
Identification of a Depolymerase Specific for K64-Serotype Klebsiella pneumoniae: Potential Applications in Capsular Typing and Treatment

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Capsular Genotyping of K. pneumoniae Strains
2.2. Serological Typing of K. pneumoniae Strains
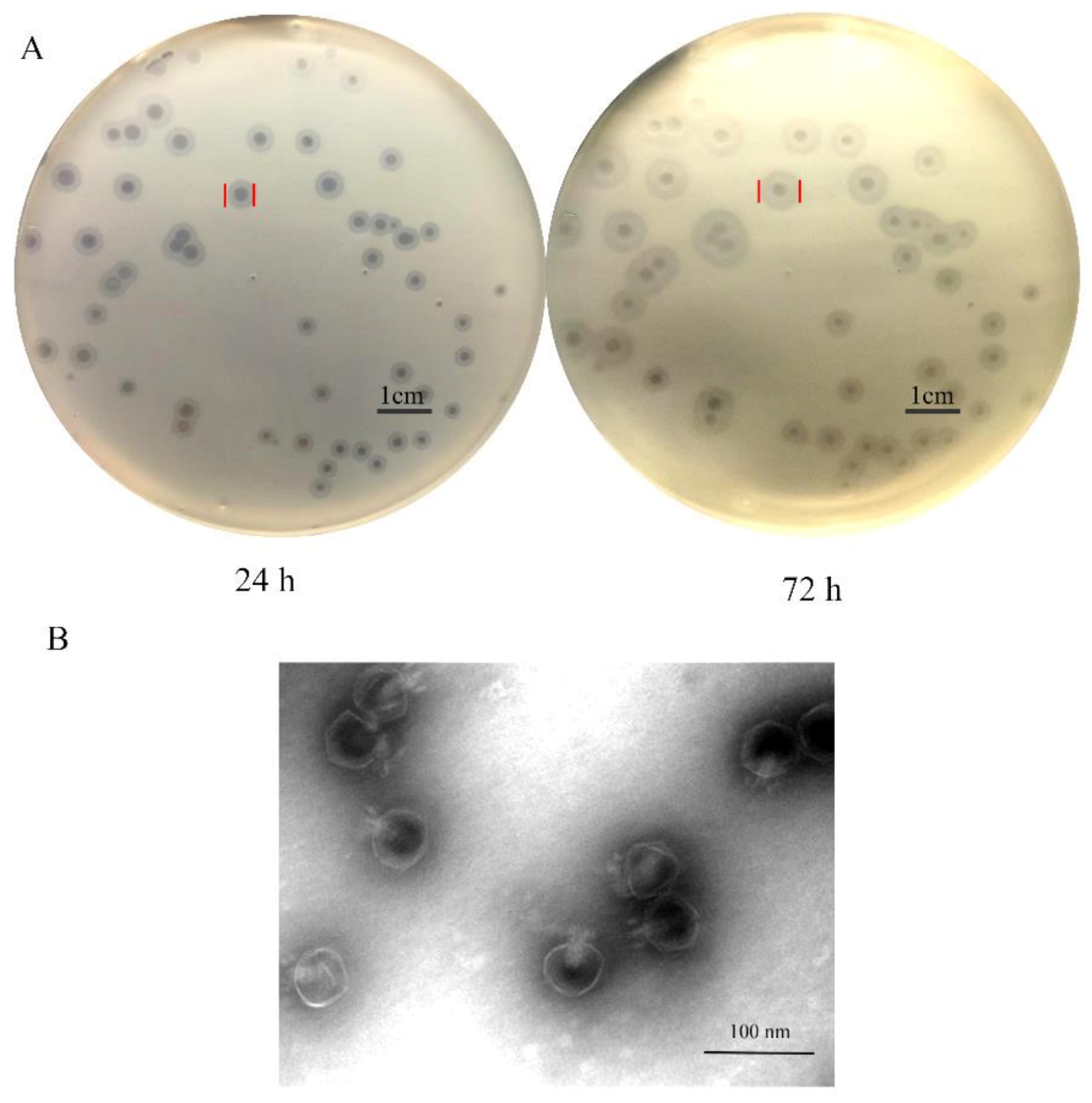
2.3. Isolation and Morphology of Phage SH-KP152410
2.4. Microbiological Characteristics and Lytic Spectrum of the SH-KP152410
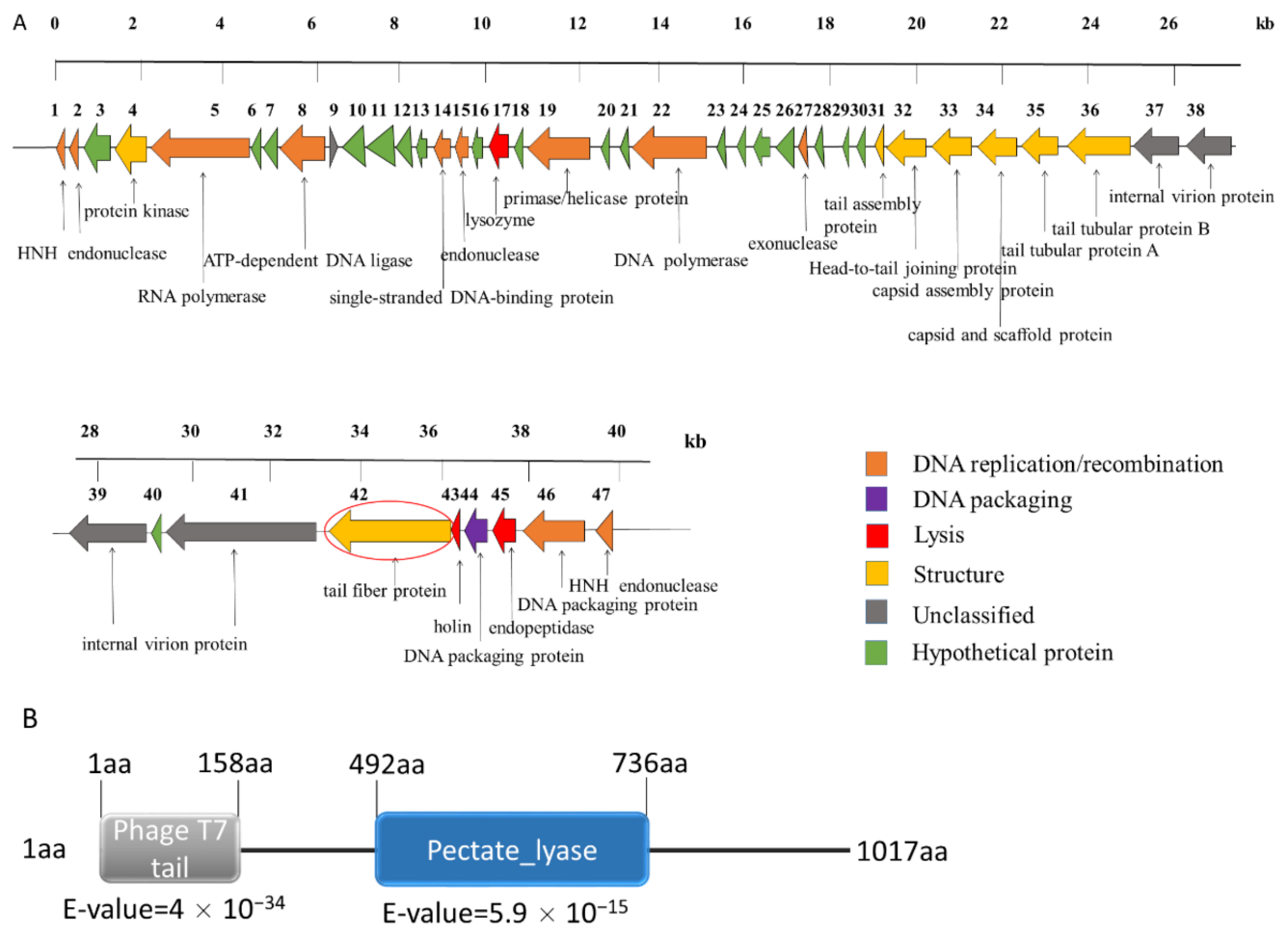
2.5. Genomic Analysis of the Phage SH-KP152410
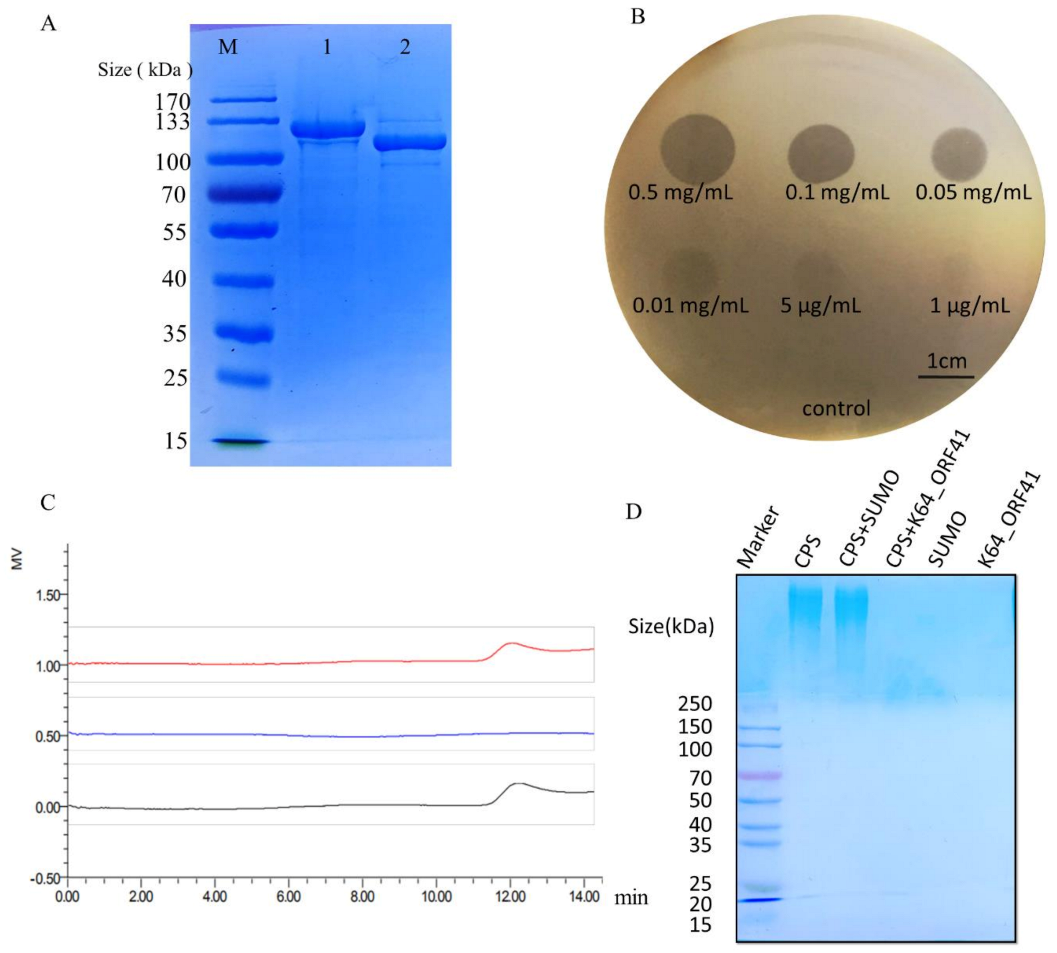
2.6. Prediction and Expression of the Putative Capsule Depolymerase
2.7. Assessment of the Recombinant Depolymerase Activity
2.8. Application of K64-ORF41 for K64 Capsular Typing
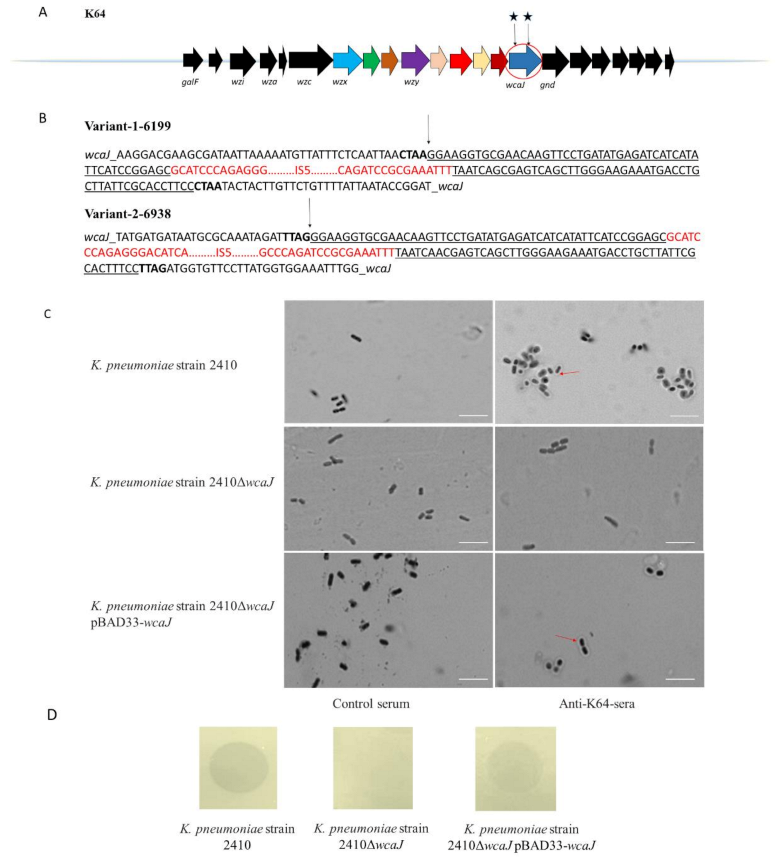
2.9. ISs Occur in the cps Region of K. pneumoniae
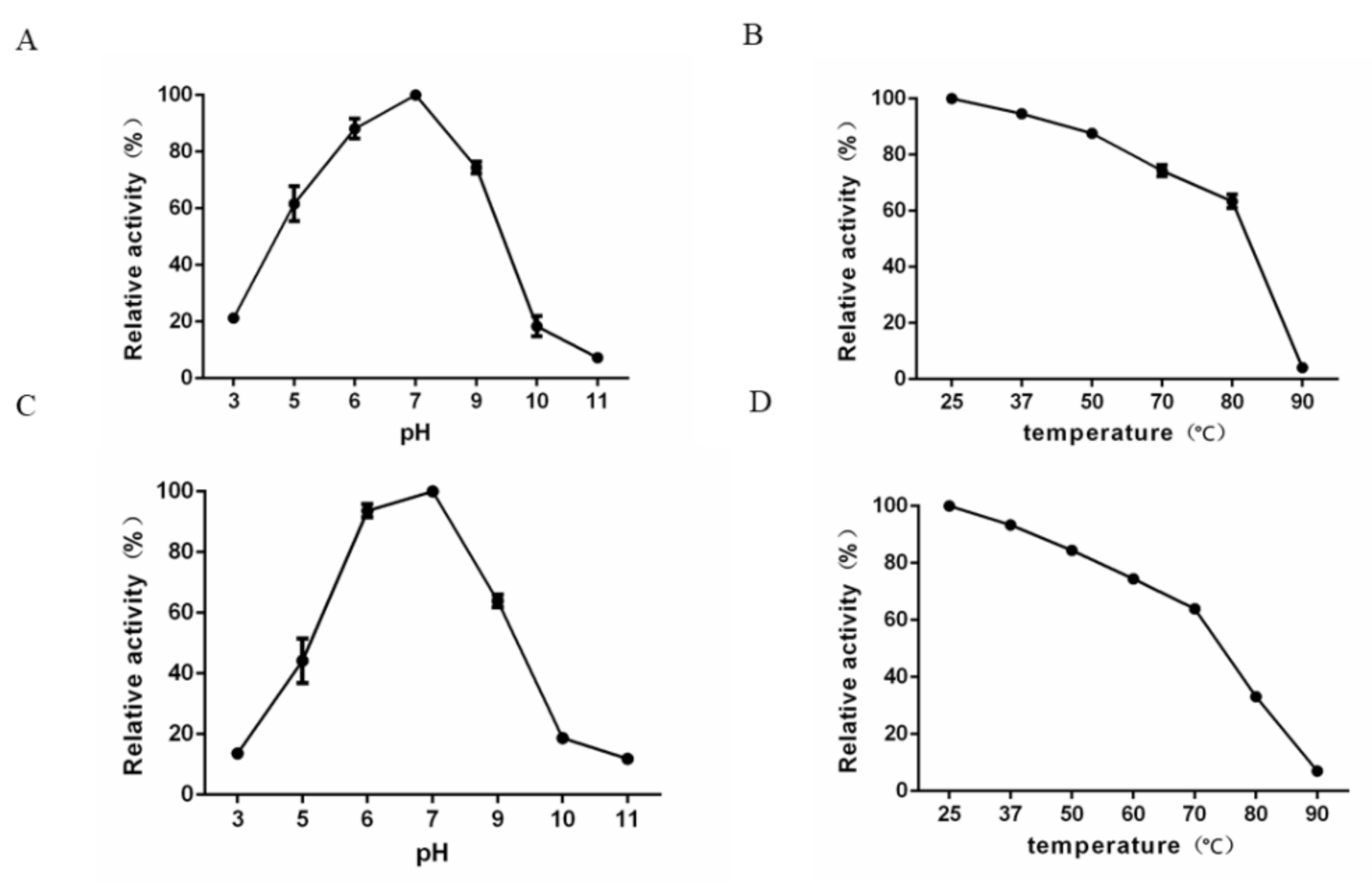
2.10. Characterization of the Depolymerase Activity and Stability at Various pH Values and Temperatures
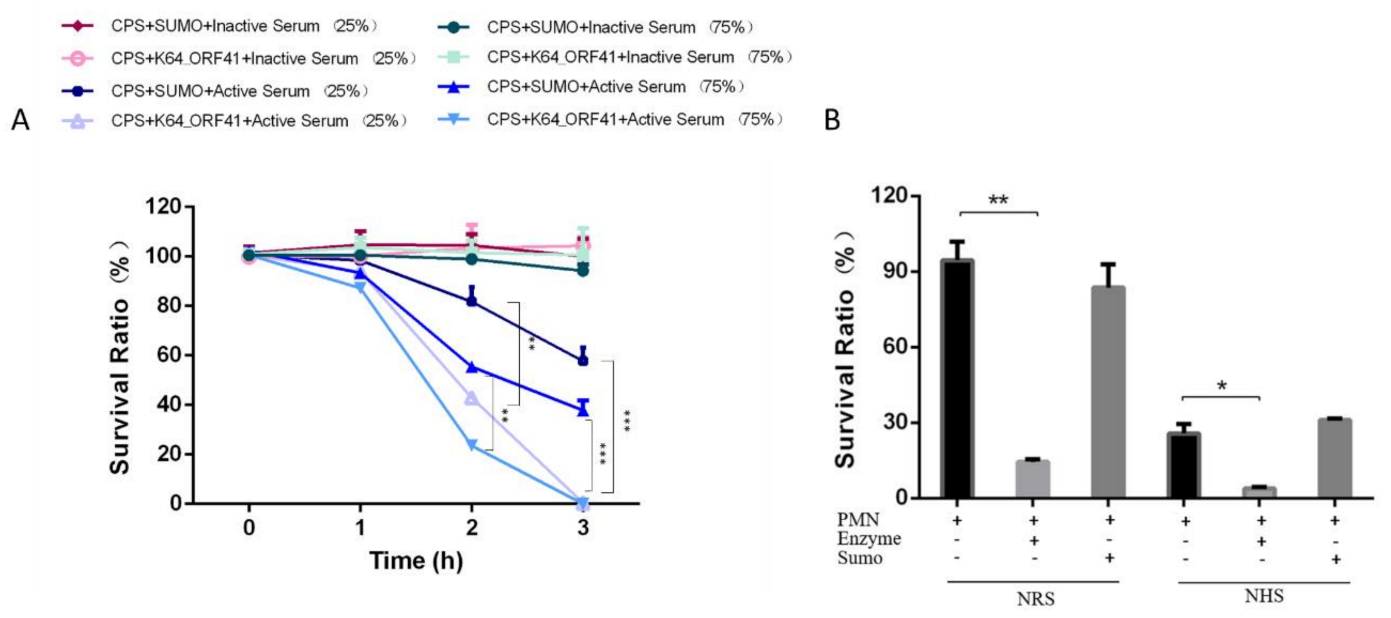
2.11. Serum Sensitivity of Depolymerase-Treated Bacteria
2.12. Neutrophil Killing of Enzyme-Treated Bacteria
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain Isolation and Identification
4.2. Bacteriophage Isolation
4.3. Transmission Electron Microscopy
4.4. Phage Activity Spectrum
4.5. Genomic DNA Sequencing and Annotation
4.6. Cloning, Expression, and Purification of the Recombinant Depolymerase
4.7. Depolymerase Activity by the Single-Spot Assay
4.8. Purification of the Capsular Polysaccharide (CPS)
4.9. Cleavage of K64 Capsular Polysaccharide by the Depolymerase in SEC-HPLC
4.10. Alcian Blue Staining
4.11. Capsular Serotyping
4.12. Sequence Analysis of the cps Region of K. pneumoniae
4.13. Effects of pH and Temperature on K64-ORF41 Activity and Stability
4.14. Serum Resistance Assay
4.15. Neutrophil Killing Assay
4.16. Preparation of K64-CPS Antiserum
4.17. Generation of an Isogenic wcaJ Deletion Mutant in K. pneumoniae
4.18. Cloning of wcaJ and Expression in K. pneumoniae
4.19. Statistics
4.20. Ethics Statement
