LC-MS Profile, Gastrointestinal and Gut Microbiota Stability and Antioxidant Activity of Rhodiola rosea Herb Metabolites: A Comparative Study with Subterranean Organs

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Chemicals
2.2. Chemical Composition Analysis of R. rosea Organs and Antioxidant Activity Assays
2.3. Total Extract Preparation from R. rosea Organs
2.4. Solid-Phase Extratcion (SPE) of Total Extract from R. rosea Organs
2.5. High-Performance Liquid Chromatography with Diode Array Detection and Electrospray Ionization Triple Quadrupole Mass Spectrometric Detection (HPLC-DAD-ESI-QQQ-MS)
2.6. Flavonol O-Glycosides Hydrolysis and HPLC-DAD-ESI-QQQ-MS Analysis
2.7. HPLC-MS Quantification
2.8. Simulated Gastrointestinal Digestion and Gut Microbiota Incubation
2.9. HPLC-DAD Assays for Gastric, Intestinal, and Gut Microbiota Media
2.10. Incubation of R. rosea Extracts with Digestive Enzymes Mixture
2.11. Trolox-Equivalent Content in Simulated Gastric, Intestinal and Gut Microbiota Media
2.12. Statistical Analysis
3. Results and Discussion
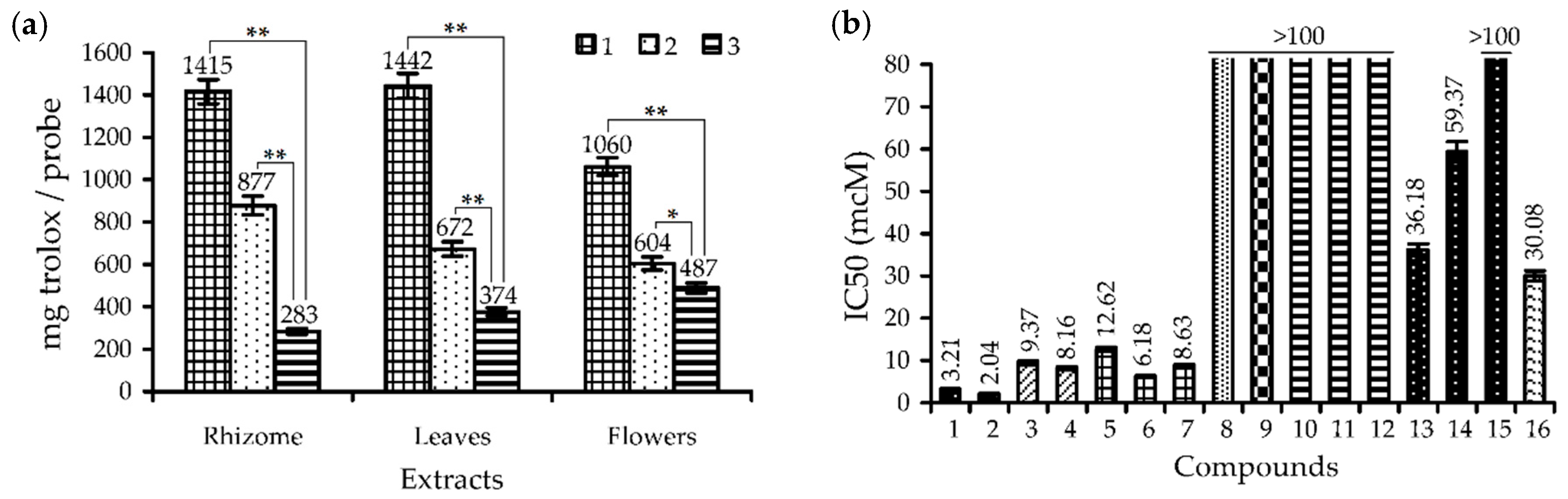
3.1. Phenolic Composition and Antioxidant Activity of Rhodiola rosea Extracts
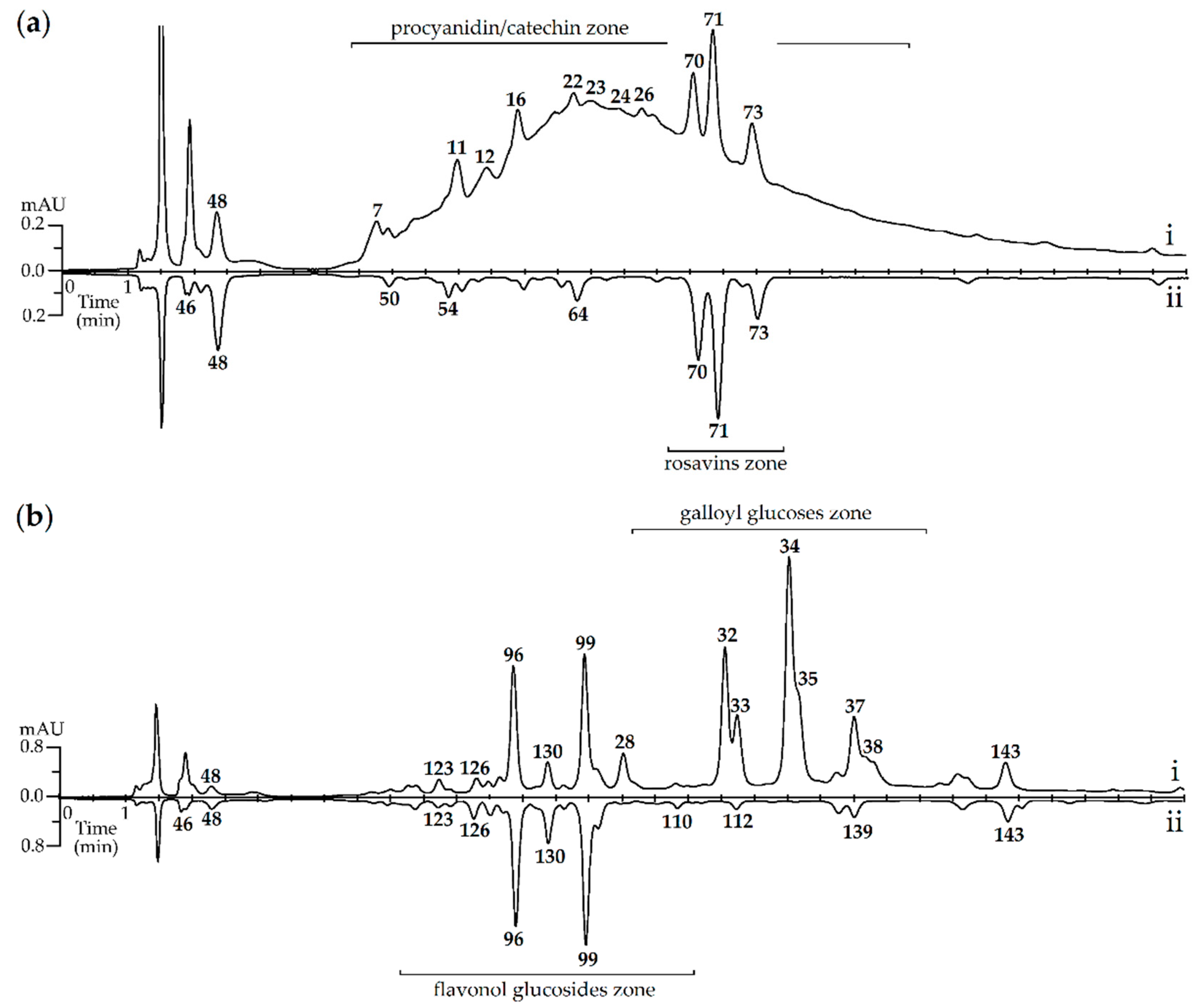
3.2. HPLC-DAD-QQQ-ESI-MS Profiles of R. rosea Organs: Quantitative and Quantitative Study
3.2.1. Galloyl O-Glycosides
3.2.2. Catechins
3.2.3. Procyanidins
3.2.4. Simple Phenolics
3.2.5. Phenethyl Alcohol Derivatives
3.2.6. (Hydroxy)Cinnamates
3.2.7. Hydroxynitrile Glucosides
3.2.8. Monoterpene O-Glucosides
3.2.9. Flavonol O-Glycosides
3.2.10. Comparative Chemodiversity of Various Organs of R. rosea
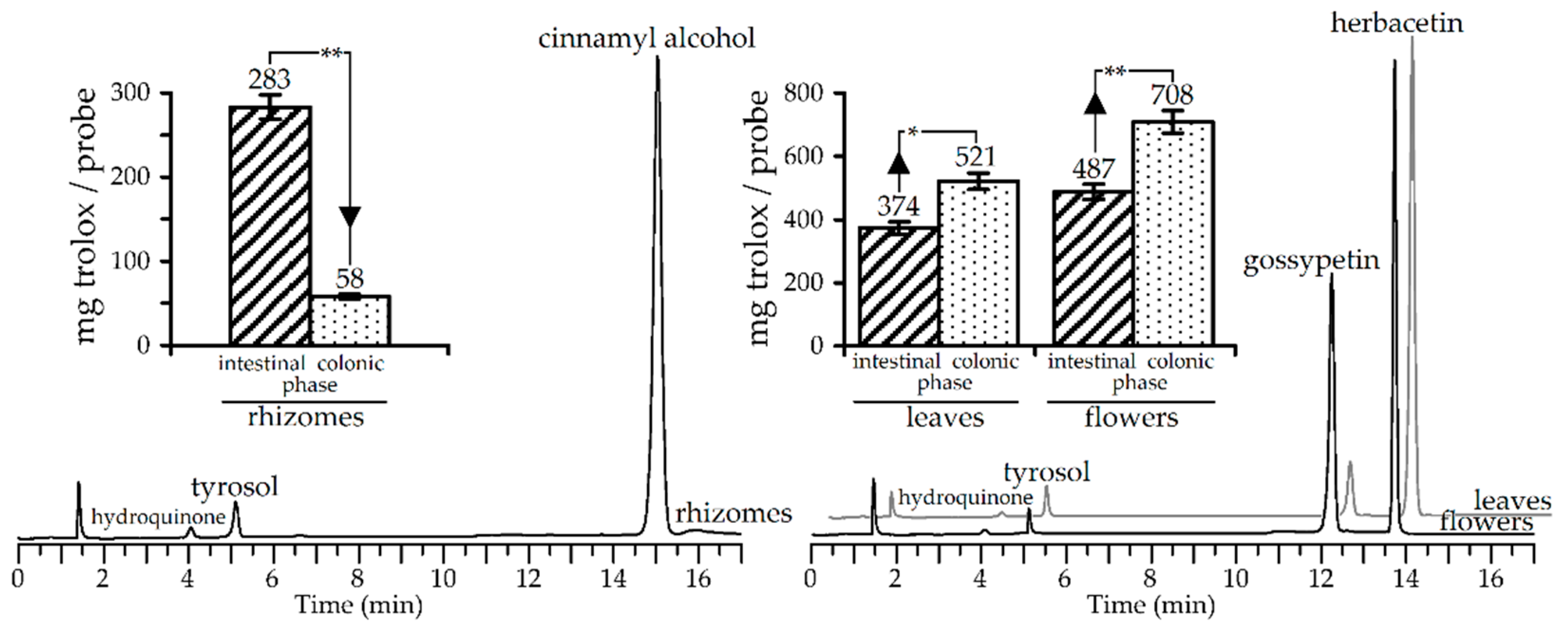
3.3. Stability of Metabolites and Antioxidant Activity of Rhodiola rosea Extracts in Simulated Gastrointestinal Digestion Model
3.4. Gut Microbiota Metabolites of R. rosea Extracts and Their Antioxidant Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cunningham, A.B.; Li, H.L.; Luo, P.; Zhao, W.J.; Long, X.C.; Brinckmann, J.A. There “ain’t no mountain high enough”?: The drivers, diversity and sustainability of China’s Rhodiola trade. J. Ethnopharmacol. 2020, 252, 112379. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Wu, X.; Cao, J.; Peng, Y.; Wang, A.; Pei, J.; Xiao, J.; Wang, S.; Wang, Y. Rhodiola species: A comprehensive review of traditional use, phytochemistry, pharmacology, toxicity, and clinical study. Med. Res. Rev. 2019, 2019, 1–72. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, J.; Shi, R.; Li, N.; Xu, Z.; Sun, M. Antioxidative effects of Rhodiola genus: Phytochemistry and pharmacological mechanisms against the diseases. Curr. Top. Med. Chem. 2017, 17, 1692–1708. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Pham, V.; Bui, M.; Song, L.; Wu, C.; Walia, A.; Uchio, E.; Smith-Liu, F.; Zi, X. Rhodiola rosea L.: An herb with anti-stress, anti-aging, and immunostimulating properties for cancer chemoprevention. Curr. Pharmacol. Rep. 2017, 3, 384–395. [Google Scholar] [CrossRef]
- Liu, Y.; Weng, W.; Gao, R.; Liu, Y.; Monacelli, F. New insights for cellular and molecular mechanisms of aging and aging-related diseases: Herbal medicine as potential therapeutic approach. Oxid. Med. Cell. Long. 2019, 2019, 4598167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anghelescu, I.-G.; Edwards, D.; Seifritz, E.; Kasper, S. Stress management and the role of Rhodiola rosea: A review. Int. J. Psych. Clin. Pract. 2018, 22, 242–252. [Google Scholar] [CrossRef] [Green Version]
- Pu, W.-L.; Zhang, M.-Y.; Bai, R.-Y.; Sun, L.-K.; Li, W.-H.; Yu, Y.-L.; Zhang, Y.; Song, L.; Wang, Z.-X.; Peng, Y.-F.; et al. Anti-inflammatory effects of Rhodiola rosea L.: A review. Biomed. Pharmacother. 2020, 121, 109552. [Google Scholar] [CrossRef]
- Panossian, A.; Wikman, G.; Sarris, J. Rosenroot (Rhodiola rosea): Traditional use, chemical composition, pharmacology and clinical efficacy. Phytomedicine 2010, 17, 481–493. [Google Scholar] [CrossRef]
- Maneev, A.G.; Khmelyova, I.R. Some aspects of Rhodiola rosea in nature and culture. In Problems of studying the plants of Siberia; Gureeva, I.I., Ed.; TGU: Tomsk, Russia, 2015; pp. 318–329. [Google Scholar]
- Kim, E.F. Rhodiola rosea and Perspectives of It Introduction. Ph.D. Biology Thesis, Central Siberian Botanic Garden, Novosibirsk, Russia, 14 October 1999. [Google Scholar]
- Platikanov, S.; Evstatieva, L. Introduction of wild golden root (Rhodiola rosea L.) as a potential economic crop in Bulgaria. Econ. Bot. 2008, 62, 621–627. [Google Scholar] [CrossRef]
- Erst, A.A.; Yakubov, V.V. Regenerative in vitro capacity of rare species Rhodiola rosea L. from various habitats. Contemp. Probl. Ecol. 2019, 12, 368–376. [Google Scholar] [CrossRef]
- Zapesochnaya, G.G.; Kurkin, V.A.; Shchavlinskii, A.N. Flavonoids of the epigeal part of Rhodiola rosea. II. Structures of new glycosides of herbacetin and of gossypetin. Chem. Nat. Compd. 1985, 21, 464–473. [Google Scholar] [CrossRef]
- Kurkin, V.A.; Zapesochnaya, G.G.; Shchavlinskii, A.N. Flavonoids of the epigeal part of Rhodiola rosea. I. Chem. Nat. Compd. 1984, 20, 623–624. [Google Scholar] [CrossRef]
