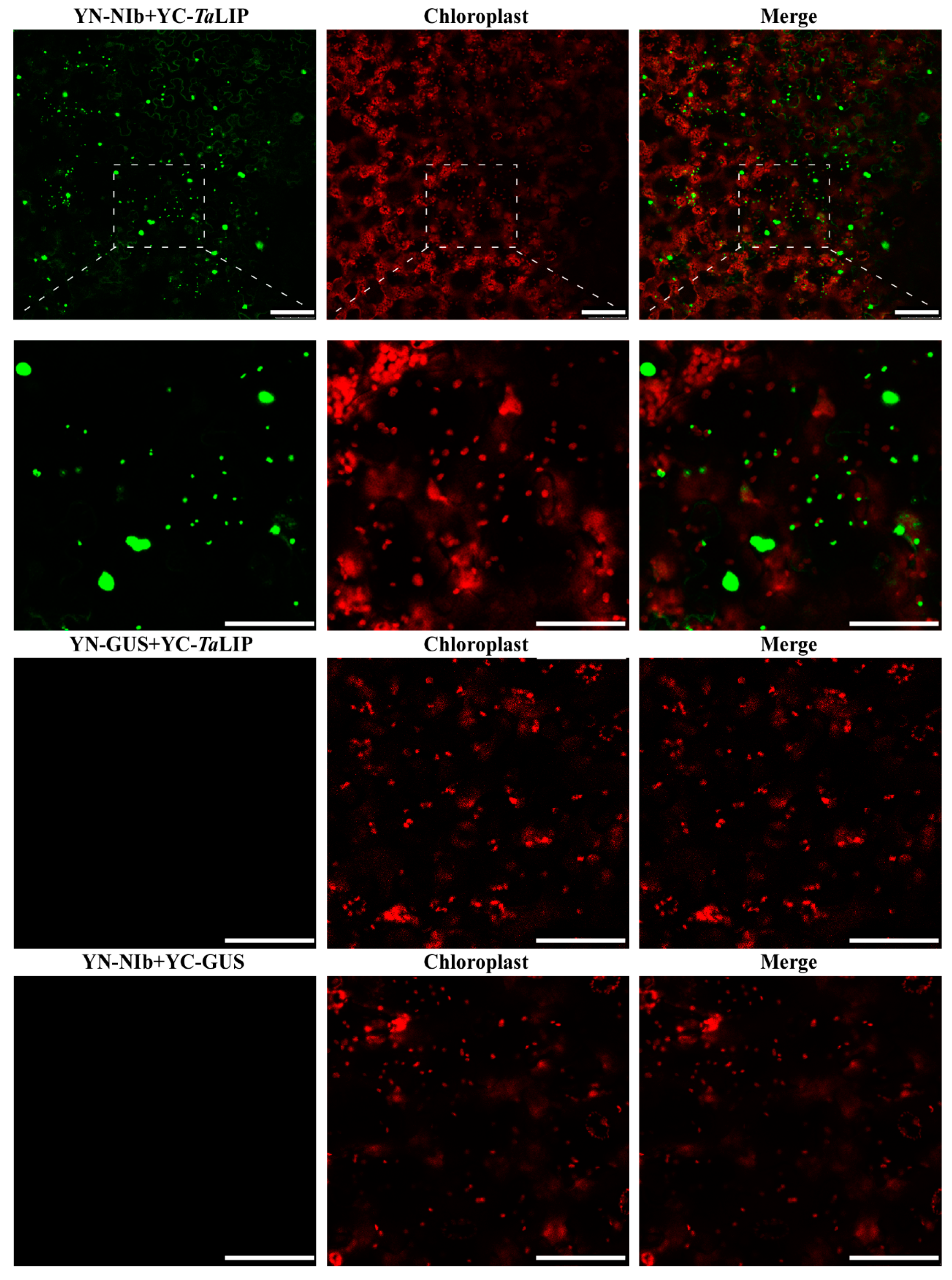
3.3. Sub-Cellular Localization of WYMV NIb and TaLIP Is Altered by Their Interaction in Nicotiana Benthamiana
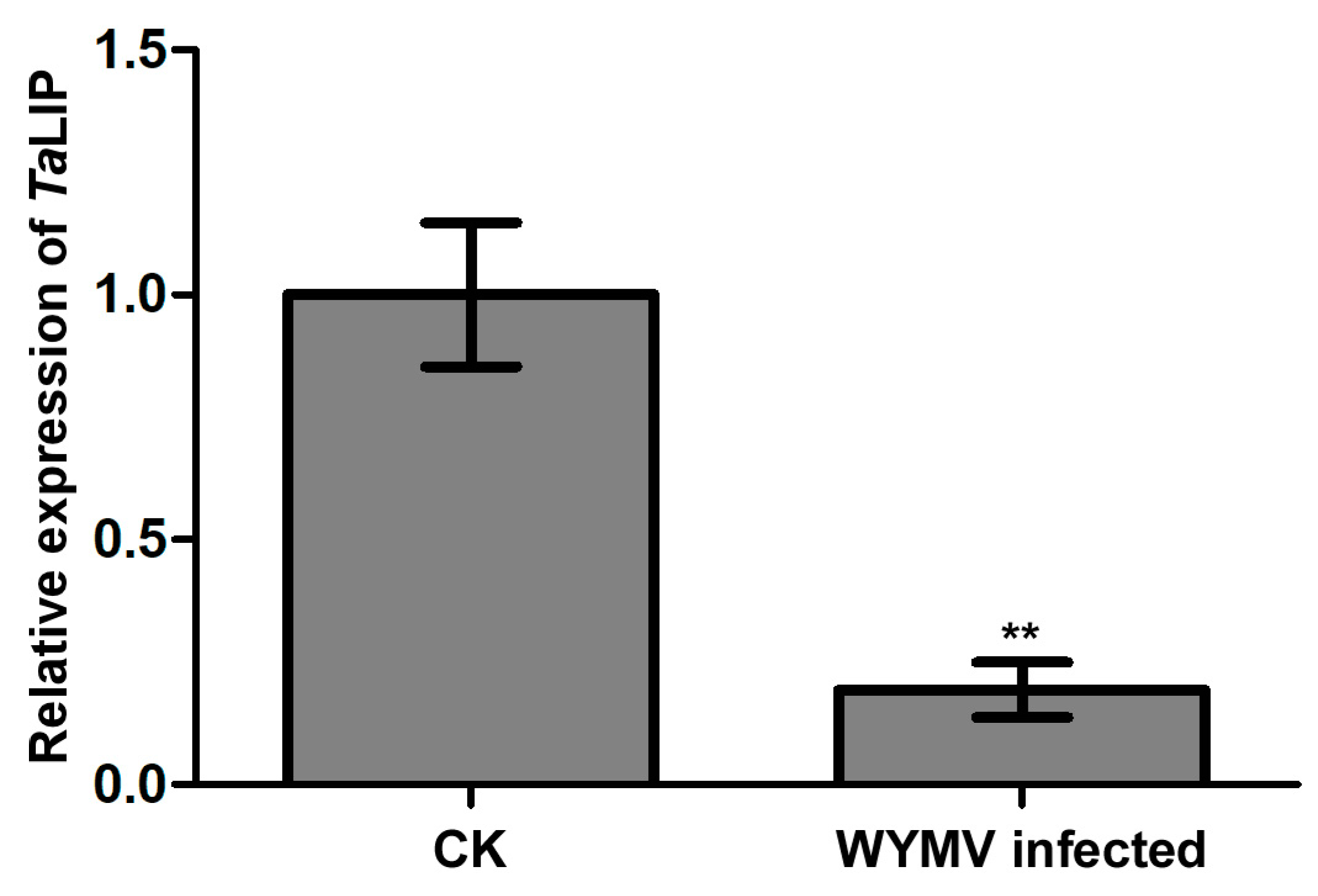
3.4. The Transcriptional Level of TaLIP Is Downregulated in WYMV-Infected Wheat
3.5. Knockdown the TaLIP Facilitate WYMV Infection in Wheat
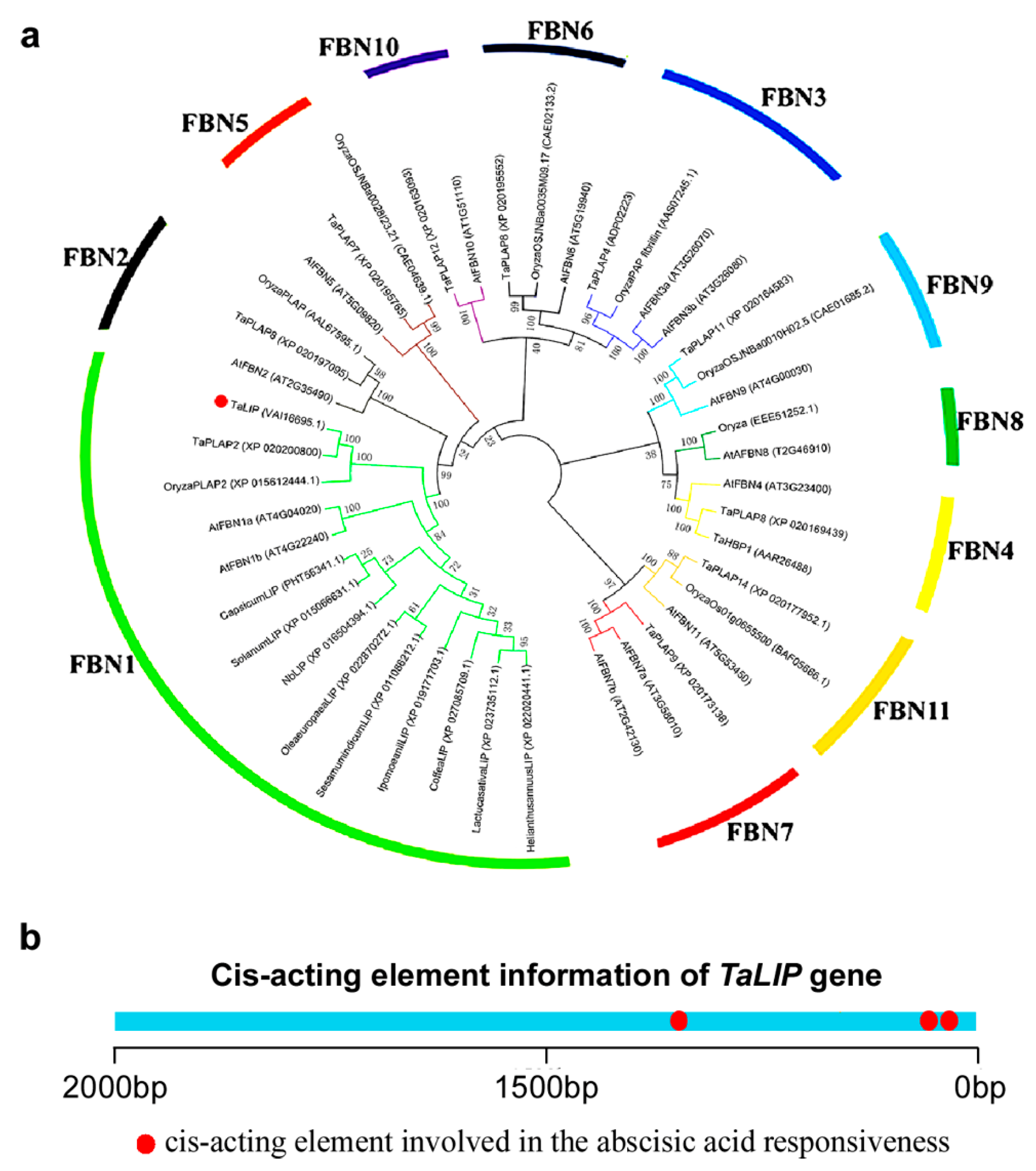
3.6. TaLIP Belongs to the FBN1 Subspecies of the Fibrillin Family and Has Three Copies of ABA Responsive Promoter Element
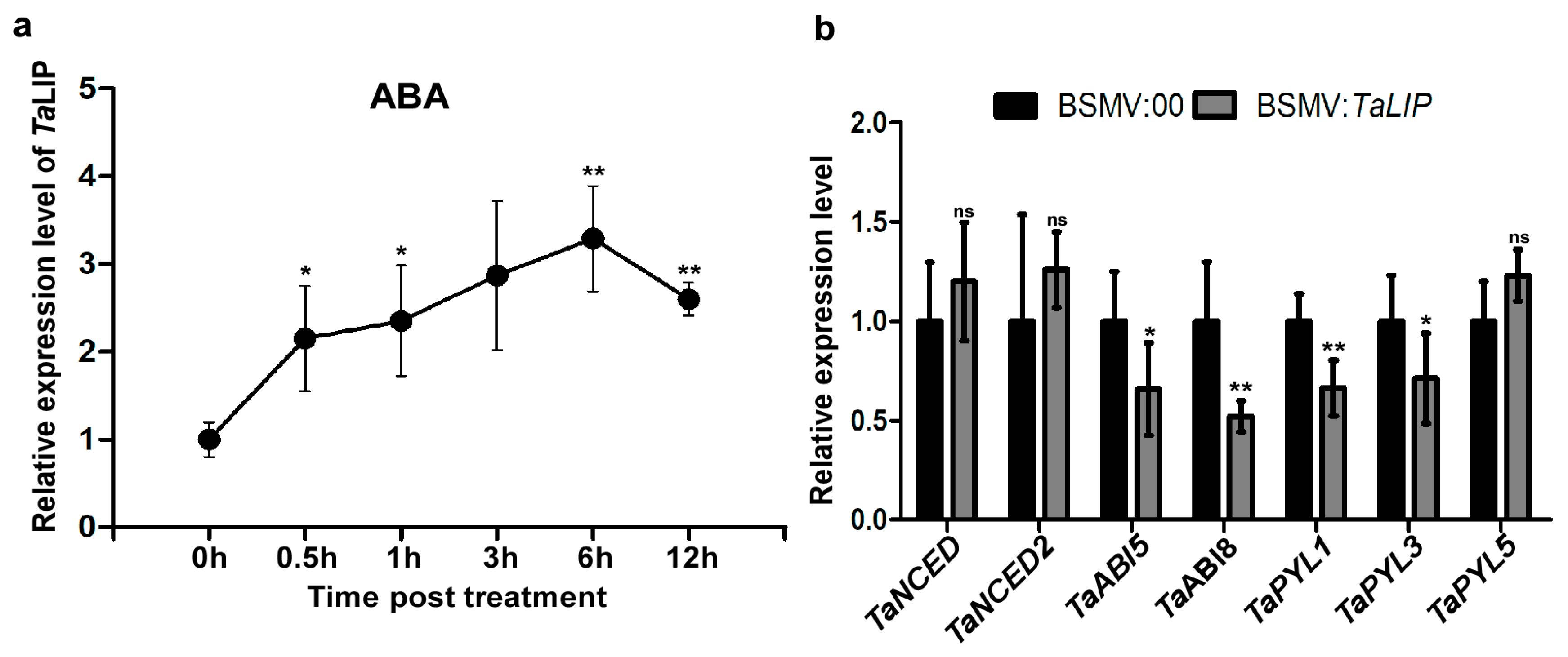
3.7. TaLIP Is Responsive to External ABA Stimuli, Silencing the TaLIP Gene Suppressing the ABA Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sanfaçon, H.J. Grand challenge in plant virology: Understanding the impact of plant viruses in model plants, in agricultural crops, and in complex ecosystems. Front. Microbiol. 2017, 8, 860. [Google Scholar] [CrossRef] [PubMed]
- García, J.A.; Pallás, V.J. Viral factors involved in plant pathogenesis. Curr. Opin. Virol. 2015, 11, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cui, H.; Cui, X.; Wang, A.J. The altered photosynthetic machinery during compatible virus infection. Curr. Opin. Virol. 2016, 17, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.J. Dissecting the molecular network of virus-plant interactions: The complex roles of host factors. Annu. Rev. Phytopathol. 2015, 53, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Torrance, L.; Andreev, I.A.; Gabrenaite-Verhovskaya, R.; Cowan, G.; Mäkinen, K.; Taliansky, M.E.J. An unusual structure at one end of potato potyvirus particles. J. Mol. Biol. 2006, 357, 1–8. [Google Scholar] [CrossRef]