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fung, C.-P.; Chang, F.-Y.; Lee, S.-C.; Hu, B.-S.; Kuo, B.I.-T.; Liu, C.-Y.; Ho, M.; Siu, L.K. A global emerging disease of Klebsiella pneumoniae liver abscess: Is serotype K1 an important factor for complicated endophthalmitis? Gut 2002, 50, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Hidron, A.I.; Edwards, J.R.; Patel, J.; Horan, T.C.; Sievert, D.M.; Pollock, D.A.; Fridkin, S.K. NHSN annual update: Antimicrobial-resistant pathogens associated with healthcare-associated infections: Annual summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2006–2007. Infect. Control Hosp. Epidemiol. 2008, 29, 996–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinert, R.R.; Low, D.E.; Rossi, F.; Zhang, X.; Wattal, C.; Dowzicky, M.J. Antimicrobial susceptibility among organisms from the Asia/Pacific Rim, Europe and Latin and North America collected as part of TEST and the in vitro activity of tigecycline. J. Antimicrob. Chemother. 2007, 60, 1018–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the Offense with a Strong Defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, C.-H.; Kuo, C.-F.; Wang, C.-H.; Wu, C.-M.; Tsao, N. Experimental Phage Therapy in TreatingKlebsiella pneumoniae-Mediated Liver Abscesses and Bacteremia in Mice. Antimicrob. Agents Chemother. 2011, 55, 1358–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saylor, C.; Dadachova, E.; Casadevall, A. Monoclonal antibody-based therapies for microbial diseases. Vaccine 2009, 27, G38–G46. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.J.; Lin, T.L.; Chen, Y.H.; Hsu, C.R.; Hsieh, P.F.; Wu, M.C.; Wang, J.T. Capsular types of Klebsiella pneumoniae revisited by wzc sequencing. PLoS ONE 2013, 8, e80670. [Google Scholar] [CrossRef]
- Edwards, P.R.; Fife, M.A. Capsule Types of Klebsiella. J. Infect. Dis. 1952, 91, 92–104. [Google Scholar] [CrossRef]
- Podschun, R.; Ullmann, U. Klebsiella spp. as Nosocomial Pathogens: Epidemiology, Taxonomy, Typing Methods, and Pathogenicity Factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.J.; Lin, T.L.; Lin, Y.T.; Su, P.A.; Chen, C.T.; Hsieh, P.F.; Hsu, C.R.; Chen, C.C.; Hsieh, Y.C.; Wang, J.T. Identification of capsular types in carbapenem-resistant Klebsiella pneumoniae strains by wzc sequencing and implications for capsule depolymerase treatment. Antimicrob. Agents Chemother. 2015, 59, 1038–1047. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.J.; Fang, H.C.; Yang, H.C.; Lin, T.L.; Hsieh, P.F.; Tsai, F.C.; Keynan, Y.; Wang, J.T. Capsular polysaccharide synthesis regions in Klebsiella pneu-moniae serotype K57 and a new capsular serotype. J. Clin. Microbiol. 