- Petsalo, A.; Jalonen, J.; Tolonen, A. Identification of flavonoids of Rhodiola rosea by liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2006, 1112, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Krasnov, E.A. Flavonoids of Rhodiola litvinovii. Chem. Nat. Compd. 1979, 15, 756. [Google Scholar] [CrossRef] [Green Version]
- Olennikov, D.N.; Chirikova, N.K. New flavonol glycosides from Rhodiola quadrifida. Chem. Nat. Compd. 2020, 55, 986–991. [Google Scholar]
- Krasnov, E.A.; Khoruzhaya, T.G.; Dranik, L.I.; Gordienko, V.G.; Kovalev, I.P. 6-O-Galloylarbutin from Rhodiola coccinea. Chem. Nat. Compd. 1976, 11, 492–494. [Google Scholar] [CrossRef]
- Khoruzhaya, T.G.; Krasnov, E.A. Phenolic compounds of Rhodiola coccinea. Chem. Nat. Compd. 1972, 8, 665. [Google Scholar] [CrossRef]
- Fan, W.; Tezuka, Y.; Ni, K.M.; Kadota, S. Prolyl endopeptidase inhibitors from the underground part of Rhodiola sachalinensis. Chem. Pharm. Bull. 2001, 49, 396–401. [Google Scholar] [CrossRef] [Green Version]
- Panossian, A.; Hovhannisyan, A.; Abrahamyan, H.; Gabrielyan, E.; Wickman, G. Pharmacokinetics of active constituents of Rhodiola rosea SHR-5 extract. In Comprehensive Bioactive Natural Products; Gupta, V.K., Ed.; Studium Press, LLC: Houston, TX, USA, 2010; Volume 2, pp. 307–329. [Google Scholar]
- Olennikov, D.N.; Kashchenko, N.I.; Akobirshoeva, A. Phenolic compounds and hydroxynitrile glycosides from roots of Rhodiola recticaulis and R. gelida. Chem. Nat. Compd. 2019, 55, 948–950. [Google Scholar] [CrossRef]
- Chirikova, N.K.; Olennikov, D.N.; Tankhaeva, L.M. Quantitative determination of flavonoid content in the aerial part of Baical scullcap (Scutellaria baicalensis Georgi). Russ. J. Bioorg. Chem. 2010, 36, 915–922. [Google Scholar] [CrossRef]
- Sun, B.; Ricardo-da-Silva, J.M.; Spranger, I. Critical factors of vanillin assay for catechins and proanthocyanidins. J. Agric. Food Chem. 1998, 46, 4267–4274. [Google Scholar] [CrossRef]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry 1986, 25, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Olennikov, D.N.; Tankhaeva, L.M. Quantitative determination of phenolic compounds in Mentha piperita leaves. Chem. Nat. Compd. 2010, 46, 22–27. [Google Scholar] [CrossRef]
- Inoue, K.H.; Hagerman, A.E. Determination of gallotannin with rhodanine. Anal. Biochem. 1988, 169, 363–369. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kruglova, M.Y. A new quercetin glycoside and other phenolic compounds from the genus Filipendula. Chem. Nat. Compd. 2013, 49, 610–616. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Fedorov, I.A.; Kashchenko, N.I.; Chirikova, N.K.; Vennos, C. Khellactone derivatives and other phenolics of Phlojodicarpus sibiricus (Apiaceae): HPLC-DAD-ESI-QQQ-MS/MS and HPLC-UV profile, and antiobesity potential of dihydrosamidin. Molecules 2019, 24, 2286. [Google Scholar] [CrossRef] [Green Version]
- Giusti, M.M.; Wrolstad, R.E. Characterization and measurement of anthocyanins by UV-visible spectroscopy. In Current Protocols in Food Analytical Chemistry; Reid, D.S., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2001; pp. F1.2.1–F1.2.13. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K. Meadowsweet teas as new functional beverages: Comparative analysis of nutrients, phytochemicals and biological effects of four Filipendula species. Molecules 2017, 22, 16. [Google Scholar] [CrossRef] [Green Version]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K.; Vasil’eva, A.G.; Gadimli, A.I.; Isaev, J.I.; Vennos, C. Caffeoylquinic acids and flavonoids of fringed sagewort (Artemisia frigida Willd.): HPLC-DAD-ESI-QQQ-MS profile, HPLC-DAD quantification, in vitro digestion stability, and antioxidant capacity. Antioxidants 2019, 8, 307. [Google Scholar] [CrossRef] [Green Version]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K. In vitro bioaccessibility, human gut microbiota metabolites and hepatoprotective potential of chebulic ellagitannins: A case of Padma Hepaten® formulation. Nutrients 2015, 7, 8456–8477. [Google Scholar] [CrossRef] [Green Version]
- Zomborszki, Z.P.; Kúsz, N.; Csupor, D.; Peschel, W. Rhodiosin and herbacetin in Rhodiola rosea preparations: Additional markers for quality control? Pharm. Biol. 2019, 57, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Staneva, J.; Todorova, M.; Neykov, N.; Evstatieva, L. Ultrasonically assisted extraction of total phenols and flavonoids from Rhodiola rosea. Nat. Prod. Commun. 2009, 4, 935–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peschel, W.; Prieto, J.M.; Karkour, C.; Williamson, E.M. Effect of provenance, plant part and processing on extract profiles from cultivated European Rhodiola rosea L. for medicinal use. Phytochemistry 2013, 86, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Marchev, A.S.; Aneva, I.Y.; Koycheva, I.K.; Georgiev, M.I. Phytochemical variations of Rhodiola rosea L. wild-grown in Bulgaria. Phytochem. Lett. 2017, 20, 386–390. [Google Scholar] [CrossRef]