- Arturo, D.; Xiaofeng, W.; Paul, A.J. Membrane-shaping host reticulon proteins play crucial roles in viral RNA replication compartment formation and function. Proc. Natl. Acad. Sci. USA 2010, 107, 16291–16296. [Google Scholar] [Green Version]
- Wang, Y.L.; Nagy, P.D.J. Tomato bushy stunt virus Co-Opts the RNA-Binding Function of a Host Metabolic Enzyme for Viral Genomic RNA Synthesis. Cell Host Microbe 2008, 3, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Revers, F.; García, J.A. Molecular biology of potyviruses. Adv. Virus Res. 2015, 92, 101–199. [Google Scholar]
- Park, S.H.; Li, F.; Renaud, J.; Shen, W.; Li, Y.; Guo, L.; Cui, H.; Sumarah, M.; Wang, A. NbEXPA1, an α-expansin, is plasmodesmata-specific and a novel host factor for potyviral infection. Plant J. 2017, 92, 846–861. [Google Scholar] [CrossRef]
- Nakamura, K.; Fushimi, K.; Kouchi, H.; Mihara, K.; Miyazaki, M.; Ohe, T.; Namba, M.J. Inhibitory effects of antioxidants on neonatal rat cardiac myocyte hypertrophy induced by tumor necrosis factor-α and angiotensin II. Circulation 1998, 98, 794–799. [Google Scholar] [CrossRef]
- Xiong, R.J. SCE1, the SUMO-conjugating enzyme in plants that interacts with NIb, the RNA-dependent RNA polymerase of Turnip mosaic virus, is required for viral infection. J. Virol. 2013, 87, 4704–4715. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Xiong, R.; Li, Y.; Li, F.; Zhou, X.; Wang, A.J. Sumoylation of turnip mosaic virus RNA polymerase promotes viral infection by counteracting the host NPR1-mediated immune response. Plant Cell 2017, 29, 508–525. [Google Scholar] [CrossRef]