2008, 46, 2231–2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisse, S.; Passet, V.; Haugaard, A.B.; Babosan, A.; Kassis-Chikhani, N.; Struve, C.; Decré, D. wzi Gene Sequencing, a Rapid Method for Determination of Capsular Type for Klebsiella Strains. J. Clin. Microbiol. 2013, 51, 4073–4078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; Wick, R.R.; Gorrie, C.; Jenney, A.; Follador, R.; Thomson, N.R.; Holt, K.E. Identification of Klebsiella capsule synthesis loci from whole genome data. Microb. Genom. 2016, 2, e000102. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.; Fang, C.; Lai, S.; Chang, S.; Wang, J. Genetic Determinants of Capsular Serotype K1 ofKlebsiella pneumoniaeCausing Primary Pyogenic Liver Abscess. J. Infect. Dis. 2006, 193, 645–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Li, P.; Niu, W.; Yuan, X.; Liu, H.; Huang, Y.; An, X.; Fan, H.; Zhangxiang, L.; Mi, L.; et al. Protective and therapeutic application of the depolymerase derived from a novel KN1 genotype of Klebsiella pneumoniae bacteriophage in mice. Res. Microbiol. 2019, 170, 156–164. [Google Scholar] [CrossRef]
- Pan, Y.J.; Lin, T.L.; Chen, Y.Y.; Lai, P.H.; Tsai, Y.T.; Hsu, C.R.; Hsieh, P.F.; Lin, Y.T.; Wang, J.T. Identification of three podoviruses infecting Klebsiella encoding capsule depolymerases that digest specific capsular types. Microb. Biotechnol. 2019, 12, 472–486. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, P.-F.; Lin, H.-H.; Lin, T.-L.; Chen, Y.-Y.; Wang, J.-T. Two T7-like Bacteriophages, K5-2 and K5-4, Each Encodes Two Capsule Depolymerases: Isolation and Functional Characterization. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Follador, R.; Heinz, E.; Wyres, K.L.; Ellington, M.J.; Kowarik, M.; Holt, K.E.; Thomson, N.R. The diversity of Klebsiella pneumoniae surface polysaccharides. Microb. Genom. 2016, 2, e000073. [Google Scholar] [CrossRef]
- Majkowska-Skrobek, G.; Latka, A.; Berisio, R.; Squeglia, F.; Maciejewska, B.; Briers, Y.; Drulis-Kawa, Z. Phage-Borne Depolymerases Decrease Klebsiella pneumoniae Resistance to Innate Defense Mechanisms. Front. Microbiol. 2018, 9, 2517. [Google Scholar] [CrossRef] [Green Version]
