Blowup of Accidental Images as a Passageway to Discovery: Insights into the Interaction between Hydroxyapatite Nanoparticles and Human Mesenchymal Stem Cells


Advanced Materials and Nanobiotechnology Laboratory, TardigradeNano, Irvine, CA 92604, USA
Appl. Sci. 2020, 10(22), 8204; https://doi.org/10.3390/app10228204
Submission received: 7 October 2020
/
Revised: 5 November 2020
/
Accepted: 16 November 2020
/
Published: 19 November 2020
(This article belongs to the Special Issue Applications of Nanoparticles in Nanomedicine: The Rising Path of Bioceramics in the Nano-Era)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Despite decades of research into the interaction between cells and nanoparticles, there is a lack of consensus regarding how specific physicochemical characteristics of the nanoparticles, including chemical composition, crystallinity, size, morphology, charge, and aspect ratio, among others, govern their internalization and intracellular fate. Methodological novelties offer new perspectives on the same old problematics, and often translate into an improved understanding of the given topic. Inspired by an analogy with the theme of the movie, Lisbon Story, a conceptually unconventional method for gaining insight into the interaction between nanoparticles and cells is proposed here. It involves the random, “Take 1” capture of an atomic force micrograph showing the interaction of human mesenchymal stem cells and clusters of spherical hydroxyapatite nanoparticles with a broad distribution of sizes and shapes, the blowup of its segments, and their detailed qualitative inspection. This method led to the derivation of three illustrative hypotheses, some of which were refuted and some corroborated. Specifically, the presupposition that there is an inverse relationship between the cellular uptake efficiency and the size of nanoparticle clusters was confirmed, both empirically and through a literature meta-analysis, but the idea that the geometry of these clusters affects the uptake was refuted. The definite presence of morphological determinants of the cellular uptake at the level of elementary particles, not clusters thereof, however, was confirmed in an alternative experiment. Likewise, immunofluorescent studies demonstrated that relatively large and irregularly shaped nanoparticle clusters do get internalized and localized to the perinuclear area, where they engage in an intimate interaction with the cell nucleus. The proposed enhancement of the binding between cells and biomaterials by increasing the surface ruffling consequential to the nanoparticle uptake - in analogy with the enhanced cell adhesion achieved by introducing topographic irregularities to smooth biomaterial surfaces - was also confirmed by showing that the uptake improves the stem cell adhesion. The uptake also augmented the stem cell viability and the proliferative capacity of cells reseeded with this internal nanoparticle cargo on a fresh surface, albeit with moderate levels of statistical significance and the caveat of its presumed dependence on the cell type, the nanoparticle chemistry and dose, and the overall stage in the transition of the multipotent cells toward an osteoprogenitor lineage.
Keywords:
AFM; film; hydroxyapatite; immunofluorescence; mesenchymal stem cells; nanoparticles; topography; uptake1. Introduction
Despite decades of research into the interaction between cells and nanoparticles, the literature is pervaded with insufficiently comprehensive, methodologically limited reports that frequently contradict each other’s findings with regard to the effect of specific physicochemical characteristics of the nanoparticles, such as chemical composition, crystallinity, size, morphology, charge, and aspect ratio, among others, on their internalization and intracellular fate. For example, for each report demonstrating that eukaryotic cells have a greater affinity for smaller nanoparticles [1,2,3], two reports can be found that demonstrate the opposite, namely a direct proportionality between the cell viability and the particle size [4,5,6,7,8,9]. Likewise, the number of literature reports arguing in favor of the greater affinity of cells for negatively charged nanoparticles [10,11,12] is counterbalanced by the reports demonstrating the greater affinity for positively charged nanoparticles [13,14], as well as reports showing nil difference between the two [15]. Even when comparisons are limited to a single cell type, such as mesenchymal stem cells (MSCs), which lie at the focus of this study, there are still considerable incoherencies pervading the literature. For example, while some reports provide evidence in favor of the inverse dependence of the degree of differentiation of MSCs on the particle size [16,17], other reports demonstrate the direct dependence [18,19], with the rest of the studies providing mixed findings, such as the greatest effect of particles in the medium size ranges [20,21], or the different effect of differently sized particles on markers dominant in early, middle, and later stages of the differentiation process [22,23].
This lack of consensus with respect to the causative grounds underlying the interaction of nanoparticles with cells indicates the complexity, and the existence of a multitude of physicochemical factors, that synergistically determine the interaction with cells and other biological entities. It also highlights the need for the assessment of this fundamental subject from new angles, using innovative experimental approaches. Methodological novelties offer new perspectives on the same old problematics, and often translate into enriched insight on the given topic. Inspired by an analogy with the theme of a movie about movies, titled Lisbon Story, one such conceptually unconventional method for gaining insight into the interaction between nanoparticles and cells is proposed here.
Wim Wenders’ Lisbon Story, a movie that this author watched at the Belgrade FEST in 1995 and that exists at the time of writing this piece only in three analog 35 mm prints, is to a large extent a story about the writer’s block, about the lack of inspiration for creative work befalling a burned-out auteur. In the sciences, such lack of inspiration for engaging in conceptualization and testing of groundbreaking ideas often drives scientists to the mainstream of conventional, paradigmatic research. In Lisbon Story, the film artist sought by the protagonist deals with this writer’s block by strapping a camera to his back and taking random images of Lisbon, with the idea that each of these accidental images would bear a greater significance for our lives than the best planned schemes. As in Michelangelo Antonioni’s Blowup, as it were, magnification of the fine details in these images, and meticulous subjection thereof to scrutiny can open whole new universes before the explorer and his community; a point that originates from the domain of arts, but even more strongly resonates with the core of the scientific method. In this conceptually experimental study, an analogous method involving a random, “Take 1” capture of an image showing the interaction of nanoparticles with cells was set to take place, and its detailed inspection was performed to see how far, and in what direction, it will take the investigator.
Numerous points can be invoked as justifications of this unorthodox method. For one, we live, globally, in times of unprecedented research productivity, where the amount of data ending in depository files destined never to be excavated, revisited, and included in meaningful analyses grows with each new day. In contrast, the conditions of war-stricken poverty, in which this author grew up, taught him how to efficiently use even the finest amount of data. Since under such adverse conditions every piece of data usually takes plenty of hard work and time to obtain, the value of data proportionally increases in the eyes of the investigator. The current obsession with gathering large sets of data, not necessarily for statistical studies, can be further said not only to be wasteful, but also intrinsically unscientific, in a sense that it disobeys the detailed, insightful focus on small details that is an integral element of an imaginative and elaborate scientific exploration of reality.
Therefore, like a snapshot photographer who departs to the other side of the world with only a dozen or so slots for images in her camera, preparing extensively for each analog shot, then using no processing techniques during the production [24], here I experiment with taking a random image on a high-resolution atomic force microscope (AFM), and test whether it can be a gateway to interesting findings. The last time one such random image was taken, it was also on an AFM; although the image was of a different sample, it was titled, symbolically, Lisbon Story [25]. Therefore, it has made sense that this continuation of this story, which tests if gateways to discovery really abide in each segment of reality, starts with the use of the same experimental technique. If proven correct, this would have valuable instructions for modern science. One is that the customary insistence on an abundance of data is not a necessary prerequisite for engaging in good science. This would revert us to the premises of poor science, of science for and of the poor, that this author has advocated over the years [26,27,28]. As per this ideology, poverty can be an instigator of creativity, whereas the access to state-of-the-art empirical tools puts this creativity to sleep by rendering us committed to the paradigm. While the lack of resources and funds promotes conceptually novel ideas and necessitates the use of intellectual inventiveness to compensate for the lack of access to tools that can produce fancy and flashy results, it also has no obligation to pay to the reigning academic system, where conformity to the paradigm and mainstream methodologies is needed lest the researcher be excluded as excessively strange from the circle of their peers who, as ever, have a final say in the decisions to support or emasculate one’s research.
Therefore, the presentation of results here starts with a single image of human mesenchymal stem cells (hMSCs) challenged with hydroxyapatite (HAp) nanoparticles, randomly taken on an AFM. This AFM image was then used as a sole source of inquiry into the specifics and generalities of the interaction between cells and nanoparticles. Although MSCs were chosen for this study because of their large potential in regenerative therapies, and thus for the overall need for understanding their interaction with nanoparticles better, the results of this approach may prove relevant for comprehending the general aspects of the interaction between cells and nanoparticles of many different kinds. The discussion that follows reveals how far the inquiry stemming from this unconventional method has taken this researcher in the direction of pertinent findings. In that sense, this study covers empirical grounds that have a large significance for the field of biomedicine, while at the same time it proposes a conceptual novelty, thus challenging the common experimental design practices in use in natural sciences. Such common methods are often dogmatically accepted by the scientific society, and their challenging is essential, lest science stiffen into a state of methodological bigotry that is diametrically opposite to its flexible and open-minded nature.
Another plane on which this study challenges mainstream scientific practices is by providing a novel and more veritable account of the thought process leading from the hypothesis to the experimental design, to the research outcome, and, ideally, a discovery. Whereas a conventionally structured scientific paper largely misrepresents the natural thread of thought along which scientific ideas originate, papers could be written in countless different styles and forms, notwithstanding that this is so easily forgotten. This particular study is written up with the goal of providing a better alignment with this natural progression of scientific thought, where “hypotheses appear along uncharted by-ways of thought; they are imaginative and inspirational in character; they are adventures of the mind” [29]. This metacognitive awareness underlying this study justifies the use of Lisbon Story, itself a cinematic metastudy in some respects, as its key reference point. In the end, the benefits of such innovative styles of technical writing are too many to fit in this study, the heart of which, including the whole discussion revolving around it, is purely scientific, albeit touched tinily by the gentle hand of poetry.
2. Materials and Methods
2.1. Nanoparticle Synthesis
To synthesize HAp nanoparticles, 400 mL of a 0.06 M aqueous NH4H2PO4 (Fisher Scientific, Pittsburgh, PA, USA) solution containing 25 mL 28% NH4OH (Sigma-Aldrich, St. Louis, MO, USA) was added dropwise to a 400 mL 0.1 M aqueous solution of Ca(NO3)2 (Fisher Scientific, Pittsburgh, PA, USA), which contained 50 mL of 28% NH4OH. The beaker was kept heated on a plate at 50 °C and stirred vigorously at 400 rpm. Once the addition of NH4H2PO4 had been completed, the suspension was brought to a boil, then immediately removed from the plate and left to air cool at room temperature. Stirring was suspended and the precipitate was left to age along with its supernatant under ambient conditions for 24 h. After 24 h, the precipitate was separated into 50 mL Falcon tubes and centrifuged/washed three times with deionized H2O (5 min at 5000 rpm), and left to dry in a vacuum oven (Accu Temp-19, Across International, Livingston, NJ, USA) (p = −20 mmHg) at 80 °C.
To synthesize brick-shaped and needle-shaped HAp microparticles, 100 mL solutions composed of 10 mM Ca(NO3)2, different concentrations of NH4H2PO4, and 20 mM urea were covered with aluminum foil to prevent evaporation, heated to 88 °C, and then subjected to an ultrasound field using an ultrasonicator (Q700 QSonica, Newtown, CT, USA), a regular, ½’’ tip, the amplitude equal to 20, and pulse on/off time of 2/1 s for different processing periods. The decomposition of urea at elevated temperatures proceeded according to Equation (1), yielding NH4OH as a product and an increase in the pH, which caused the precipitation of HAp, as per Equations (2) and (3).
CO(NH2)2 + H2O → CNOH + NH4+ + OH−
10Ca2+ + 6HPO42− + 8OH- + 3H2O → Ca10(PO4)6(OH)2 + 9H2O
10Ca2+ + 6H2PO4− + 11OH− → Ca10(PO4)6(OH)2 + 9H2O + 3H+
To yield brick-shaped or needle-shaped particles, the concentration of NH4H2PO4 in the reaction solution was 40 mM or 60 mM, respectively [30]. After the reaction times, ranging from 1–3 h, the samples were cooled down to room temperature and aged in air for 12 h. The precipitates were then sedimented in the centrifugal field (3500 rpm) for 5 min, separated from the supernatant and dried in a vacuum oven (p = −20 mmHg) at 60 °C.
2.2. Cell Culture
hMSCs were derived from human bone marrow and approved for use by the relevant institutional review boards at the University of Illinois in Chicago (UIC PAF No. 201405459) and the Tulane Center for Stem Cell Research and Regenerative Medicine [31]. The cells were maintained in MesenPro RS culture medium, composed of 10 mL MesenPRO RS growth supplement and 5 mL GlutaMAX-I in 500 mL of MesenPRO RS basal medium. Cell lines were grown to confluency before being plated on 12 mm circular glass coverslips or in 48-well culture plates. The cells were grown at 37 °C and 5% CO2, and were regularly monitored for possible contamination. Passages 3–5 were used for all the experiments.
2.3. AFM and SEM Imaging
To prepare hMSCs for AFM imaging, the cells were grown on glass coverslips in the aforementioned medium. Near the confluency, the cells were treated with 5 mg/mL HAp nanoparticles. After the incubation period of 72 h, the cells were fixed for 15 min in 4% paraformaldehyde, and transported to the measurement site. AFM images were taken on a Pacific Nano Technology (Santa Clara, CA, UAS) Nano-R Atomic Force Microscope (Target Technology Company, Irvine, CA, USA). The close contact mode was used to acquire all the images on the Nanorule+ software platform (Pacific Nano Technology, Santa Clara, CA, USA). Gold-coated tips were placed onto the cantilevers prior to the imaging. The closed-loop control system of the instrument was set to compensate for the creep of the piezoelectric translator of the scanner with position sensors attached to it. The resolutions in the x and y directions were <2 nm and the noise level in the z direction was <0.13 nm. The maximum range of the scanner was 80 µm in the x and y directions, and 8 µm in the z direction. No image processing was performed, aside from the artificial coloring by adjusting the color saturation and hue. Field emission scanning electron microscopy (FE-SEM) was performed on a Carl Zeiss (Oberkochen, Germany) ULTRA Plus microscope at the electron acceleration voltage of 3 kV. The samples were sputtered with gold prior to the SEM analysis of the nanoparticle size and morphology.