- Dufresne, P.J.; Thivierge, K.; Cotton, S.; Beauchemin, C.; Ide, C.; Ubalijoro, E.; Laliberté, J.F.; Fortin, M.G.J. Heat shock 70 protein interaction with Turnip mosaic virus RNA-dependent RNA polymerase within virus-induced membrane vesicles. Virology 2008, 374, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.S.; Wei, T.; Laliberté, J.F.; Wang, A.J. A host RNA helicase-like protein, AtRH8, interacts with the potyviral genome-linked protein, VPg, associates with the virus accumulation complex, and is essential for infection. Plant Physiol. 2010, 152, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiong, R.; Bernards, M.; Wang, A.J. Recruitment of Arabidopsis RNA helicase AtRH9 to the viral replication complex by viral replicase to promote turnip mosaic virus replication. Sci. Rep. 2016, 6, 30297. [Google Scholar] [CrossRef] [PubMed]
- Andrés, M.S.A.; Arteaga, M.L.; Ortega, R.G.; Voorrips, R.E. New genes related to PVY resistance in C. annuum L. ‘Serrano Criollo de Morelos-334’. In Proceedings of the Eucarpia Meeting on Genetics & Breeding of Capsicum & Eggplant, Noordwijkerhout, The Netherlands, 17–19 May 2004. [Google Scholar]
- Cook, A.A.J. A mutation for resistance to Potato virus Y in pepper. Phytopathology 1960, 51, 632. [Google Scholar]
- Janzac, B.; Montarry, J.; Palloix, A.; Navaud, O.; Moury, B.J. A point mutation in the polymerase of Potato virus Y confers virulence toward the Pvr4 resistance of pepper and a high competitiveness cost in susceptible cultivar. Mol. Plant Microbe Interact. 2010, 23, 823–830. [Google Scholar] [CrossRef]