- Bellich, B.; Ravenscroft, N.; Rizzo, R.; Lagatolla, C.; D’Andrea, M.M.; Rossolini, G.M.; Cescutti, P. Structure of the capsular polysaccharide of the KPC-2-producing Klebsiella pneumoniae strain KK207-2 and assignment of the glycosyltransferases functions. Int. J. Biol. Macromol. 2019, 130, 536–544. [Google Scholar] [CrossRef]
- Heidary, M.; Nasiri, M.J.; Dabiri, H.; Tarashi, S. Prevalence of Drug-resistant Klebsiella pneumoniae in Iran: A Review Article. Iran. J. Public Health 2018, 47, 317–326. [Google Scholar] [PubMed]
- Li, B.; Zhao, Y.; Liu, C.; Chen, Z.; Zhou, D. Molecular pathogenesis of Klebsiella pneumoniae. Future Microbiol. 2014, 9, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.R.; Lin, T.L.; Pan, Y.J.; Hsieh, P.F.; Wang, J.T. Isolation of a bacteriophage specific for a new capsular type of Klebsiella pneu-moniae and characterization of its polysaccharide depolymerase. PLoS ONE 2013, 8, e70092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majkowska-Skrobek, G.; Łątka, A.; Berisio, R.; Maciejewska, B.; Squeglia, F.; Romano, M.; Lavigne, R.; Struve, C.; Drulis-Kawa, Z. Capsule-Targeting Depolymerase, Derived from Klebsiella KP36 Phage, as a Tool for the Development of Anti-Virulent Strategy. Viruses 2016, 8, 324. [Google Scholar] [CrossRef]
- Chaturongakul, S.; Ounjai, P. Phage-host interplay: Examples from tailed phages and Gram-negative bacterial pathogens. Front. Microbiol. 2014, 5, 442. [Google Scholar] [CrossRef] [Green Version]
- Maszewska, A. Phage associated polysaccharide depolymerases—characteristics and application. Postepy Hig. Med. Dosw. (Online) 2015, 69, 690–702. [Google Scholar] [CrossRef]
- Lin, T.-L.; Hsieh, P.-F.; Huang, Y.-T.; Lee, W.-C.; Tsai, Y.-T.; Su, P.-A.; Pan, Y.-J.; Hsu, C.-R.; Wu, M.-C.; Wang, J.-T. Isolation of a Bacteriophage and Its Depolymerase Specific for K1 Capsule of Klebsiella pneumoniae: Implication in Typing and Treatment. J. Infect. Dis. 2014, 210, 1734–1744. [Google Scholar] [CrossRef] [Green Version]
- Teng, T.; Li, Q.; Liu, Z.; Li, X.; Liu, Z.; Liu, H.; Liu, F.; Xie, L.; Wang, H.; Zhang, L.; et al. Characterization and genome analysis of novel Klebsiella phage Henu1 with lytic activity against clinical strains of Klebsiella pneumoniae. Arch. Virol. 2019, 164, 2389–2393. [Google Scholar] [CrossRef]