2.4. Immunofluorescent Staining
Cell cultures on coverslips were treated with 250 µg/mL of HAp nanoparticles for 24 h before being washed, fixed, permeabilized, and stained. Cells were fixed with 4% paraformaldehyde, then washed with the wash buffer (0.1 wt.% Triton-X and 0.1 wt.% bovine serum albumin in phosphate buffered saline, PBS) and stained with 0.2 mL per well of the staining solution containing 1:4000 v/v AlexaFluor 568 phalloidin as an actin-staining reagent and 1:100 v/v Lonza (Basel, Switzerland) OsteoImage as HAp-staining reagent, along with one drop of NucBlue ReadyProbes as a cell nucleus-staining reagent. Fixed cells were incubated at room temperature for 2 h before being mounted and cured with ProLong Diamond antifade mounting agent. Control cells that did not contain HAp were stained using the same protocol and reagents, but excluding OsteoImage.
Cells evaluated for their proliferation in response to nanoparticle uptake were detached from the polymeric growth surfaces after the exposure to HAp nanoparticles being embedded in them, resuspended, and reseeded on glass coverslips. After 7 days of incubation, as a measure of the proliferation degree, the cell number was counted and compared for different sample groups, including the untreated control cells and the cells treated with the nanoparticles. A portion of the cells was stained after 7 days of incubation for collagen type I and nuclei. The staining procedure began with washing the cells with PBS (pH 7.4) and fixing them for 15 min in 3.7% paraformaldehyde. The cells were then washed with PBS 3 × 5 min and then with the aforementioned blocking solution 2 × 5 min. The cells were then blocked and permeabilized in the blocking solution for 48 h, and then incubated in 500 μL/well of the primary antibody, 10 μg/mL rabbit anti-collagen-type-1 (Abcam, Cambridge, MA, USA) in the blocking solution for 1 h. The cells were then washed with PBS 3 × 10 min and incubated with 500 μL/well of the secondary antibody, 10 μg/mL AlexaFluor 555 goat anti-rabbit IgG (Invitrogen, Carlsbad, CA, USA), and 20 μg/mL 4′,6-diamidino-2-phenylindole dihydrochloride nuclear counterstain (DAPI, Invitrogen, Carlsbad, CA, USA), all in the blocking solution for 1 h and then washed with PBS 3 × 5 min. The coverslips containing the fixed and stained cells were mounted onto glass slides using hard set vectashield and nail polish. To visualize the lysosomes, the cells were transfected transiently with LAMP1-YFP (Addgene plasmid #1816) [32] using jetPRIME transfection reagent (Polyplus-transfection, Illkirch-Graffenstaden, France). Immunofluorescent images were collected on Nikon T1-S/L100 and C1si confocal optical microscopes at 60 × magnification in oil.
Circularity (φ) of the nanoparticle clusters was defined based on Cox’s formula [33,34], where A is the surface area of the 2D projection of the particle, and P is the particle perimeter, where a perfect circle is indicated by φ = 1.0, while the particle increasingly resembles an elongated polygon as φ approaches 0.0:
φ = 4πA/P2
2.5. Cell Viability Assay
MTT (3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide) solution was prepared according to the manufacturer’s instructions (Vybrant® MTT Cell Proliferation Assay Kit V-13154). Cells were plated and challenged with 1 mg/mL HAp nanoparticles, following the steps described in Section 2.3 and Section 2.4. The assay was performed according to the manufacturer’s instructions, and absorbance was measured at 540 nm using a microplate reader (FLUOstar Omega, BMG LABTECH). Normalization to the control was used to convert the absorbance values to cell viabilities.
2.6. Flow Cytometry
To measure the cellular uptake using fluorescent activated cell sorting, cells were grown in 24 well plates (Corning) and treated overnight with the nanoparticles at 5 mg/mL. Prior studies have shown that saturation in the cellular uptake of nanoparticles is usually reached by 24 h of incubation [35,36,37]. After the incubation, the cells were washed to remove the nanoparticles and trypsinized. The cellular uptake of the nanoparticles was determined by assessing the granularity of the cells, based on increased side scatter (SSC) of cells compared to the controls (FACSVerse, Becton Dickinson, Laguna Hills, CA, USA).
3. Results and Discussion
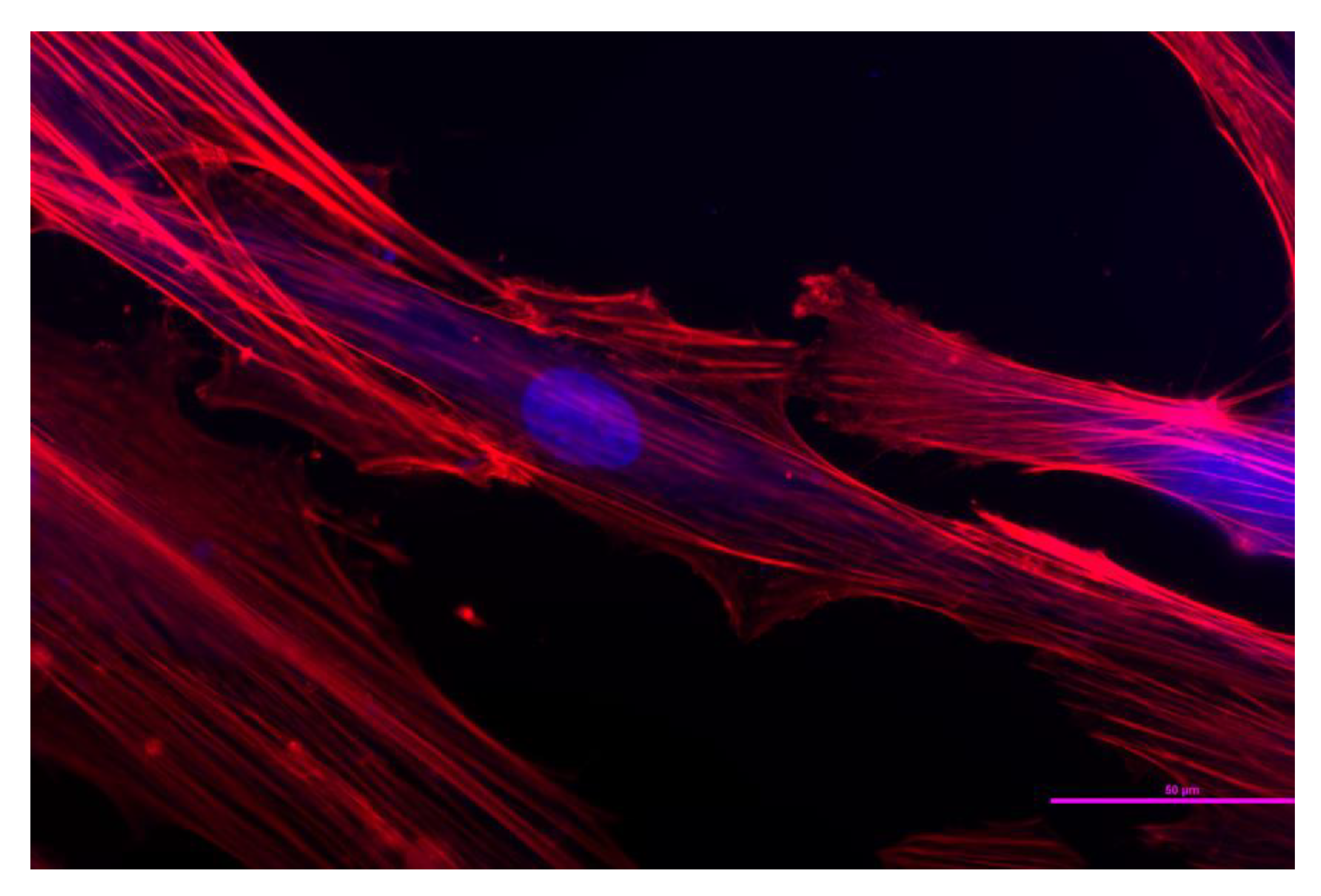
The random image was taken (Figure 1), hung on the wall, like in Antonioni’s Blowup, and gazed at for days. Then the little details began to pop up, inspiring thought-provoking observations.
The AFM image shown in Figure 1 represents a handful of fixed hMSCs challenged with HAp nanoparticles. The cells adhering onto the bottom of the cell culture well display lower aspect ratios than those of spindly fibroblasts, including the control hMSCs (Figure 2), which may be indicative of their being impelled to enter the osteoblastic differentiation path after the exposure to bone mineral particles. Osteoblasts are derived from MSCs in a process paralleled by the expression of bone morphogenetic proteins, members of the Wingless pathways, and proteins including Runt-related transcription factors 2, Distal-less homeobox 5, and Osterix [38]. In the presence of HAp and other calcium phosphate nanoparticles, hMSCs have demonstrated the overwhelming tendency to transform to osteoblasts rather than to adipocytes or chondrocytes [39,40,41]. Accompanying this transition toward an osteoprogenitor lineage, the pronounced degree of elongation of hMSCs gets reduced, and cells begin to increasingly adopt a round appearance and the squared shapes characteristic for osteoblasts [42]. The differentiation process, however, usually requires timescales longer than 72 h [43], meaning that the majority of cells must have maintained their mesenchymal phenotype by the point of their fixation, and that no endogenous mineral nodules have yet begun to form. Imaged under the AFM, the cells also display typical features of fixed cells, primarily in terms of the lack of distinct cytoskeletal fibers, the reason being the crosslinking of the membrane proteins during the fixation procedure and the resulting stiffening of the cell membrane, which, as such, becomes more pronounced than the actomyosin microfibers [44].
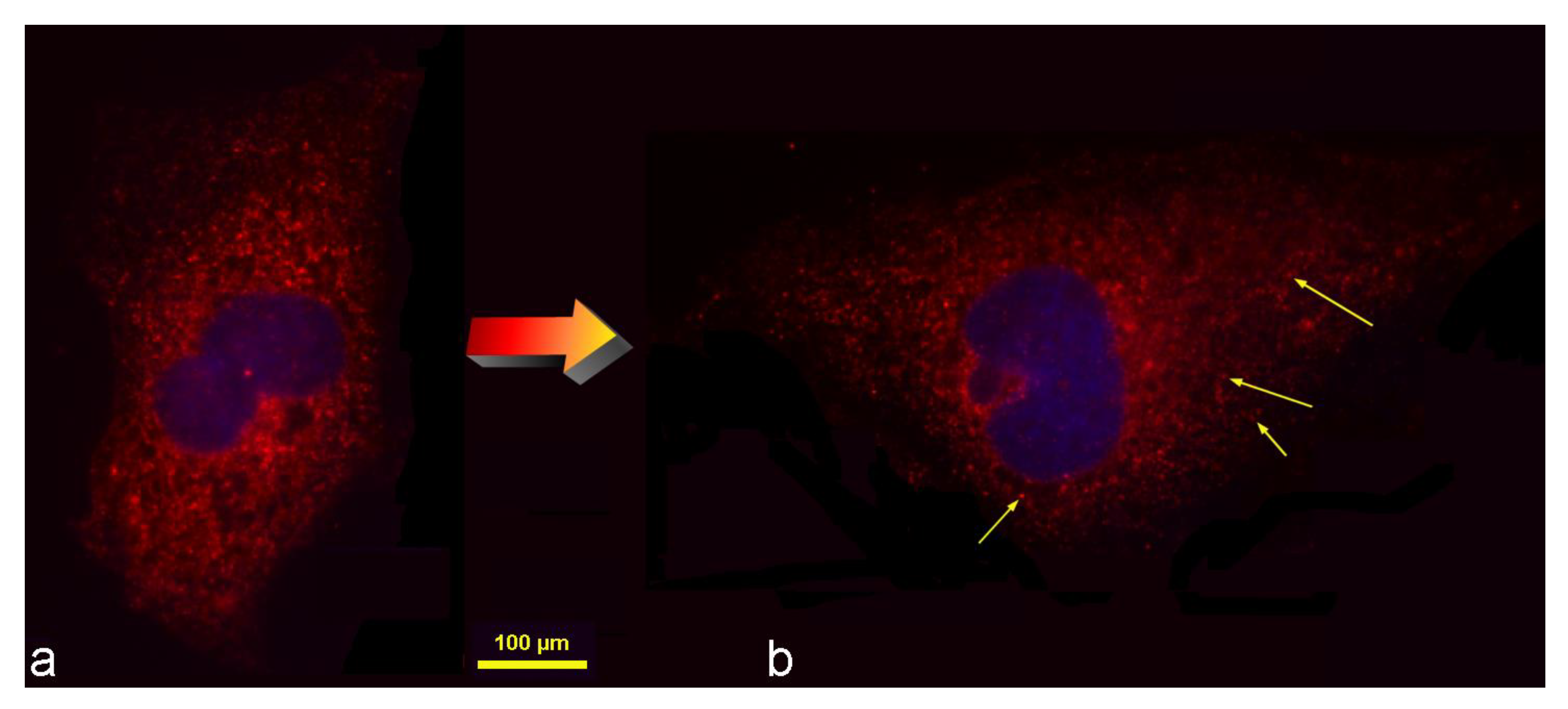
The cell sizes vary and in this image decrease along the diagonal connecting the bottom left corner with the upper right one. HAp nanoparticle agglomerates visible in the image vary in size, too. Since the primary particles in this experiment were well under 100 nm in size (Figure 3e), such elementary particular entities would be invisible at the resolution of 185 nm employed to capture both the cells and the particles, with the entire image shown in Figure 1 having dimensions of 93.40 × 93.40 µm. Therefore, all the distinguishable particulate entities in Figure 1 are agglomerates of HAp nanoparticles varying in size and shape. The most prominent and largest of them, visible near the upper central edge of the image frame, is magnified in Figure 3a. This nanoparticle cluster is ~6.3 × 4.1 µm in size and falls right around the 5 μm upper limit for the nonspecific uptake pathway, also called micropinocytosis [45]. This size limit is significantly higher than the 50–500 nm upper limits established for the two specific, receptor-mediated uptake pathways, namely clathrin-mediated endocytosis (<200 nm) and caveolin-mediated endocytosis (<500 nm) [46], as well as for the clathrin- and caveolin-independent uptake routes, such as the lipid-raft mediated and ARF6-mediated GEEC pathways. Despite looking very bright, this specific nanoparticle aggregate has only a 1.7 µm height, meaning that it is likely to be prone to macropinocytosis, at least by the sizable cells specializing in the uptake of particles and particle debris, such as macrophages or osteoclasts. Based solely on AFM imaging, however, it is impossible to precisely conclude whether this particle lies on the inside or the outside of the cell. Given the prominence of the object, its considerable size and sharp features, it is most likely that it rests on the surface of the cell, which may not be able to uptake this foreign object considering that it is only 5–10 times larger than it.