- Kyle, M.M.; Palloix, A.J. Proposed revision of nomenclature for potyvirusresistance genes in Capsicum. Euphytica 1997, 97, 183–188. [Google Scholar] [CrossRef]
- Murphy, J.F.; Blauth, J.R.; Livingstone, K.D.; Lackney, V.K.; Jahn, M.K. Trichome density of main stem is tightly linked to PepMoV resistance in chili pepper (Capsicum annuum L.). Mol. Plant Microbe Interact. 2011, 122, 1051–1058. [Google Scholar]
- Chen, M.; Sun, L.; Wu, H.; Chen, J.; Ma, Y.; Zhang, X.; Du, L.; Cheng, S.; Zhang, B.; Ye, X.J. Durable field resistance to Wheat yellow mosaic virus in transgenic wheat containing the antisense virus polymerase gene. Plant Biotechnol. J. 2014, 12, 447–456. [Google Scholar] [CrossRef]
- Sun, L.; Andika, I.B.; Shen, J.; Yang, D.; Chen, J.J. The P2 of Wheat yellow mosaic virus rearranges the endoplasmic reticulum and recruits other viral proteins into replication-associated inclusion bodies. Mol. Plant Pathol. 2014, 15, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.K.; Mcnellis, T.W.J. Fibrillin protein function: The tip of the iceberg? Trends Plant Sci. 2011, 16, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Walter, M.; Chaban, C.; Schutze, K.O.; Weckermann, K.; Nake, C.; Blazevic, D.; Grefen, C.; Schumacher, K.; Oecking, C.J. Visualization of protein interactions in living plant cells using bimolecular fluorescence complementation. Plant J. 2010, 40, 428–438. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Cheng, Y.; Cui, L.; Lijie, Y.; Jackson, A.O.; Zhiyong, L.; Chenggui, H.; Jialin, Y.; Dawei, L.J. A high throughput barley stripe mosaic virus vector for virus induced gene silencing in monocots and dicots. PLoS ONE 2011, 6, e26468. [Google Scholar]