- Gryszczyńska, A.; Krajewska-Patan, A.; Buchwald, W.; Czerny, B.; Mielcarek, S.; Rudzińska, K.; Mrozikiewicz, P.M. Comparison of proanthocyanidins content in Rhodiola kirilowii and Rhodiola rosea roots-application of UPLC-MS/MS method. Herba Pol. 2012, 58, 5–15. [Google Scholar]
- Pooja; Anilakumar, K.R.; Khanum, F.; Bawa, A.S. Phytoconstituents and antioxidant potency of Rhodiola rosea—A versatile adaptogen. J. Food Biochem. 2006, 30, 203–214. [Google Scholar] [CrossRef]
- Elameen, A.; Dragland, S.; Klemsdal, S.S. Bioactive compounds produced by clones of Rhodiola rosea maintained in the Norwerian germplasm collection. Pharmazie 2010, 65, 618–623. [Google Scholar] [CrossRef]
- Sun, B.; Leandro, C.; da Silva, J.M.R.; Spranger, J. Separation of grape and wine proanthocyanidins according to their degree of polymerization. J. Agric. Food Chem. 1998, 46, 1390–1396. [Google Scholar] [CrossRef]
- Kashchenko, N.I.; Chirikova, N.K.; Olennikov, D.N. Agrimoniin, an active ellagitannin from Comarum palustre herb with anti-α-glucosidase and antidiabetic potential in streptozotocin-induced diabetic rats. Molecules 2017, 22, 73. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K.; Koryakina, L.P.; Vladimirov, L.N. Bitter gentian teas: Nutritional and phytochemical profiles, polysaccharide characterisation and bioactivity. Molecules 2015, 20, 20014–20030. [Google Scholar] [CrossRef] [Green Version]
- Olennikov, D.N.; Chirikova, N.K.; Kashchenko, N.I.; Nikolaev, V.M.; Kim, S.-W.; Vennos, C. Bioactive phenolics of the genus Artemisia (Asteraceae): HPLC-DAD-ESI-TQ-MS/MS profile of the Siberian species and their inhibitory potential against α-amylase and α-glucosidase. Front. Pharmacol. 2018, 9, 756. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Gadimli, A.I.; Isaev, J.I.; Kashchenko, N.I.; Prokopyev, A.S.; Katayeva, T.N.; Chirikova, N.K.; Vennos, C. Caucasian Gentiana species: Untargeted LC-MS metabolic profiling, antioxidant and digestive enzyme inhibiting activity of six plants. Metabolites 2019, 9, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.H.; Hsu, C.C.; Hsiao, G.; Fang, J.Y.; Liu, W.M.; Lee, C.K. Anti-MMP-2 activity and skin-penetrating capability of the chemical constituents from Rhodiola rosea. Planta Med. 2016, 82, 698–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockenbach, I.I.; Jungfer, E.; Ritter, C.; Santiago-Schübel, B.; Thiele, B.; Fett, R.; Galensa, R. Characterization of flavan-3-ols in seeds of grape pomace by CE, HPLC-DAD-MSn and LC-ESI-FTICR-MS. Food Res. Int. 2012, 48, 848–855. [Google Scholar] [CrossRef] [Green Version]
- Troshchenko, A.T.; Kutikova, G.A. Rhodioloside from Rhodiola rosea and R. quadrifida I. Chem. Nat. Compd. 1967, 3, 204–207. [Google Scholar] [CrossRef]
- Furmanowa, M.; Skopińska-Rozewska, E.; Rogala, E.; Hartwich, M. Rhodiola rosea in vitro culture—Phytochemical analysis and antioxidant action. Acta Soc. Bot. Polon. 1998, 67, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Zapesochnaya, G.G.; Kurkin, V.A. Glycosides of cinnamyl alcohol from the rhizomes of Rhodiola rosea. Chem. Nat. Compd. 1982, 18, 685–688. [Google Scholar] [CrossRef]
- Kurkin, V.A.; Zapesochnaya, G.G.; Shchavlinskii, A.N. Terpenoids of the rhizomes of Rhodiola rosea. Chem. Nat. Compd. 1985, 21, 593–597. [Google Scholar] [CrossRef]
- Tolonen, A.; Hohtola, A.; Jalonen, J. Comparison of electrospray ionization and atmospheric pressure chemical ionization techniques in the analysis of the main constituents from Rhodiola rosea extracts by liquid chromatography/mass spectrometry. J. Mass Spectrom. 2003, 38, 845–853. [Google Scholar] [CrossRef]
- Hillhouse, B.J.; Ming, D.S.; French, C.J.; Towers, G.H.N. Acetylcholine esterase inhibitors in Rhodiola rosea. Pharm. Biol. 2004, 42, 68–72. [Google Scholar] [CrossRef]
- Ma, C.; Hu, L.; Fu, Q.; Gu, X.; Tao, G.; Wang, H. Separation of four flavonoids from Rhodiola rosea by on-line combination of sample preparation and counter-current chromatography. J. Chromatogr. A 2013, 1306, 12–19. [Google Scholar] [CrossRef]
- Zapesochnaya, G.G.; Kurkin, V.A. The flavonoids of the rhizomes of Rhodiola rosea. II. A flavonolignan and glycosides of herbacetin. Chem. Nat. Compd. 1983, 19, 21–29. [Google Scholar] [CrossRef]
- Kurkin, V.A.; Zapesochnaya, G.G.; Shchavlinskii, A.N. Flavonoids of the rhizomes of Rhodiola rosea. III. Chem. Nat. Compd. 1984, 20, 367–368. [Google Scholar] [CrossRef]
- Berardini, N.; Carle, R.; Schieber, A. Characterization of gallotannins and benzophenone derivatives from mango (Mangifera indica L. cv. ‘Tommy Atkins’) peels, pulp and kernels by high-performance liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 2208–2216. [Google Scholar] [CrossRef] [PubMed]
- Yu, W. Polyphenol constituents of Rhodiola crenulata. Nat. Prod. Res. Dev. 1992, 4, 26–31. [Google Scholar]
- Yousef, G.G.; Grace, M.H.; Cheng, D.M.; Belolipov, I.V.; Raskin, I.; Lila, M.A. Comparative phytochemical characterization of three Rhodiola species. Phytochemistry 2006, 67, 2380–2391. [Google Scholar] [CrossRef]