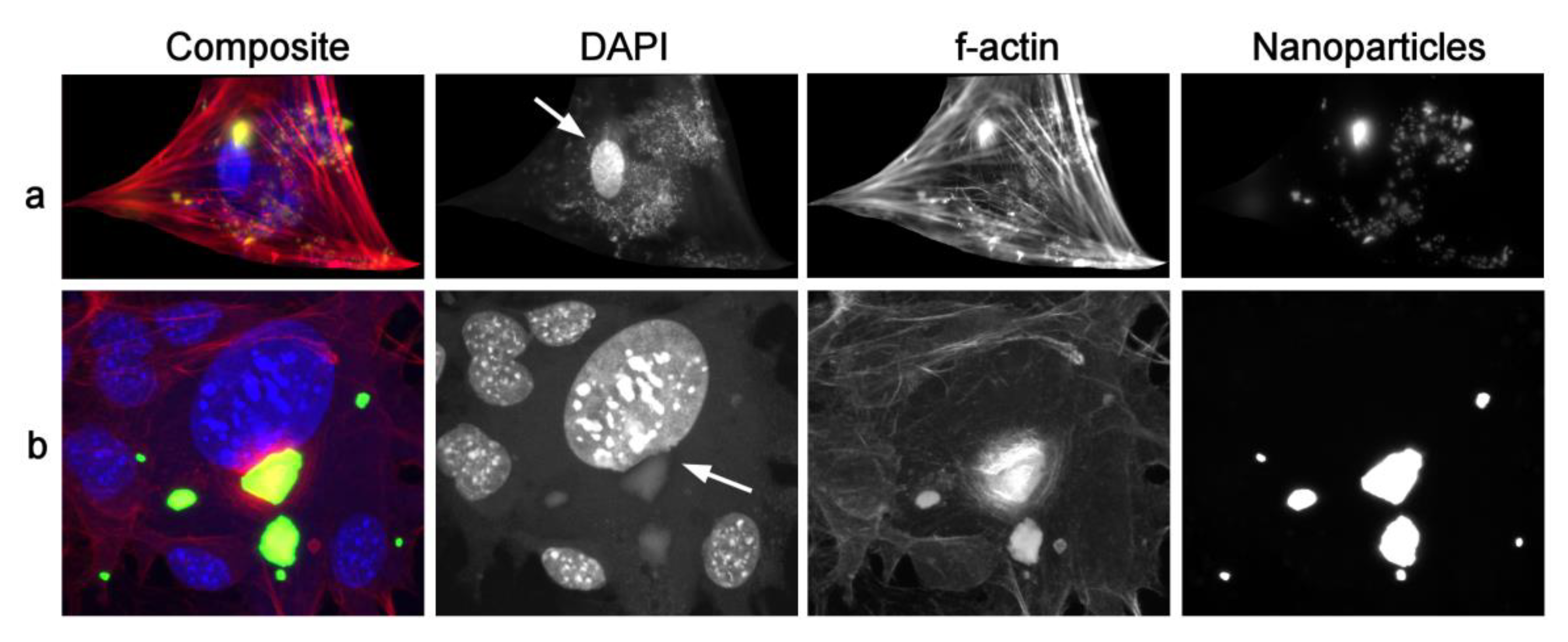
Figure 4, however, shows two interesting scenarios derived from optical fluorescent imaging. In both cases, a somewhat smaller, but equally irregularly shaped HAp nanoparticle cluster as that in Figure 3a coincides with the cell nucleus. In one case, however, it does not directly interfere with the nucleus (Figure 4a), while in the other case (Figure 4b) it obviously indents the nucleus due to its localization in the direct vicinity of it. This is to say that very large objects do get internalized by the cells, without necessarily getting molded to softer, more curved geometries during the passage through the cytoplasm towards different posts in the cell. Nanoparticle agglomerates can preserve their sharp outlines and this will be an important premise for the analysis that will be reported in Section 3.2.
Another interesting cluster of nanoparticles to pay attention to is the one magnified in Figure 3b, apparently resting tangled with either the cell membrane or the microvilli extensions at the interface between the cell and its microenvironment, and having a pronouncedly elongated shape. This particle adheres parallel to the cell membrane, in a configuration that imposes a comparatively large membrane deformation energy cost to the cell upon its uptake [47], for which reason its rotation by ~90° is expected to occur prior to the uptake [48]. In contrast to it, the particles appearing to be localized inside another cell, magnified in Figure 3c, appear rounder. The edges of these seemingly internalized particles are soft and curved, unlike the sharper edges of a set of particles tangled within the cellular filopodia and apparently resting outside the cell boundaries (Figure 3d).
Inspired by this brief discussion of features intrinsic to a randomly taken AFM image, one question emerges: may the overly large particles as well as those with overly sharp edges be less prone to cellular internalization than their smaller and more softly shaped counterparts? This question is divisible to two distinct hypotheses, which will be treated and discussed in Section 3.1 and Section 3.2, respectively: (1) the uptake is reduced as the size of the nanoparticle clusters increases, and (2) the uptake is diminished for nanoparticle clusters whose shapes deviate from the perfect spherical symmetry.
A third question also arises: what exactly constitutes the observed round topographic features deemed to be nanoparticle agglomerates? Given the working principle behind AFM imaging, where the signal is formed by the detection of changes to the topographic elevations by a nanoscopic, atomically sized probe scanning the cell surface, it is conceivable that any internalized particles would be invisible to the AFM probe. However, as seen by the comparison between the portion of the AFM image from Figure 1 magnified in Figure 3c and the optical micrograph presented in Figure 4a, the localization of the particles in the perinuclear region is specific for these nanoparticles. This may suggest that, like the bulging belly of a boa constrictor after swallowing an elephant [49], AFM can detect topographic irregularities on the cell surface that are secondary to the uptake of the nanoparticles. The significantly higher stiffness of nanoparticles, compared to that of the softer cytoplasm, supports this possibility. A prior AFM study demonstrated an increase in the surface roughness of SKOV-3 ovarian cancer cells following their uptake of poly-L-lactide-co-glycolide (PLGA) nanoparticles and the washing of any remnants thereof adhering to the membrane [50]. Furthermore, if internalization of nanoparticles produces fine corrugations on the cell surface, would the uptake then possibly facilitate the binding of the cells onto materials in the same way that the creation of topographic irregularities on smooth surfaces promotes cell adhesion and proliferation on them? [51]. Section 3.3 describes a preliminary effort to test this hypothesis, that the cellular uptake of nanoparticles could enhance cell adhesion and proliferation, notwithstanding that any confirmation of this effect would in no way be directly and unequivocally indicative of the mechanism outlined in this paragraph. Likewise, it is possible that the opposite effect compared to the one hypothesized here would be observed in case the uptake and the surface growth turn out to be mutually antagonistic.
Here, the author should be excused for the naïveté of these presuppositions emerging from the analysis of Figure 1, because even the bluntest and plainly erroneous hypotheses at this instant could still lead to valuable insights. Besides, given the conceptually innovative nature of this study, it primarily serves the purpose of demonstrating a route worth taking in cell biology, biomaterials, or any other research field as an alternative to the standard, objective-driven research plans, whose programmatic nature can be said to have largely obscured almost everything that is pure and absolute in science as we have known it.
3.1. Smaller Particle Conglomerates Are More Likely to Be Internalized by the Cells
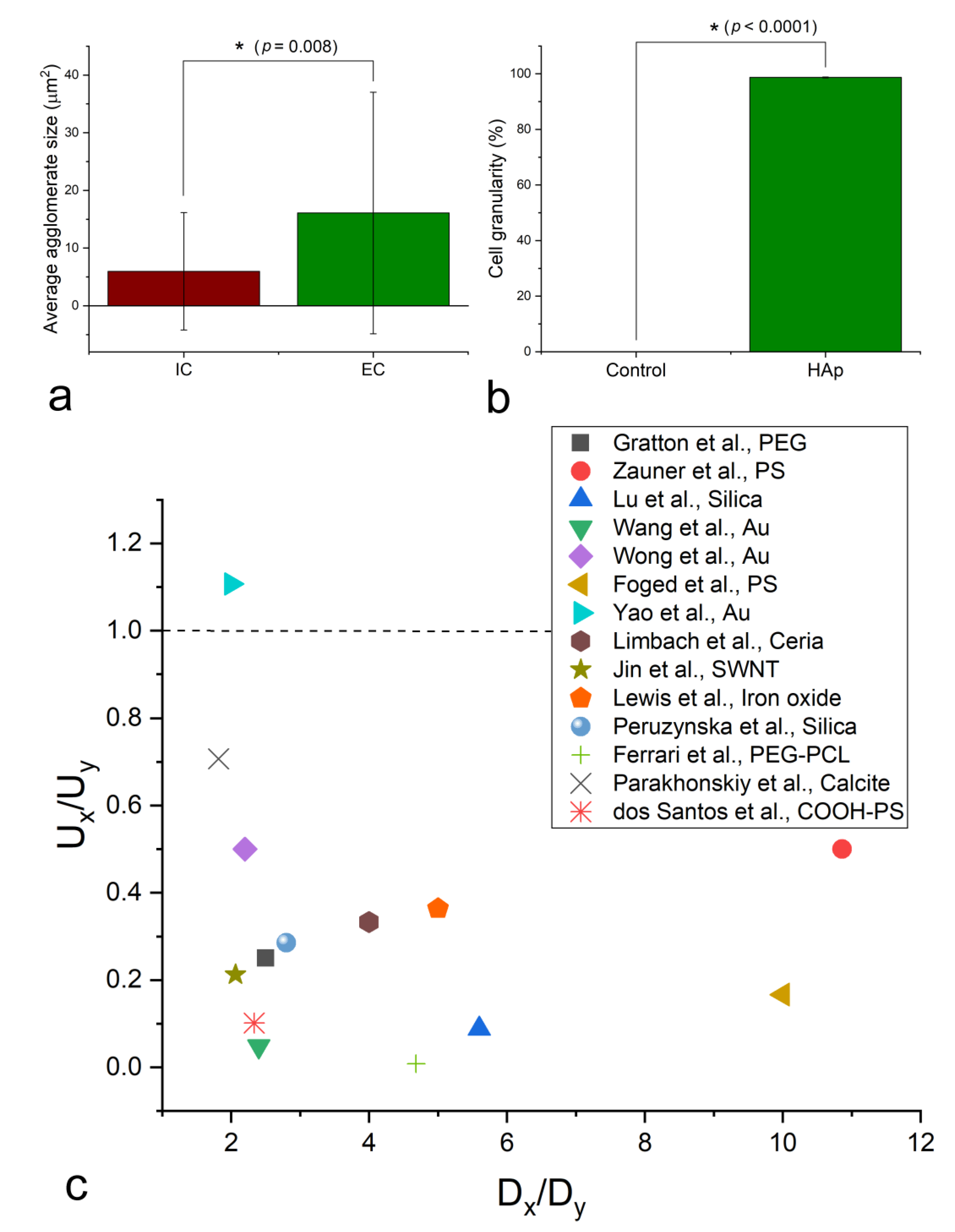
Owing to the ambiguities surrounding the exact location and geometries of nanoparticle clusters overlapping with cellular boundaries, all measurements used to test the proposed hypotheses were performed on fluorescent optical images of cells in interaction with HAp nanoparticles. The results of the testing of the relationship between the cellular uptake of the nanoparticle agglomerates and their sizes demonstrated a clear tendency for the smaller clusters to undergo a more abundant intracellular localization than the larger ones. Specifically, as seen in Figure 5a, the average size of intracellularly located nanoparticle agglomerates was significantly lower than that of the extracellularly located ones. Meanwhile, the cell granularity following the overnight treatment with HAp nanoparticles was in excess of 98% (Figure 5b), indicating that the great majority of cells did uptake large enough amounts of nanoparticles to display significantly higher granularities than the untreated cells. The literature data overwhelmingly agree with this trend, as indicated by the results of the meta-analysis presented in Figure 5c and showing that in all but one report analyzed, the smaller particles were taken up more efficiently than the larger ones. By this, in both the experimental and the literature analyses, it is meant that the smaller particles outnumber the larger ones inside the cells, not that their absolute amounts are larger, given than a single large nanoparticle cluster can outweigh thousands or millions of smaller ones, which would present a misleading metric. In addition to this, a number of studies have demonstrated diametrically opposite effects of the particle size on the uptake efficiency in different cell lines. In one of them, for example, the uptake increased with the PLGA particle size in RAW264.7 macrophages, and decreased in human epidermoid cancer KB cells [52]. Likewise, in another study, the uptake percent of rhodamine B labeled carboxymethyl chitosan nanoparticles increased with the particle size in murine macrophages and decreased in L02 and SMMC-7721 cells [53]. Other studies [54,55] have confirmed that macrophages, the comparatively large cells specializing in particle uptake, usually deviate from the trend observed, both here and in the literature, to apply to fibroblasts, osteoblasts, epithelial cells, MSCs, and multiple other cell types, for which reason they, alongside particles lower than 20 nm in size, were excluded from the literature meta-analysis reported in Figure 5c. This does not go against the definite presence of a lower critical limit for nanoparticle uptake [56], for which reason cells often wait for nanoparticles adhering onto their membranes to cluster together before they uptake them within a single endocytic vesicle [57]. Typically, thus, around the nanoparticle size of 50 nm the maximum uptake efficiency is reached, below which it begins to drop with a further reduction in the particle size [58,59].
