L-Tryptophan Enhances Intestinal Integrity in Diquat-Challenged Piglets Associated with Improvement of Redox Status and Mitochondrial Function


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals and Treatments
2.3. Performance Measurement and Sample Collection
2.4. Serum Intestinal Permeability Biomarker Concentrations
2.5. Ex Vivo Measurement of Intestinal Barrier Function
2.6. Intestinal Redox Status Measurements
2.7. Isolation of Intestinal Mitochondria
2.8. Intestinal Mitochondrial ROS Assay
2.9. Intestinal Mitochondrial Membrane Potential (ΔΨm) Assay
2.10. Determination of Jejunal Mitochondrial DNA (mtDNA) Content
2.11. RNA Isolation and Reverse Transcription Quantitative PCR (RT-qPCR)
2.12. Gel Electrophoresis and Western Blotting
2.13. Statistical Analysis
3. Results
3.1. Growth Performance
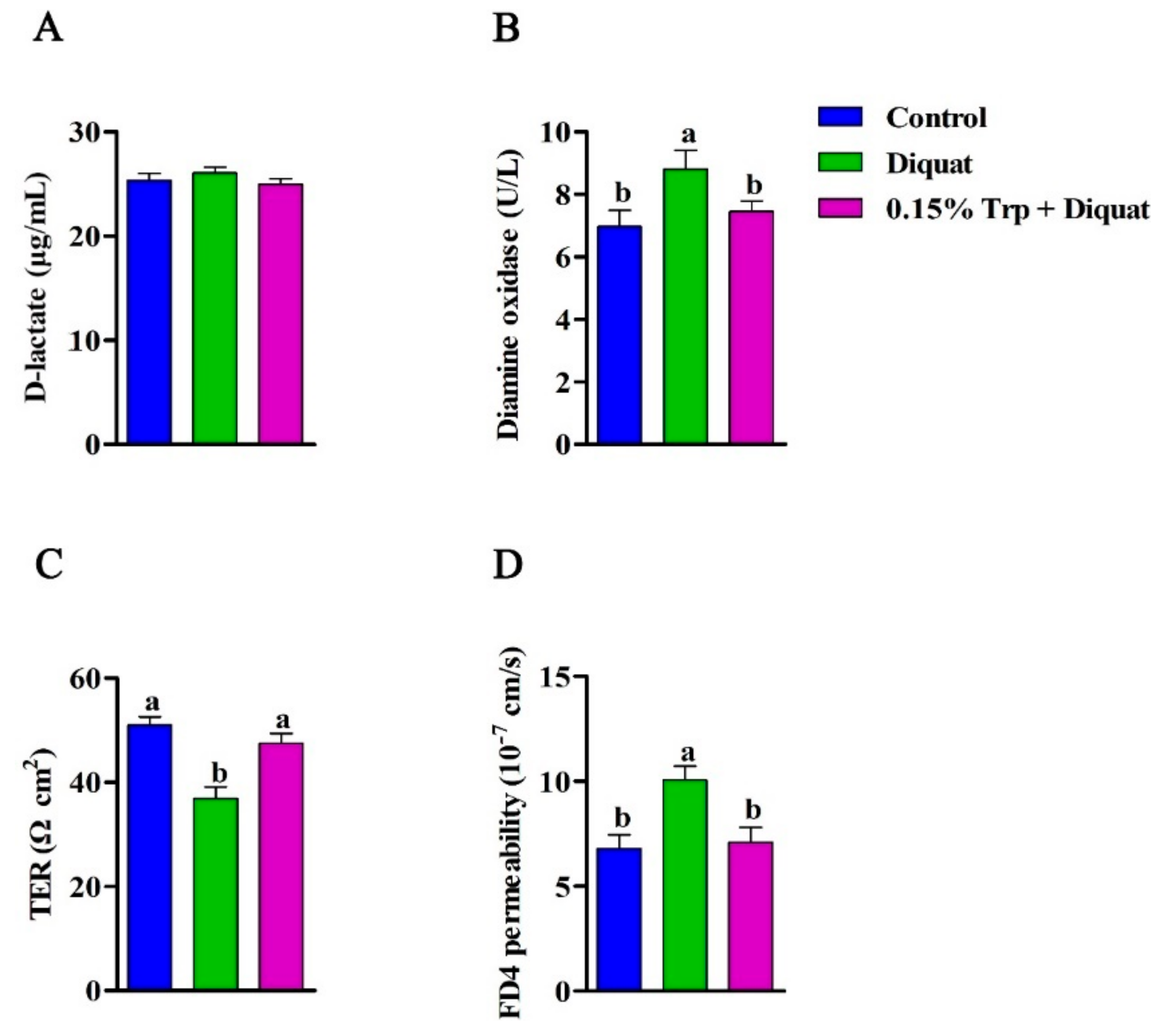
3.2. Intestinal Permeability
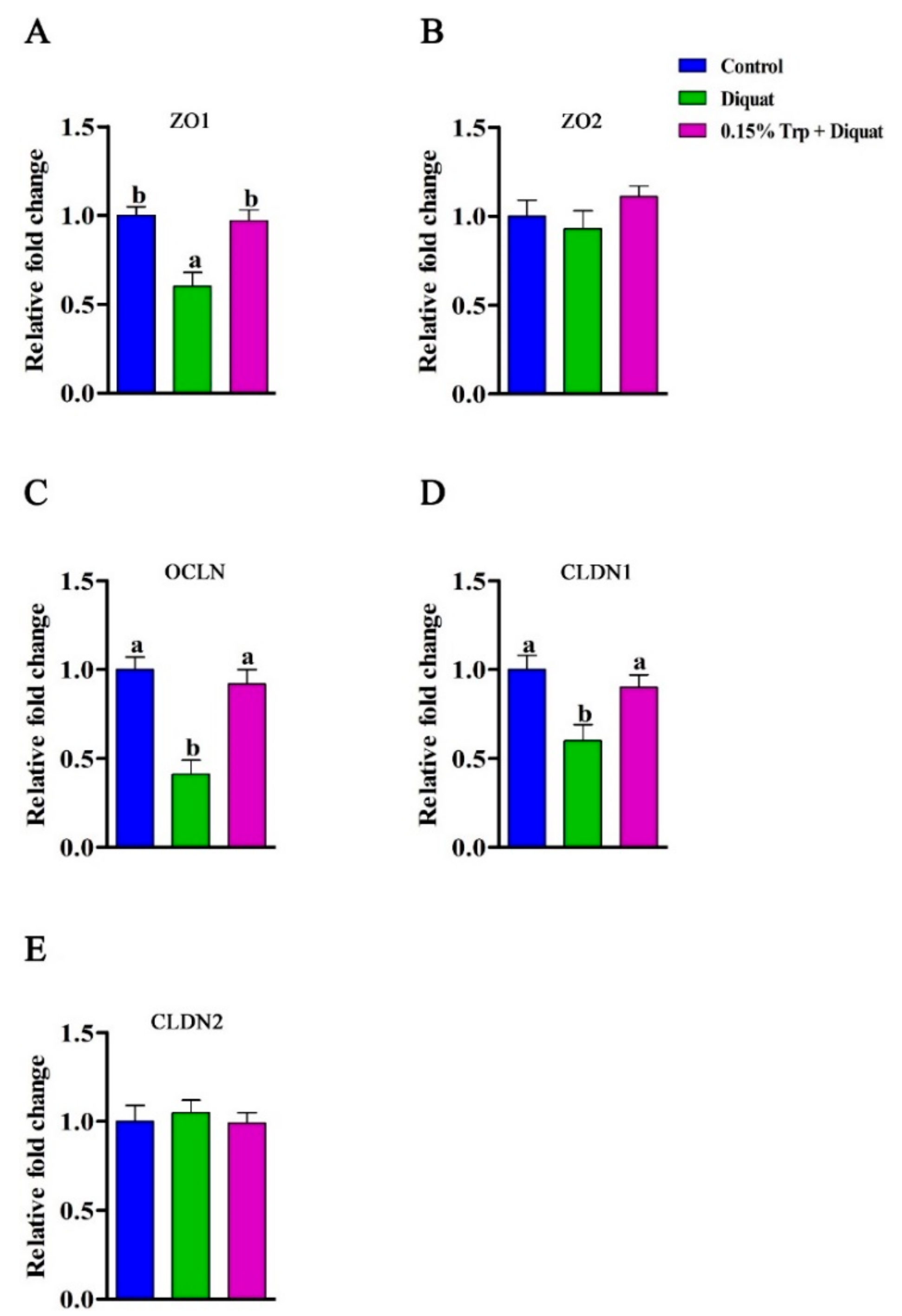
