
The 4-Hydroxynonenal–Protein Adducts and Their Biological Relevance: Are Some Proteins Preferred Targets?
 , ,
, ,
Abstract
:1. Introduction
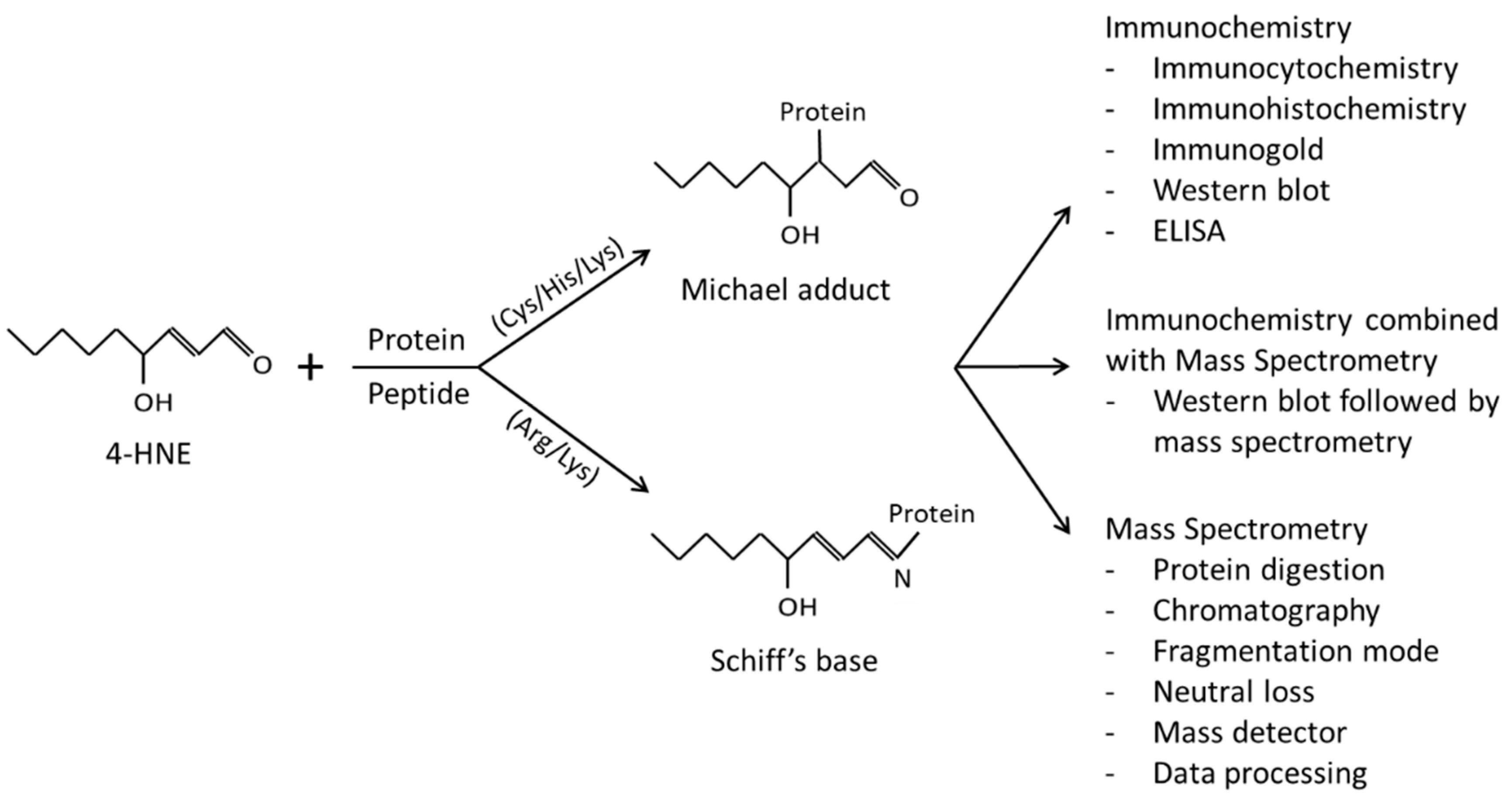
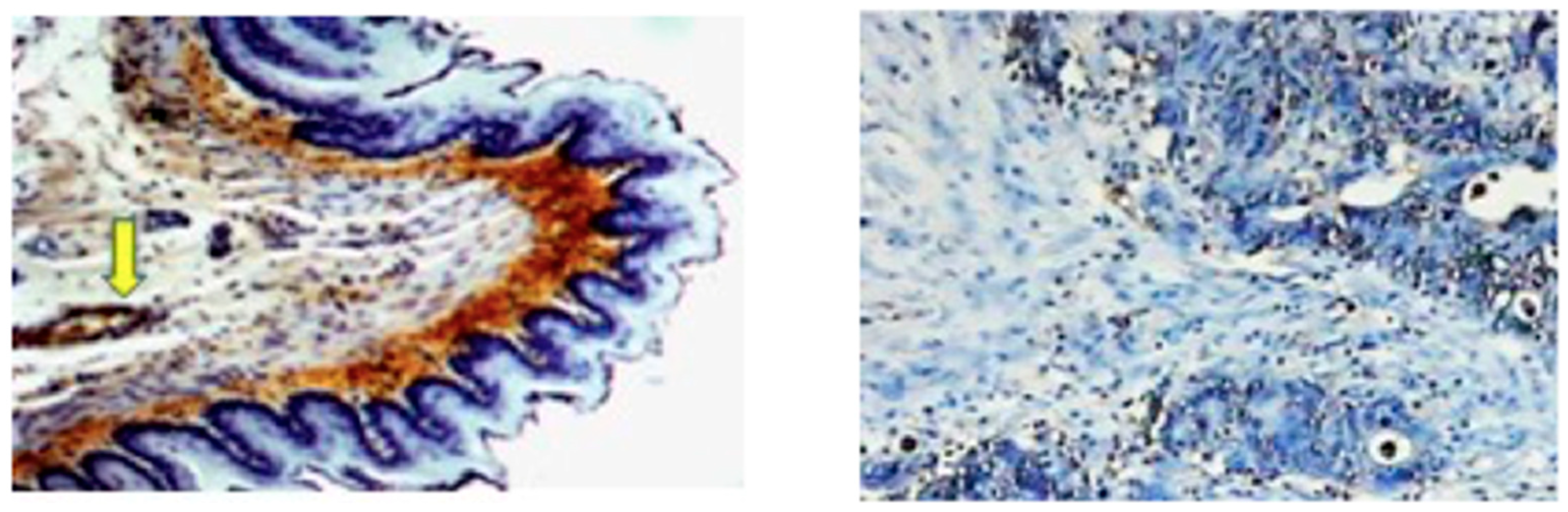
2. Immunochemical Methods Employed for the Detection of 4-HNE–Protein Adducts
3. 4-HNE Adduction to Proteins—Selective or Random Event?