- Bowers, J.R.; Kitchel, B.; Driebe, E.M.; MacCannell, D.R.; Roe, C.; Lemmer, D.; De Man, T.; Rasheed, J.K.; Engelthaler, D.M.; Keim, P.; et al. Genomic Analysis of the Emergence and Rapid Global Dissemination of the Clonal Group 258 Klebsiella pneumoniae Pandemic. PLoS ONE 2015, 10, e0133727. [Google Scholar] [CrossRef] [Green Version]
- Koh, T.H.; Cao, D.; Shan, Q.Y.; Bacon, A.; Hsu, L.Y.; Ooi, E.E. Acquired carbapenemases in Enterobactericeae in Singapore, 1996–2012. Pathology 2013, 45, 600–603. [Google Scholar] [CrossRef]
- Huang, Y.H.; Chou, S.H.; Liang, S.W.; Ni, C.E.; Lin, Y.T.; Huang, Y.W.; Yang, T.C. Emergence of an XDR and carbapenemase-producing hypervirulent Klebsiella pneumoniae strain in Taiwan. J. Antimicrob. Chemother. 2018, 73, 2039–2046. [Google Scholar] [CrossRef] [PubMed]
- Cryz, S.J., Jr.; Mortimer, P.M.; Mansfield, V.; Germanier, R. Seroepidemiology of Klebsiella bacteremic isolates and implications for vaccine development. J. Clin. Microbiol. 1986, 23, 687–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, K.; Xiao, T.; David, S.; Wang, Q.; Zhou, Y.; Guo, L.; Aanensen, D.; Holt, K.E.; Thomson, N.R.; Grundmann, H.; et al. Novel Subclone of Carbapenem-Resistant Klebsiella pneumoniae Sequence Type 11 with Enhanced Virulence and Transmissibility, China. Emerg. Infect. Dis. 2020, 26, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.Y.; Qin, R.; Lu, Y.H.; Shen, J.; Zhang, S.; Wang, C.Y.; Yang, Y.Q.; Hu, F.P.; He, P. Capsular polysaccharide and lipopolysaccharide O type analysis of Klebsiella pneumoniae isolates by genotype in China. Epidemiol. Infect. 2020, 148, 1–13. [Google Scholar] [CrossRef]
- Kassa, T.; Chhibber, S. Thermal treatment of the bacteriophage lysate of Klebsiella pneumoniae B5055 as a step for the purifi-cation of capsular depolymerase enzyme. J. Virol. Methods 2012, 179, 135–141. [Google Scholar] [CrossRef]
- Majkowska-Skrobek, G.; Maciejewska, B. Bacteriophages and Phage-Derived Proteins—Application Approaches. Curr. Med. Chem. 2015, 22, 1757–1773. [Google Scholar] [CrossRef]