- Fan, W.; Tezuka, Y.; Komatsu, K.; Namba, T.; Kadota, S. Prolyl endopeptidase inhibitors from the underground part of Rhodiola sacra S. H. Fu. Biol. Pharm. Bull. 1999, 22, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Wiedenfeld, H.; Zych, M.; Buchwald, W.; Furmanow, M. New compounds from Rhodiola kirilowii. Sci. Pharm. 2007, 75, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Mudge, E.; Lopes-Lutz, D.; Brown, P.N.; Schieber, A. Purification of phenylalkanoids and monoterpene glycosides from Rhodiola rosea L. roots by high-speed counter-current chromatography. Phytochem Anal. 2013, 24, 129–134. [Google Scholar] [CrossRef]
- Avula, B.; Wang, Y.H.; Ali, Z.; Smillie, T.J.; Filion, V.; Cuerrier, A.; Arnason, J.T.; Khan, I.A. RP-HPLC determination of phenylalkanoids and monoterpenoids in Rhodiola rosea and identification by LC-ESI-TOF. Biomed. Chromatogr. 2009, 23, 865–872. [Google Scholar] [CrossRef]
- Ali, Z.; Fronczek, F.R.; Khan, I.A. Phenylalkanoids and monoterpene analogues from the roots of Rhodiola rosea. Planta Med. 2008, 74, 178–181. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, F. Über die Vergärung des Tyrosins zu p-Oxyphenyl-äthylalkohol (Tyrosol). Ber. Dtsch. Chem. Ges. 1911, 44, 139–146. [Google Scholar] [CrossRef]
- Marković, A.K.; Torić, J.; Barbarić, M.; Brala, C.J. Hydroxytyrosol, tyrosol and derivatives and their potential effects on human health. Molecules 2019, 24, 2001. [Google Scholar] [CrossRef] [Green Version]
- Wiedenfeld, H.; Dumaa, M.; Malinowski, M.; Furmanowa, M.; Narantuya, S. Phytochemical and analytical studies of extracts from Rhodiola rosea and Rhodiola quadrifida. Pharmazie 2007, 62, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Bjarnholt, N.; Rook, F.; Motawia, M.S.; Cornett, C.; Jørgensen, C.; Olsen, C.E.; Jaroszewski, J.W.; Bak, S.; Møller, B.L. Diversification of an ancient theme: Hydroxynitrile glucosides. Phytochemistry 2008, 69, 1507–1516. [Google Scholar] [CrossRef] [PubMed]
- Daikonya, A.; Kitanaka, S. Constituents isolated from the roots of Rhodiola sacra S. H. Fu. Japan J. Food Chem. Saf. 2011, 18, 183–190. [Google Scholar] [CrossRef]
- Kurkin, V.A.; Zapesochnaya, G.G.; Klyaznika, V.G. Flavonoids of the rhizomes of Rhodiola rosea. I. Tricin glucosides. Chem. Nat. Compd. 1982, 18, 550–552. [Google Scholar] [CrossRef]
- Stevens, J.F.; ‘t Hart, H.; Elema, E.T.; Bolck, A. Flavonoid variation in Eurasian Sedum and Sempervivum. Phytochemistry 1996, 41, 503–512. [Google Scholar] [CrossRef]
- Van Diermen, D. Phytochemical Investigation of Two Crassulaceae Species: Rhodiola rosea L., the New “herbal Stress Buster”, and Sedum dasyphyllum L. Thèse de Doctorat, University of Genève, Genève, Switzerland, 2009; 220p. [Google Scholar] [CrossRef]
- Van Diermen, D.; Pierreclos, M.; Hostettmann, K.; Cuendet, M. Cancer chemopreventive activity of constituents from Sedum dasyphyllum L. Planta Med. 2010, 76, P078. [Google Scholar] [CrossRef]
- Krasnov, E.A.; Demidenko, L.A. Flavone glycosides of Rhodiola krylovii. Chem. Nat. Compd. 1984, 20, 103–104. [Google Scholar] [CrossRef]
- Hussein, S.A.M.; Barakat, H.H.; Nawar, M.A.M.; Willuhn, G. Flavonoids from Ephedra aphylla. Phytochemistry 1997, 45, 1529–1532. [Google Scholar] [CrossRef]
- Pangarova, T.T.; Zapesochnaya, G.G. The structure of the flavonoids from Rhodiola algida. II. Chem. Nat. Compd. 1975, 11, 744–750. [Google Scholar] [CrossRef]
- Krasnov, E.A.; Demidenko, L.A. New flavonol glycosides of Rhodiola algida. Chem. Nat. Compd. 1979, 15, 353–354. [Google Scholar] [CrossRef]
- Adamczyk, B.; Simon, J.; Kitunen, V.; Adamczyk, S.; Smolander, A. Tannins and their complex interaction with different organic nitrogen compounds and enzymes: Old paradigms versus recent advances. ChemistryOpen 2017, 6, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Justesen, U.; Arrigoni, E.; Larsen, B.R.; Amado, R. Degradation of flavonoid glycosides and aglycones during in vitro fermentation with human faecal flora. LWT–Food Sci. Technol. 2000, 33, 424–430. [Google Scholar] [CrossRef]
- Guo, N.; Zhu, M.; Han, X.; Sui, D.; Wang, Y.; Yang, Q. The metabolism of salidroside to its aglycone p-tyrosol in rats following the administration of salidroside. PLoS ONE 2014, 9, e103648. [Google Scholar] [CrossRef] [PubMed]
- Schindler, G.; Patzak, U.; Brinkhaus, B.; von Niecieck, A.; Wittig, J.; Krähmer, N.; Glöckl, I.; Veit, M. Urinary excretion and metabolism of arbutin after oral administration of Arctostaphylos uvae ursi extract as film-coated tablets and aqueous solution in healthy humans. J. Clin. Pharmacol. 2002, 42, 920–927. [Google Scholar] [CrossRef]
- Krook, M.A.; Hagerman, A.E. Stability of polyphenols epigallocatechin gallate and pentagalloyl glucose in a simulated digestive system. Food Res. Int. 2012, 49, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Quatrin, A.; Rampelotto, C.; Pauletto, R.; Maurer, L.H.; Nichelle, S.M.; Klein, B.; Rodrigues, R.F.; Maróstica Junior, M.R.; Fonseca, B.D.S.; de Menezes, C.R.; et al. Bioaccessibility and catabolism of phenolic compounds from jaboticaba (Myrciaria trunciflora) fruit peel during in vitro gastrointestinal digestion and colonic fermentation. J. Funct. Foods 2020, 65, 103714. [Google Scholar] [CrossRef]
