
VTC4 Polyphosphate Polymerase Knockout Increases Stress Resistance of Saccharomyces cerevisiae Cells

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Growth Conditions
2.2. PolyP Extraction and Measurement
2.3. Enzymatic Assay of PolyPs
2.4. PolyP Electrophoresis
2.5. Determination of Yeast Sensitivity to Peroxide, Alkali, and Heavy Metal Ions
2.6. Pi Accumulation
2.7. Quantitative PCR
2.8. Statistics
3. Results
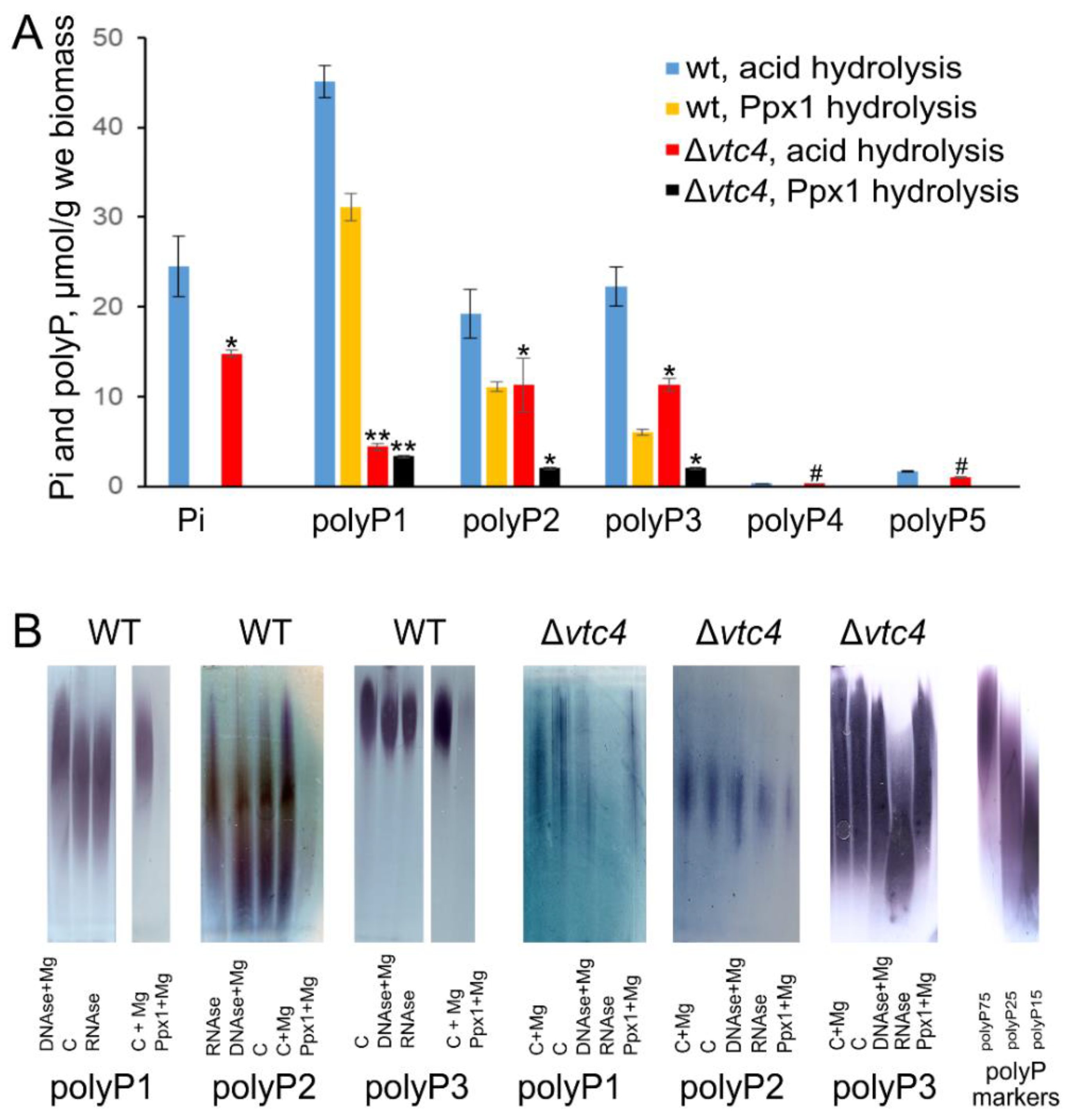
3.1. VTC4 Knockout Strain Has Decreased but Detectable PolyP Level
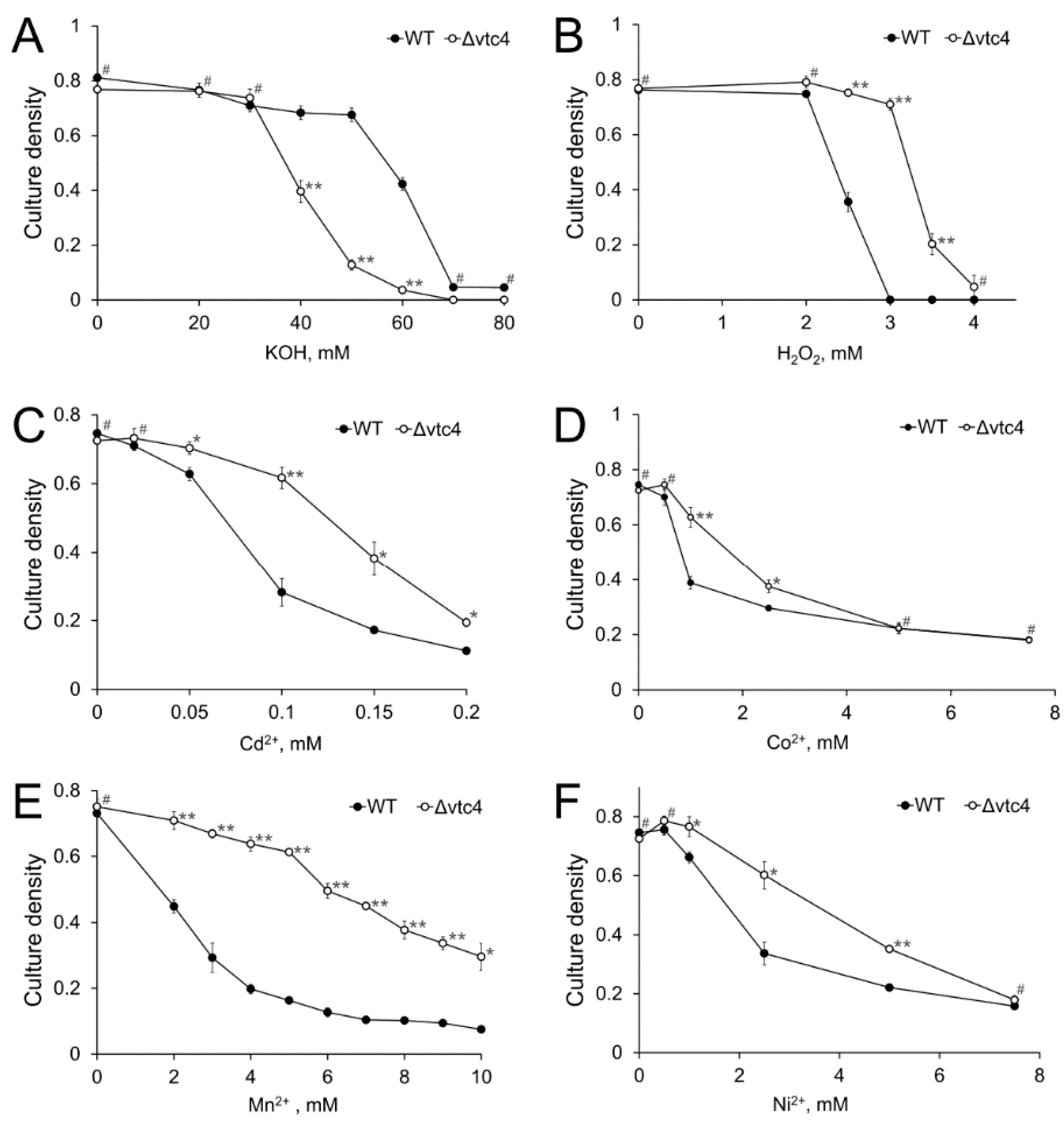
3.2. The Δvtc4 Strain Is More Sensitive to Alkali but Resistant to Peroxide and Heavy Metals
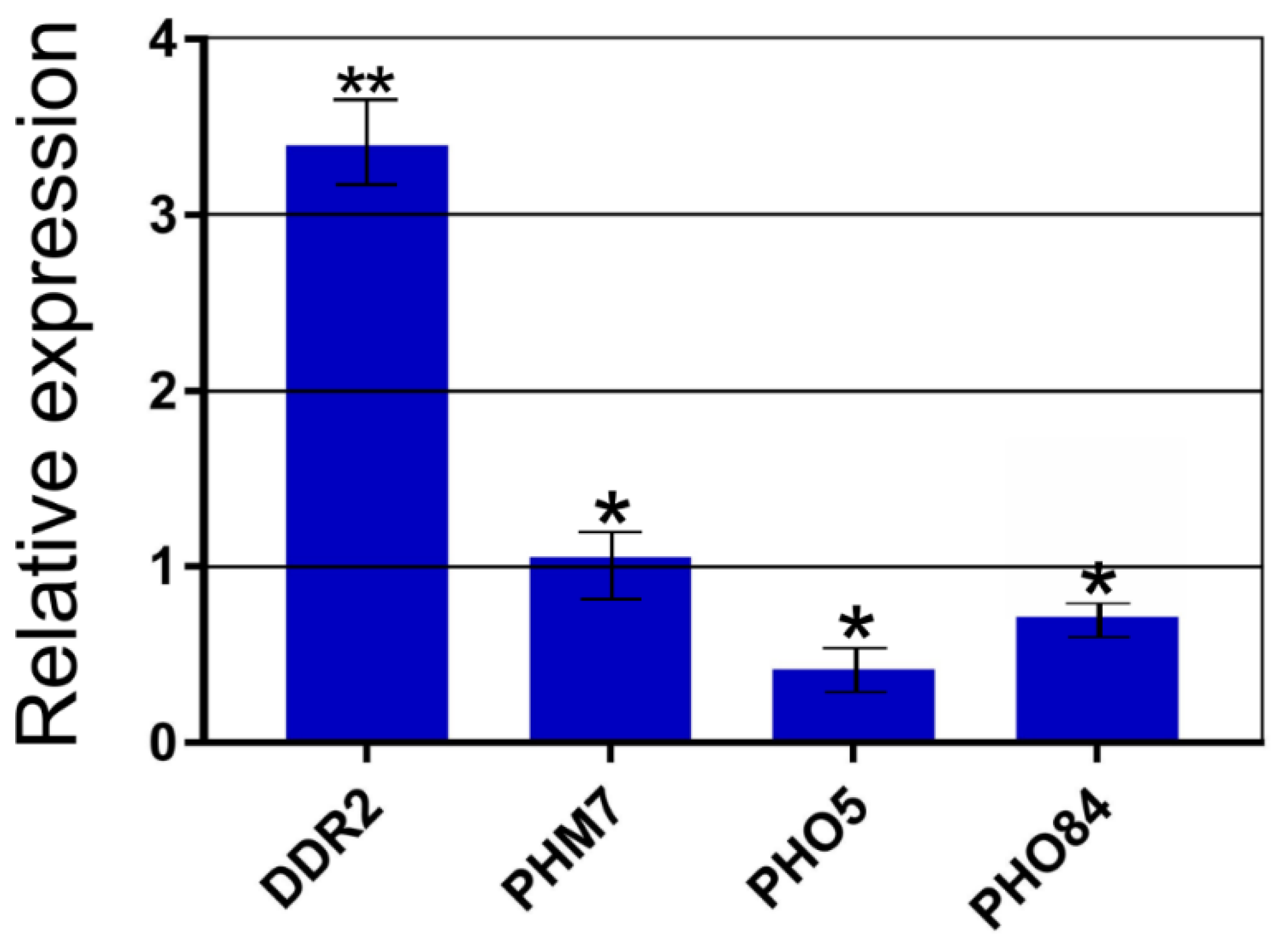
3.3. Differential Expression of Selected Genes Allows to Explain Stress Resistance of Δvtc4 Cells
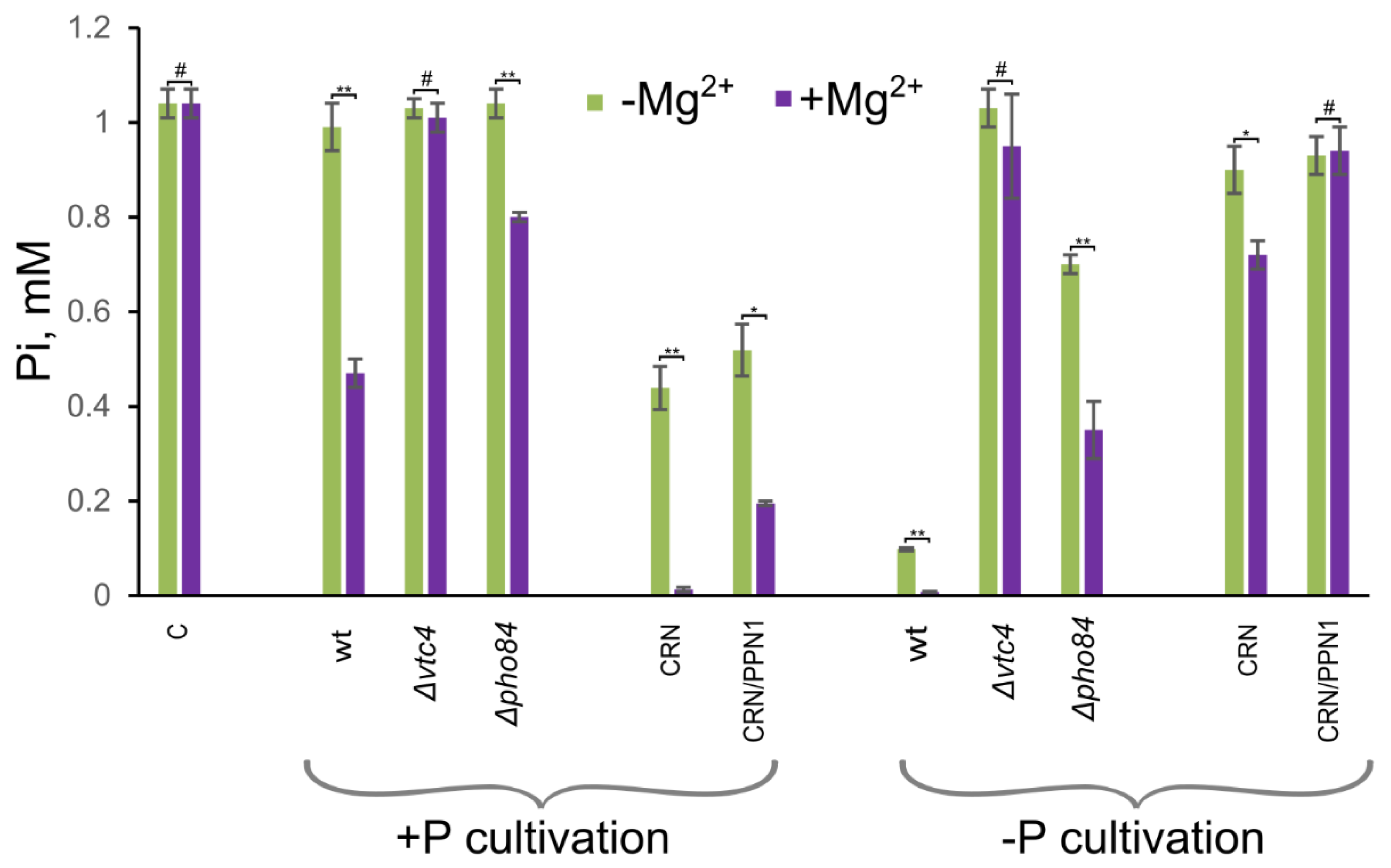
3.4. Δvtc4 Cells Show Decreased Pi Accumulation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rao, N.N.; Gómez-García, M.R.; Kornberg, A. Inorganic Polyphosphate: Essential for Growth and Survival. Annu. Rev. Biochem. 2009, 78, 605–647. [Google Scholar] [CrossRef] [PubMed]
- Albi, T.; Serrano, A. Inorganic polyphosphate in the microbial world. Emerging roles for a multifaceted biopolymer. World J. Microbiol. Biotechnol. 2016, 32, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Jakob, U. Inorganic polyphosphate, a multifunctional polyanionic protein scaffold. J. Biol. Chem. 2019, 294, 2180–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desfougères, Y.; Saiardi, A.; Azevedo, C. Inorganic polyphosphate in mammals: Where’s Wally? Biochem. Soc. Trans. 2020, 48, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, P.; Gosain, T.P.; Singh, M.; Sankhe, G.D.; Arora, G.; Kidwai, S.; Agarwal, S.; Chugh, S.; Saini, D.K.; Singh, R. Inorganic polyphosphate accumulation suppresses the dormancy response and virulence in Mycobacterium tuberculosis. J. Biol. Chem. 2019, 294, 10819–10832. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.J. Interactions between DksA and Stress-Responsive Alternative Sigma Factors Control Inorganic Polyphosphate Accumulation inEscherichia coli. J. Bacteriol. 2020, 202, e00133-20. [Google Scholar] [CrossRef]