- Shu, H.-Y.; Fung, C.-P.; Liu, Y.-M.; Wu, K.-M.; Chen, Y.-T.; Li, L.-H.; Liu, T.-T.; Kirby, R.; Tsai, S.-F. Genetic diversity of capsular polysaccharide biosynthesis in Klebsiella pneumoniae clinical isolates. Microbiology 2009, 155, 4170–4183. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Guo, Y.; Li, J.; Zhang, Y.; Yang, Y.; Dong, N.; Zhu, D.; He, P.; Hu, F. In vitro and in vivo bactericidal activity of ceftazidime-avibactam against Carbapenemase–producing Klebsiella pneumoniae. Antimicrob. Resist. Infect. Control. 2018, 7, 142–149. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Jeng, Y.-Y.; Chen, T.-L.; Fung, C.-P. Bacteremic community-acquired pneumonia due to Klebsiella pneumoniae: Clinical and microbiological characteristics in Taiwan, 2001–2008. BMC Infect. Dis. 2010, 10, 307. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, F.M.; Dib-Hajj, F.; Shang, W.; Gootz, T.D. High-level carbapenem resistance in a Klebsiella pneumoniae clinical isolate is due to the combination of blaACT-1 β-lactamase production, porin OmpK35/36 insertional inactivation, and down-regulation of the phosphate transport porin PhoE. Antimicrob. Agents Chemother. 2006, 50, 3396–3406. [Google Scholar] [CrossRef] [Green Version]
- Squeglia, F.; Maciejewska, B.; Łątka, A.; Ruggiero, A.; Briers, Y.; Drulis-Kawa, Z.; Berisio, R. Structural and Functional Studies of a Klebsiella Phage Capsule Depolymerase Tailspike: Mechanistic Insights into Capsular Degradation. Structure 2020, 28, 613–624.e4. [Google Scholar] [CrossRef] [PubMed]
- Ernst, C.M.; Braxton, J.R.; Rodriguez-Osorio, C.A.; Zagieboylo, A.P.; Li, L.; Pironti, A.; Manson, A.L.; Nair, A.V.; Benson, M.; Cummins, K.; et al. Adaptive evolution of virulence and per-sistence in carbapenem-resistant Klebsiella pneumoniae. Nat. Med. 2020, 26, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Venturini, C.; Ben Zakour, N.L.; Bowring, B.; Morales, S.; Cole, R.; Kovach, Z.; Branston, S.; Kettle, E.; Thomson, N.; Iredell, J. Fine capsule variation affects bacteriophage susceptibility in Klebsiella pneumoniae ST258. FASEB J. 2020, 34, 10801–10817. [Google Scholar] [CrossRef] [PubMed]
- Kropinski, A.M.; Mazzocco, A.; Waddell, T.E.; Lingohr, E.; Johnson, R.P. Enumeration of Bacteriophages by Double Agar Overlay Plaque Assay. Methods Mol. Biol. 2009, 501, 69–76. [Google Scholar]