Here it should be pointed out that the particle uptake and colloidal stability are tightly related in a sense that smaller particles that tend to be more stable in suspension tend also to come into less contact with the cells seeded at the bottom of the cell culture plates compared to the larger particles, which are more prone to sedimentation. However, since this effect favors the uptake of larger clusters of nanoparticles, which, as per these results, get uptaken less than the finer ones, it can be concluded that cells do definitely prefer to uptake the smaller particles. In any case, 2D culture is not always a good proxy for 3D in vivo scenarios, for which reason deviations from the trend observed may occur in real-life conditions.
3.2. More Irregular Particle Clusters Are Equally Likely to Be Internalized as the Less Irregular Ones
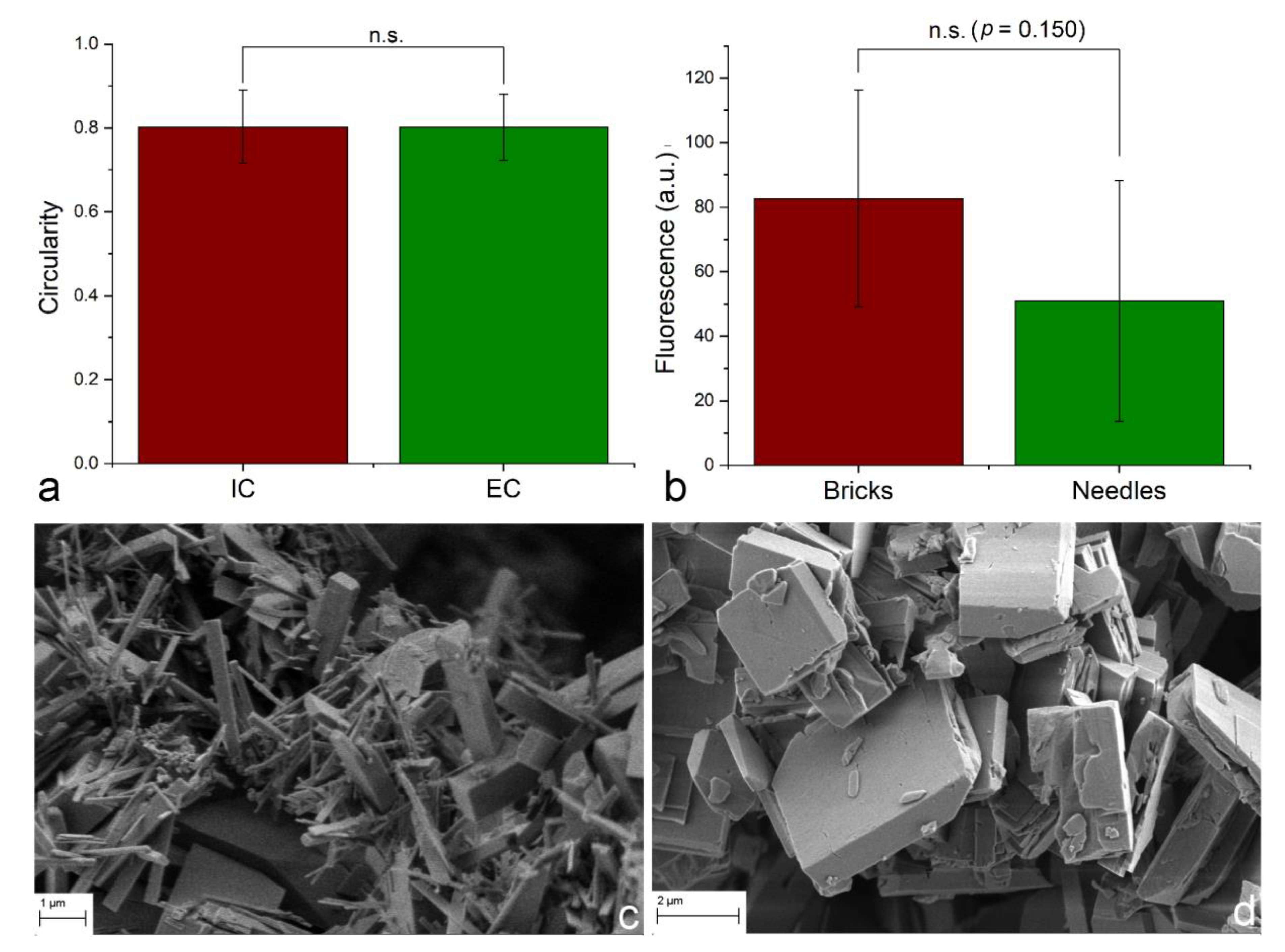
Particle shape is an essential determinant of the cellular uptake and of the overall response of the cells to the nanoparticle treatment. A number of studies have demonstrated that nanoparticles with different morphologies can be uptaken, accumulated, and/or eliminated from the cells at thoroughly different rates [74,75,76]. For example, displaying a high level of semblance with biogenic apatite platelets, HAp flakes upregulated the expression of genes and augmented the production of proteins that initiate osteogenic differentiation more efficiently than HAp needles or rods [77]. Another study contrasted these findings by showing that HAp spheres enhance the osteogenic differentiation of MSCs more than HAp needles or flakes [78]. However, a study showing that the uptake of gold nano-spheres by RAW264.7 and HeLa cells is inversely proportional to their size in the 22–66 nm range, while the uptake of gold nano-triangles is directly proportional to their size in the 46–94 nm range [79] may be particularly illustrative here, by its demonstrating that shape can outweigh size as a factor determining the uptake efficiency. Inspired by the rod-shaped nanoparticle cluster shown in Figure 3b, and its ambiguous localization halfway between the inside and the outside of a cell, an analysis was conducted based on measuring the deviation of the intracellular and extracellular HAp nanoparticle clusters in hMSC cultures from the perfect circularity, as defined by Equation (4). Against expectations, the circularity of internalized and external clusters was practically identical at 80.14 ± 0.08 for the intracellular clusters, and 80.22 ± 0.09 for the extracellular ones (Figure 6a). This indifference of the cell to the cluster geometry when it comes to the decision to internalize it or not, however, applies to a sample composed of spherical nanoparticles constituting the clusters. When the primary particles comprising separate samples differ in morphology, the situation might be different. To test this, two distinct powders were synthesized, differing in the primary particle shapes, both being in the micrometer size range, but one being highly elongated and uniaxial (Figure 6c), and the other being planar (Figure 6d). The fact that the uptake was more pronounced for particles with more isotropic shapes compared to those that looked more needle-like in morphology (Figure 6b) suggests that the cells do have a way of distinguishing between more and less preferential particle geometries. These findings may be read as a pointer toward the ability of cells to sense the fine, nanostructured topography of clusters and thereupon discard any geometric differences between them, which they do take into account when larger particles, containing no such nanoscale substructure, are brought into an interaction with them.
These results combined suggest that in a sample where nanoparticles of uniform morphologies form a broad distribution of aggregates of different shapes, the uptake process is stochastic and all shapes have an equal probability of uptake, assuming that their sizes are identical. However, when the elementary particles differ in morphology, the uptake is bound to be shape-dependent. A lateral conclusion may be that the oval shape of vesicles internalizing the particles upon the uptake is not exclusive when it comes to the internalization of particle geometries deviating from perfect circularity. In fact, as shown in Figure 7, although spherically shaped lysosomes present a great majority of the lysosomal population in both control cells and cells undergoing the nanoparticle uptake, a somewhat higher percentage of liposomes exhibits asymmetric shapes in the latter cells. This insight coincides with the earlier findings, where discoid vesicles were transformed into tubular vesicles under pH changes, such as those facilitated by the partial dissolution of HAp inside the lysosomal compartments [80]. This also bears resemblance to the earlier demonstrated adjustment of the reverse micelles to the shape of nanocrystals precipitated inside their aqueous interiors [81]. Therefore, although endosomes, lysosomes, caveosomes, macropinosomes, phagosomes, and other vacuoles are usually depicted as spherical, they appear to have the capability to partially adjust their shape to that of the solid particles transported therein. There are certain limitations, of course, as to how far a vesicle can stretch laterally, for which reason the typical trend with endocytosis is such that the internalization rate of spherical entities is higher than that of those with higher aspect ratios [82]. This agrees with the virtual absence of any evidently rod-shaped, let alone more exotically shaped lysosomes visualized in Figure 7.
3.3. Uptake of HAp Nanoparticles Can Facilitate the Cell Adhesion and Proliferation on the Surface
The observed corrugations on the cell surface visualized under AFM (Figure 1) were likely to have originated from the uptaken solid entities, which are considerably stiffer than the cytoplasm. This inspired the question of whether such topographic irregularities might cause an improved cell adhesion onto the surface in analogy with the way the introduction of similar topographic irregularities to the surface of biomaterials promotes the cell adhesion and proliferation, especially when the cells are boney in nature [83].