- Omelon, S.; Georgiou, J.; Henneman, Z.J.; Wise, L.M.; Sukhu, B.; Hunt, T.; Wynnyckyj, C.; Holmyard, D.; Bielecki, R.; Grynpas, M.D. Control of Vertebrate Skeletal Mineralization by Polyphosphates. PLoS ONE 2009, 4, e5634. [Google Scholar] [CrossRef] [Green Version]
- Müller, W.E.G.; Wang, X.; Schröder, H.C. New Target Sites for Treatment of Osteoporosis. Blue Biotechnology 2017, 55, 187–219. [Google Scholar] [CrossRef]
- Baker, C.J.; Smith, S.A.; Morrissey, J.H. Polyphosphate in thrombosis, hemostasis, and inflammation. Res. Pr. Thromb. Haemost. 2018, 3, 18–25. [Google Scholar] [CrossRef]
- Angelova, P.R.; Iversen, K.Z.; Teschemacher, A.G.; Kasparov, S.; Gourine, A.V.; Abramov, A.Y. Signal transduction in astrocytes: Localization and release of inorganic polyphosphate. Glia 2018, 66, 2126–2136. [Google Scholar] [CrossRef] [Green Version]
- Seidlmayer, L.K.; Juettner, V.V.; Kettlewell, S.; Pavlov, E.V.; Blatter, L.A.; Dedkova, E.N. Distinct mPTP activation mechanisms in ischaemia–reperfusion: Contributions of Ca2+, ROS, pH, and inorganic polyphosphate. Cardiovasc. Res. 2015, 106, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Bru, S.; Martínez-Laínez, J.M.; Hernández-Ortega, S.; Quandt, E.; Torres-Torronteras, J.; Martí, R.; Canadell, D.; Ariño, J.; Sharma, S.; Jiménez, J.; et al. Polyphosphate is involved in cell cycle progression and genomic stability in Saccharomyces cerevisiae. Mol. Microbiol. 2016, 101, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Andreeva, N.; Kulakovskaya, T.; Zvonarev, A.; Penin, A.; Eliseeva, I.; Teterina, A.; Lando, A.; Kulakovskiy, I.V. Transcriptome profile of yeast reveals the essential role of PMA2 and uncharacterized gene YBR056W-A (MNC1) in adaptation to toxic manganese concentration. Metallomics 2017, 9, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Trilisenko, L.; Zvonarev, A.; Valiakhmetov, A.; Penin, A.A.; Eliseeva, I.A.; Ostroumov, V.; Kulakovskiy, I.V.; Kulakovskaya, T. The Reduced Level of Inorganic Polyphosphate Mobilizes Antioxidant and Manganese-Resistance Systems in Saccharomyces cerevisiae. Cells 2019, 8, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hovnanyan, K.; Marutyan, S.; Marutyan, S.; Hovnanyan, M.; Navasardyan, L.; Trchounian, A. Ultrastructural investigation of acidocalcisomes and ATPase activity in yeast Candida guilliermondii NP-4 as ‘complementary’ stress-targets. Lett. Appl. Microbiol. 2020, 71, 413–419. [Google Scholar] [CrossRef]