- Neilson, A.P.; Hopf, A.S.; Cooper, B.R.; Pereira, M.A.; Bomser, J.A.; Ferruzzi, M.J. Catechin degradation with concurrent formation of homo- and heterocatechin dimers during in vitro digestion. J. Agric. Food Chem. 2007, 55, 8941–8949. [Google Scholar] [CrossRef]
- Soares, S.; Brandao, E.; Mateus, N.; de Freitas, V. Interaction between red wine procyanidins and salivary proteins: Effect of stomach digestion on the resulting complexes. RSC Adv. 2015, 5, 12664. [Google Scholar] [CrossRef]
- Fernández, K.; Labra, J. Simulated digestion of proanthocyanidins in grape skin and seed extracts and the effects of digestion on the angiotensin I-converting enzyme (ACE) inhibitory activity. Food Chem. 2013, 139, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Rha, C.-S.; Seong, H.; Jung, Y.S.; Jang, D.; Kwak, J.-G.; Kim, D.-O.; Han, N.S. Stability and fermentability of green tea flavonols in in-vitro-simulated gastrointestinal digestion and human fecal fermentation. Int. J. Mol. Sci. 2019, 20, 5890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, L.M.L.; Barlow, P.J. Flavonoid recovery and stability from Ginkgo biloba subjected to a simulated digestion process. Food Chem. 2004, 86, 195–202. [Google Scholar] [CrossRef]
- Hanske, L.; Loh, G.; Sczesny, S.; Blaut, M.; Braune, A. The bioavailability of apigenin-7-glucoside is influenced by human intestinal microbiota in rats. J. Nutr. 2009, 139, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Che, Q.M.; Huang, X.L.; Li, Y.M.; Zhang, K.; Akao, T.; Hattori, M. Studies on metabolites of baicalin in human urine. China J. Chin. Mater. Med. 2001, 26, 768–769. [Google Scholar]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Papers | Year | Papers | Year | Papers | Year | Papers | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| R | H | R | H | R | H | R | H | ||||
| 1966 | 1 | 0 | 1987 | 2 | 0 | 1999 | 2 | 0 | 2011 | 26 | 0 |
| 1967 | 1 | 0 | 1988 | 1 | 0 | 2000 | 7 | 0 | 2012 | 31 | 0 |
| 1968 | 1 | 0 | 1989 | 3 | 0 | 2001 | 2 | 0 | 2013 | 53 | 0 |
| 1973 | 1 | 0 | 1990 | 1 | 0 | 2002 | 7 | 0 | 2014 | 40 | 0 |
| 1977 | 1 | 0 | 1991 | 7 | 0 | 2003 | 13 | 0 | 2015 | 42 | 0 |
| 1980 | 1 | 0 | 1992 | 1 | 0 | 2004 | 11 | 1 | 2016 | 50 | 0 |
| 1981 | 3 | 0 | 1993 | 3 | 0 | 2005 | 11 | 0 | 2017 | 35 | 0 |
| 1982 | 1 | 0 | 1994 | 1 | 0 | 2006 | 18 | 0 | 2018 | 47 | 0 |
| 1983 | 2 | 0 | 1995 | 1 | 0 | 2007 | 23 | 0 | 2019 | 69 | 0 |
| 1984 | 1 | 1 | 1996 | 2 | 0 | 2008 | 25 | 0 | Total | 626 | 3 |
| 1985 | 1 | 1 | 1997 | 7 | 0 | 2009 | 40 | 0 | |||
| 1986 | 2 | 0 | 1998 | 3 | 0 | 2010 | 26 | 0 | |||
| Organ | Collection Place | Collection Date | Coordinates | Height (m a.s.l.) | Voucher Specimens No |
|---|---|---|---|---|---|
| Herbal: leaves, flowers, stems | Chulman, Aldanskii Ulus, Sakha (Yakutia) Republic | 25.VII.2019 | 57°00′37″ N, 124°49′02″ E | 960 | YA/CRA-0719/22-106 |
| Subterranean: roots, rhizomes | Chulman, Aldanskii Ulus, Sakha (Yakutia) Republic | 02.IX.2019 | 57°00′37″ N, 124°49′02″ E | 960 | YA/CRA-0919/38-471 |
| Organ | Flavonoids | Catechins | Procyanidins | Phenylpropanoids | Gallotannins | Total Phenolics |
|---|---|---|---|---|---|---|
| Roots | 1.89 ± 0.03 | 10.84 ± 0.80 | 9.21 ± 0.39 | 21.89 ± 0.43 | 1.04 ± 0.02 | 44.87 |
| Rhizomes | 0.75 ± 0.02 | 61.30 ± 1.22 | 31.37 ± 0.61 | 46.45 ± 0.92 | 0.73 ± 0.02 | 140.60 |
| Flowers | 46.36 ± 0.95 | 0.66 ± 0.02 | 1.52 ± 0.11 | 1.14 ± 0.02 | 63.11 ± 1.26 | 112.79 |
| Leaves | 16.71 ± 0.31 | 0.30 ± 0.01 | 0.17 ± 0.00 | 8.27 ± 0.16 | 97.53 ± 2.02 | 122.98 |
| Stems | 2.96 ± 0.05 | 0.32 ± 0.01 | 1.10 ± 0.02 | 5.11 ± 0.10 | 12.35 ± 0.25 | 21.84 |
| Extracts | DPPH• a,b | ABTS•+ b,c | O2•− b,d | •OH b,e | CBA b,f |
|---|---|---|---|---|---|
| Roots | 11.71 ± 0.23 ** | 5.37 ± 0.11 ** | 45.11 ± 0.90 ** | 37.58 ± 0.75 ** | 87.50 ± 2.62 ** |
| Rhizomes | 2.96 ± 0.06 ** | 0.62 ± 0.01 ** | 8.63 ± 0.17 ** | 6.02 ± 0.11 ** | 44.79 ± 1.41 ** |
| Flowers | 3.95 ± 0.08 ** | 0.98 ± 0.02 ** | 23.19 ± 0.45 ** | 12.83 ± 0.25 * | 25.06 ± 0.63 * |
| Leaves | 2.91 ± 0.06 ** | 0.53 ± 0.01 ** | 10.38 ± 0.21 ** | 7.39 ± 0.14 ** | 11.86 ± 0.23 * |
| Stems | 8.20 ± 0.16 | 2.98 ± 0.05 * | 37.69 ± 0.75 ** | 21.18 ± 0.42 ** | 37.15 ± 1.11 ** |
| Trolox g | 8.38 ± 0.17 | 3.18 ± 0.06 | 125.11 ± 2.50 | 14.06 ± 0.28 | 21.05 ± 0.62 |
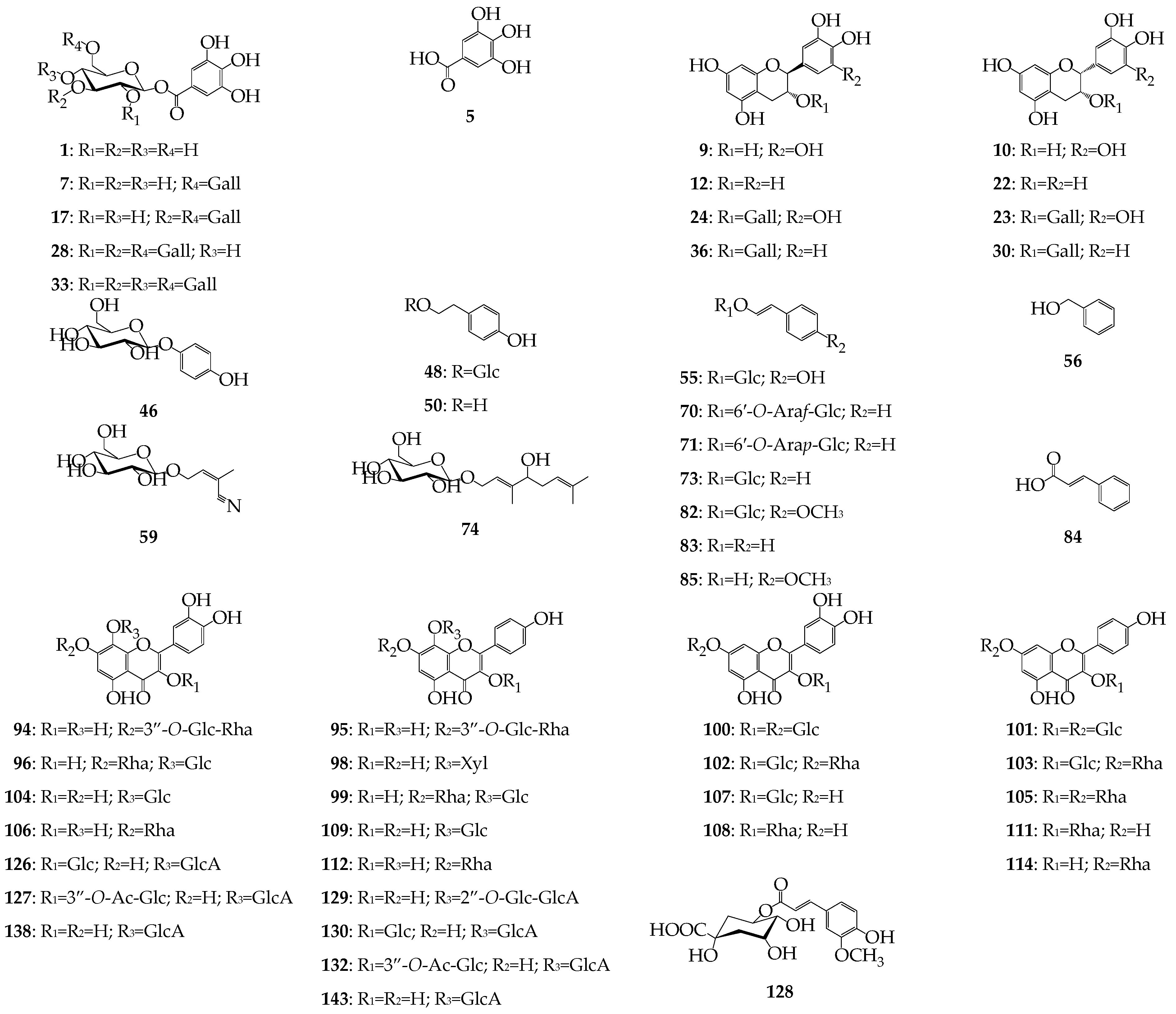
| No. | tR, min a | ESI-MS, m/z b | Group c | Compound d | Presence (+) in Organs, mg/g ± S.D. e | ||||
|---|---|---|---|---|---|---|---|---|---|
| Roots | Rhizome | Leaves | Flowers | Stems | |||||
| 1 | 1.91 i | 331 N | GT | 1-O-Galloyl glucose S | + | + | + | + | + |
| 2 | 2.18 i | 331 N | GT | O-Galloyl glucose L | + | + | + | + | + |
| 3 | 2.33 i | 331 N | GT | O-Galloyl glucose L | + | + | |||
| 4 | 2.57 i | 331 N | GT | O-Galloyl glucose L | + | + | |||
| 5 | 2.94 i | 169 N | GT | Gallic acid S | [46] | [46] | + | + | + |
| 6 | 4.61 i | 483 N | GT | Di-O-galloyl glucose L | + | ||||
| 7 | 4.78 i | 483 N | GT | 1,6-Di-O-galloyl glucose S | + | + | + | + | + |