4. Mass Spectrometry as a Tool to Study the 4-HNE–Protein Adductome
5. Biological Relevance of Protein Residue Modification by 4-HNE
6. 4-HNE–Protein Adducts Impacting Cellular Homeostasis
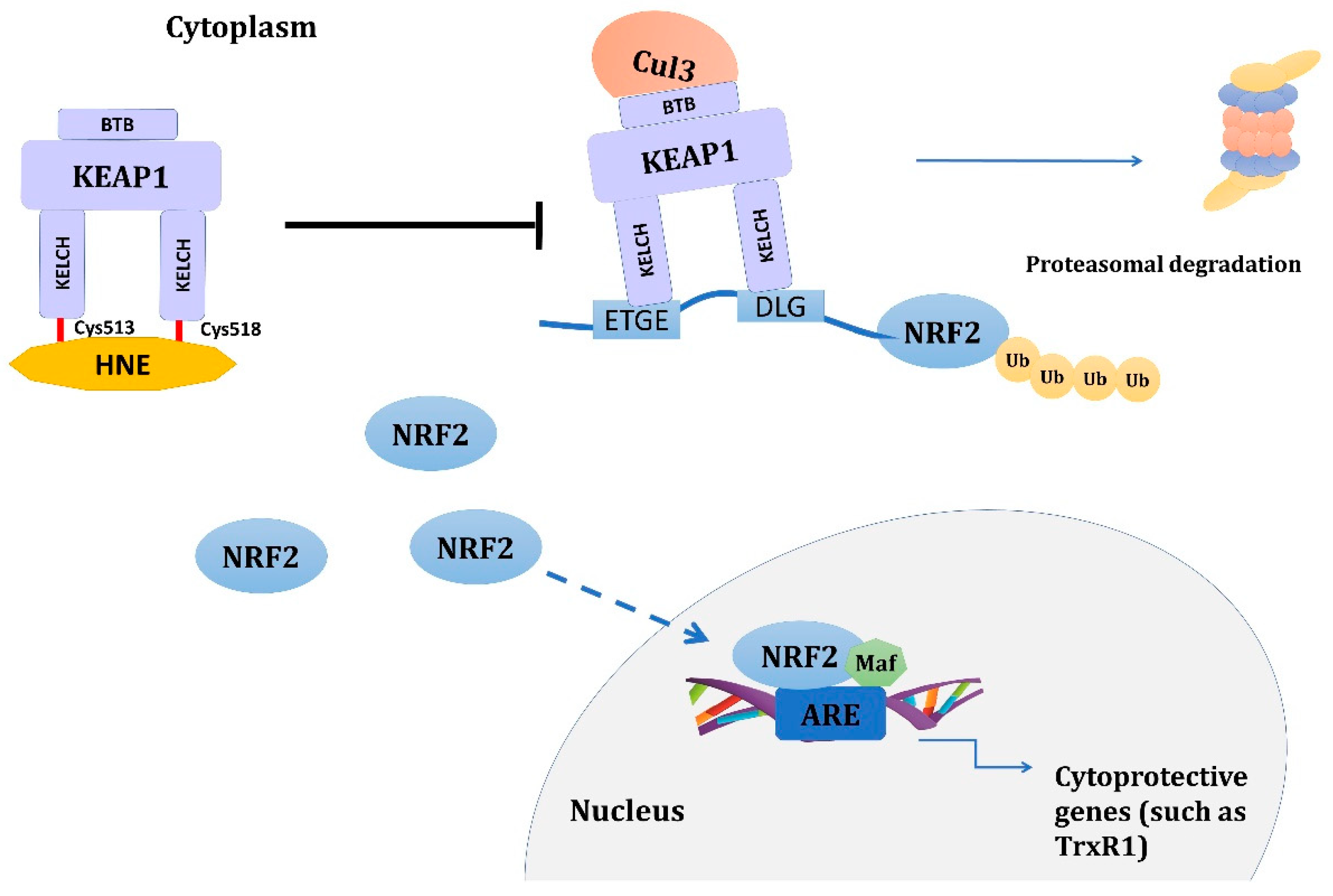
6.1. HNE–KEAP1 Adducts and Adaptive Response
6.2. 4-HNE–Protein Adducts in Ferroptosis
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sies, H. Oxidative Stress: Introductory Remarks. In Oxidative Stress; Sies, H., Ed.; Academic Press: London, UK, 1985; pp. 1–8. [Google Scholar]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [Green Version]
- Ramani, S.; Pathak, A.; Dalal, V.; Paul, A.; Biswas, S. Oxidative Stress in Autoimmune Diseases: An Under Dealt Malice. Curr. Protein Pept. Sci. 2020, 21, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Milkovic, L.; Cipak Gasparovic, A.; Cindric, M.; Mouthuy, P.A.; Zarkovic, N. Short Overview of ROS as Cell Function Regulators and Their Implications in Therapy Concepts. Cells 2019, 8, 793. [Google Scholar] [CrossRef] [Green Version]
- Sies, H. On the history of oxidative stress: Concept and some aspects of current development. Curr. Opin. Toxicol. 2018, 7, 122–126. [Google Scholar] [CrossRef]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Cindrić, M.; Jakovčević, A.; Žarković, K.; Žarković, N. Lipid peroxidation in brain tumors. Neurochem. Int. 2021, 149, 105118. [Google Scholar] [CrossRef]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Zheng, Y.; Brash, A.R. Demonstration of HNE-related aldehyde formation via lipoxygenase-catalyzed synthesis of a bis-allylic dihydroperoxide intermediate. Chem. Res. Toxicol. 2013, 26, 896–903. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.; Tallman, K.A.; Porter, N.A.; Brash, A.R. Two distinct pathways of formation of 4-hydroxynonenal. Mechanisms of nonenzymatic transformation of the 9- and 13-hydroperoxides of linoleic acid to 4-hydroxyalkenals. J. Biol. Chem. 2001, 276, 20831–20838. [Google Scholar] [CrossRef] [Green Version]
- Bilska-Wilkosz, A.; Iciek, M.; Górny, M. Chemistry and Biochemistry Aspects of the 4-Hydroxy-2,3-trans-nonenal. Biomolecules 2022, 12, 145. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Porter, N.A.; Schneider, C.; Brash, A.R.; Yin, H. Formation of 4-hydroxynonenal from cardiolipin oxidation: Intramolecular peroxyl radical addition and decomposition. Free Radic. Biol. Med. 2011, 50, 166–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaur, R.J.; Siems, W.; Bresgen, N.; Eckl, P.M. 4-Hydroxy-nonenal-A Bioactive Lipid Peroxidation Product. Biomolecules 2015, 5, 2247–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siems, W.; Grune, T. Intracellular metabolism of 4-hydroxynonenal. Mol. Aspects Med. 2003, 24, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Bruenner, B.A.; Jones, A.D.; German, J.B. Direct characterization of protein adducts of the lipid peroxidation product 4-hydroxy-2-nonenal using electrospray mass spectrometry. Chem. Res. Toxicol. 1995, 8, 552–559. [Google Scholar] [CrossRef]
- Doorn, J.A.; Petersen, D.R. Covalent modification of amino acid nucleophiles by the lipid peroxidation products 4-hydroxy-2-nonenal and 4-oxo-2-nonenal. Chem. Res. Toxicol. 2002, 15, 1445–1450. [Google Scholar] [CrossRef]
- Lesgards, J.-F.; Frayne, I.R.; Comte, B.; Busseuil, D.; Rhéaume, E.; Tardif, J.-C.; Rosiers, C. Des Differential distribution of 4-hydroxynonenal adducts to sulfur and nitrogen residues in blood proteins as revealed using Raney nickel and gas chromatography-mass spectrometry. Free Radic. Biol. Med. 2009, 47, 1375–1385. [Google Scholar] [CrossRef]
- LoPachin, R.M.; Geohagen, B.C.; Gavin, T. Synaptosomal Toxicity and Nucleophilic Targets of 4-Hydroxy-2-Nonenal. Toxicol. Sci. 2009, 107, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Sauerland, M.; Mertes, R.; Morozzi, C.; Eggler, A.L.; Gamon, L.F.; Davies, M.J. Kinetic assessment of Michael addition reactions of alpha, beta-unsaturated carbonyl compounds to amino acid and protein thiols. Free Radic. Biol. Med. 2021, 169, 1–11. [Google Scholar] [CrossRef]
- Aslebagh, R.; Pfeffer, B.A.; Fliesler, S.J.; Darie, C.C. Mass spectrometry-based proteomics of oxidative stress: Identification of 4-hydroxy-2-nonenal (HNE) adducts of amino acids using lysozyme and bovine serum albumin as model proteins. Electrophoresis 2016, 37, 2615–2623. [Google Scholar] [CrossRef] [Green Version]
- Annangudi, S.P.; Deng, Y.; Gu, X.; Zhang, W.; Crabb, J.W.; Salomon, R.G. Low-density lipoprotein has an enormous capacity to bind (E)-4-hydroxynon-2-enal (HNE): Detection and characterization of lysyl and histidyl adducts containing multiple molecules of HNE. Chem. Res. Toxicol. 2008, 21, 1384–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakita, C.; Maeshima, T.; Yamazaki, A.; Shibata, T.; Ito, S.; Akagawa, M.; Ojika, M.; Yodoi, J.; Uchida, K. Stereochemical configuration of 4-hydroxy-2-nonenal-cysteine adducts and their stereoselective formation in a redox-regulated protein. J. Biol. Chem. 2009, 284, 28810–28822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niki, E. Oxidative stress and antioxidants: Distress or eustress? Arch. Biochem. Biophys. 2016, 595, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.P.; Jung, T.; Grune, T.; Siems, W. 4-Hydroxynonenal (HNE) modified proteins in metabolic diseases. Free Radic. Biol. Med. 2017, 111, 309–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, D.L.; Doorn, J.A.; Kiebler, Z.; Petersen, D.R. Cysteine modification by lipid peroxidation products inhibits protein disulfide isomerase. Chem. Res. Toxicol. 2005, 18, 1324–1331. [Google Scholar] [CrossRef]
- Milkovic, L.; Cipak Gasparovic, A.; Zarkovic, N. Overview on major lipid peroxidation bioactive factor 4-hydroxynonenal as pluripotent growth-regulating factor. Free Radic. Res. 2015, 49, 850–860. [Google Scholar] [CrossRef]
- Jaganjac, M.; Milkovic, L.; Gegotek, A.; Cindric, M.; Zarkovic, K.; Skrzydlewska, E.; Zarkovic, N. The relevance of pathophysiological alterations in redox signaling of 4-hydroxynonenal for pharmacological therapies of major stress-associated diseases. Free Radic. Biol. Med. 2020, 157, 128–153. [Google Scholar] [CrossRef]
- Zhang, H.; Forman, H.J. Signaling by 4-hydroxy-2-nonenal: Exposure protocols, target selectivity and degradation. Arch. Biochem. Biophys. 2017, 617, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Riahi, Y.; Cohen, G.; Shamni, O.; Sasson, S. Signaling and cytotoxic functions of 4-hydroxyalkenals. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E879–E886. [Google Scholar] [CrossRef] [Green Version]