- Ramos, C.L.; Gomes, F.M.; Girard-Dias, W.; Almeida, F.P.; Albuquerque, P.; Kretschmer, M.; Kronstad, J.W.; Frases, S.; De Souza, W.; Rodrigues, M.L.; et al. Phosphorus-rich structures and capsular architecture in Cryptococcus neoformans. Futur. Microbiol. 2017, 12, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Hothorn, M.; Neumann, H.; Lenherr, E.D.; Wehner, M.; Rybin, V.; Hassa, P.O.; Uttenweiler, A.; Reinhardt, M.; Schmidt, A.; Seiler, J.; et al. Catalytic Core of a Membrane-Associated Eukaryotic Polyphosphate Polymerase. Science 2009, 324, 513–516. [Google Scholar] [CrossRef]
- Müller, O.; Bayer, M.J.; Peters, C.; Andersen, J.S.; Mann, M.; Mayer, A. The Vtc proteins in vacuole fusion: Coupling NSF activity to V0trans-complex formation. EMBO J. 2002, 21, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Müller, O.; Neumann, H.; Bayer, M.J.; Mayer, A. Role of the Vtc proteins in V-ATPase stability and membrane trafficking. J. Cell Sci. 2003, 116, 1107–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desfougères, Y.; Gerasimaitė, R.; Jessen, H.J.; Mayer, A. Vtc5, a Novel Subunit of the Vacuolar Transporter Chaperone Complex, Regulates Polyphosphate Synthesis and Phosphate Homeostasis in Yeast. J. Biol. Chem. 2016, 291, 22262–22275. [Google Scholar] [CrossRef] [Green Version]
- Gomes-Vieira, A.L.; Wideman, J.G.; Paes-Vieira, L.; Gomes, S.L.; Richards, T.A.; Meyer-Fernandes, J.R. Evolutionary conservation of a core fungal phosphate homeostasis pathway coupled to development in Blastocladiella emersonii. Fungal Genet. Biol. 2018, 115, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, N.; DeRisi, J.; Brown, P.O. New Components of a System for Phosphate Accumulation and Polyphosphate Metabolism in Saccharomyces cerevisiae Revealed by Genomic Expression Analysis. Mol. Biol. Cell 2000, 11, 