3.3. mRNA Abundances and Protein Expression of Tight Junction Proteins in the Jejunum
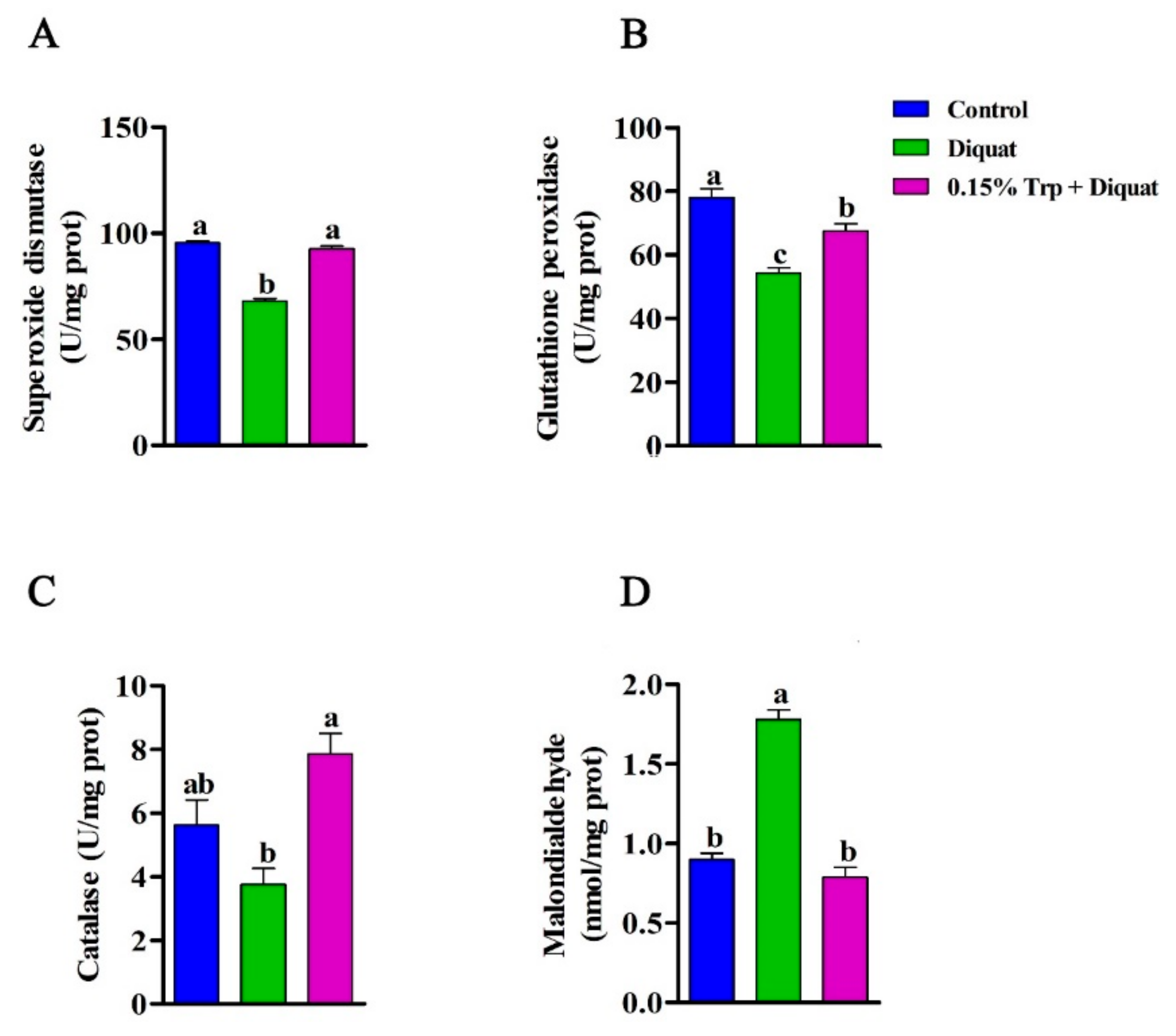
3.4. Antioxidant Enzymes Activities and MDA Content in the Jejunum
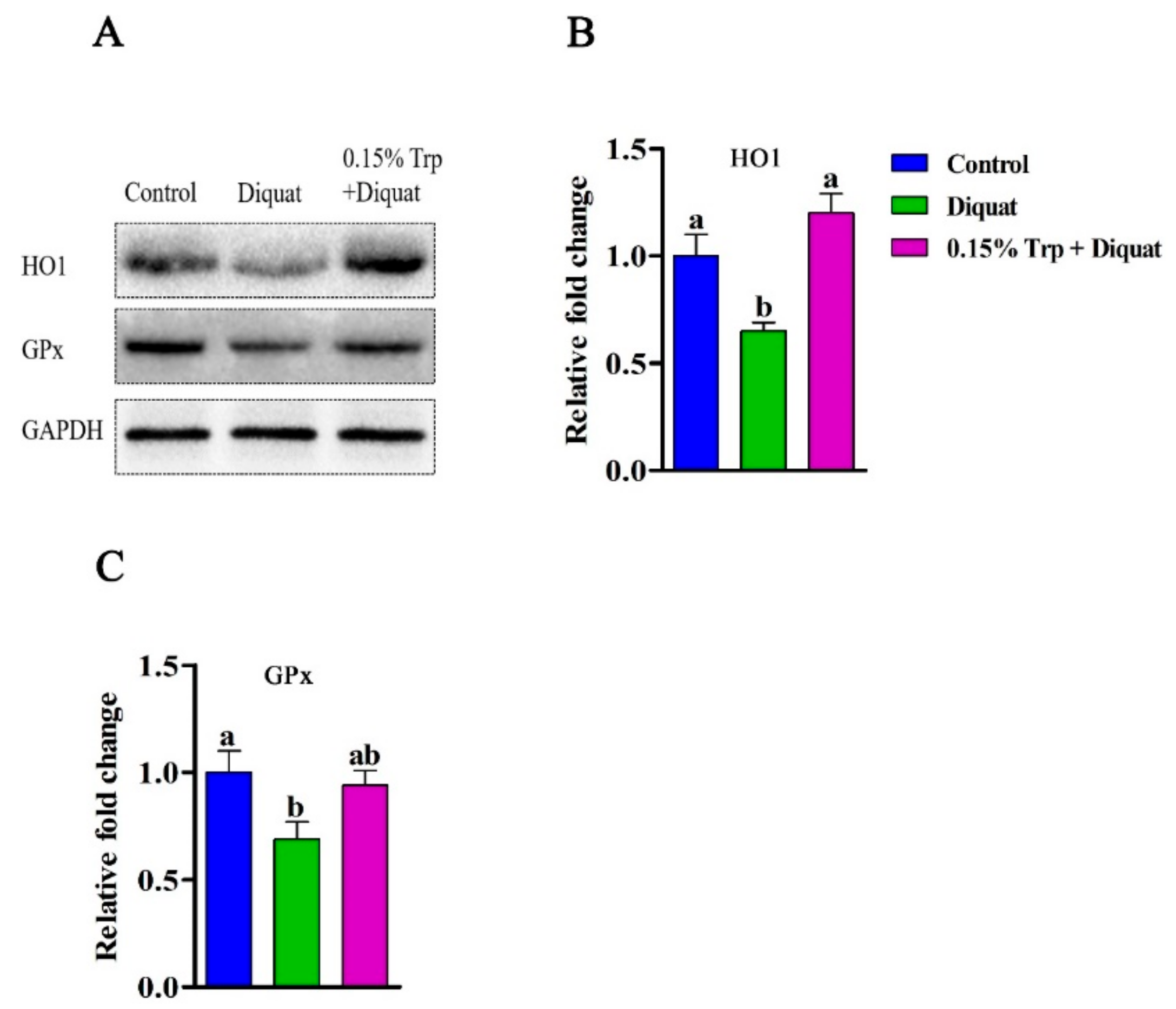
3.5. Redox-Sensitive Genes Expression in the Jejunum
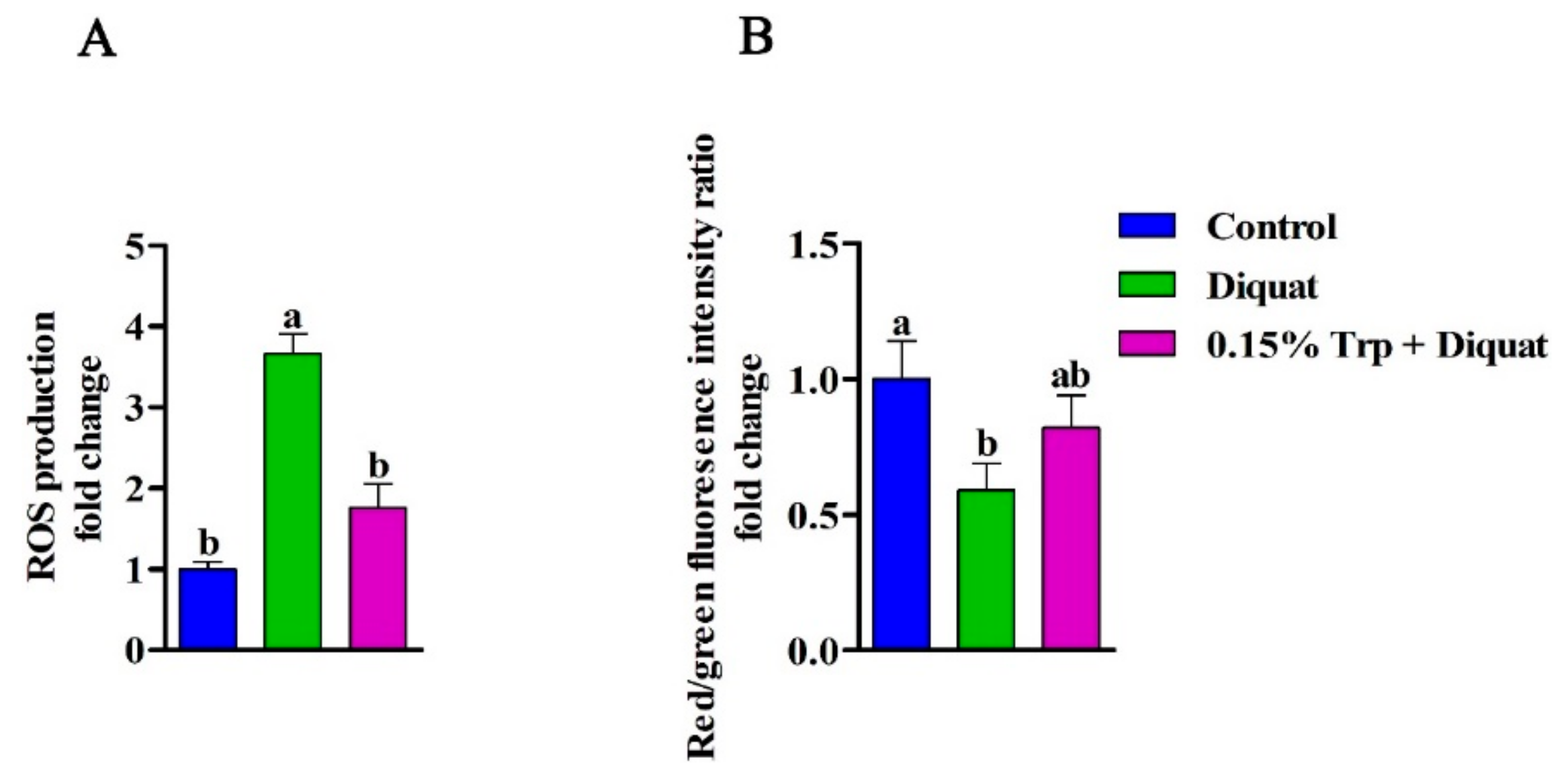
3.6. Intestinal Mitochondrial ROS Production and ΔΨm
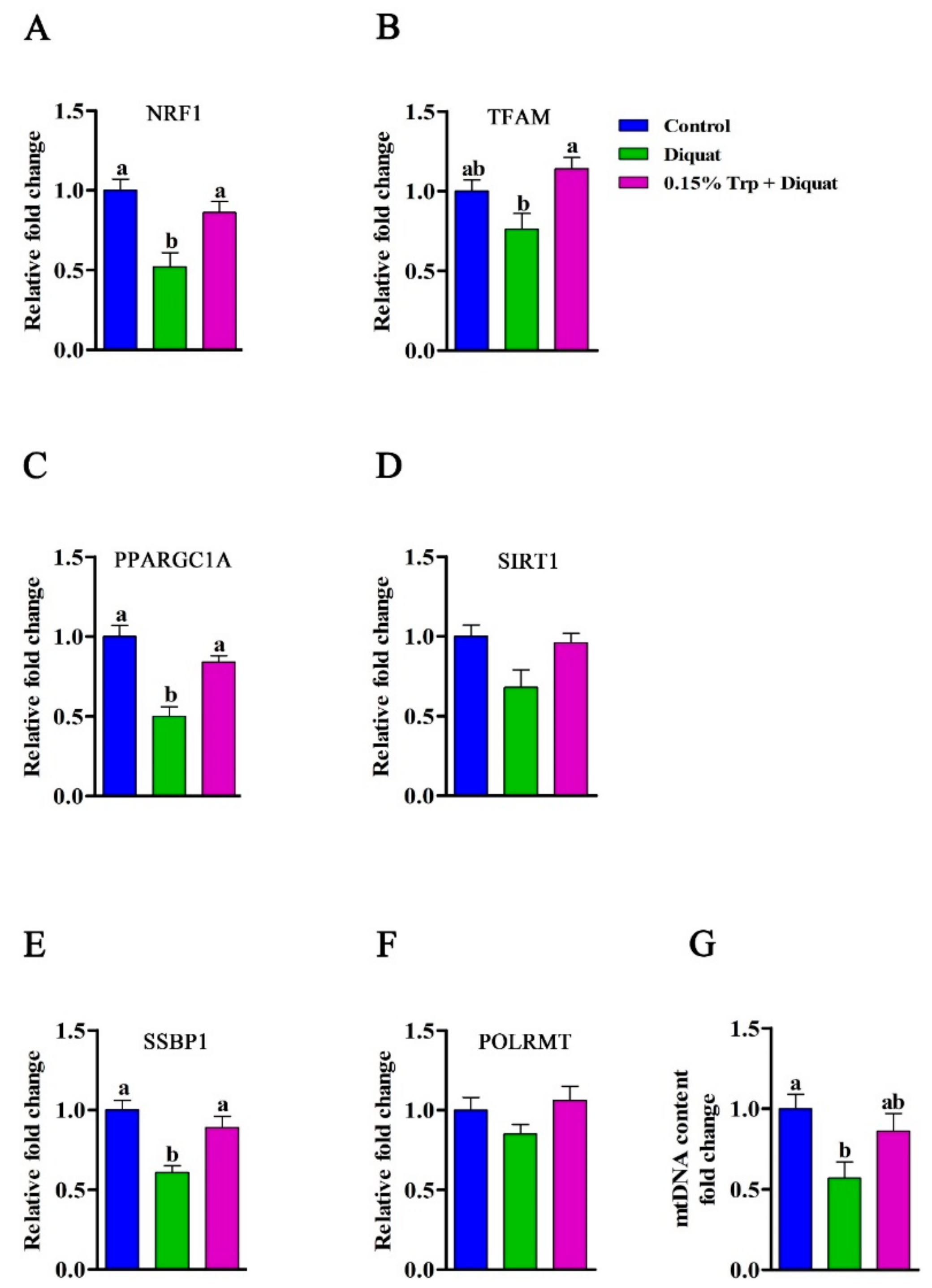
3.7. Expression of Genes Related to Mitochondrial Biogenesis in the Jejunum
3.8. mtDNA Content in the Jejunum
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Campbell, J.M.; Crenshaw, J.D. The biological stress of early-weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [PubMed]
- Pluske, J.R.; Hampson, D.J. Factors influencing the structure and function of the small intestine in the weaned pig: A review. Livest. Prod. Sci. 1997, 51, 215–236. [Google Scholar] [CrossRef]