- Palinski, W.; Ylä-Herttuala, S.; Rosenfeld, M.E.; Butler, S.W.; Socher, S.A.; Parthasarathy, S.; Curtiss, L.K.; Witztum, J.L. Antisera and monoclonal antibodies specific for epitopes generated during oxidative modification of low density lipoprotein. Arterioscler. Off. J. Am. Heart Assoc. Inc. 1990, 10, 325–335. [Google Scholar] [CrossRef] [Green Version]
- Jürgens, G.; Ashy, A.; Esterbauer, H. Detection of new epitopes formed upon oxidation of low-density lipoprotein, lipoprotein (a) and very-low-density lipoprotein. Use of an antiserum against 4-hydroxynonenal-modified low-density lipoprotein. Biochem. J. 1990, 265, 605–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Esterbauer, H.; Jürgens, G. Studies on epitopes on low-density lipoprotein modified by 4-hydroxynonenal. Biochemical characterization and determination. Biochem. J. 1992, 288 Pt 1, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Uchida, K.; Toyokuni, S.; Nishikawa, K.; Kawakishi, S.; Oda, H.; Hiai, H.; Stadtman, E.R. Michael Addition-Type 4-Hydroxy-2-nonenal Adducts in Modified Low-Density Lipoproteins: Markers for Atherosclerosis. Biochemistry 1994, 33, 12487–12494. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Szweda, L.I.; Chae, H.Z.; Stadtman, E.R. Immunochemical detection of 4-hydroxynonenal protein adducts in oxidized hepatocytes. Proc. Natl. Acad. Sci. USA 1993, 90, 8742–8746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyokuni, S.; Miyake, N.; Hiai, H.; Hagiwara, M.; Kawakishi, S.; Osawa, T.; Uchida, K. The monoclonal antibody specific for the 4-hydroxy-2-nonenal histidine adduct. FEBS Lett. 1995, 359, 189–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waeg, G.; Dimsity, G.; Esterbauer, H. Monoclonal antibodies for detection of 4-hydroxynonenal modified proteins. Free Radic. Res. 1996, 25, 149–159. [Google Scholar] [CrossRef]
- Uchida, K.; Stadtman, E.R. Modification of histidine residues in proteins by reaction with 4-hydroxynonenal. Proc. Natl. Acad. Sci. USA 1992, 89, 4544–4548. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.; Milkovic, L.; Bennett, S.J.; Griffiths, H.R.; Zarkovic, N.; Grune, T. Measurement of HNE-protein adducts in human plasma and serum by ELISA-Comparison of two primary antibodies. Redox Biol. 2013, 1, 226–233. [Google Scholar] [CrossRef] [Green Version]
- Hartley, D.P.; Kroll, D.J.; Petersen, D.R. Prooxidant-initiated lipid peroxidation in isolated rat hepatocytes: Detection of 4-hydroxynonenal- and malondialdehyde-protein adducts. Chem. Res. Toxicol. 1997, 10, 895–905. [Google Scholar] [CrossRef]
- Campos-Pinto, I.; Méndez, L.; Schouten, J.; Wilkins, J.; Fedorova, M.; Pitt, A.R.; Davis, P.; Spickett, C.M. Epitope mapping and characterization of 4-hydroxy-2-nonenal modified-human serum albumin using two different polyclonal antibodies. Free Radic. Biol. Med. 2019, 144, 234–244. [Google Scholar] [CrossRef]
- Zarkovic, K.; Jakovcevic, A.; Zarkovic, N. Contribution of the HNE-immunohistochemistry to modern pathological concepts of major human diseases. Free Radic. Biol. Med. 2017, 111, 110–126. [Google Scholar] [CrossRef] [PubMed]
- Živković, M.; Žarković, K.; Škrinjar, L.; Waeg, G.; Poljak-Blaži, M.; Šunjić, S.B.; Schaur, R.J.; Žarković, N. A new method for detection of HNE-histidine conjugates in rat inflammatory cells. Croat. Chem. Acta 2005, 78, 91–98. [Google Scholar]
- Borovic, S.; Rabuzin, F.; Waeg, G.; Zarkovic, N. Enzyme-linked immunosorbent assay for 4-hydroxynonenal-histidine conjugates. Free Radic. Res. 2006, 40, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Miriyala, S.; Miao, L.; Mitov, M.; Schnell, D.; Dhar, S.K.; Cai, J.; Klein, J.B.; Sultana, R.; Butterfield, D.A.; et al. Redox proteomic identification of HNE-bound mitochondrial proteins in cardiac tissues reveals a systemic effect on energy metabolism after doxorubicin treatment. Free Radic. Biol. Med. 2014, 72, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spickett, C.M.; Wiswedel, I.; Siems, W.; Zarkovic, K.; Zarkovic, N. Advances in methods for the determination of biologically relevant lipid peroxidation products. Free Radic. Res. 2010, 44, 1172–1202. [Google Scholar] [CrossRef] [PubMed]
- Breusing, N.; Grune, T.; Andrisic, L.; Atalay, M.; Bartosz, G.; Biasi, F.; Borovic, S.; Bravo, L.; Casals, I.; Casillas, R.; et al. An inter-laboratory validation of methods of lipid peroxidation measurement in UVA-treated human plasma samples. Free Radic. Res. 2010, 44, 1203–1215. [Google Scholar] [CrossRef]
- Andersen, C.; Grønnemose, A.L.; Pedersen, J.N.; Nowak, J.S.; Christiansen, G.; Nielsen, J.; Mulder, F.A.A.; Otzen, D.E.; Jørgensen, T.J.D. Lipid Peroxidation Products HNE and ONE Promote and Stabilize Alpha-Synuclein Oligomers by Chemical Modifications. Biochemistry 2021, 60, 3644–3658. [Google Scholar] [CrossRef]
- Williams, M.V.; Wishnok, J.S.; Tannenbaum, S.R. Covalent adducts arising from the decomposition products of lipid hydroperoxides in the presence of cytochrome c. Chem. Res. Toxicol. 2007, 20, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Tallman, K.A.; Porter, N.A.; Liebler, D.C. Quantitative chemoproteomics for site-specific analysis of protein alkylation by 4-hydroxy-2-nonenal in cells. Anal. Chem. 2015, 87, 2535–2541. [Google Scholar] [CrossRef] [Green Version]
- Tzeng, S.-C.; Maier, C.S. Label-Free Proteomics Assisted by Affinity Enrichment for Elucidating the Chemical Reactivity of the Liver Mitochondrial Proteome toward Adduction by the Lipid Electrophile 4-hydroxy-2-nonenal (HNE). Front. Chem. 2016, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Arashiki, N.; Otsuka, Y.; Ito, D.; Yang, M.; Komatsu, T.; Sato, K.; Inaba, M. The covalent modification of spectrin in red cell membranes by the lipid peroxidation product 4-hydroxy-2-nonenal. Biochem. Biophys. Res. Commun. 2010, 391, 1543–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultana, R.; Perluigi, M.; Butterfield, D.A. Lipid peroxidation triggers neurodegeneration: A redox proteomics view into the Alzheimer disease brain. Free Radic. Biol. Med. 2013, 62, 157–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atalay, S.; Gęgotek, A.; Skrzydlewska, E. Protective Effects of Cannabidiol on the Membrane Proteome of UVB-Irradiated Keratinocytes. Antioxidants 2021, 10, 402. [Google Scholar] [CrossRef]
- Just, J.; Jung, T.; Friis, N.A.; Lykkemark, S.; Drasbek, K.; Siboska, G.; Grune, T.; Kristensen, P. Identification of an unstable 4-hydroxynoneal modification on the 20S proteasome subunit α7 by recombinant antibody technology. Free Radic. Biol. Med. 2015, 89, 786–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colzani, M.; Criscuolo, A.; Casali, G.; Carini, M.; Aldini, G. A method to produce fully characterized ubiquitin covalently modified by 4-hydroxy-nonenal, glyoxal, methylglyoxal, and malondialdehyde. Free Radic. Res. 2016, 50, 328–336. [Google Scholar] [CrossRef]