4309–4321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerasimaitė, R.; Sharma, S.; Desfougères, Y.; Schmidt, A.; Mayer, A. Coupled synthesis and translocation restrains polyphosphate to acidocalcisome-like vacuoles and prevents its toxicity. J. Cell Sci. 2014, 127, 5093–5104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerasimaitė, R.; Mayer, A. Enzymes of yeast polyphosphate metabolism: Structure, enzymology and biological roles. Biochem. Soc. Trans. 2016, 44, 234–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wild, R.; Gerasimaite, R.; Jung, J.-Y.; Truffault, V.; Pavlovic, I.; Schmidt, A.; Saiardi, A.; Jessen, H.J.; Poirier, Y.; Hothorn, M.; et al. Control of eukaryotic phosphate homeostasis by inositol polyphosphate sensor domains. Science 2016, 352, 986–990. [Google Scholar] [CrossRef] [Green Version]
- Gerasimaite, R.; Pavlovic, I.; Capolicchio, S.; Hofer, A.; Schmidt, A.; Jessen, H.J.; Mayer, A. Inositol Pyrophosphate Specificity of the SPX-Dependent Polyphosphate Polymerase VTC. ACS Chem. Biol. 2017, 12, 648–653. [Google Scholar] [CrossRef]
- Mouillon, J.-M.; Persson, B.L. New aspects on phosphate sensing and signalling in Saccharomyces cerevisiae. FEMS Yeast Res. 2006, 6, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Eskes, E.; Deprez, M.-A.; Wilms, T.; Winderickx, J. pH homeostasis in yeast; the phosphate perspective. Curr. Genet. 2018, 64, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Potapenko, E.; Cordeiro, C.D.; Huang, G.; Storey, M.; Wittwer, C.; Dutta, A.K.; Jessen, H.J.; Starai, V.J.; Docampo, R. 5-Diphosphoinositol pentakisphosphate (5-IP7) regulates phosphate release from acidocalcisomes and yeast vacuoles. J. Biol. Chem. 2018, 293, 19101–19112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbagh, Y. Phosphate as a sensor and signaling molecule. Clin. Nephrol. 2013, 79, 57–65. [Google Scholar] [CrossRef]
- Tomar, P.; Sinha, H. Conservation of PHO pathway in ascomycetes and the role of Pho84. J. Biosci. 2014, 39, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Rajagopal, A.; Xu, Y.-F.; Rabinowitz, J.D.; O’Shea, E.K. A systematic genetic screen for genes involved in sensing inorganic phosphate availability in Saccharomyces cerevisiae. PLoS ONE 2017, 12, e0176085. [Google Scholar] [CrossRef]