- Holzberg, S.; Brosio, P.; Gross, C.; Pogue, G.P. Barley stripe mosaic virus-induced gene silencing in a monocot plant. Plant J. 2002, 30, 315–327. [Google Scholar] [CrossRef]
- Zhou, T.; Zhang, H.; Lai, T.; Cheng, Q.; Shi, N.; Wang, H.; Jin, M.; Zhong, S.; Fan, Z.; Liu, Y.J. Virus-induced gene complementation reveals a transcription factor network in modulation of tomato fruit ripening. Sci. Rep. 2012, 2, 836. [Google Scholar] [CrossRef]
- Yael, L.D.; Marianna, O.; Elena, S.; Yigal, E.; Dalia, R.D.; Alexander, V.J. Expression and functional analyses of the plastid lipid-associated protein CHRC suggest its role in chromoplastogenesis and stress. Plant Physiol. 2006, 142, 233–244. [Google Scholar]
- Laizet, Y.; Pontier, D.; Mache, R.; Kuntz, M.J. Subfamily organization and phylogenetic origin of genes encoding plastid lipid-associated proteins of the fibrillin type. J. Genome Sci. Technol. 2004, 3, 19–28. [Google Scholar] [CrossRef]
- Bret, C.; Clarke, J.D.; Paul, B.; Joel, K.; Don, H.; Sylvia, P.; Sonia, G.; Molly, D.; Peter, L.; Cinzia, E.J. A network of rice genes associated with stress response and seed development. Proc. Natl. Acad. Sci. USA 2003, 100, 4945–4950. [Google Scholar] [Green Version]