- Pillon, N.J.; Vella, R.E.; Souleere, L.; Becchi, M.; Lagarde, M.; Soulage, C.O. Structural and functional changes in human insulin induced by the lipid peroxidation byproducts 4-hydroxy-2-nonenal and 4-hydroxy-2-hexenal. Chem. Res. Toxicol. 2011, 24, 752–762. [Google Scholar] [CrossRef]
- Hauck, A.K.; Zhou, T.; Hahn, W.; Petegrosso, R.; Kuang, R.; Chen, Y.; Bernlohr, D.A. Obesity-induced protein carbonylation in murine adipose tissue regulates the DNA-binding domain of nuclear zinc finger proteins. J. Biol. Chem. 2018, 293, 13464–13476. [Google Scholar] [CrossRef] [Green Version]
- Galligan, J.J.; Fritz, K.S.; Backos, D.S.; Shearn, C.T.; Smathers, R.L.; Jiang, H.; MacLean, K.N.; Reigan, P.R.; Petersen, D.R. Oxidative stress-mediated aldehyde adduction of GRP78 in a mouse model of alcoholic liver disease: Functional independence of ATPase activity and chaperone function. Free Radic. Biol. Med. 2014, 73, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Jaganjac, M.; Milkovic, L.; Zarkovic, N.; Zarkovic, K. Oxidative stress and regeneration. Free Radic. Biol. Med. 2022, 181, 154–165. [Google Scholar] [CrossRef]
- Geib, T.; Iacob, C.; Jribi, R.; Fernandes, J.; Benderdour, M.; Sleno, L. Identification of 4-hydroxynonenal-modified proteins in human osteoarthritic chondrocytes. J. Proteomics 2021, 232, 104024. [Google Scholar] [CrossRef]
- Al-Menhali, A.S.; Anderson, C.; Gourine, A.V.; Abramov, A.Y.; D’Souza, A.; Jaganjac, M. Proteomic Analysis of Cardiac Adaptation to Exercise by High Resolution Mass Spectrometry. Front. Mol. Biosci. 2021, 8, 723858. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cong, Y.; Quan, B.; Lan, T.; Chu, X.; Ye, Z.; Hou, X.; Wang, C. Chemoproteomic profiling of targets of lipid-derived electrophiles by bioorthogonal aminooxy probe. Redox Biol. 2017, 12, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Domingues, P.; Wroński, A.; Ambrożewicz, E.; Skrzydlewska, E. The Proteomic Profile of Keratinocytes and Lymphocytes in Psoriatic Patients. Proteomics. Clin. Appl. 2019, 13, e1800119. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Zhang, Y.; Xiong, Y.; Tao, T.; Wang, Y.; Yao, J.; Zhang, L.; Yan, G.; Bao, H.; Lu, H. Highly Selective and Large Scale Mass Spectrometric Analysis of 4-Hydroxynonenal Modification via Fluorous Derivatization and Fluorous Solid-Phase Extraction. Anal. Chem. 2017, 89, 3093–3100. [Google Scholar] [CrossRef]
- Wang, C.; Weerapana, E.; Blewett, M.M.; Cravatt, B.F. A chemoproteomic platform to quantitatively map targets of lipid-derived electrophiles. Nat. Methods 2014, 11, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Hauck, A.K.; Zhou, T.; Upadhyay, A.; Sun, Y.; O’connor, M.B.; Chen, Y.; Bernlohr, D.A. Histone carbonylation is a redox-regulated epigenomic mark that accumulates with obesity and aging. Antioxidants 2020, 9, 1210. [Google Scholar] [CrossRef]
- Gęgotek, A.; Domingues, P.; Wroński, A.; Wójcik, P.; Skrzydlewska, E. Proteomic plasma profile of psoriatic patients. J. Pharm. Biomed. Anal. 2018, 155, 185–193. [Google Scholar] [CrossRef]
- Golizeh, M.; Geib, T.; Sleno, L. Identification of 4-hydroxynonenal protein targets in rat, mouse and human liver microsomes by two-dimensional liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2016, 30, 1488–1494. [Google Scholar] [CrossRef] [PubMed]
- Tanito, M.; Haniu, H.; Elliott, M.H.; Singh, A.K.; Matsumoto, H.; Anderson, R.E. Identification of 4-hydroxynonenal-modified retinal proteins induced by photooxidative stress prior to retinal degeneration. Free Radic. Biol. Med. 2006, 41, 1847–1859. [Google Scholar] [CrossRef]
- Camarillo, J.M.; Rose, K.L.; Galligan, J.J.; Xu, S.; Marnett, L.J. Covalent Modification of CDK2 by 4-Hydroxynonenal as a Mechanism of Inhibition of Cell Cycle Progression. Chem. Res. Toxicol. 2016, 29, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Lavilla, C.J.; Billacura, M.P.; Hanna, K.; Boocock, D.J.; Coveney, C.; Miles, A.K.; Foulds, G.A.; Murphy, A.; Tan, A.; Jackisch, L.; et al. Carnosine protects stimulus-secretion coupling through prevention of protein carbonyl adduction events in cells under metabolic stress. Free Radic. Biol. Med. 2021, 175, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Codreanu, S.G.; Zhang, B.; Sobecki, S.M.; Billheimer, D.D.; Liebler, D.C. Global analysis of protein damage by the lipid electrophile 4-hydroxy-2-nonenal. Mol. Cell. Proteomics 2009, 8, 670–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila, A.; Tallman, K.A.; Jacobs, A.T.; Liebler, D.C.; Porter, N.A.; Marnett, L.J. Identification of Protein Targets of 4-Hydroxynonenal Using Click Chemistry for ex Vivo Biotinylation of Azido and Alkynyl Derivatives. Chem. Res. Toxicol. 2008, 21, 432–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.-Y.H.; Tallman, K.A.; Liebler, D.C.; Porter, N.A. An azido-biotin reagent for use in the isolation of protein adducts of lipid-derived electrophiles by streptavidin catch and photorelease. Mol. Cell. Proteomics 2009, 8, 2080–2089. [Google Scholar] [CrossRef] [Green Version]
- Connor, R.E.; Marnett, L.J.; Liebler, D.C. Protein-selective capture to analyze electrophile adduction of hsp90 by 4-hydroxynonenal. Chem. Res. Toxicol. 2011, 24, 1275–1282. [Google Scholar] [CrossRef]
- Zarkovic, N.; Cipak, A.; Jaganjac, M.; Borovic, S.; Zarkovic, K. Pathophysiological relevance of aldehydic protein modifications. J. Proteomics 2013, 92, 239–247. [Google Scholar] [CrossRef]
- Al-Menhali, A.S.; Banu, S.; Angelova, P.R.; Barcaru, A.; Horvatovich, P.; Abramov, A.Y.; Jaganjac, M. Lipid peroxidation is involved in calcium dependent upregulation of mitochondrial metabolism in skeletal muscle. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129487. [Google Scholar] [CrossRef]
- Maniti, O.; François-Moutal, L.; Lecompte, M.-F.; Vial, C.; Lagarde, M.; Guichardant, M.; Marcillat, O.; Granjon, T. Protein “amyloid-like” networks at the phospholipid membrane formed by 4-hydroxy-2-nonenal-modified mitochondrial creatine kinase. Mol. Membr. Biol. 2015, 32, 1–10. [Google Scholar] [CrossRef]
- Larroque-Cardoso, P.; Mucher, E.; Grazide, M.-H.; Josse, G.; Schmitt, A.-M.; Nadal-Wolbold, F.; Zarkovic, K.; Salvayre, R.; Nègre-Salvayre, A. 4-Hydroxynonenal impairs transforming growth factor-β1-induced elastin synthesis via epidermal growth factor receptor activation in human and murine fibroblasts. Free Radic. Biol. Med. 2014, 71, 427–436. [Google Scholar] [CrossRef]
- Mihalas, B.P.; Bromfield, E.G.; Sutherland, J.M.; De Iuliis, G.N.; McLaughlin, E.A.; John Aitken, R.; Nixon, B. Oxidative damage in naturally aged mouse oocytes is exacerbated by dysregulation of proteasomal activity. J. Biol. Chem. 2018, 293, 18944–18964. [Google Scholar] [CrossRef] [Green Version]