- Auesukaree, C.; Homma, T.; Tochio, H.; Shirakawa, M.; Kaneko, Y.; Harashima, S. Intracellular Phosphate Serves as a Signal for the Regulation of the PHO Pathway in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 17289–17294. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, C.; Saiardi, A. Eukaryotic Phosphate Homeostasis: The Inositol Pyrophosphate Perspective. Trends. Biochem. Sci. 2017, 42, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Pinson, B.; Merle, M.; Franconi, J.-M.; Daignan-Fornier, B. Low Affinity Orthophosphate Carriers Regulate PHO Gene Expression Independently of Internal Orthophosphate Concentration in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 35273–35280. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.R.; O’Shea, E.K. An intracellular phosphate buffer filters transient fluctuations in extracellular phosphate levels. Proc. Natl. Acad. Sci. USA 2005, 102, 9565–9570. [Google Scholar] [CrossRef] [Green Version]
- Boyce, K.J.; Kretschmer, M.; Kronstad, J.W. The vtc4 Gene Influences Polyphosphate Storage, Morphogenesis, and Virulence in the Maize Pathogen Ustilago maydis. Eukaryot. Cell 2006, 5, 1399–1409. [Google Scholar] [CrossRef] [Green Version]
- Vagabov, V.M.; Trilisenko, L.V.; Kulaev, I.S. Dependence of inorganic polyphosphate chain length on the orthophosphate content in the culture medium of the yeast Saccharomyces cerevisiae. Biochemistry (Moscow) 2000, 65, 349–354. [Google Scholar] [PubMed]
- Vagabov, V.M.; Trilisenko, L.V.; Kulakovskaya, T.; Kulaev, I.S. Effect of a carbon source on polyphosphate accumulation in Saccharomyces cerevisiae. FEMS Yeast Res. 2008, 8, 877–882. [Google Scholar] [CrossRef] [Green Version]
- Lichko, L.; Kulakovskaya, T.; Pestov, N.; Kulaev, I. Inorganic Polyphosphates and Exopolyphosphatases in Cell Compartments of the Yeast Saccharomyces cerevisiae under Inactivation of PPX1 and PPN1 Genes. Biosci. Rep. 2006, 26, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Kulaev, I.S.; Vagabov, V.M.; Kulakovskaya, T.V. The Biochemistry of Inorganic Polyphosphates; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2004. [Google Scholar] [CrossRef]