- Ramos, J.-L.; Duque, E.; Huertas, M.J.; Haïdour, A. Isolation and expansion of the catabolic potential of a Pseudomonas putida strain able to grow in the presence of high concentrations of aromatic hydrocarbons. J. Bacteriol. 1995, 177, 3911–3916. [Google Scholar] [CrossRef] [Green Version]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. Erratum: SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. GigaScience 2015, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Delcher, A.L.; Harmon, D.; Kasif, S.; White, O.; Salzberg, S.L. Improved microbial gene identification with GLIMMER. Nucleic Acids Res. 1999, 27, 4636–4641. [Google Scholar] [CrossRef]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef] [Green Version]
- Bales, P.M.; Renke, E.M.; May, S.L.; Shen, Y.; Nelson, D.C. Purification and Characterization of Biofilm-Associated EPS Exopolysac-charides from ESKAPE Organisms and Other Pathogens. PLoS ONE 2013, 8, e67950. [Google Scholar] [CrossRef] [Green Version]
- Rennie, R.P.; Duncan, I.B.R. Combined biochemical and serological typing of clinical isolates of Klebsiella. Appl. Microbiol. 1974, 28, 534–539. [Google Scholar] [CrossRef]
- Matsen, J.M.; Blazevic, D.J. Characterization of ornithine decarboxylase-positive, nonmotile strains of the Klebsiella-Enterobacter group. Appl. Microbiol. 1969, 18, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.F.; Yang, C.Y.; Lin, T.L.; Wang, J.T.; Yang, F.L.; Wu, S.H.; Hu, B.S.; Chou, T.Y.; Tsai, M.D.; Lin, C.H.; et al. Humoral immunity against capsule polysaccharide protects the host from magA+ Klebsiella pneumoniae-induced lethal disease by evading Toll-like receptor 4 signaling. Infect. Immun. 2009, 77, 615–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Fang, C.-T.; Chuang, Y.-P.; Shun, C.-T.; Chang, S.-C.; Wang, J.-T. A Novel Virulence Gene in Klebsiella pneumoniae Strains Causing Primary Liver Abscess and Septic Metastatic Complications. J. Exp. Med. 2004, 199, 697–705. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.D.; Porter, A.R.; Dorward, D.W.; Brinkworth, A.J.; Chen, L.; Kreiswirth, B.N.; DeLeo, F.R. Phagocytosis and Killing of Car-bapenem-Resistant ST258 Klebsiella pneumoniae by Human Neutrophils. J. Infect. Dis. 2016, 213, 1615–1622. [Google Scholar] [CrossRef] [Green Version]
- Nauseef, W.M. Isolation of human neutrophils from venous blood. Methods Mol. Biol. 2007, 412, 15–20. [Google Scholar]
- Sukhija, K.; Pyne, M.; Ali, S.; Orr, V.; Abedi, D.; Moo-Young, M.; Chou, C.P. Developing an Extended Genomic Engineering Approach Based on Recombineering to Knock-in Heterologous Genes to Escherichia coli Genome. Mol. Biotechnol. 2011, 51, 109–118. [Google Scholar] [CrossRef]