- Smith, F.; Clark, J.E. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, 352–363. [Google Scholar]
- Shen, Y.B.; Voilqué, G. Effects of increasing tryptophan intake on growth and physiological changes in nursery pigs. J. Anim. Sci. 2012, 90, 2264–2275. [Google Scholar] [CrossRef]
- Liang, H.W.; Dai, Z.L. Dietary L-tryptophan supplementation enhances the intestinal mucosal barrier function in weaned piglets: Implication of tryptophan-metabolizing microbiota. Int. J. Mol. Sci. 2019, 20, 20. [Google Scholar] [CrossRef]
- Trevisi, P.; Melchior, D. A tryptophan-enriched diet improves feed intake and growth performance of susceptible weanling pigs orally challenged with Escherichia coli K881. J. Anim. Sci. 2009, 87, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Koopmans, S.J.; Guzik, A.C. Effects of supplemental L-tryptophan on serotonin, cortisol, intestinal integrity, and behavior in weanling piglets. J. Anim. Sci. 2006, 84, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Jiang, M. Tryptophan increases intestinal permeability and decreases intestinal tight junction protein expression in weanling piglets. J. Anim. Sci. 2016, 94, 87–90. [Google Scholar]
- Tossou, M.C.B.; Liu, H.; Bai, M.; Chen, S.; Cai, Y.; Duraipandiyan, V.; Liu, H.; Adebowale, T.O.; Al-Dhabi, N.A.; Long, L.; et al. Effect of high dietary tryptophan on intestinal morphology and tight junction protein of weaned pig. Biomed. Res. Int. 2016, 2016, 2912418. [Google Scholar] [CrossRef]
- Cao, S.T.; Wang, C.C. Weaning disrupts intestinal antioxidant status, impairs intestinal barrier and mitochondrial function, and triggers mitophagy in piglets. J. Anim. Sci. 2018, 96, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Wu, M.M. Development of an antioxidant system after early weaning in piglets. J. Anim. Sci. 2014, 92, 612–619. [Google Scholar] [CrossRef]