- Retamal, M.A.; Fiori, M.C.; Fernandez-Olivares, A.; Linsambarth, S.; Peña, F.; Quintana, D.; Stehberg, J.; Altenberg, G.A. 4-Hydroxynonenal induces Cx46 hemichannel inhibition through its carbonylation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158705. [Google Scholar] [CrossRef] [PubMed]
- Elrayess, M.A.; Almuraikhy, S.; Kafienah, W.; Al-Menhali, A.; Al-Khelaifi, F.; Bashah, M.; Zarkovic, K.; Zarkovic, N.; Waeg, G.; Alsayrafi, M.; et al. 4-hydroxynonenal causes impairment of human subcutaneous adipogenesis and induction of adipocyte insulin resistance. Free Radic. Biol. Med. 2017, 104, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaganjac, M.; Almuraikhy, S.; Al-Khelaifi, F.; Al-Jaber, M.; Bashah, M.; Mazloum, N.A.; Zarkovic, K.; Zarkovic, N.; Waeg, G.; Kafienah, W.; et al. Combined metformin and insulin treatment reverses metabolically impaired omental adipogenesis and accumulation of 4-hydroxynonenal in obese diabetic patients. Redox Biol. 2017, 12, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žarković, N.; Orehovec, B.; Milković, L.; Baršić, B.; Tatzber, F.; Wonisch, W.; Tarle, M.; Kmet, M.; Mataić, A.; Jakovčević, A.; et al. Preliminary Findings on the Association of the Lipid Peroxidation Product 4-Hydroxynonenal with the Lethal Outcome of Aggressive COVID-19. Antioxidants 2021, 10, 1341. [Google Scholar] [CrossRef]
- Zarkovic, N.; Jakovcevic, A.; Mataic, A.; Jaganjac, M.; Vukovic, T.; Waeg, G.; Zarkovic, K. Post-mortem Findings of Inflammatory Cells and the Association of 4-Hydroxynonenal with Systemic Vascular and Oxidative Stress in Lethal COVID-19. Cells 2022, 11, 444. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, A.; Collado, A.; Tengbom, J.; Jiao, T.; Wodaje, T.; Johansson, N.; Farnebo, F.; Färnert, A.; Yang, J.; Lundberg, J.O.; et al. Erythrocytes Induce Vascular Dysfunction in COVID-19. JACC. Basic Transl. Sci. 2022, 7, 193–204. [Google Scholar] [CrossRef]
- Casós, K.; Zaragozá, M.C.; Zarkovic, N.; Zarkovic, K.; Andrisic, L.; Portero-Otín, M.; Cacabelos, D.; Mitjavila, M.T. A fish-oil-rich diet reduces vascular oxidative stress in apoE(-/-) mice. Free Radic. Res. 2010, 44, 821–829. [Google Scholar] [CrossRef]
- Zarkovic, K.; Larroque-Cardoso, P.; Pucelle, M.; Salvayre, R.; Waeg, G.; Nègre-Salvayre, A.; Zarkovic, N. Elastin aging and lipid oxidation products in human aorta. Redox Biol. 2015, 4, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Perkovic, M.N.; Jaganjac, M.; Milkovic, L.; Horvat, T.; Rojo, D.; Zarkovic, K.; Ćorić, M.; Hudolin, T.; Waeg, G.; Orehovec, B.; et al. Relationship between 4-Hydroxynonenal (4-HNE) as Systemic Biomarker of Lipid Peroxidation and Metabolomic Profiling of Patients with Prostate Cancer. Biomolecules 2023, 13, 145. [Google Scholar] [CrossRef]
- Wakita, C.; Honda, K.; Shibata, T.; Akagawa, M.; Uchida, K. A method for detection of 4-hydroxy-2-nonenal adducts in proteins. Free Radic. Biol. Med. 2011, 51, 1–4. [Google Scholar] [CrossRef]
- Kumano-Kuramochi, M.; Shimozu, Y.; Wakita, C.; Ohnishi-Kameyama, M.; Shibata, T.; Matsunaga, S.; Takano-Ishikawa, Y.; Watanabe, J.; Goto, M.; Xie, Q.; et al. Identification of 4-hydroxy-2-nonenal-histidine adducts that serve as ligands for human lectin-like oxidized LDL receptor-1. Biochem. J. 2012, 442, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavez, J.; Chung, W.-G.; Miranda, C.L.; Singhal, M.; Stevens, J.F.; Maier, C.S. Site-Specific Protein Adducts of 4-Hydroxy-2(E)-Nonenal in Human THP-1 Monocytic Cells: Protein Carbonylation Is Diminished by Ascorbic Acid. Chem. Res. Toxicol. 2010, 23, 37–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codreanu, S.G.; Ullery, J.C.; Zhu, J.; Tallman, K.A.; Beavers, W.N.; Porter, N.A.; Marnett, L.J.; Zhang, B.; Liebler, D.C. Alkylation damage by lipid electrophiles targets functional protein systems. Mol. Cell. Proteomics 2014, 13, 849–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimsrud, P.A.; Picklo, M.J.S.; Griffin, T.J.; Bernlohr, D.A. Carbonylation of adipose proteins in obesity and insulin resistance: Identification of adipocyte fatty acid-binding protein as a cellular target of 4-hydroxynonenal. Mol. Cell. Proteomics 2007, 6, 624–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearn, C.T.; Fritz, K.S.; Reigan, P.; Petersen, D.R. Modification of Akt2 by 4-hydroxynonenal inhibits insulin-dependent Akt signaling in HepG2 cells. Biochemistry 2011, 50, 3984–3996. [Google Scholar] [CrossRef]
- Eliuk, S.M.; Renfrow, M.B.; Shonsey, E.M.; Barnes, S.; Kim, H. Active site modifications of the brain isoform of creatine kinase by 4-hydroxy-2-nonenal correlate with reduced enzyme activity: Mapping of modified sites by Fourier transform-ion cyclotron resonance mass spectrometry. Chem. Res. Toxicol. 2007, 20, 1260–1268. [Google Scholar] [CrossRef]
- Crabb, J.W.; O’Neil, J.; Miyagi, M.; West, K.; Hoff, H.F. Hydroxynonenal inactivates cathepsin B by forming Michael adducts with active site residues. Protein Sci. 2002, 11, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Isom, A.L.; Barnes, S.; Wilson, L.; Kirk, M.; Coward, L.; Darley-Usmar, V. Modification of Cytochrome c by 4-hydroxy-2-nonenal: Evidence for histidine, lysine, and arginine-aldehyde adducts. J. Am. Soc. Mass Spectrom. 2004, 15, 1136–1147. [Google Scholar] [CrossRef] [Green Version]
- Musatov, A.; Carroll, C.A.; Liu, Y.-C.; Henderson, G.I.; Weintraub, S.T.; Robinson, N.C. Identification of bovine heart cytochrome c oxidase subunits modified by the lipid peroxidation product 4-hydroxy-2-nonenal. Biochemistry 2002, 41, 8212–8220. [Google Scholar] [CrossRef]
- Bennaars-Eiden, A.; Higgins, L.; Hertzel, A.V.; Kapphahn, R.J.; Ferrington, D.A.; Bernlohr, D.A. Covalent modification of epithelial fatty acid-binding protein by 4-hydroxynonenal in vitro and in vivo. Evidence for a role in antioxidant biology. J. Biol. Chem. 2002, 277, 50693–50702. [Google Scholar] [CrossRef] [Green Version]
- Sampey, B.P.; Carbone, D.L.; Doorn, J.A.; Drechsel, D.A.; Petersen, D.R. 4-Hydroxy-2-nonenal adduction of extracellular signal-regulated kinase (Erk) and the inhibition of hepatocyte Erk-Est-like protein-1-activating protein-1 signal transduction. Mol. Pharmacol. 2007, 71, 871–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szweda, L.I.; Uchida, K.; Tsai, L.; Stadtman, E.R. Inactivation of glucose-6-phosphate dehydrogenase by 4-hydroxy-2-nonenal. Selective modification of an active-site lysine. J. Biol. Chem. 1993, 268, 3342–3347. [Google Scholar] [CrossRef] [PubMed]
- Backos, D.S.; Fritz, K.S.; Roede, J.R.; Petersen, D.R.; Franklin, C.C. Posttranslational modification and regulation of glutamate-cysteine ligase by the α,β-unsaturated aldehyde 4-hydroxy-2-nonenal. Free Radic. Biol. Med. 2011, 50, 14–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, A.E.; Morin, D.; Lamé, M.W.; Jones, A.D. Purification, Mass Spectrometric Characterization, and Covalent Modification of Murine Glutathione S-Transferases. Chem. Res. Toxicol. 1995, 8, 1054–1062. [Google Scholar] [CrossRef]