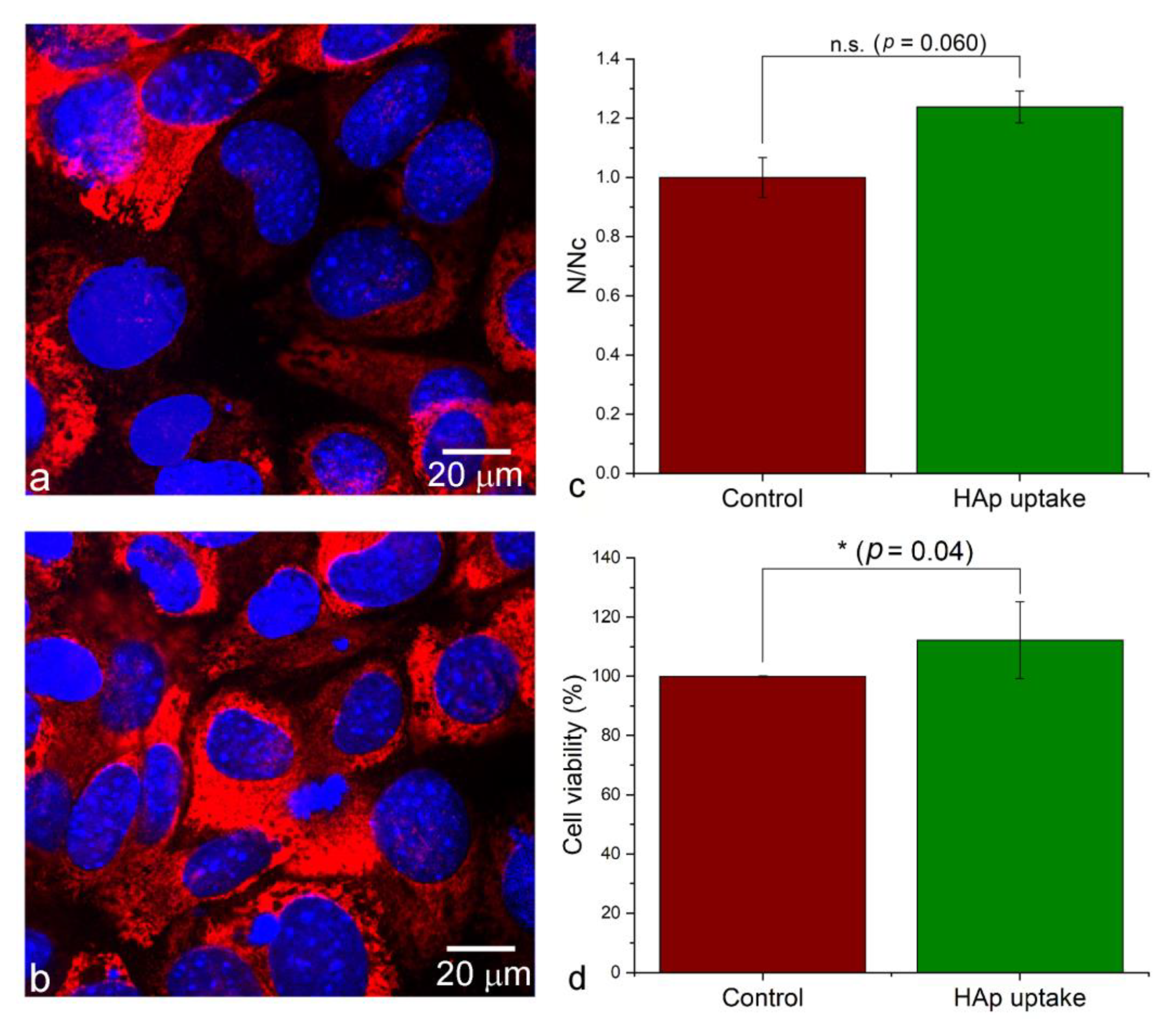
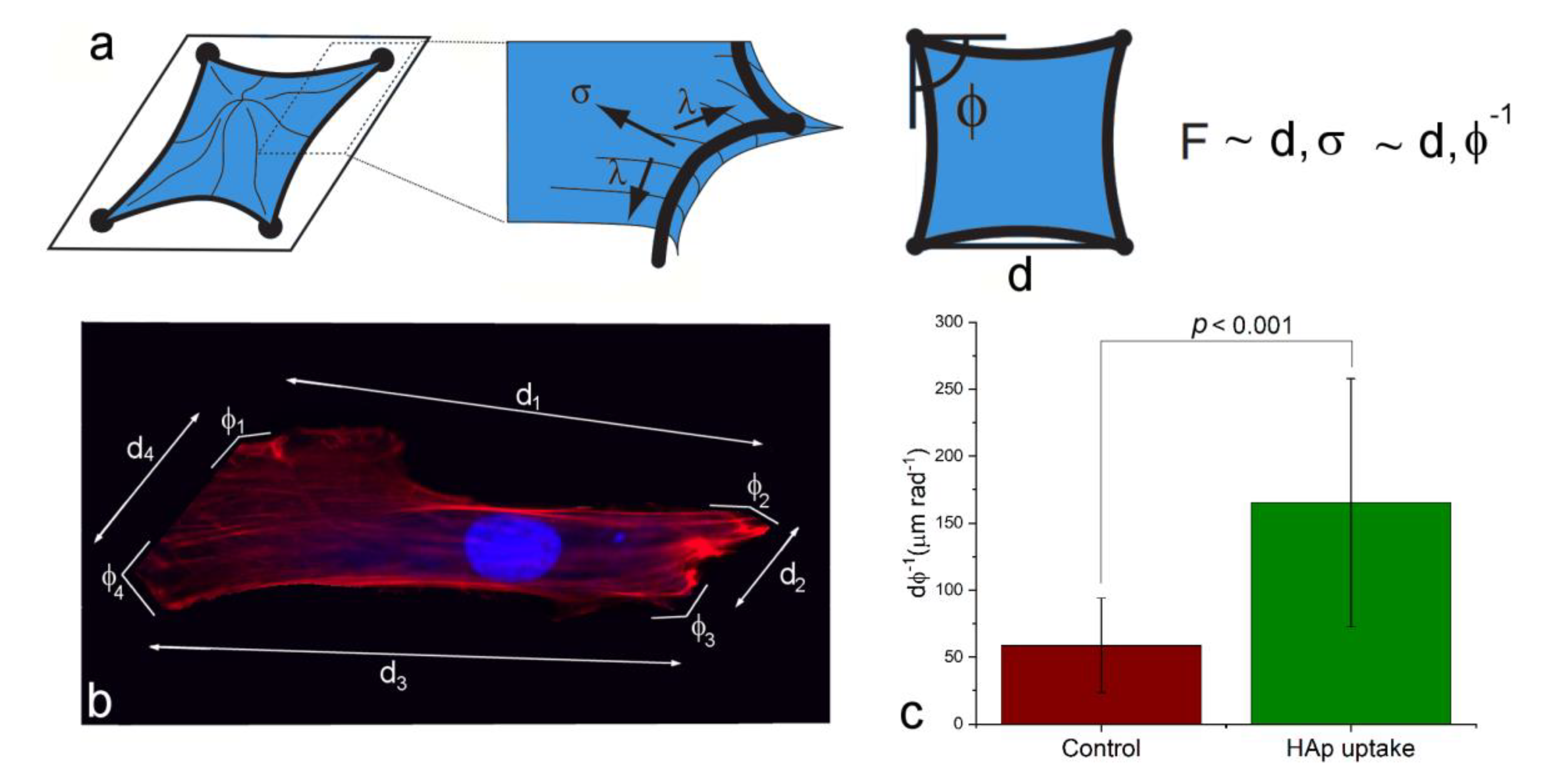
Albeit scarce, prior literature reports were tentatively not in favor of this hypothesis. For example, an earlier study showed that the osteoblast detachment force actually dropped after the uptake of HAp [84]. Moreover, the treatment of suspended hepatoma cells with HAp diminished their ability to adhere to the cell culture plates, and resulted in cell apoptosis [85]. Yet another study reiterated this diminishment of the cell adhesion onto the surface consequential to the uptake, by showing that the internalization of 40–80 nm sized HAp nanoparticles by hepatoma cells suspended the proliferation and weakened the adhesion of the cells onto the bottoms of the cell culture flasks [86]. Since the interaction between integrins and nanoparticles has been shown to favor endocytosis [87], it can be expected that integrin-mediated mechanotransduction and integrin-mediated endocytosis may compete with one another, thus rendering the uptake and the adhesion inversely proportional to one another. This scenario, however, need not apply to osteoblastic cells that MSCs can transition to. Namely, these cells are migratory, but the activation of mineralization binds them, at least temporarily, to a particular locus, where the intensified binding onto the surface is expected. This effect was first indirectly evidenced by the increased intracellular and pericellular collagen production by cells treated with HAp (Figure 8b), than by the control cells (Figure 8a). Collagen was used here instead of HAp as an immunofluorescent proxy for gauging the mineralization activity, because the endogenous and the administered HAp are indistinguishable in a stain assay such as Alizarin Red. Furthermore, both the cell density as an indicator of the proliferation rate (Figure 8c), and the viability measured as the extent of the mitochondrial metabolic activity (Figure 8d) were higher in the population of cells treated with HAp than in the control population. Here it should be noted that compared to the statistically significant increase in cell viability following the uptake of HAp, the uptake provided some, but not a fully statistically significant advantage for cell proliferation (Figure 8c-d). Finally, the estimation of the change in the cellular adhesion force with the uptake of HAp nanoparticles was made on the basis of the methodology postulated by Bischofs et al [88]. As described in Figure 9a,b, this method correlates the adhesion force with the specific parameters of the cell geometry, including the circumference and the opening angles at the cell corners as discrete sites of adhesion. The results of an analysis carried out on a sample of 16 cells for each sample group demonstrated a definitely larger adhesion force of hMSCs subjected to the uptake of HAp, compared to the control cellular population, as indicated by the almost three-times higher ratio of the average side length of the cell contours to the average opening angle (Figure 9c), which is directly proportional to the adhesion force (Figure 9a).
These findings suggest that infusing the cells with the mineral nodules can be expected to commit the cell to a particular location, at least until these intracellular nodules do not become secreted and fused with the extracellular mineral phase being built. Since HAp nanoparticles, upon internalization, alter the actin cytoskeleton architecture in the direction of increased cell adhesion favoring osteogenic differentiation, and through increased Ca2+ levels activate Ca2+-mediated signaling cascades, e.g., Ras/cAMP/Rap1/MAPK, which also set the stem cells on the osteogenic differentiation pathway [89], this scenario is quite realistic and is supported by the observed morphological semblance of HAp-exposed hMSCs and round osteoblastic cells evidenced in Figure 1. This effect, however, is expected to be greatly dependent and perhaps even antagonistic, depending on the cell type, the nanoparticle dose, and the cell seeding density. Moreover, the normally inverse relationship between the metabolic activity and the proliferation rate often predisposes the treated cells to display opposite effects relative to the control at different incubation time points. For example, the proliferation of cells seeded on HAp-containing polymeric surfaces with distinct topographic features was initially slowed because of their enhanced differentiation and augmented metabolic activity, but it later caught up with the control, and even surpassed it [90]. Osteoblasts originating from MSCs, however, may be immune to this antagonism; an earlier study, for example, demonstrated the faster proliferation of such cells than that of their undifferentiated MSC precursors [91].
HAp owes its excellent cellular uptake propensities to multiple effects, including the bioactivity of its elemental composition [92], the low net surface charge [93], the superb adsorption capacity [94], and the fine release of calcium ions [95]; the exocytotic activators, fostering the opening of the complementary endocytic pathways [96]. HAp does not have the mechanical rigidity of metallic nanoparticles or oxides such as silica, which is known to be a physical factor favoring the uptake [97], but it has a diffusive surface character thanks to its hydroxylated, clay-like interface [98,99,100], which endows the interface with the cells with a certain “stickiness” that is of benefit for the particle internalization. The topographic irregularities, consequential to the nanostructured nature of the particles, also contribute to the affinity of cells for them, resulting in the copious capture of even as large of entities as that observed leaning onto the cell nucleus in Figure 4b. This propensity for cellular uptake can be harnessed to elicit various therapeutic effects in cells. In the past, the uptake of HAp nanoparticles was used with success to increase the activity of a chemotherapy, by delivering its molecular mediator closer to its intracellular target site [101], to alter the cancer vs. healthy cell selectivity of the response of in vitro culture to magnetic nanoparticles [102], and, of course, to promote the differentiation of stem cells into the osteoblastic lineage [103]. Here, this repertoire of effects is extended by showing that the uptake of HAp can significantly improve the adhesion of hMSCs onto the biomaterial or tissue surface. This uptake also augments the cell viability and mildly improves the proliferative capacity of cells reseeded with this internal nanoparticle cargo on a fresh surface.
The findings presented here, with respect to the relationship between the cellular uptake and the cell adhesion, viability, and proliferation, however, should be treated with caution because they may not necessarily apply to different nanoparticle chemistries, cell types, or growth conditions. Still, this establishment of a positive relationship for at least one cell type and particle concentration speaks in favor of the unorthodox research approach followed here. This approach is such that it starts from an accidentally recorded piece of data and then subjects it to a detailed inspection. Despite the blurredness and the comparatively rudimentary form that the exemplary image in Figure 1 assumed, the insights that it led this investigator to, have been nothing short of copious and surprising. To allow the data to surprise the analyst and shake his convictions speaks of the grounds from which scientific inquiry should be carried out more often, especially in a heavily objective-driven research climate, where data are usually but bricks used to edify the paradigm.
4. Conclusions
Film, like all forms of art, can provide an inspiration for the conceptualization of research in the natural sciences [104]. Here, a thread of analogy was drawn between the method used by the filmmaker in Wim Wenders’ Lisbon Story and the method for research in natural sciences. Like the randomly captured photographs of a city in search of an artistic effect in this film, a single random AFM image of hMSCs interacting with HAp nanoparticles was recorded and used as a sole starting point of inquiry into the nature of this interaction.
Overall, a gaze at the randomly captured image inspired the derivation of hypotheses, some of which were refuted, and some of which were confirmed after testing. Specifically, the presupposition that there is an inverse relationship between the uptake efficiency and the size of nanoparticle clusters was confirmed, but the idea that the geometry of these clusters affects the uptake was refuted, notwithstanding that the definite presence of morphological determinants of the cellular uptake at the level of elementary particles, not clusters thereof, was confirmed in an alternative experiment. Finally, the proposed enhancement of the binding between cells and biomaterials by increasing the surface ruffling consequential to the nanoparticle uptake was tentatively confirmed, albeit being in need of more in-depth research.
In the broad frame, it is impossible to tell whether the refutation or the confirmation of these hypotheses bears a greater significance for the evolution of knowledge in this sphere. In any case, when it comes to the scientific insights that resulted from what can be derived from an accidentally taken AFM image like the one posed at the opening of this study, only the surface was grazed. Overall, the findings of this study that pertain to the field of nanomedicine, in need of significant deepening, may be considered secondary compared to the conceptual novelty of the unconventional research method from which they stemmed. To reiterate, this study, from the conceptual standpoint, has challenged the mainstream, objective-driven research methods dogmatically embraced by the scientific society, wishing to act as a remedy against the stiffening of science into states of methodological monotony that are fundamentally at odds with the open-mindedness underlying insightful scientific research. If this striving for conceptual innovation succeeds in inspiring fellow researchers to engage themselves and their study groups in similar analyses as that performed here, countless novel findings may result, and the ambitions attached to this work by the author may be considered fulfilled. For, by showing that a fine detail, invisible to the naked eye, can be more important than insatiable collections of data in this adventure of the human mind that we call science, something beautiful is being told. It is of particular relevance to a world that cranks out, craves, and swallows more and more of the expressions, impressions, and stimulations with each new day, when little is enough.
Besides, in the times of a viral pandemic, during which this work has been drafted, when most labs are closed but the wheel of knowledge must keep spinning, it is a humbling insight that sometimes a single image, perhaps resting buried in some long forgotten folders, conceals the source of innumerable interesting and innovative ideas, if it only becomes inspected patiently. A random flicker of light, a stray snap of the electron beam, or the swift scan by a nanoscale tip thus become potential gateways to whole new areas of human knowledge. Random, not so random at the end of the day, in this world that staggers with its atomic laws and its Platonist aesthetics and beauties that leave tongues tied and languages silent.
Funding
This research received no external funding.
Acknowledgments
Thanked are Victoria M. Wu for the experimental assistance and Target Technology Company in Irvine, CA for allowing the use of AFM.
Conflicts of Interest
The author declares no conflict of interest.