- Christ, J.J.; Blank, L.M. Saccharomyces cerevisiae containing 28% polyphosphate and production of a polyphosphate-rich yeast extract thereof. FEMS Yeast Res. 2019, 19, 011. [Google Scholar] [CrossRef]
- Azevedo, C.; Desfougères, Y.; Jiramongkol, Y.; Partington, H.; Trakansuebkul, S.; Singh, J.; Steck, N.; Jessen, H.J.; Saiardi, A. Development of a yeast model to study the contribution of vacuolar polyphosphate metabolism to lysine polyphosphorylation. J. Biol. Chem. 2020, 295, 1439–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethuraman, A.; Rao, N.N.; Kornberg, A. The endopolyphosphatase gene: Essential in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2001, 98, 8542–8547. [Google Scholar] [CrossRef] [Green Version]
- Eldarov, M.A.; Baranov, M.V.; Dumina, M.V.; Shgun, A.A.; Andreeva, N.A.; Trilisenko, L.V.; Kulakovskaya, T.V.; Ryasanova, L.P.; Kulaev, I.S. Polyphosphates and exopolyphosphatase activities in the yeast Saccharomyces cerevisiae under overexpression of homologous and heterologous PPN1 genes. Biochemistry (Moscow) 2013, 78, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Muller-Felter, S.; Ebel, J.P. Separation des Acides Riboiuicleiques el des Polyphosphates Inorganqucs. II. Misc an Point d’une Technqe de Separation par Adsorbtion Differentielle sur Charbon. Bull. Soc. Chim. Biol. 1962, 44, 1175–1184. [Google Scholar]
- Kulakovskaya, T.; A Andreeva, N.; Karpov, A.V.; A Sidorov, I.; Kulaev, I.S. Hydrolysis of tripolyphosphate by purified exopolyphosphatase from Saccharomyces cerevisiae cytosol: Kinetic model. Biochemistry (Moscow) 1999, 64, 990–993. [Google Scholar] [PubMed]
- Andreeva, N.; Ledova, L.; Ryazanova, L.; Tomashevsky, A.; Kulakovskaya, T.; Eldarov, M. Ppn2 endopolyphosphatase overexpressed in Saccharomyces cerevisiae: Comparison with Ppn1, Ppx1, and Ddp1 polyphosphatases. Biochimie 2019, 163, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Garinot-Schneider, C.; Pommer, A.J.; Moore, G.R.; Kleanthous, C.; James, R. Identification of Putative Active-site Residues in the DNase Domain of Colicin E9 by Random Mutagenesis. J. Mol. Biol. 1996, 260, 731–742. [Google Scholar] [CrossRef]