- Ishii, T.; Tatsuda, E.; Kumazawa, S.; Nakayama, T.; Uchida, K. Molecular basis of enzyme inactivation by an endogenous electrophile 4-hydroxy-2-nonenal: Identification of modification sites in glyceraldehyde-3-phosphate dehydrogenase. Biochemistry 2003, 42, 3474–3480. [Google Scholar] [CrossRef] [PubMed]
- Carbone, D.L.; Doorn, J.A.; Kiebler, Z.; Sampey, B.P.; Petersen, D.R. Inhibition of Hsp72-mediated protein refolding by 4-hydroxy-2-nonenal. Chem. Res. Toxicol. 2004, 17, 1459–1467. [Google Scholar] [CrossRef] [Green Version]
- Carbone, D.L.; Doorn, J.A.; Kiebler, Z.; Ickes, B.R.; Petersen, D.R. Modification of heat shock protein 90 by 4-hydroxynonenal in a rat model of chronic alcoholic liver disease. J. Pharmacol. Exp. Ther. 2005, 315, 8–15. [Google Scholar] [CrossRef]
- Aldini, G.; Gamberoni, L.; Orioli, M.; Beretta, G.; Regazzoni, L.; Maffei Facino, R.; Carini, M. Mass spectrometric characterization of covalent modification of human serum albumin by 4-hydroxy-trans-2-nonenal. J. Mass Spectrom. 2006, 41, 1149–1161. [Google Scholar] [CrossRef]
- Smathers, R.L.; Fritz, K.S.; Galligan, J.J.; Shearn, C.T.; Reigan, P.; Marks, M.J.; Petersen, D.R. Characterization of 4-HNE modified L-FABP reveals alterations in structural and functional dynamics. PLoS ONE 2012, 7, e38459. [Google Scholar] [CrossRef] [Green Version]
- Doorn, J.A.; Hurley, T.D.; Petersen, D.R. Inhibition of human mitochondrial aldehyde dehydrogenase by 4-hydroxynon-2-enal and 4-oxonon-2-enal. Chem. Res. Toxicol. 2006, 19, 102–110. [Google Scholar] [CrossRef]
- Roede, J.R.; Carbone, D.L.; Doorn, J.A.; Kirichenko, O.V.; Reigan, P.; Petersen, D.R. In vitro and in silico characterization of peroxiredoxin 6 modified by 4-hydroxynonenal and 4-oxononenal. Chem. Res. Toxicol. 2008, 21, 2289–2299. [Google Scholar] [CrossRef] [PubMed]
- Shearn, C.T.; Smathers, R.L.; Backos, D.S.; Reigan, P.; Orlicky, D.J.; Petersen, D.R. Increased carbonylation of the lipid phosphatase PTEN contributes to Akt2 activation in a murine model of early alcohol-induced steatosis. Free Radic. Biol. Med. 2013, 65, 680–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camarillo, J.M.; Ullery, J.C.; Rose, K.L.; Marnett, L.J. Electrophilic Modification of PKM2 by 4-Hydroxynonenal and 4-Oxononenal Results in Protein Cross-Linking and Kinase Inhibition. Chem. Res. Toxicol. 2017, 30, 635–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritz, K.S.; Galligan, J.J.; Smathers, R.L.; Roede, J.R.; Shearn, C.T.; Reigan, P.; Petersen, D.R. 4-hydroxynonenal inhibits SIRT3 via thiol-specific modification. Chem. Res. Toxicol. 2011, 24, 651–662. [Google Scholar] [CrossRef] [Green Version]
- Jang, E.J.; Jeong, H.O.; Park, D.; Kim, D.H.; Choi, Y.J.; Chung, K.W.; Park, M.H.; Yu, B.P.; Chung, H.Y. Src Tyrosine Kinase Activation by 4-Hydroxynonenal Upregulates p38, ERK/AP-1 Signaling and COX-2 Expression in YPEN-1 Cells. PLoS ONE 2015, 10, e0129244. [Google Scholar] [CrossRef] [Green Version]
- Stewart, B.J.; Doom, J.A.; Petersen, D.R. Residue-specific adduction of tubulin by 4-hydroxynonenal and 4-oxononenal causes cross-linking and inhibits polymerization. Chem. Res. Toxicol. 2007, 20, 1111–1119. [Google Scholar] [CrossRef]
- Dalleau, S.; Baradat, M.; Guéraud, F.; Huc, L. Cell death and diseases related to oxidative stress:4-hydroxynonenal (HNE) in the balance. Cell Death Differ. 2013, 20, 1615–1630. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Chen, X.; Kang, R.; Kroemer, G. Ferroptosis: Molecular mechanisms and health implications. Cell Res. 2021, 31, 107–125. [Google Scholar] [CrossRef]
- Biasi, F.; Vizio, B.; Mascia, C.; Gaia, E.; Zarkovic, N.; Chiarpotto, E.; Leonarduzzi, G.; Poli, G. c-Jun N-terminal kinase upregulation as a key event in the proapoptotic interaction between transforming growth factor-beta1 and 4-hydroxynonenal in colon mucosa. Free Radic. Biol. Med. 2006, 41, 443–454. [Google Scholar] [CrossRef]
- Bauer, G.; Zarkovic, N. Revealing mechanisms of selective, concentration-dependent potentials of 4-hydroxy-2-nonenal to induce apoptosis in cancer cells through inactivation of membrane-associated catalase. Free Radic. Biol. Med. 2015, 81, 128–144. [Google Scholar] [CrossRef]
- Zhong, H.; Xiao, M.; Zarkovic, K.; Zhu, M.; Sa, R.; Lu, J.; Tao, Y.; Chen, Q.; Xia, L.; Cheng, S.; et al. Mitochondrial control of apoptosis through modulation of cardiolipin oxidation in hepatocellular carcinoma: A novel link between oxidative stress and cancer. Free Radic. Biol. Med. 2017, 102, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Sunjic, S.B.; Gasparovic, A.C.; Jaganjac, M.; Rechberger, G.; Meinitzer, A.; Grune, T.; Kohlwein, S.D.; Mihaljevic, B.; Zarkovic, N. Sensitivity of Osteosarcoma Cells to Concentration-Dependent Bioactivities of Lipid Peroxidation Product 4-Hydroxynonenal Depend on Their Level of Differentiation. Cells 2021, 10, 269. [Google Scholar] [CrossRef] [PubMed]
- Žarković, N.; Jaganjac, M.; Žarković, K.; Gęgotek, A.; Skrzydlewska, E. Spontaneous Regression of Cancer: Revealing Granulocytes and Oxidative Stress as the Crucial Double-edge Sword. Front. Biosci. (Landmark Ed.) 2022, 27, 119. [Google Scholar] [CrossRef] [PubMed]
- Shoeb, M.; Ansari, N.H.; Srivastava, S.K.; Ramana, K. V 4-Hydroxynonenal in the pathogenesis and progression of human diseases. Curr. Med. Chem. 2014, 21, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Poll, G.; Leonarduzzi, G.; Charpotto, E. 4-Hydroxynonenal Signaling. In Signal Transduction by Reactive Oxygen and Nitrogen Species: Pathways and Chemical Principles; Forman, H.J., Fukuto, J., Torres, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 180–193. [Google Scholar]
- Sonowal, H.; Ramana, K. V 4-Hydroxy-Trans-2-Nonenal in the Regulation of Anti-Oxidative and Pro-Inflammatory Signaling Pathways. Oxid. Med. Cell. Longev. 2019, 2019, 5937326. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Sharma, P.; Bailey, T.; Bhattarai, S.; Subedi, U.; Miller, C.; Ara, H.; Kidambi, S.; Sun, H.; Panchatcharam, M.; et al. Electrophilic Aldehyde 4-Hydroxy-2-Nonenal Mediated Signaling and Mitochondrial Dysfunction. Biomolecules 2022, 12, 1555. [Google Scholar] [CrossRef]
- Shireman, L.M.; Kripps, K.A.; Balogh, L.M.; Conner, K.P.; Whittington, D.; Atkins, W.M. Glutathione transferase A4-4 resists adduction by 4-hydroxynonenal. Arch. Biochem. Biophys. 2010, 504, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Huycke, M.M.; Herman, T.S.; Wang, X. Glutathione S-transferase alpha 4 induction by activator protein 1 in colorectal cancer. Oncogene 2016, 35, 5795–5806. [Google Scholar] [CrossRef]