References
- Poormoghadam, D.; Almasi, A.; Ashrafizadeh, M.; Vishkaei, A.S.; Reyazat, S.M.; Tavakol, S. The particle size of drug nanocarriers dictates the fate of neurons; critical points in neurological therapeutics. Nanotechnology 2020, 31, 335101. [Google Scholar] [CrossRef] [PubMed]
- Perez-Garnes, M.; Gutierrez-Salmeron, M.; Morales, V.; Chocarro-Calvo, A.; Sanz, R.; Garcia-Jimenez, C.; Garcia-Munoz, R.A. Engineering hollow mesoporous silica nanoparticles to increase cytotoxicity. Mat. Sci. Eng. C 2020, 112, 110935. [Google Scholar] [CrossRef] [PubMed]
- Shanei, A.; Akbari-Zadeh, H.; Attaran, N.; Salamat, M.R.; Baradaran-Ghahfarokhi, M. Effect of targeted gold nanoparticles size on acoustic cavitation: An in vitro study on melanoma cells. Ultrasonics 2020, 102, 106061. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-D.; Wu, D.; Shen, X.; Liu, P.-X.; Yang, N.; Zhao, B.; Zhang, H.; Sun, Y.-M.; Zhang, L.-A.; Fan, F.-Y. Size-dependent in vivo toxicity of PEG-coated gold nanoparticles. Int. J. Nanomed. 2011, 6, 2071–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.J.; Liu, N.; Liu, Q.S.; Zhang, J.Q.; Zhou, Q.F.; Jiang, G.B. Silver nanoparticles induce size-dependent and particle-specific neurotoxicity to primary cultures of rat cerebral cortical neurons. Ecotoxicol. Environ. Saf. 2020, 198, 110674. [Google Scholar] [CrossRef] [PubMed]
- Arrieta-Sandoval, N.; Rojas, P.E.; Olivas-Armendariz, I.; Gomez, L.V.E.; Paz, J.H.F.; Cordero, B.M.E.; Gonzalez, C.R.A. Effect of Ag2S-BSA nanoparticle size on 3T3 fibroblast cell line cytotoxicity. J. Nanopart. Res. 2020, 22, 106. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, S.; Shen, Y.R.; Xiao, Y.Q.; Gao, L.Z.; Shi, S.R. Cytotoxicity studies of Fe3O4 nanoparticles in chicken macrophage cells. R. Soc. Open Sci. 2020, 7, 191561. [Google Scholar] [CrossRef] [Green Version]
- Thiramanas, R.; Jiang, S.; Simon, J.; Landfester, K.; Mailander, V. Silica nanocapsules with different sizes and physicochemical properties as suitable nanocarriers for uptake in T-cells. Int. J. Nanomed. 2020, 15, 6069–6084. [Google Scholar] [CrossRef]
- Colaco, M.; Marques, A.P.; Jesus, S.; Duarte, A.; Borges, O. Safe-by-Design of Glucan nanoparticles: Size matters when assessing the immunotoxicity. Chem. Res. Toxicol. 2020, 33, 915–932. [Google Scholar] [CrossRef]
- Wroblewska-Wolna, A.M.; Harvie, A.J.; Rowe, S.F.; Critchley, K.; Butt, J.N.; Jeuken, L.J.C. Quantum dot interactions with and toxicity to Shewanella oneidensis MR-1. Nanotechnology 2020, 31, 134005. [Google Scholar] [CrossRef]
- Chen, Y.H.; D’Amario, C.; Gee, A.; Duong, H.T.T.; Shimoni, O.; Valenzuela, S.M. Dispersion stability and biocompatibility of four ligand-exchanged NaYF4:Yb, Er upconversion nanoparticles. Acta Biomater. 2020, 102, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, S.; de Haan, L.H.; Evers, N.M.; Jiang, X.; Marcelis, A.T.; Zuilhof, H.; Rietjens, I.M.; Alink, G.M. Role of surface charge and oxidative stress in cytotoxicity of organic monolayer-coated silicon nanoparticles towards macrophage NR8383 cells. Part Fibre Toxicol. 2010, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tada-Oikawa, S.; Eguchi, M.; Yasuda, M.; Izouka, K.; Ikegami, A.; Vranic, S.; Boland, S.; Tran, L.; Ichihara, G.; Ichihara, S. Functionalized surface-charged SiO2 nanoparticles induce pro-inflammatory responses, but are not lethal to Caco-2 cells. Chem. Res. Toxicol. 2020, 33, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Rikimaru-Kaneko, A.; Hashiguchi, K.; Shirotake, S. Effect of anionic and cationic n-butylcyanoacrylate nanoparticles on NO and cytokine production in Raw264.7 cells. Immunopharmacol. Immunotoxicol. 2011, 33, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Pavičić, I.; Milić, M.; Pongrac, I.M.; Ahmed, L.B.; Glavan, T.M.; Ilić, K.; Zapletal, E.; Ćurlin, M.; Mitrečić, D.; Vrček, I.V. Neurotoxicity of silver nanoparticles stabilized with different coating agents: In vitro response of neuronal precursor cells. Food Chem. Toxicol. 2020, 136, 110935. [Google Scholar] [CrossRef]
- Zhai, C.; Zhang, X.; Chen, J.; He, J.; Fei, H.; Liu, Y.; Luo, C.; Fan, W. The effect of cartilage extracellular matrix particle size on the chondrogenic differentiation of bone marrow mesenchymal stem cells. Regen. Med. 2019, 14, 663–680. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Liu, X.; Zhang, R.; Feng, Q. In Vitro Uptake of Hydroxyapatite Nanoparticles and Their Effect on Osteogenic Differentiation of Human Mesenchymal Stem Cells. Stem Cells Int. 2018, 2018, 2036176. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.-S.; Song, I.; Kim, D.-S.; Lee, U.; Kim, J.-K.; Son, H.; Min, J.; Kim, T.-H. Size-dependent effects of graphene oxide on the osteogenesis of human adipose-derived mesenchymal stem cells. Colloids Surf B Biointerfaces 2018, 169, 20–29. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Huang, Q.; Liu, X.; Zhang, R.; Feng, Q. The effect of hydroxyapatite nanoparticles on adipogenic differentiation of human mesenchymal stem cells. J. Biomed. Mater. Res. Part A 2018, 106, 1822–1831. [Google Scholar] [CrossRef]
- Ko, W.-K.; Heo, D.N.; Moon, H.-J.; Lee, S.J.; Bae, M.S.; Lee, J.B.; Sun, I.-C.; Jeon, H.B.; Park, H.K.; Kwon, I.K. The effect of gold nanoparticle size on osteogenic differentiation of adipose-derived stem cells. J. Colloid Interface Sci. 2015, 438, 68–76. [Google Scholar] [CrossRef]
- Lv, L.; Liu, Y.; Zhang, P.; Zhang, X.; Liu, J.; Chen, T.; Su, P.; Li, H.; Zhou, Y. The nanoscale geometry of TiO2 nanotubes influences the osteogenic differentiation of human adipose-derived stem cells by modulating H3K4 trimethylation. Bio-Mater. 2015, 39, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Weissenboeck, M.; Stein, E.; Undt, G.; Ewers, R.; Lauer, G.; Turhani, D. Particle size of hydroxyapatite granules calcified from red algae affects the osteogenic potential of human mesenchymal stem cells in vitro. Cells Tissues Organs 2006, 182, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Guo, Y.Q.; He, X.; Chen, H.Q. Effect of Titanium Particle Size on Osteogenic Differentiation of Bone Marrow–Derived Mesenchymal Stem Cells. Key Eng. Mater. 2011, 474, 1939–1942. [Google Scholar] [CrossRef]
- Smith, P. M. Train; Alfred A. Knopf: New York, NY, USA, 2015. [Google Scholar]
- Uskoković, V. A Collection of Micrographs: Where Science and Art Meet. Technoetic Arts J. Specul. Res. 2010, 7, 231–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uskoković, T.; Uskoković, E.; Wu, V.; Uskoković, V. Calcium Phosphate and Senescence of Orange Jubilees in the Summertime. ACS Appl. Bio Mater. 2020, 3, 3770–3784. [Google Scholar] [CrossRef]
- Uskoković, E.; Uskoković, T.; Wu, V.M.; Uskoković, V. … And All the World a Dream: Memory Outlining the Mysterious Temperature-Dependency of Crystallization of Water, aka the Mpemba Effect. Substantia 2020, 4, 59–117. [Google Scholar]
- Uskoković, V. Ion-Doped Hydroxyapatite: An Impasse or the Road to Follow? Ceram. Int. 2020, 46, 11443–11465. [Google Scholar] [CrossRef]
- Medawar, P. Is the Scientific Paper a Fraud? Listener, 12 September 1963; 377–378. [Google Scholar]
- Uskoković, V.; Batarni, S.S.; Schweicher, J.; King, A.; Desai, T.A. Effect of Calcium Phosphate Particle Shape and Size on their Antibacterial and Osteogenic Activity in the Delivery of Antibiotics in vitro. ACS Appl. Mater. Interfaces 2013, 5, 2422–2431. [Google Scholar] [CrossRef]
- Sekiya, I.; Larson, B.L.; Smith, J.R.; Pochampally, R.; Cui, J.G.; Prockop, D.J. Expansion of human adult stem cells from bone marrow stroma: Conditions that maximize the yields of early progenitors and evaluate their quality. Stem Cells 2002, 20, 530–541. [Google Scholar] [CrossRef]
- Sherer, N.M.; Lehmann, M.J.; Jimenez-Soto, L.F.; Ingmundson, A.; Horner, S.M.; Cicchetti, G.; Allen, P.G.; Pypaert, M.; Cunningham, J.M.; Mothes, W. Visualization of retroviral replication in living cells reveals building into multivesicular bodies. Traffic 2003, 4, 785–801. [Google Scholar] [CrossRef]
- Cox, E.P. A method of assigning numerical and percentage values to the degree of roundness of sand grains. J. Paleontol. 1927, 1, 179–183. [Google Scholar]
- Bagheri, G.; Bonadonna, C. Aerodynamics of Volcanic Particles: Characterization of Size, Shape, and Settling Velocity. In Volcanic Ash; Mackie, S., Cashman, K., Ricketts, H., Rust, A., Watson, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Geng, Y.; Dalhaimer, P.; Cai, S.; Tsai, R.; Tewari, M.; Minko, T.; Discher, D.E. Shape effects of filaments versus spherical particles in flow and drug delivery. Nat. Nanotechnol. 2007, 2, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Treuel, L.; Jiang, X.; Nienhaus, G.U. New views on cellular uptake and trafficking of manufactured nanoparticles. J. Royal Soc. Interface 2013, 10, 20120939. [Google Scholar] [CrossRef] [PubMed]
- Loos, C.; Syrovets, T.; Musyanovych, A.; Mailander, V.; Landfester, K.; Nienhaus, G.U.; Simmet, T. Functionalized polystyrene nanoparticles as a platform for studying bio-nano interactions. Beilstein. J. Nanotechnol. 2014, 5, 2403–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florencio-Silva, R.; Sasso, G.R.S.; Sasso-Cerri, E.; Simões, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. Biomed. Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef] [Green Version]
- Rau, J.V.; Fosca, M.; Fadeeva, I.V.; Kalay, S.; Culha, M.; Raucci, M.G.; Fasolino, I.; Ambrosio, L.; Antoniac, I.V.; Uskoković, V. Tricalcium Phosphate Bone Cement Supplemented with Boron Nitride Nanotubes with Enhanced Biological Properties. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 114, 111044. [Google Scholar] [CrossRef]
- Thorpe, A.A.; Creasey, S.; Sammon, C.; le Maitre, C.L. Hydroxyapatite nanoparticle injectable hydrogel scaffold to support osteogenic differentiation of human mesenchymal stem cells. Eur. Cell Mater. 2016, 5, 1–23. [Google Scholar] [CrossRef]
- Xu, S.J.; Qiu, Z.Y.; Wu, J.J.; Kong, X.D.; Weng, X.S.; Cui, F.Z.; Wang, X.M. Osteogenic Differentiation Gene Expression Profiling of hMSCs on Hydroxyapatite and Mineralized Collagen. Tissue Eng. Part A 2016, 22, 170–181. [Google Scholar] [CrossRef]
- Pino, A.M.; Rosen, C.J.; Rodríguez, J.P. In Osteoporosis, differentiation of mesenchymal stem cells (MSCs) improves bone marrow adipogenesis. Biol. Res. 2012, 45, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Tian, X.; Kim, J.P.; Xie, D.; Ao, X.; Shan, D.; Lin, Q.; Hudock, M.R.; Bai, X.; Yang, J. Citrate-based materials fuel human stem cells by metabonegenic regulation. PNAS 2018, 115, 11741–11750. [Google Scholar] [CrossRef] [Green Version]
- Hoh, J.H.; Schoenenberger, C.A. Surface morphology and mechanical properties of MDCK monolayers by atomic force microscopy. J. Cell Sci. 1994, 107, 1105–1114. [Google Scholar] [PubMed]
- Swanson, J.A.; Watts, C. Macropinocytosis. Trends Cell Biol. 1995, 5, 424–428. [Google Scholar] [CrossRef]
- Rejman, J.; Oberle, V.; Zuhorn, I.S.; Hoekstra, D. Size-dependent internalization of particles via the pathways of clathrin- and caveolae-mediated endocytosis. Biochem. J. 2004, 377 Pt 1, 159–169. [Google Scholar] [CrossRef]
- Yi, X.; Shi, X.H.; Gao, H.J. A universal law for cell uptake of one-dimensional nanomaterials. Nano Lett. 2014, 14, 1049–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Gao, H.; Bao, G. Physical principles of nanoparticle cellular endocytosis. ACS Nano 2015, 9, 8655–8671. [Google Scholar] [CrossRef] [Green Version]
- De Saint-Exupery, A. The Little Prince; Reynal & Hitchcock: New York, NY, USA, 1943. [Google Scholar]
- Guduru, R. In Situ AFM Imaging of Nanoparticle-Cellular Membrane Interactions for a Drug Delivery Study. Master of Science Thesis, Florida International University, Miami, FL, USA, 2011. [Google Scholar]
- Uskoković, V. When 1 + 1 > 2: Nanostructured Composite Materials for Hard Tissue Engineering Applications. Mater. Sci. Eng. C Mater. Biol. Appl. 2015, 57, 434–451. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.S.; Cao, J.; Naeem, M.; Noh, J.; Hasan, N.; Choi, H.K.; Yoo, J.W. Size-controlled biodegradable nanoparticles: Preparation and size-dependent cellular uptake and tumor cell growth inhibition. Coll. Surf. B 2014, 122, 545–551. [Google Scholar] [CrossRef]