- Kumble, K.D.; Kornberg, A. Inorganic Polyphosphate in Mammalian Cells and Tissues. J. Biol. Chem. 1995, 270, 5818–5822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, G.M. The Nucleotide Sequence of Saccharomyces cerevisiae 5.8 S ribosomal Ribonucleic Acid. J. Biol. Chem. 1973, 11, 3860–3875. [Google Scholar] [CrossRef]
- Collart, M.A.; Oliviero, S. Preparation of Yeast RNA. Curr. Protoc. Mol. Biol. 2001, 23, 13.12.1–13.12.5. [Google Scholar] [CrossRef] [PubMed]
- Nadai, C.; Campanaro, S.; Giacomini, A.; Corich, V. Selection and validation of reference genes for quantitative real-time PCR studies during Saccharomyces cerevisiae alcoholic fermentation in the presence of sulfite. Int. J. Food Microbiol. 2015, 215, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lichko, L.; Kulakovskaya, T. Polyphosphatase PPX1 of Saccharomyces cerevisiae as a Tool for Polyphosphate Assay. Adv. Enzym. Res. 2015, 3, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Andreeva, N.A.; Okorokov, L.A. Purification and characterization of highly active and stable polyphosphatase from Saccharomyces cerevisiae cell envelope. Yeast 1993, 9, 127–139. [Google Scholar] [CrossRef]
- Parzych, K.R.; Klionsky, D.J. Vacuolar hydrolysis and efflux: Current knowledge and unanswered questions. Autophagy 2019, 15, 212–227. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, N.; McClanahan, T.K.; Simon, J.R.; Treger, J.M.; McEntee, K. Structure and Functional Analysis of the Multistress Response GeneDDR2 from Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 1996, 229, 540–547. [Google Scholar] [CrossRef]
- Jensen, L.T.; Ajua-Alemanji, M.; Culotta, V.C. The Saccharomyces cerevisiae High Affinity Phosphate Transporter Encoded by PHO84 Also Functions in Manganese Homeostasis. J. Biol. Chem. 2003, 278, 42036–42040. [Google Scholar] [CrossRef] [Green Version]
- Reddi, A.R.; Jensen, L.T.; Culotta, V.C. Manganese Homeostasis in Saccharomyces cerevisiae. Chem. Rev. 2009, 109, 4722–4732. [Google Scholar] [CrossRef] [Green Version]