- Jaganjac, M.; Milkovic, L.; Sunjic, S.B.; Zarkovic, N. The NRF2, Thioredoxin, and Glutathione System in Tumorigenesis and Anticancer Therapies. Antioxidants 2020, 9, 1151. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V.; Canning, P. Keap1, the cysteine-based mammalian intracellular sensor for electrophiles and oxidants. Arch. Biochem. Biophys. 2017, 617, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.-H.; Saito, Y.; Yoshida, Y.; Sekine, A.; Noguchi, N.; Niki, E. 4-Hydroxynonenal induces adaptive response and enhances PC12 cell tolerance primarily through induction of thioredoxin reductase 1 via activation of Nrf2. J. Biol. Chem. 2005, 280, 41921–41927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, X.; Fu, Y.; Long, M.J.C.; Haegele, J.A.; Ge, E.J.; Parvez, S.; Aye, Y. Temporally controlled targeting of 4-hydroxynonenal to specific proteins in living cells. J. Am. Chem. Soc. 2013, 135, 14496–14499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gall Trošelj, K.; Tomljanović, M.; Jaganjac, M.; Matijević Glavan, T.; Čipak Gašparović, A.; Milković, L.; Borović Šunjić, S.; Buttari, B.; Profumo, E.; Saha, S.; et al. Oxidative Stress and Cancer Heterogeneity Orchestrate NRF2 Roles Relevant for Therapy Response. Molecules 2022, 27, 1468. [Google Scholar] [CrossRef]
- Malhotra, D.; Portales-Casamar, E.; Singh, A.; Srivastava, S.; Arenillas, D.; Happel, C.; Shyr, C.; Wakabayashi, N.; Kensler, T.W.; Wasserman, W.W.; et al. Global mapping of binding sites for Nrf2 identifies novel targets in cell survival response through ChIP-Seq profiling and network analysis. Nucleic Acids Res. 2010, 38, 5718–5734. [Google Scholar] [CrossRef]
- Gao, Q.; Zhang, G.; Zheng, Y.; Yang, Y.; Chen, C.; Xia, J.; Liang, L.; Lei, C.; Hu, Y.; Cai, X.; et al. SLC27A5 deficiency activates NRF2/TXNRD1 pathway by increased lipid peroxidation in HCC. Cell Death Differ. 2020, 27, 1086–1104. [Google Scholar] [CrossRef]
- Hong, F.; Freeman, M.L.; Liebler, D.C. Identification of sensor cysteines in human Keap1 modified by the cancer chemopreventive agent sulforaphane. Chem. Res. Toxicol. 2005, 18, 1917–1926. [Google Scholar] [CrossRef]
- McMahon, M.; Lamont, D.J.; Beattie, K.A.; Hayes, J.D. Keap1 perceives stress via three sensors for the endogenous signaling molecules nitric oxide, zinc, and alkenals. Proc. Natl. Acad. Sci. USA 2010, 107, 18838–18843. [Google Scholar] [CrossRef] [Green Version]
- Parvez, S.; Fu, Y.; Li, J.; Long, M.J.C.; Lin, H.-Y.; Lee, D.K.; Hu, G.S.; Aye, Y. Substoichiometric hydroxynonenylation of a single protein recapitulates whole-cell-stimulated antioxidant response. J. Am. Chem. Soc. 2015, 137, 10–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, B.; Chen, X.B.; Ying, M.D.; He, Q.J.; Cao, J.; Yang, B. The Role of Ferroptosis in Cancer Development and Treatment Response. Front. Pharmacol. 2017, 8, 992. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Huang, J.; Yu, C.; Liu, J.; Gao, W.; Li, J.; Song, X.; Zhou, Z.; Li, C.; Xie, Y.; et al. A noncanonical function of EIF4E limits ALDH1B1 activity and increases susceptibility to ferroptosis. Nat. Commun. 2022, 13, 6318. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Y.; Lan, T.; Qin, W.; Zhu, Y.; Qin, K.; Gao, J.; Wang, H.; Hou, X.; Chen, N.; et al. Quantitative Profiling of Protein Carbonylations in Ferroptosis by an Aniline-Derived Probe. J. Am. Chem. Soc. 2018, 140, 4712–4720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Luo, M.; Zhang, K.; Zhang, J.; Gao, T.; Connell, D.O.; Yao, F.; Mu, C.; Cai, B.; Shang, Y.; et al. Nedd4 ubiquitylates VDAC2/3 to suppress erastin-induced ferroptosis in melanoma. Nat. Commun. 2020, 11, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.W.; Chhoy, P.; Mukhopadhyay, D.; Karner, E.R.; Mercurio, A.M. Targeting prominin2 transcription to overcome ferroptosis resistance in cancer. EMBO Mol. Med. 2021, 13, e13792. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.T.; Marnett, L.J. Heat Shock Factor 1 Attenuates 4-Hydroxynonenal-mediated Apoptosis. J. Biol. Chem. 2007, 282, 33412–33420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cyran, A.M.; Zhitkovich, A. Heat Shock Proteins and HSF1 in Cancer. Front. Oncol. 2022, 12, 860320. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Clone | Immunogen | Applications | Description | Specificity |
|---|---|---|---|---|
| HNEJ-2 | 4-HNE-modified KLH | IHC, WB, ELISA | mouse IgG1 kappa | Recognizes 4-HNE; Negligible reactivity with proteins that were treated with other aldehydes, such as 2-nonenal, 2-hexenal, 1-hexanal, 4-hydroxyhexenal, formaldehyde, or glutaraldehyde. |
| 12F7 | 4-HNE-modified KLH | WB, IHC, ICC/IF, ELISA | mouse IgG1 | Specific for 4-HNE-modified proteins. Does not detect free 4-HNE. Does not react with 4-hydroxyhexenal, acrolein, crotonaldehyde, hexanoyl lys, MDA, or methylglyoxal-modified proteins |
| 198960 | 4-HNE-modified KLH | WB, IHC, Simple Western | mouse IgG2b | Detects 4-hydroxynonenal adducts of histidine residues. No cross-reactivity with nitrotyrosine, formaldehyde, glutaraldehyde, or 4-hydroxy hexenaldehyde adducts is observed. |
| 9H132 | 4-HNE-modified KLH | WB | mouse IgG2b | Recognizes 4-HNE adducts of histidine residues. It does not cross-react with nitrotyrosine, formaldehyde, glutaraldehyde, or 4-hydroxy hexenaldehyde adducts. |
| IOFK-3 | 4-HNE-modified KLH | WB, IHC | mouse | This antibody shows almost negligible reactivity with proteins that were treated with other aldehydes such as: 2-nonenal, 2-hexenal, 1-hexenal, 4-hydroxy-2-hexenal, formaldehyde, or glutaraldehyde. |
| Sample Type | Sample Preparation | Method (Mass Analyzer) | Adducts Searched | Ref. |
|---|---|---|---|---|
| Adipocytes | Nuclear proteins, immunoprecipitation of carbonylated proteins, trypsin digestion | NanoLC-ESI-CID-MS/MS (Orbitrap) | 4-HNE–Cys, 4-HNE–His, 4-HNE–Lys | [58] |
| Chondrocytes | Whole cell lysates, nuclear and cytosolic protein extracts, trypsin or pepsin digestion followed by SPE | UHPLC-QTOF-MS/MS, TOF-MS | 4-HNE–Cys, 4-HNE–His, 4-HNE–Lys | [61] |
| Heart (left ventricle) samples | Whole tissue lysate, SDS-PAGE, trypsin digestion | NanoLC-ESI-HCD-MS/MS (Orbitrap and Iontrap), neutral ion loss (46, 52, 69, 78 m/z) triggered ETD-Iontrap MS/MS | 4-HNE–Cys, 4-HNE–His, 4-HNE–Lys, 4-HNE–Arg | [62] |
| HEK293T cells | Cell lysate, protein labeling, click chemistry, protein digestion | LC-HCD-MS/MS (Orbitrap) | Modification of Cys, His, and Lys | [63] |
| Keratinocytes and lymphocytes | Whole cell lysates, SDS-PAGE, trypsin digestion | LC-ESI-HCD-MS/MS (Quadrupole-Orbitrap) | 4-HNE–Cys, 4-HNE–His, 4-HNE–Lys | [64] |
| MCF-7 cells | Whole cell lysate, trypsin digestion, fluorous derivatization, fluorous SPE | UPLC-ESI-HCD-MS/MS (Quadrupole-Orbitrap) | 4-HNE–Cys, 4-HNE–His, 4-HNE–Lys, 4-HNE–Arg | [65] |