- Bi, D.; Jiang, X.; Sheng, Z.K.; Ngmenterebo, D.; Tai, C.; Wang, M.; Deng, Z.; Rajakumar, K.; Ou, H.Y. Mapping the resistance-associated mobilome of a car-bapenem-resistant Klebsiella pneumoniae strain reveals insights into factors shaping these regions and facilitates generation of a ‘resistance-disarmed’ model organism. J. Antimicrob. Chemother. 2015, 70, 2770–2774. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate No. | Genotype (wzi) | SH-KP152410 | K64-ORF41 | Quellung Test | Agglutination | |
|---|---|---|---|---|---|---|
| Plaques | Halos | |||||
| 13,882 1, 2410 2, Y8 2, 5085, 6003, 6088,6089, 6233, 6353, 6368, 6603, 6682, 6718, 6810, 6954, 6974, 7323, 7366, 7761,7804, 7871, 7880, 7888, 7901, 7982, 8004, 8046, 8047, 8061, 8171, 8207, 8213, 8310, 8315, 8328, 8417, 8422, 8456, 8497, 8608 | KL64 | + | + | + | + | + |
| 6082, 6199, 6483, 6821, 6938, 6938, 6990, 8206 | KL64 | − | − | − | − | − |
| 6118, 6876, 7168 | KL47 | − | − | − | − | − |
| 5146, 5170 | KL2 | − | − | − | − | − |
| 5080, 5174 | KL1 | − | − | − | − | − |
| 7830, 6409 | KL24 | − | − | − | − | − |
| 6795, 7072 | KL60 | − | − | − | − | − |
| 8191, 8666 | KL54 | − | − | − | − | − |
| 7607, 7616 | KL61 | − | − | − | − | − |
| 7485, 6422, 7321, 7485 | KL19 | − | − | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Sheng, Y.; Ma, R.; Xu, M.; Liu, F.; Qin, R.; Zhu, M.; Zhu, X.; He, P. Identification of a Depolymerase Specific for K64-Serotype Klebsiella pneumoniae: Potential Applications in Capsular Typing and Treatment. Antibiotics 2021, 10, 144. https://doi.org/10.3390/antibiotics10020144
Li J, Sheng Y, Ma R, Xu M, Liu F, Qin R, Zhu M, Zhu X, He P. Identification of a Depolymerase Specific for K64-Serotype Klebsiella pneumoniae: Potential Applications in Capsular Typing and Treatment. Antibiotics. 2021; 10(2):144. https://doi.org/10.3390/antibiotics10020144
Chicago/Turabian StyleLi, Jiayin, Yueying Sheng, Ruijing Ma, Mengsha Xu, Fuli Liu, Rong Qin, Mingxi Zhu, Xianchao Zhu, and Ping He. 2021. "Identification of a Depolymerase Specific for K64-Serotype Klebsiella pneumoniae: Potential Applications in Capsular Typing and Treatment" Antibiotics 10, no. 2: 144. https://doi.org/10.3390/antibiotics10020144