- Breus, N.A.; Ryazanova, L.P.; Dmitriev, V.V.; Kulakovskaya, T.; Kulaev, I.S. Accumulation of phosphate and polyphosphate by Cryptococcus humicola and Saccharomyces cerevisiae in the absence of nitrogen. FEMS Yeast Res. 2012, 12, 617–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulaev, I.S.; Bobyk, M.A. Detection in Neurospora crassa of a new enzyme-1,3-diphosphoglycerate: Polyphosphate phosphotransferase. Biokhimiia 1971, 36, 426–429. [Google Scholar]
- Shabalin, Y.A.; Kulaev, I.S. Solubilization and Properties of Yeast Dolichyl pyrophosphate:polyphosphate phosphotransferase. Biokhimia (Moscow) 1989, 54, 68–75. [Google Scholar]
- Reusch, R.N. Poly- -hydroxybutyrate/Calcium Polyphosphate Complexes in Eukaryotic Membranes. Exp. Biol. Med. 1989, 191, 377–381. [Google Scholar] [CrossRef]
- Kornberg, A. Inorganic polyphosphate: Toward making a forgotten polymer unforgettable. J. Bacteriol. 1995, 177, 491–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, M.J.; Jakob, U. Oxidative stress protection by polyphosphate—new roles for an old player. Curr. Opin. Microbiol. 2015, 24, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Lonetti, A.; Szijgyarto, Z.; Bosch, D.; Loss, O.; Azevedo, C.; Saiardi, A. Identification of an Evolutionarily Conserved Family of Inorganic Polyphosphate Endopolyphosphatases. J. Biol. Chem. 2011, 286, 31966–31974. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomashevsky, A.; Kulakovskaya, E.; Trilisenko, L.; Kulakovskiy, I.V.; Kulakovskaya, T.; Fedorov, A.; Eldarov, M. VTC4 Polyphosphate Polymerase Knockout Increases Stress Resistance of Saccharomyces cerevisiae Cells. Biology 2021, 10, 487. https://doi.org/10.3390/biology10060487
Tomashevsky A, Kulakovskaya E, Trilisenko L, Kulakovskiy IV, Kulakovskaya T, Fedorov A, Eldarov M. VTC4 Polyphosphate Polymerase Knockout Increases Stress Resistance of Saccharomyces cerevisiae Cells. Biology. 2021; 10(6):487. https://doi.org/10.3390/biology10060487
Chicago/Turabian StyleTomashevsky, Alexander, Ekaterina Kulakovskaya, Ludmila Trilisenko, Ivan V. Kulakovskiy, Tatiana Kulakovskaya, Alexey Fedorov, and Mikhail Eldarov. 2021. "VTC4 Polyphosphate Polymerase Knockout Increases Stress Resistance of Saccharomyces cerevisiae Cells" Biology 10, no. 6: 487. https://doi.org/10.3390/biology10060487