- Singh, D.K.; Maximova, S.N.; Jensen, P.J.; Lehman, B.L.; Ngugi, H.K.; Mcnellis, T.W. FIBRILLIN4 is required for plastoglobule development and stress resistance in apple and Arabidopsis. Plant Physiol. 2010, 154, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Vishnevetsky, M.; Ovadis, M.H.; Levy, M.; Libal, W.Y.; Adam, Z.; Vainstein, A.J. Molecular cloning of a carotenoid-associated protein from Cucumis sativus corollas: Homologous genes involved in carotenoid sequestration in chromoplasts. Plant J. 2010, 10, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sulpice, R.; Himmelbach, A.; Meinhard, M.; Christmann, A.; Grill, E.J. Fibrillin expression is regulated by abscisic acid response regulators and is involved in abscisic acid-mediated photoprotection. Proc. Natl. Acad. Sci. USA 2006, 103, 6061–6066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alazem, M.; Lin, K.Y.; Lin, N.S.J. The abscisic acid pathway has multifaceted effects on the accumulation of Bamboo mosaic virus. Mol. Plant Microbe Interact. 2014, 27, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Fraser, R.S.S.; Whenham, R.J.J. Abscisic acid metabolism in tomato plants infected with tobacco mosaic virus: Relationships with growth, symptoms and the Tm-1 gene for TMV resistance. Physiol. Mol. Plant Pathol. 1989, 34, 215–226. [Google Scholar] [CrossRef]
- Iriti, M.; Faoro, F.J. Abscisic acid is involved in chitosan-induced resistance to tobacco necrosis virus (TNV). Plant Physiol. Biochem. 2008, 46, 1106–1111. [Google Scholar] [CrossRef]
- Deruère, J.; Römer, S.; d’Harlingue, A.; Backhaus, R.A.; Kuntz, M.; Camara, B. Fibril assembly and carotenoid overaccumulation in chromoplasts: A model for supramolecular lipoprotein structures. Plant Cell 1994, 6, 119–133. [Google Scholar]