| 8 | 5.11 i | 609 N | PC | Procyanidin dimer (EGC-EGC) L | + | + | + | + | |
| 9 | 5.27 i | 305 N | CT | Gallocatechin S | + | + | + | ||
| 10 | 5.52 i | 305 N | CT | Epigallocatechin S | [38] | + | |||
| 11 | 5.76 i | 761 N | PC | Procyanidin dimer (EGC-EGCG) L | + | [47] | + | ||
| 12 | 5.85 i | 289 N | CT | Catechin S | [38] | + | |||
| 13 | 6.27 i | 635 N | GT | Tri-O-galloyl glucose L | + | + | |||
| 14 | 6.31 i | 913 N | PC | Procyanidin dimer (EGCG-EGCG) L | + | [47] | |||
| 15 | 6.35 i | 913 N | PC | Procyanidin trimer (EGC-EGC-EGC) L | + | + | |||
| 16 | 6.48 i | 1065 N | PC | Procyanidin trimer (EGC-EGC-EGCG) L | + | [47] | |||
| 17 | 6.54 i | 635 N | GT | 1,3,6-Tri-O-galloyl glucose S | + | + | |||
| 18 | 6.78 i | 635 N | GT | Tri-O-galloyl glucose L | + | + | + | ||
| 19 | 6.82 i | 1217 N | PC | Procyanidin trimer (EGC-EGCG-EGCG) L | + | [47] | |||
| 20 | 6.89 i | 1369 N | PC | Procyanidin trimer (EGCG-EGCG-EGCG) L | + | [47] | |||
| 21 | 6.92 i | 1369 N | PC | Procyanidin tetramer (EGC-EGC-EGC-EGCG) L | + | + | |||
| 22 | 7.02 i | 289 N | CT | Epicatechin S | [38] | + | |||
| 23 | 7.12 i | 457 N | CT | Epigallocatechin gallate S | [38] | + | |||
| 24 | 7.22 i | 457 N | CT | Gallocatechin gallate S | + | + | |||
| 25 | 7.41 i | 1521 N | PC | Procyanidin tetramer (EGC-EGC-EGCG-EGCG) L | + | [47] | |||
| 26 | 7.53 i | 1673 N | PC | Procyanidin tetramer (EGC-EGCG-EGCG-EGCG) L | + | [47] | |||
| 27 | 7.55 i | 787 N | GT | Tetra-O-galloyl glucose L | + | ||||
| 28 | 7.63 i | 787 N | GT | 1,2,3,6-Tetra-O-galloyl glucose S | + | + | + | ||
| 29 | 7.83 i | 787 N | GT | Tetra-O-galloyl glucose L | + | ||||
| 30 | 8.07 i | 441 N | CT | Epicatechin gallate S | [38] | + | |||
| 31 | 8.14 i | 787 N | GT | Tetra-O-galloyl glucose L | + | ||||
| 32 | 8.26 i | 939 N | GT | Penta-O-galloyl glucose L | + | + | + | ||
| 33 | 8.42 i | 939 N | GT | 1,2,3,4,6-Penta-O-galloyl glucose S | + | + | + | ||
| 34 | 8.63 i | 1091 N | GT | Hexa-O-galloyl glucose L | + | + | + | ||
| 35 | 8.72 i | 1091 N | GT | Hexa-O-galloyl glucose L | + | + | + | ||
| 36 | 8.92 i | 441 N | CT | Catechin gallate S | + | + | |||
| 37 | 8.98 i | 1243 N | GT | Hepta-O-galloyl glucose L | + | + | + | ||
| 38 | 9.14 i | 1243 N | GT | Hepta-O-galloyl glucose L | + | + | + | ||
| 39 | 9.63 i | 1395 N | GT | Octa-O-galloyl glucose L | + | ||||
| 40 | 9.87 i | 1395 N | GT | Octa-O-galloyl glucose L | + | + | |||
| 41 | 10.04 i | 1825 N | PC | Procyanidin tetramer (EGCG-EGCG-EGCG-EGCG) L | + | [47] | + | + | |
| 42 | 10.41 i | 1673 N | PC | Procyanidin pentamer (EGC-EGC-EGC-EGC-EGCG) L | + | + | + | + | |
| 43 | 11.18 i | 1825 N | PC | Procyanidin pentamer (EGC-EGC-EGC-EGCG-EGCG) L | + | + | + | ||
| 44 | 11.43 i | 1977 N | PC | Procyanidin pentamer (EGC-EGC-EGCG-EGCG-EGCG) L | + | [47] | + | ||
| 45 | 1.90 ii | 403 N | PG | Hydroquinone O-Hex-O-Pent L | + | + | + | + | + |
| 46 | 1.98 ii | 271 N | PG | Hydroquinone O-Glc (=arbutin) S | + | + | + | + | + |
| 47 | 2.09 ii | 431 N | PE | Tyrosol O-Hex-O-Pent L | + | + | + | + | + |
| 48 | 2.37 ii | 299 N | PE | Tyrosol O-Glc (=salidroside) S | [48] | [48] | + | + | + |
| 49 | 4.59 ii | 283 N | PE | Tyrosol O-dHex L | + | + | + | ||
| 50 | 4.95 ii | 137 N | PE | Tyrosol (=p-hydroxyphenethyl alcohol) S | [48] | [48] | + | + | + |
| 51 | 5.11 ii | 401 N | PG | Benzyl alcohol O-Hex-O-Pent L | + | + | + | + | + |
| 52 | 5.23 ii | 401 N | PG | Benzyl alcohol O-Hex-O-Pent L | + | + | + | + | + |
| 53 | 5.56 ii | 269 N | PG | Benzyl alcohol O-Hex L | + | + | + | + | + |
| 54 | 5.82 ii | 443 N | HC | p-Hydroxycinnamyl alcohol O-Hex-O-Pent L | + | + | |||
| 55 | 6.02 ii | 311 N | HC | p-Hydroxycinnamyl alcohol O-Glc (=triandrin) S | [49] | [49] | + | + | + |
| 56 | 6.27 ii | 107 N | PG | Benzyl alcohol S | + | + | + | + | + |
| 57 | 6.55 ii | 390 N | HNG | Rhodiocyanoside A/D O-Pent L | + | + | |||
| 58 | 6.76 ii | 258 N | HNG | Rhodiocyanoside D L | + | + | + | + | + |
| 59 | 6.97 ii | 258 N | HNG | Rhodiocyanoside A S | + | + | + | + | + |
| 60 | 7.18 ii | 625 N | TG | Rosiridol di-O-Hex-O-Pent L | + | + | + | ||
| 61 | 7.27 ii | 595 N | TG | Rosiridol O-Hex-di-O-Pent L | + | + | + | ||
| 62 | 7.47 ii | 595 N | TG | Rosiridol O-Hex-di-O-Pent L | |||||
| 63 | 7.52 ii | 463 N | TG | Rosiridol O-Hex-O-Pent L | + | + | + | + | + |
| 64 | 7.81 ii | 463 N | TG | Rosiridol O-Hex-O-Pent L | + | + | + | + | + |
| 65 | 8.18 ii | 341 N | PE | Tyrosol O-Hex-O-Ac L | + | + | + | + | + |
| 66 | 8.72 ii | 341 N | PE | Tyrosol O-Hex-O-Ac L | + | + | + | ||
| 67 | 8.98 ii | 383 N | PE | Tyrosol O-Hex-di-O-Ac L | + | + | + | ||
| 68 | 9.02 ii | 559 N | HC | Cinnamyl alcohol O-Hex-di-O-Pent L | + | + | |||
| 69 | 9.26 ii | 559 N | HC | Cinnamyl alcohol O-Hex-di-O-Pent L | + | + | |||
| 70 | 9.57 ii | 427 N | HC | Cinnamyl alcohol O-(6′-O-Araf)-Glc (=rosarin) S | [50] | [50] | |||
| 71 | 9.95 ii | 427 N | HC | Cinnamyl alcohol O-(6′-O-Arap)-Glc (=rosavin) S | [50] | [50] | |||
| 72 | 10.26 ii | 427 N | HC | Cinnamyl alcohol O-Hex-O-Pent L | + | + | |||
| 73 | 10.51 ii | 295 N | HC | Cinnamyl alcohol O-Glc (=rosin) S | [50] | [50] | |||