| MDA-MB-231 | Cell lysate, protein labeling, click chemistry, protein digestion | LC-MS/MS | 4-HNE–Cys | [66] |
| Mice white adipose depots; Drosophila Melanogaster Flies | Histone purification, immunoprecipitation of 4-HNE-modified proteins, SDS-PAGE, trypsin digestion | NanoLC-ESI-CID-MS/MS (Orbitrap and Iontrap) | 4-HNE–Cys, 4-HNE–His, 4-HNE–Lys | [67] |
| Plasma samples | Albumin removal, SDS-PAGE, trypsin digestion | LC-ESI-HCD-MS/MS (Quadrupole-Orbitrap) | 4-HNE–Cys, 4-HNE–His, 4-HNE–Lys | [68] |
| Rat, mouse, and human liver microsomes | Trypsin digestion followed by SPE clean-up, fractionation by HPLC-DAD | UHPLC-ESI-MS/MS (Quadrupole-TOF) | 4-HNE–Cys, 4-HNE–His, 4-HNE–Lys | [69] |
| Rat retina | S2-DE, WB, band excision, digestion with digested with 0.005% tosylsulfonyl phenylalanyl chloromethyl ketone-treated trypsin | MALDI-TOF/TOF | n.s. | [70] |
| RKO human colorectal cancer cells | Lysate of transfected cells and purification of CDK2–His, SDS-PAGE, selection of gel bands corresponding to CDK2–His, trypsin digestion | HPLC-ESI-MS/MS (Orbitrap and linear trap quadrupole) | 4-HNE–Cys, 4-HNE–His, 4-HNE–Lys, 4-HNE–Arg | [71] |
| Serum samples; Skeletal muscle cells; mouse pancreatic islets | Immunoprecipitation of 4-HNE-modified proteins, trypsin digestion | HPLC-ESI-TOF MS/MS | n.s. | [72] |
| Protein | Site Modified (Detected) | The Effect of 4-HNE Adduction | Ref. |
|---|---|---|---|
| Adipocyte fatty-acid-binding protein (A-FABP) | Cys117 | Decreases affinity for fatty acids. | [95] |
| Akt2 | His196, His267, Cys311 | Inhibits insulin-dependent Akt signaling by restricting substrate binding to Akt2. | [96] |
| Alpha-synuclein | His50 ** | Induces α-synuclein oligomerization and stabilizes it against dissociation to monomers. | [48] |
| Brain cytosolic isoform of creatine kinase (CK-BB) | His7, His26, His29, His66 *, Lys86, His97, Lys101, Cys141, Cys145, His191 *, His234, Cys254, His276, Cys283 *, His296 *, His305 | Dose-dependent reduction in enzyme activity; Cys283 is readily modified by 4-HNE even at low concentrations. | [97] |
| Cathepsin B | Cys29 (A chain), His150 (B chain) | Inactivation of the enzyme and loss of protease activity. | [98] |
| Cyclin-dependent kinase 2 (CDK2) | His60, His71 *, Lys129, His161 *, His268, His283, His295 | Decreases kinase activity and leads to cell cycle arrest. | [71] |
| Cytochrome c | Arg38, His33, Lys87 | Decreases protein pI and structure and could affect mitochondrial function. | [99] |
| Cytochrome c oxidase subunit VIII | His36 | Inactivation of the enzyme | [100] |
| Epithelial fatty acid-binding protein (E-FABP) | Lys115, Cys120 *, Cys127 | Stabilizes the E-FABP structure against chemical denaturation by guanidine hydrochloride. | [101] |
| Extracellular signal regulated kinase (Erk-1/2) | His178 | Modification of inactive cytosolic monomers results in inhibition of Erk-1/2 phosphorylation and activity. | [102] |
| Glucose-6-phosphate dehydrogenase (G6PD) | Lys (n.s.) | Inactivation of the enzyme | [103] |
| Glutamate–cysteine ligase (GCL) | GCLC subunit (Cys553) GCLM subunit (Cys35) | Increases the activity of the cellular enzyme. Increased activity of monomeric GCLC. Modified GCLM may affect formation of GCL holoenzyme. | [104] |
| Glutathione S-transferases (GST) isoforms: alpha (GSTA), mu (GSTM) and pi (GSTP) | n.s. | Decreases the catalytic activity of GST with 1-chloro-2,4-dinitrobenzene as substrate. The strongest effect was recorded for GSTP, where extensive modification with 4-HNE was accompanied by an almost complete loss of enzyme activity. | [105] |
| Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | His164, Cys244, Cys281, His327, Lys331 | The modifications detected indicate that the inactivation of GAPDH by 4-HNE is not due to modification of the active site but is likely due to impairment of the protein structure. | [106] |
| Heat shock protein 72-kDa (Hsp72) | Cys267 | Reduced Hsp72-mediated protein refolding efficiency and decreased affinity for ATP. | [107] |
| Heat shock protein 90-kDa (Hsp90) | Cys572 | Inhibition of Hsp90-mediated chaperone activity | [108] |
| Human serum albumin (HSA) | Cys34, His67, His146, Lys 195, Lys 199, His242, His288, His510, Lys525 | Cys34 is the residue most susceptible to 4-HNE adduction, which could impair HSA function. | [109] |
| Liver fatty acid-binding protein (L-FABP), unbound apo and lipid-bound holo | Apo L-FABP (Lys57, Cys69) Holo L-FABP (Lys6, Lys31, His43, Lys46, Lys57, Cys69) | Binding capacity for natural ligands is reduced. | [110] |
| Mitochondrial aldehyde dehydrogenase (ALDH2) | Cys302 * | Irreversible inhibition of enzyme activity at very high concentrations of the aldehyde when the active site is modified (500 μM), but reversible at lower concentrations (50 μM). | [111] |
| Peroxiredoxin 6 (PRX6) | Cys91 | Induces a distortion in the tertiary protein structure and alters the conformation of the active site. | [112] |
| Phosphatase and tensin homolog deleted on chromosome 10 (PTEN) | Cys71 *, Cys136, Lys147, Lys223, Cys250, Lys254, Lys313, Lys327 *, Lys344 | Inhibition of enzyme activity | [113] |
| Pyruvate kinase M2 isoform (PKM2) | Cys49, His274, Cys424 *, His439 *, Lys256 | Inhibits kinase activity. Cys424 is involved in protein-protein interactions, whereas it is His439 in fructose 1,6-bis-phosphate binding. | [114] |
| Sirtuin 3 (SIRT3) | Cys280 | Allosteric inhibition of SIRT3 activity. | [115] |
| Src Tyrosine Kinase (Src) | His236, Cys241, Cys248 * | Activation of Src by adduct formation at Cys248, which induces autophosphorylation of Tyr416. | [116] |
| Tubulin | α-Tubulin (Cys347, Cys376) β-Tubulin (Cys303) | Impairment of tubulin polymerization | [117] |
| ZAK kinase | Cys22 * | Inhibition of enzyme activity, resulting in a negative feedback mechanism that may reduce activation of the JNK pathway. | [66] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milkovic, L.; Zarkovic, N.; Marusic, Z.; Zarkovic, K.; Jaganjac, M. The 4-Hydroxynonenal–Protein Adducts and Their Biological Relevance: Are Some Proteins Preferred Targets? Antioxidants 2023, 12, 856. https://doi.org/10.3390/antiox12040856
Milkovic L, Zarkovic N, Marusic Z, Zarkovic K, Jaganjac M. The 4-Hydroxynonenal–Protein Adducts and Their Biological Relevance: Are Some Proteins Preferred Targets? Antioxidants. 2023; 12(4):856. https://doi.org/10.3390/antiox12040856
Chicago/Turabian StyleMilkovic, Lidija, Neven Zarkovic, Zlatko Marusic, Kamelija Zarkovic, and Morana Jaganjac. 2023. "The 4-Hydroxynonenal–Protein Adducts and Their Biological Relevance: Are Some Proteins Preferred Targets?" Antioxidants 12, no. 4: 856. https://doi.org/10.3390/antiox12040856