- Bai, Y.; Zhu, W.; Hu, X.; Sun, C.; Li, Y.; Wang, D.; Wang, Q.; Pei, G.; Zhang, Y.; Guo, A.J. Genome-wide analysis of the bZIP gene family identifies two ABI5-like bZIP transcription factors, BrABI5a and BrABI5b, as positive modulators of ABA signalling in Chinese cabbage. PLoS ONE 2016, 11, e0158966. [Google Scholar] [CrossRef]
- Yang, T.; Yao, S.; Hao, L.; Zhao, Y.; Lu, W.; Xiao, K.J. Wheat bHLH-type transcription factor gene TabHLH1 is crucial in mediating osmotic stresses tolerance through modulating largely the ABA-associated pathway. Plant Cell Rep. 2016, 35, 2309–2323. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Liu, P.; Zhong, K.; Zhang, F.; Xu, M.; He, L.; Jin, P.; Chen, J.; Yang, J. Wheat Yellow Mosaic Virus NIb Interacting with Host Light Induced Protein (LIP) Facilitates Its Infection through Perturbing the Abscisic Acid Pathway in Wheat. Biology 2019, 8, 80. https://doi.org/10.3390/biology8040080
Zhang T, Liu P, Zhong K, Zhang F, Xu M, He L, Jin P, Chen J, Yang J. Wheat Yellow Mosaic Virus NIb Interacting with Host Light Induced Protein (LIP) Facilitates Its Infection through Perturbing the Abscisic Acid Pathway in Wheat. Biology. 2019; 8(4):80. https://doi.org/10.3390/biology8040080
Chicago/Turabian StyleZhang, Tianye, Peng Liu, Kaili Zhong, Fan Zhang, Miaoze Xu, Long He, Peng Jin, Jianping Chen, and Jian Yang. 2019. "Wheat Yellow Mosaic Virus NIb Interacting with Host Light Induced Protein (LIP) Facilitates Its Infection through Perturbing the Abscisic Acid Pathway in Wheat" Biology 8, no. 4: 80. https://doi.org/10.3390/biology8040080