| 74 | 10.98 ii | 331 N | TG | Rosiridol 1-O-Glc (=rosiridin) S | [51] | [51] | + | + | + |
| 75 | 11.27 ii | 353 N | HC | p-Hydroxycinnamyl alcohol O-Hex-O-Ac L | + | + | + | ||
| 76 | 11.67 ii | 353 N | HC | p-Hydroxycinnamyl alcohol O-Hex O-Ac L | + | + | + | ||
| 77 | 12.14 ii | 373 N | TG | Rosiridol O-Hex-O-Ac L | + | + | + | ||
| 78 | 12.48 ii | 415 N | TG | Rosiridol O-Hex-di-O-Ac L | + | + | + | ||
| 79 | 12.81 ii | 415 N | TG | Rosiridol O-Hex-di-O-Ac L | + | + | + | ||
| 80 | 13.65 ii | 457 N | HC | p-Methoxycinnamyl alcohol O-Hex-O-Pent L | + | + | |||
| 81 | 13.94 ii | 457 N | HC | p-Methoxycinnamyl alcohol O-Hex-O-Pent L | + | + | |||
| 82 | 14.57 ii | 325 N | HC | p-Methoxycinnamyl alcohol O-Glc (=vimalin) S | [52] | [52] | |||
| 83 | 15.06 ii | 133 N | HC | Cinnamyl alcohol S | [50] | [50] | |||
| 84 | 15.47 ii | 147 N | HC | Cinnamic acid S | + | + | |||
| 85 | 16.58 ii | 163 N | HC | p-Methoxycinnamyl alcohol S | + | + | |||
| 86 | 5.02 iii | 919 P | NFG | Kaempferol tri-O-Hex-O-dHex (S37) L | + | ||||
| 87 | 5.41 iii | 935 P | NFG | Herbacetin tri-O-Hex-O-dHex (S78) L | + | + | |||
| 88 | 5.56 iii | 789 P | NFG | Gossypetin di-O-Hex-O-dHex (S78) L | + | + | |||
| 89 | 5.63 iii | 773 P | NFG | Quercetin di-O-Hex-O-dHex (S37) L | + | + | |||
| 90 | 5.97 iii | 757 P | NFG | Kaempferol di-O-Hex-O-dHex (S37) L | + | + | + | + | |
| 91 | 6.18 iii | 643 P | NFG | Gossypetin di-O-Hex (S8) L | + | ||||
| 92 | 6.21 iii | 773 P | NFG | Herbacetin di-O-Hex-O-dHex (S78) L | + | + | |||
| 93 | 6.32 iii | 643 P | NFG | Gossypetin di-O-Hex (S38) L | + | ||||
| 94 | 6.43 iii | 627 P | NFG | Gossypetin 7-O-(3″-O-Glc)-Rha (=rhodioflavonoside) S | [53] | [53] | + | + | |
| 95 | 6.63 iii | 611 P | NFG | Herbacetin 7-O-(3″-O-Glc)-Rha (=rhodiosin) S | + | + | |||
| 96 | 6.82 iii | 627 P | NFG | Gossypetin 7-O-Rha-8-O-Glc (=rhodiolgidin) S | [13] | [13] | + | ||
| 97 | 7.02 iii | 595 P | NFG | Kaempferol O-Hex-O-dHex (S7) L | + | + | |||
| 98 | 7.21 iii | 435 P | NFG | Herbacetin-8-O-Xyl (=rhodalin) S | + | + | [14] | [14] | |
| 99 | 7.35 iii | 611 P | NFG | Herbacetin 7-O-Rha-8-O-Glc (=rhodionidin) S | + | + | [14] | [14] | + |
| 100 | 7.43 iii | 627 P | NFG | Quercetin 3,7-di-O-Glc S | + | ||||
| 101 | 7.54 iii | 611 P | NFG | Kaempferol 3,7-di-O-Glc S | + | + | [15] | ||
| 102 | 7.75 iii | 611 P | NFG | Quercetin 3-O-Glc-7-O-Rha S | + | + | |||
| 103 | 7.97 iii | 595 P | NFG | Kaempferol 3-O-Glc-7-O-Rha S | [54] | [54] | + | + | |
| 104 | 8.01 iii | 481 P | NFG | Gossypetin 8-O-Glc (=gossypin) S | + | ||||
| 105 | 8.11 iii | 579 P | NFG | Kaempferol 3,7-di-O-Rha (=kaempferitrin) S | + | ||||
| 106 | 8.41 iii | 465 P | NFG | Gossypetin 7-O-Rha (=rhodiolgin) S | [13] | ||||
| 107 | 8.57 iii | 465 P | NFG | Quercetin 3-O-Glc (=isoquercitrin) S | + | + | + | + | |
| 108 | 8.98 iii | 449 P | NFG | Quercetin 3-O-Rha (=quercitrin) S | [2] | [2] | + | ||
| 109 | 9.22 iii | 465 P | NFG | Herbacetin 8-O-Glc (=herbacin) S | [54] | [54] | + | + | |
| 110 | 9.43 iii | 449 P | NFG | Herbacetin O-dHex (S8) L | + | + | + | ||
| 111 | 10.12 iii | 433 P | NFG | Kaempferol 3-O-Rha (=afzelin) S | + | + | + | ||
| 112 | 10.46 iii | 449 P | NFG | Herbacetin 7-O-Rha (=rhodionin) S | [55] | [55] | + | ||
| 113 | 11.27 iii | 449 P | NFG | Herbacetin O-dHex (S3) L | + | ||||
| 114 | 11.45 iii | 433 P | NFG | Kaempferol 7-O-Rha S | [56] | [56] | + | ||
| 115 | 1.97 iv | 835 P | AFG | Hibiscetin di-O-Hex-O-HexA (S38) L | + | + | |||
| 116 | 2.58 iv | 819 P | AFG | Gossypetin di-O-Hex-O-HexA (S378) L | + | + | |||
| 117 | 2.82 iv | 861 P | AFG | Gossypetin O-Ac-di-O-Hex-O-HexA (S378) L | + | + | |||
| 118 | 3.15 iv | 861 P | AFG | Gossypetin O-Ac-di-O-Hex-O-HexA (S378) L | + | + | |||
| 119 | 4.42 iv | 803 P | AFG | Herbacetin di-O-Hex-O-HexA (S38) L | + | + | |||
| 120 | 4.63 iv | 803 P | AFG | Herbacetin di-O-Hex-O-HexA (S378) L | + | + | + | + | |
| 121 | 5.02 iv | 889 P | AFG | Herbacetin O-Mal-di-O-Hex-O-HexA (S378) L | + | + | + | + | + |
| 122 | 5.27 iv | 845 P | AFG | Herbacetin O-Ac-di-O-Hex-O-HexA (S378) L | + | + | |||
| 123 | 5.52 iv | 673 P | AFG | Hibiscetin O-Hex-O-HexA (S38) L | + | + | + | + | + |
| 124 | 6.03 iv | 759 P | AFG | Hibiscetin O-Mal-O-Hex-O-HexA (S38) L | + | + | + | ||
| 125 | 6.18 iv | 715 P | AFG | Hibiscetin O-Ac-O-Hex-O-HexA (S38) L | + | + | |||
| 126 | 6.53 iv | 657 P | AFG | Gossypetin 3-O-Glc-8-O-GlcA S | + | + | + | + | + |
| 127 | 6.97 iv | 699 P | AFG | Gossypetin 3-O-(3″-O-Ac)-Glc-8-O-GlcA (=rhodiquadrin B) S | + | + | + | + | + |
| 128 | 7.27 iv | 367 N | HC | 3-O-Feruloylquinic acid S | + | + | + | + | + |
| 129 | 7.51 iv | 641 P | AFG | Herbacetin 8-O-(2″-O-Glc)-GlcA (=rhodiquadrin C) S | + | + | + | + | + |
| 130 | 7.62 iv | 641 P | AFG | Herbacetin 3-O-Glc-8-O-GlcA S | + | + | + | + | + |
| 131 | 7.90 iv | 683 P | AFG | Herbacetin O-Ac-O-Hex-O-HexA (S3,8) L | + | ||||
| 132 | 8.23 iv | 683 P | AFG | Herbacetin 3-O-(3″-O-Ac)-Glc-8-O-GlcA S | + | + | + | + | + |
| 133 | 8.48 iv | 741 P | AFG | Gossypetin di-O-Ac-O-Hex-O-HexA (S3,8) L | + | + | + | + | |