- He, C.; Hu, Y.; Yin, L.; Tang, C.; Yin, C. Effects of particle size and surface charge on cellular uptake and biodistribution of polymeric nanoparticles. Biomaterials 2010, 31, 3657–3666. [Google Scholar] [CrossRef]
- Chono, S.; Tanino, T.; Seki, T.; Morimoto, K. Uptake characteristics of liposomes by rat alveolar macrophages: Influence of particle size and surface mannose modification. J. Pharm. Pharmacol. 2007, 59, 75–80. [Google Scholar] [CrossRef]
- Dalzon, B.; Torres, A.; Reymond, S.; Gallet, B.; Saint-Antonin, F.; Collin-Faure, V.; Moriscot, C.; Fenel, D.; Schoehn, G.; Aude-Garcia, C.; et al. Influences of Nanoparticles Characteristics on the Cellular Responses: The Example of Iron Oxide and Macrophages. Nanomaterials 2020, 10, 226. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Shi, W.; Freund, L.B. Mechanics of receptor-mediated endocytosis. Proc. Natl. Acad. Sci. USA 2005, 102, 9469–9474. [Google Scholar] [CrossRef] [Green Version]
- Shang, L.; Nienhaus, K.; Jiang, X.; Yang, L.; Landfester, L.; Mailander, V.; Simmet, T.; Nienhaus, G.U. Nanoparticles interactions with live cells: Quantitative fluorescence microscopy of nanoparticles size effects. Beilstein J. Nanotechnol. 2014, 5, 2388–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chithrani, B.D.; Chan, W.C.W. Elucidating the mechanism of cellular uptake and removal of protein-coated gold nanoparticles of different sizes and shapes. Nano Lett. 2007, 7, 1542–1550. [Google Scholar] [CrossRef] [PubMed]
- Osaki, F.; Kanamori, T.; Sando, S.; Sera, T.; Aoyama, Y. A quantum dot conjugated sugar ball and its cellular uptake. On the size effects of endocytosis in the subviral region. J. Am. Chem. Soc. 2004, 126, 6520–6521. [Google Scholar] [CrossRef] [PubMed]
- Gratton, S.E.A.; Ropp, P.A.; Pohlhaus, P.D.; Luft, J.C.; Madden, V.J.; Napier, M.E.; DeSimone, J.M. The effect of particle design on cellular internalization pathways. Proc. Natl. Acad. Sci. USA 2008, 105, 11613–11618. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Wu, S.; Hung, Y.; Mou, C.Y. Size effect on cell uptake in well-suspended, uniform mesoporous silica nanoparticles. Small 2009, 5, 1408–1413. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.C.; Wright, D.W. Size-dependent cellular uptake of DNA functionalized gold nanoparticles. Small 2016, 12, 5592–5600. [Google Scholar] [CrossRef]
- Yao, M.; He, L.; McClements, D.J.; Xiao, H. Uptake of gold nanoparticles by intestinal epithelial cells: Impact of particle size on their absorption, accumulation, and toxicity. J. Agric. Food Chem. 2015, 63, 8044–8049. [Google Scholar] [CrossRef]
- Limbach, L.K.; Li, Y.; Grass, R.N.; Brunner, T.J.; Hintermann, M.A.; Muller, M.; Gunther, D.; Stark, W.J. Oxide nanoparticle uptake in human lung fibroblasts: Effects of particle size, agglomeration, and diffusion at low concentrations. Environ. Sci. Technol. 2005, 39, 9370–9376. [Google Scholar] [CrossRef]
- Lewis, E.E.L.; Child, H.W.; Hursthouse, A.; Stirling, D.; McCully, M.; Paterson, D.; Mullin, M.; Berry, C.C. The influence of particle size and static magnetic fields on the uptake of magnetic nanoparticles into three dimensional cell-seeded collagen gel cultures. J. Biomed. Mat. Res. 2015, 103, 1294–1301. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.H.; Lee, C.W.; Chiou, A.; Wei, P.K. Size-dependent endocytosis of gold nanoparticles studied by three-dimensional mapping of plasmonic scattering images. J. Nanobiotechnol. 2010, 8, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foged, C.; Brodin, B.; Frokjaer, S.; Sundblad, A. Particle size and surface charge affect particle uptake by human dendritic cells in an in vitro model. Int. J. Pharm. 2005, 298, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Zauner, W.; Farrow, N.A.; Haines, A.M.R. In vitro uptake of polystyrene microspheres: Effect of particle size, cell line and cell density. J. Controlled Release 2001, 71, 39–51. [Google Scholar] [CrossRef]
- Peruzynska, M.; Cendrowski, K.; Barylak, M.; Roginska, D.; Tarnowski, M.; Tkacz, M.; Kurzawski, M.; Machalinski, B.; Mijowska, E.; Drozdzik, M. Study on size effect of the silica nanospheres with solid core and mesoporous shell on cellular uptake. Biomed. Mater. 2015, 10, 065012. [Google Scholar] [CrossRef]
- Ferrari, R.; Lupi, M.; Colombo, C.; Morbidelli, M.; D’Incalci, M.; Moscatelli, D. Investigation of size, surface charge, PEGylation degree and concentration on the cellular uptake of polymer nanoparticles. Coll. Surf. B 2014, 123, 639–647. [Google Scholar] [CrossRef]
- Dos Santos, T.; Varela, J.; Lynch, I.; Salvati, A.; Dawson, K.A. Quantitative assessment of the comparative nanoparticle-uptake efficiency of a range of cell lines. Small 2011, 7, 3341–3349. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Heller, D.A.; Sharma, R.; Strano, M.S. Size-dependent cellular uptake and expulsion of single-walled carbon nanotubes: Single particle tracking and a generic uptake model for nanoparticles. ACS Nano 2009, 3, 149–158. [Google Scholar] [CrossRef]
- Parakhonskiy, B.; Zyuzin, M.V.; Yashchenok, A.; Carregal-Romero, S.; Rejman, J.; Mohwald, H.; Parak, W.J.; Skirtach, A.G. The influence of the size and aspect ratio of anisotropic, porous CaCO3 particles on their uptake by cells. J. Nanobiotechnol. 2015, 12, 53. [Google Scholar] [CrossRef] [Green Version]
- Kashani, A.S.; Badilescu, S.; Piekny, A.; Packirisamy, M. Differing Affinities of Gold Nanostars and Nanospheres toward HeLa and HepG2 Cells: Implications for Cancer Therapy. ACS Appl. Nano Mater. 2020, 3, 4114–4126. [Google Scholar] [CrossRef]
- Zhang, Z.K.; Liu, C.R.; Li, C.; Wu, W.; Jiang, X.Q. Shape Effects of Cylindrical versus Spherical Unimolecular Polymer Nanomaterials on in Vitro and in Vivo Behaviors. Research 2019, 2019, 2391484. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, J.; Wang, D.D.; Li, D.; Yang, Y.Q.; Lu, Y.; Wu, W.; Wu, W.; Qi, J.P. The influence of nanoparticle shape on bilateral exocytosis from Caco-2 cells. Chin. Chem. Lett. 2018, 29, 1815–1818. [Google Scholar] [CrossRef]
- Xu, D.; Wan, Y.X.; Li, Z.H.; Wang, C.B.; Zou, Q.X.; Du, C.; Wang, Y.J. Tailorable hierarchical structures of biomimetic hydroxyapatite micro/nano particles promoting endocytosis and osteogenic differentiation of stem cells. Biomater. Sci. 2020, 8, 3286–3300. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Liu, H.; Hu, X.; Chen, Z.; Friis, T.E.; Wang, J.; Xiao, Y.; Zhang, S. Bio-inspired hybrid nanoparticles promote vascularized bone regeneration in a morphology-dependent manner. Nanoscale 2017, 9, 5794–5805. [Google Scholar] [CrossRef] [PubMed]
- Nambara, K.; Nikura, K.; Mitomo, H.; Ninomiya, T.; Takeuchi, C.; Wei, J.; Matsuo, Y.; Ijiro, K. Reverse size dependences of the cellular uptake of triangular and spherical gold nanoparticles. Langmuir 2016, 32, 12559–12567. [Google Scholar] [CrossRef]
- Farge, E.; Devaux, P.F. Shape changes of giant liposomes induced by an asymmetric transmembrane distribution of phospholipids. Biophys. J. 1992, 61, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Uskoković, V.; Drofenik, M. Reverse Micelles: Inert Nano-Reactors or Physico-Chemically Active Guides of the Capped Reactions. Adv. Colloid Interface Sci. 2007, 133, 23–34. [Google Scholar] [CrossRef]
- Champion, J.A.; Mitragotri, S. Shape induced inhibition of phagocytosis of polymer particles. Pharm. Res. 2009, 26, 244–249. [Google Scholar] [CrossRef] [Green Version]
- Uskoković, V. Nanostructured Platforms for the Sustained and Local Delivery of Antibiotics in the Treatment of Osteomyelitis. Crit. Rev. Ther. Drug Carr. Syst. 2015, 32, 1–59. [Google Scholar] [CrossRef]
- Rasel, M.A.I.; Singh, S.; Nguyen, T.D.; Afara, I.O.; Gu, Y. Impact of Nanoparticle Uptake on the Biophysical Properties of Cell for Biomedical Engineering Applications. Sci. Rep. 2019, 9, 5859. [Google Scholar] [CrossRef] [Green Version]
- Yin, M.; Xu, W.G.; Cui, B.C.; Dai, H.L.; Han, Y.C.; Yin, Y.X.; Li, S.P. Effects of the interaction between hydroxyapatite nanoparticles and hepatoma cells. J. Wuhan Univ. Technol. Mat. Sci. Ed. 2014, 29, 635–642. [Google Scholar] [CrossRef]
- Yin, M.; Yin, Y.; Han, Y.; Dai, H.; Li, S. Effects of Uptake of Hydroxyapatite Nanoparticles into Hepatoma Cells on Cell Adhesion and Proliferation. J. Nanomater. 2014, 2014, 731897. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, T.; Fratini, M.; Frey, F.; Yserentant, K.; Liu, Y.; Weber, E.; Galior, K.; Ohmes, J.; Braun, F.; Herten, D.P.; et al. Forces during cellular uptake of viruses and nanoparticles at the ventral side. Nat. Commun. 2020, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Bischofs, I.B.; Schmidt, S.S.; Schwarz, U.S. Effect of Adhesion Geometry and Rigidity on Cellular Force Distributions. Phys. Rev. Lett. 2009, 103, 048101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonelli, F.M.P.; Santos, A.K.; Gomes, D.A.; da Silva, S.L.; Gomes, K.N.; Ladeira, L.O.; Resende, R.R. Stem cells and calcium signaling. Adv. Exp. Med. Biol. 2012, 740, 891–916. [Google Scholar] [PubMed] [Green Version]
- Uskoković, V.; Desai, T.A. Does Translational Symmetry Matter on the Micro Scale? Fibroblastic and Osteoblastic Interactions with the Topographically Distinct Poly(ε-Caprolactone)/Hydroxyapatite Thin Films. ACS Appl. Mater. Interfaces 2014, 6, 13209–13220. [Google Scholar] [CrossRef]
- Hanna, H.; Mir, L.M.; Andre, F.M. In vitro osteoblastic differentiation of mesenchymal stem cells generates cell layers with distinct properties. Stem Cell Res. Ther. 2018, 9, 203. [Google Scholar] [CrossRef]
- Ghiasi, B.; Sefidbakht, Y.; Mozaffari-Jovin, S.; Gharachloo, B.; Mehraria, M.; Khodadadi, A.; Rezaei, M.; Ranaei-Siadat, S.O.; Uskoković, V. Hydroxyapatite as a Biomaterial–A Gift that Keeps on Giving. Drug Dev. Ind. Pharm. 2020, 46, 1035–1062. [Google Scholar] [CrossRef]
- Khan, M.A.; Wu, V.M.; Ghosh, S.; Uskoković, V. Gene Delivery Using Calcium Phosphate Nanoparticles: Optimization of the Transfection Process and the Effects of Citrate and Poly(L-Lysine) as Additives. J. Colloid Interface Sci. 2016, 471, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Wu, V.M.; Uskoković, V. Waiting for Aπαταο: 250 Years Later. Found. Sci. 2019, 24, 617–640. [Google Scholar] [CrossRef]
- Wu, V.M.; Uskoković, V. Is There a Relationship between Solubility and Resorbability of Different Calcium Phosphate Phases in vitro? Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 2157–2168. [Google Scholar] [CrossRef] [Green Version]
- Hallermann, S. Calcium channels for endocytosis. J. Physiol. 2014, 592 (Pt 16), 3343–3344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.Y.; Chen, H.M.; Xie, J.; Becton, M.; Wang, X.Q. Interplay of Nanoparticle Rigidity and Its Translocation Ability through Cell Membrane. J. Phys. Chem. B 2019, 123, 8923–8930. [Google Scholar] [CrossRef] [PubMed]
- Uskoković, V. The Role of Hydroxyl Channel in Defining Selected Physicochemical Peculiarities Exhibited by Hydroxyapatite. RSC Adv. 2015, 5, 36614–36633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey, C.; Combes, C.; Drouet, C.; Glimcher, M.J. Bone mineral: Update on chemical composition and structure. Osteoporos. Int. 2009, 20, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Uskoković, V.; Tang, S.; Nikolić, M.G.; Marković, S.; Wu, V.M. Calcium Phosphate Nanoparticles as Intrinsic Inorganic Antimicrobials: In Search of the Key Particle Property. Biointerphases 2019, 14, 031001. [Google Scholar] [CrossRef]
- Wu, V.M.; Mickens, J.; Uskoković, V. Bisphosphonate-Functionalized Calcium Phosphate Nanoparticles for the Delivery of the Bromodomain Inhibitor JQ1 in the Treatment of Osteosarcoma. ACS Appl. Mater. Interfaces 2017, 9, 25887–25904. [Google Scholar] [CrossRef]
- Pernal, S.P.; Wu, V.M.; Uskoković, V. Hydroxyapatite as a Vehicle for the Selective Effect of Superparamagnetic Iron Oxide Nanoparticles against Human Glioblastoma Cells. ACS Appl. Mater. Interfaces 2017, 9, 39283–39302. [Google Scholar] [CrossRef]
- Shi, X.X.; Zhou, K.; Huang, F.; Zhang, J.; Wang, C. Endocytic mechanisms and osteoinductive profile of hydroxyapatite nanoparticles in human umbilical cord Wharton’s jelly-derived mesenchymal stem cells. Int. J. Nanomed. 2018, 13, 1457–1470. [Google Scholar] [CrossRef] [Green Version]
- Uskoković, V. Celeste’s Plight: What Can Film Teach Natural Science? Film Int. 2019, 17, 69–88. [Google Scholar] [CrossRef]
Figure 1.