| 134 | 8.73 iv | 741 P | AFG | Gossypetin di-O-Ac-O-Hex-O-HexA (S3,8) L | + | + | + | + | + |
| 135 | 8.98 iv | 683 P | AFG | Herbacetin O-Ac-O-Hex-O-HexA (S3,8) L | + | + | + | + | + |
| 136 | 9.49 iv | 683 P | AFG | Herbacetin O-Ac-O-Hex-O-HexA (S3,8) L | + | + | + | + | + |
| 137 | 10.47 iv | 725 P | AFG | Herbacetin di-O-Ac-O-Hex-O-HexA (S3,8) L | + | + | + | + | + |
| 138 | 11.25 iv | 495 P | AFG | Gossypetin 8-O-GlcA (=hibifolin) S | + | + | |||
| 139 | 11.59 iv | 581 P | AFG | Gossypetin O-Mal-O-HexA (S8) L | + | + | |||
| 140 | 11.98 iv | 581 P | AFG | Gossypetin O-Mal-O-HexA (S8) L | + | + | + | ||
| 141 | 12.26 iv | 537 P | AFG | Gossypetin O-Ac-O-HexA (S8) L | + | + | + | + | |
| 142 | 12.61 iv | 537 P | AFG | Gossypetin O-Ac-O-HexA (S8) L | + | + | |||
| 143 | 14.01 iv | 479 P | AFG | Herbacetin 8-O-GlcA (=melocorin) S | + | + | + | + | + |
| 144 | 14.50 iv | 565 P | AFG | Herbacetin O-Mal-O-HexA (S8) L | + | + | + | + | |
| 145 | 15.37 iv | 521 P | AFG | Herbacetin O-Ac-O-HexA (S8) L | + | + | + | ||
| 146 | 16.11 iv | 521 P | AFG | Herbacetin O-Ac-O-HexA (S8) L | + | ||||
| Total compounds found | 90 | 90 | 87 | 100 | 69 | ||||
| Total known compounds found | 21 | 25 | 3 | 5 | 0 | ||||
| Total compounds found for the first time | 69 | 65 | 84 | 95 | 69 | ||||
| Compound | Roots | Rhizome | Leaves | Flowers | Stems |
|---|---|---|---|---|---|
| Total galloyl glucoses: | 5.45 | 3.08 | 100.63 | 68.19 | 10.98 |
| incl. mono-galloyl glucoses | 0.61 | 0.30 | 1.05 | 0.48 | 0.60 |
| di-galloyl glucoses | 2.29 | 1.17 | 10.35 | 1.56 | 3.89 |
| tri-galloyl glucoses | 0.83 | 0.42 | 1.95 | 0.00 | 1.04 |
| tetra-galloyl glucoses | 0.00 | 0.00 | 4.77 | 7.99 | 1.42 |
| penta-galloyl glucoses | 0.00 | 0.00 | 25.60 | 12.21 | 0.94 |
| hexa-galloyl glucoses | 0.00 | 0.00 | 37.79 | 24.02 | 2.22 |
| hepta-galloyl glucoses | 0.00 | 0.00 | 17.45 | 19.10 | 0.51 |
| octa-galloyl glucoses | 0.00 | 0.00 | 0.00 | 1.31 | 0.09 |
| Total catechins | 10.58 | 57.40 | 0.00 | 0.00 | 0.12 |
| Total procyanidins: | 8.77 | 34.81 | 0.00 | 3.02 | 0.23 |
| incl. dimers | 3.50 | 15.48 | 0.00 | 0.19 | 0.15 |
| trimers | 3.47 | 12.29 | 0.00 | 0.00 | 0.00 |
| tetramers | 0.24 | 1.14 | 0.00 | 1.10 | 0.00 |
| pentamers | 1.56 | 5.90 | 0.00 | 1.73 | 0.08 |
| Hydroquinone glucosides | 4.64 | 8.87 | 16.62 | 24.04 | 2.67 |
| p-Hydroxyphenethyl alcohol glucosides | 1.97 | 20.12 | 0.62 | 1.87 | traces |
| Hydroxynitrile glucosides | 0.97 | 2.64 | 1.42 | 2.66 | 0.58 |
| Rosiridol glucosides | traces | 5.18 | 0.53 | 6.45 | traces |
| Cinnamyl alcohols glucosides: | 19.89 | 55.76 | 0.58 | 1.26 | 0.39 |
| incl. p-hydroxycinnamyl alcohol glucosides | traces | 6.30 | 0.58 | 1.26 | 0.39 |
| cinnamyl alcohol glucosides | 19.89 | 47.42 | 0.00 | 0.00 | 0.00 |
| p-methoxycinnamyl alcohol glucosides | traces | 2.04 | 0.00 | 0.00 | 0.00 |
| Total flavonol glucosides: | 1.69 | 0.50 | 26.85 | 55.55 | 6.30 |
| incl. neutral glucoside | 0.56 | 0.08 | traces | 42.88 | 1.74 |
| acidic glucoside | 1.13 | 0.42 | 26.85 | 12.67 | 4.56 |
| or incl. herbacetin glucosides | 0.95 | 0.33 | 23.99 | 33.74 | 5.51 |
| gossypetin glucosides | 0.28 | 0.11 | 1.97 | 20.60 | 0.72 |
| hibiscetin glucosides | 0.00 | 0.00 | 0.89 | 0.92 | 0.07 |
| kaempferol glucosides | 0.46 | 0.06 | traces | 0.21 | traces |
| quercetin glucosides | traces | traces | traces | 0.08 | traces |
| Compounds | Rhizomes Extract | Leaves Extract | Flowers Extract | |||
|---|---|---|---|---|---|---|
| After Gastric Phase | After Intestinal Phase | After Gastric Phase | After Intestinal Phase | After Gastric Phase | After Intestinal Phase | |
| Galloyl glucoses | n.d. | n.d. | 65.15 | 90.20 | 74.07 | 94.17 |
| Procyanidins | 29.11 | 94.49 | n.d. | n.d. | n.d. | n.d. |
| Catechins | 39.63 | 96.74 | n.d. | n.d. | n.d. | n.d. |
| Hydroquinone glucosides | 7.30 | 23.71 | 9.03 | 28.02 | 6.03 | 24.02 |
| p-Hydroxyphenethyl alcohol glucosides | 3.07 | 12.52 | n.d. | n.d. | n.d. | n.d. |
| Cinnamyl alcohol glucosides | 0.07 | 1.15 | n.d. | n.d. | n.d. | n.d. |
| Flavonol glucosides | n.d. | n.d. | 0.44 | 0.92 | 1.39 | 2.04 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olennikov, D.N.; Chirikova, N.K.; Vasilieva, A.G.; Fedorov, I.A. LC-MS Profile, Gastrointestinal and Gut Microbiota Stability and Antioxidant Activity of Rhodiola rosea Herb Metabolites: A Comparative Study with Subterranean Organs. Antioxidants 2020, 9, 526. https://doi.org/10.3390/antiox9060526
Olennikov DN, Chirikova NK, Vasilieva AG, Fedorov IA. LC-MS Profile, Gastrointestinal and Gut Microbiota Stability and Antioxidant Activity of Rhodiola rosea Herb Metabolites: A Comparative Study with Subterranean Organs. Antioxidants. 2020; 9(6):526. https://doi.org/10.3390/antiox9060526
Chicago/Turabian StyleOlennikov, Daniil N., Nadezhda K. Chirikova, Aina G. Vasilieva, and Innokentii A. Fedorov. 2020. "LC-MS Profile, Gastrointestinal and Gut Microbiota Stability and Antioxidant Activity of Rhodiola rosea Herb Metabolites: A Comparative Study with Subterranean Organs" Antioxidants 9, no. 6: 526. https://doi.org/10.3390/antiox9060526