The randomly taken atomic force microscope (AFM) image of hMSCs challenged with HAp nanoparticles, used as the starting point for inquiry into the interaction between nanoparticles and cells.
Figure 1.
The randomly taken atomic force microscope (AFM) image of hMSCs challenged with HAp nanoparticles, used as the starting point for inquiry into the interaction between nanoparticles and cells.

Figure 2.
Representative optical fluorescent micrograph of control hMSCs, not subjected to interaction with the nanoparticles. Cells were stained for nucleus using DAPI (blue) and for f-actin cytoskeletal microfilaments using phalloidin (red). Scale bar equals 50 μm.
Figure 2.
Representative optical fluorescent micrograph of control hMSCs, not subjected to interaction with the nanoparticles. Cells were stained for nucleus using DAPI (blue) and for f-actin cytoskeletal microfilaments using phalloidin (red). Scale bar equals 50 μm.

Figure 3.
Magnified portions of Figure 1 (a–d), along with an SEM image of HAp nanoparticles (e), confirming that what is visible in Figure 1 are agglomerates of individual nanoparticles rather than nanoparticles per se. White arrows point at the key particles discussed in the text. The dashed rectangle in (c) denotes an area brimming with spherical entities, which are fine nanoparticle agglomerates resting on or inside the cell.
Figure 3.
Magnified portions of Figure 1 (a–d), along with an SEM image of HAp nanoparticles (e), confirming that what is visible in Figure 1 are agglomerates of individual nanoparticles rather than nanoparticles per se. White arrows point at the key particles discussed in the text. The dashed rectangle in (c) denotes an area brimming with spherical entities, which are fine nanoparticle agglomerates resting on or inside the cell.

Figure 4.
Fluorescent optical images of stained hMSCs and HAp nanoparticles. Both cells in the foci of images (a,b) display the uptake of finer nanoparticle aggregates, but also a single large agglomerate on the edge of the cell nucleus. Arrow in (a) points at the intact cell nucleus area overlapping with the nanoparticle agglomerate signal, while the arrow in (b) points at the indented cell nucleus area in contact with the apparently intracellular irregularly shaped and sizable nanoparticle agglomerate. DAPI was used to stain the cell nuclei in blue, phalloidin to stain f-actin microfilaments in red, and OsteoImage to stain HAp nanoparticles in green.
Figure 4.
Fluorescent optical images of stained hMSCs and HAp nanoparticles. Both cells in the foci of images (a,b) display the uptake of finer nanoparticle aggregates, but also a single large agglomerate on the edge of the cell nucleus. Arrow in (a) points at the intact cell nucleus area overlapping with the nanoparticle agglomerate signal, while the arrow in (b) points at the indented cell nucleus area in contact with the apparently intracellular irregularly shaped and sizable nanoparticle agglomerate. DAPI was used to stain the cell nuclei in blue, phalloidin to stain f-actin microfilaments in red, and OsteoImage to stain HAp nanoparticles in green.

Figure 5.
The significantly lower average size of the surface projection of optically imaged intracellular (IC) HAp nanoparticle agglomerates in interaction with hMSCs (n = 40) compared to that of the extracellular (EC) nanoparticle agglomerates (a). Granularity of hMSCs challenged with HAp nanoparticles approaching 100%, showing that practically all cells uptake the nanoparticles (n = 3) (b). Meta-analysis of literature data representing the ratio between the uptaken concentration of larger particles and the uptaken concentration of smaller particles (Ux/Uy), as a function of the ratio between the size of the larger particles and the size of the smaller particles (Dx/Dy) in various cell types, excluding macrophages (c). Physical qualities measured to assess the difference in the uptake efficiency between differently sized particles varied between studies and included percent internalization (Gratton et al. [60]), weight of the exogenous ion determined by ICP-MS per cell (Lu et al. [61], Wong et al. [62], Yao et al. [63], Limbach et al. [64], Lewis et al. [65]), the number of nanoparticles per cell derived from plasmonic scattering (Wang et al. [66]), uptake rate via flow cytometry (Foged et al. [67], Zauner et al. [68] Peruzynska et al. [69], Ferrari et al. [70], dos Santos et al. [71]), or fluorescence signal intensity (Jin et al. [72], Parakhonskiy et al. [73]). No information on particles lower in size than 20 nm was included in the analysis. The dashed line demarcates the better uptake of larger particles above it from the better uptake of smaller particles below it. Abbreviations: PS = polystyrene, PEG = poly(ethylene glycol), SWNT = single-walled carbon nanotubes, PEG-PCL = PEGylated polycaprolactone, COOH-PS = carboxylated polystyrene. Data points in (a,b) represent averages, error bars represent standard deviation, and p-values represent the confidence intervals for the statistical significance of the difference between the sample groups.
Figure 5.
The significantly lower average size of the surface projection of optically imaged intracellular (IC) HAp nanoparticle agglomerates in interaction with hMSCs (n = 40) compared to that of the extracellular (EC) nanoparticle agglomerates (a). Granularity of hMSCs challenged with HAp nanoparticles approaching 100%, showing that practically all cells uptake the nanoparticles (n = 3) (b). Meta-analysis of literature data representing the ratio between the uptaken concentration of larger particles and the uptaken concentration of smaller particles (Ux/Uy), as a function of the ratio between the size of the larger particles and the size of the smaller particles (Dx/Dy) in various cell types, excluding macrophages (c). Physical qualities measured to assess the difference in the uptake efficiency between differently sized particles varied between studies and included percent internalization (Gratton et al. [60]), weight of the exogenous ion determined by ICP-MS per cell (Lu et al. [61], Wong et al. [62], Yao et al. [63], Limbach et al. [64], Lewis et al. [65]), the number of nanoparticles per cell derived from plasmonic scattering (Wang et al. [66]), uptake rate via flow cytometry (Foged et al. [67], Zauner et al. [68] Peruzynska et al. [69], Ferrari et al. [70], dos Santos et al. [71]), or fluorescence signal intensity (Jin et al. [72], Parakhonskiy et al. [73]). No information on particles lower in size than 20 nm was included in the analysis. The dashed line demarcates the better uptake of larger particles above it from the better uptake of smaller particles below it. Abbreviations: PS = polystyrene, PEG = poly(ethylene glycol), SWNT = single-walled carbon nanotubes, PEG-PCL = PEGylated polycaprolactone, COOH-PS = carboxylated polystyrene. Data points in (a,b) represent averages, error bars represent standard deviation, and p-values represent the confidence intervals for the statistical significance of the difference between the sample groups.

Figure 6.
Identical circularities of HAp nanoparticle agglomerates localized intracellularly (IC) and extracellularly (EC) with respect to the hMSCs (a). Statistically insignificantly lower fluorescence originating from internalized needle-shaped HAp nanoparticles (c) compared to that of their brick-shaped counterparts (d), indicating that the uptake is morphologically dependent for samples composed of nanoparticles with unique morphologies (b), despite being morphologically independent with respect to geometries of agglomerates of nanoparticles in single samples. Data points in (a,b) represent averages, error bars represent standard deviation, and p-values represent the confidence intervals for the statistical significance of the difference between the sample groups.
Figure 6.
Identical circularities of HAp nanoparticle agglomerates localized intracellularly (IC) and extracellularly (EC) with respect to the hMSCs (a). Statistically insignificantly lower fluorescence originating from internalized needle-shaped HAp nanoparticles (c) compared to that of their brick-shaped counterparts (d), indicating that the uptake is morphologically dependent for samples composed of nanoparticles with unique morphologies (b), despite being morphologically independent with respect to geometries of agglomerates of nanoparticles in single samples. Data points in (a,b) represent averages, error bars represent standard deviation, and p-values represent the confidence intervals for the statistical significance of the difference between the sample groups.

Figure 7.
Lysosomal-associated membrane protein 1 (LAMP1) red stains achieved by the transient transfection of control cells (a) and cells challenged with nanoparticles (b) with LAMP1-YFP. Nuclear DNA was stained in blue using DAPI. Yellow arrows in (b) indicate the slightly elongated or misshaped lysosomes coexisting with their spherical counterparts and mostly prominent in the cells undergoing the nanoparticle uptake.
Figure 7.
Lysosomal-associated membrane protein 1 (LAMP1) red stains achieved by the transient transfection of control cells (a) and cells challenged with nanoparticles (b) with LAMP1-YFP. Nuclear DNA was stained in blue using DAPI. Yellow arrows in (b) indicate the slightly elongated or misshaped lysosomes coexisting with their spherical counterparts and mostly prominent in the cells undergoing the nanoparticle uptake.

Figure 8.
Control cells (a) and cells treated with the nanoparticles (b) stained for their nuclei (blue) and collagen production (red). Proliferation of control cells reseeded in cell culture dishes and cells that underwent the uptake of HAp nanoparticles prior to reseeding (c). hMSC viability of population subjected to HAp nanoparticle uptake relative to the viability of the control population (d). Data points in (c,d) represent averages (n = 2 − 4), error bars represent standard deviation, and p-values represent the confidence intervals for the statistical significance of the difference of the value of the sample group compared to the control.
Figure 8.
Control cells (a) and cells treated with the nanoparticles (b) stained for their nuclei (blue) and collagen production (red). Proliferation of control cells reseeded in cell culture dishes and cells that underwent the uptake of HAp nanoparticles prior to reseeding (c). hMSC viability of population subjected to HAp nanoparticle uptake relative to the viability of the control population (d). Data points in (c,d) represent averages (n = 2 − 4), error bars represent standard deviation, and p-values represent the confidence intervals for the statistical significance of the difference of the value of the sample group compared to the control.

Figure 9.
Schematization of the discrete adhesion model proposed by Bischofs et al., and applied to estimate the force of adhesion of the cells to the underlying surface (a). The model predicts the direct dependence of the adhesion force on the cell dimensions defined by the cell side length, d, and on the surface tension, σ. The surface tension can be further expressed as inversely proportional to the opening angle at the cell corners, ϕ, as direct adhesion sites. A representative hMSC stained for f-actin in red and the nucleus in blue, along with the derived parameters d and ϕ (b). Comparison of d/ϕ ratio directly indicative of the adhesion force for the control, untreated hMSCs and for hMSCs that underwent HAp nanoparticle uptake (c). Data points represent averages (n = 16), error bars represent standard deviation, and p-value represents the confidence interval for the statistical significance of the difference of the value of the sample group compared to the control.
Figure 9.
Schematization of the discrete adhesion model proposed by Bischofs et al., and applied to estimate the force of adhesion of the cells to the underlying surface (a). The model predicts the direct dependence of the adhesion force on the cell dimensions defined by the cell side length, d, and on the surface tension, σ. The surface tension can be further expressed as inversely proportional to the opening angle at the cell corners, ϕ, as direct adhesion sites. A representative hMSC stained for f-actin in red and the nucleus in blue, along with the derived parameters d and ϕ (b). Comparison of d/ϕ ratio directly indicative of the adhesion force for the control, untreated hMSCs and for hMSCs that underwent HAp nanoparticle uptake (c). Data points represent averages (n = 16), error bars represent standard deviation, and p-value represents the confidence interval for the statistical significance of the difference of the value of the sample group compared to the control.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Uskoković, V. Blowup of Accidental Images as a Passageway to Discovery: Insights into the Interaction between Hydroxyapatite Nanoparticles and Human Mesenchymal Stem Cells. Appl. Sci. 2020, 10, 8204. https://doi.org/10.3390/app10228204
AMA Style
Uskoković V. Blowup of Accidental Images as a Passageway to Discovery: Insights into the Interaction between Hydroxyapatite Nanoparticles and Human Mesenchymal Stem Cells. Applied Sciences. 2020; 10(22):8204. https://doi.org/10.3390/app10228204
Chicago/Turabian StyleUskoković, Vuk. 2020. "Blowup of Accidental Images as a Passageway to Discovery: Insights into the Interaction between Hydroxyapatite Nanoparticles and Human Mesenchymal Stem Cells" Applied Sciences 10, no. 22: 8204. https://doi.org/10.3390/app10228204
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.