- Mao, X.B.; Lv, M. The effect of dietary tryptophan levels on oxidative stress of liver induced by diquat in weaned piglets. J. Anim. Sci. Biotechnol. 2014, 5, 49. [Google Scholar] [CrossRef]
- Cao, S.T.; Wu, H. Diquat-induced oxidative stress increases intestinal permeability, impairs mitochondrial function, and triggers mitophagy in piglets. J. Anim. Sci. 2018, 96, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Dixit, P.; Jalin, D.K. Differential effect of oxidative stress on intestinal apparent permeability of drugs transported by paracellular and transcellular route. Eur. J. Drug Metab. Pharmacokinet. 2012, 37, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Miyazono, Y. Oxidative stress and enhanced paracellular permeability in the small intestine of methotrexate-treated rats. Cancer Chemother. Pharmacol. 2010, 65, 1117–1123. [Google Scholar] [CrossRef]
- Marcu, R.; Zheng, Y. Mitochondria and angiogenesis. Adv. Exp. Med. Biol. 2017, 982, 371–406. [Google Scholar] [PubMed]
- Jeong, E.M.; Chung, J. Role of mitochondrial oxidative stress in glucose tolerance, insulin resistance, and cardiac diastolic dysfunction. J. Am. Heart Assoc. 2016, 5, 173. [Google Scholar] [CrossRef] [PubMed]
- Thummasorn, S.; Kumfu, S. Granulocyte-colony stimulating factor attenuates mitochondrial dysfunction induced by oxidative stress in cardiac mitochondria. Mitochondrion 2011, 11, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Janssenduijghuijsen, L.; Grefte, S. Mitochondrial ATP production and intestinal epithelial permeability—An in vitro model. BBA Bioenerg. 2016, 1857, e119. [Google Scholar] [CrossRef]
- Liu, J.B.; Yan, H.L. The response of performance in grower and finisher pigs to diets formulated to different tryptophan to lysine ratios. Livest. Sci. 2019, 222, 25–30. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012; 400p. [Google Scholar]
- Wang, J.; Chen, L. Gene expression is altered in piglet small intestine by weaning and dietary glutamine supplementation. J. Nutr. 2008, 138, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.Q.; Yao, K. Effects of a-ketoglutarate on energy status in the intestinal mucosa of weaned piglets chronically challenged with lipopolysaccharide. Br. J. Nutr. 2011, 106, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.H.; Yan, H.L. Mycotoxin binder improves growth rate in piglets associated with reduction of toll-like receptor-4 and increase of tight junction protein gene expression in gut mucosa. J. Anim. Sci. Biotechnol. 2017, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.B.; Yan, H.L. Effects of dietary energy and protein content and lipid source on growth performance and carcass traits in Pekin ducks. Poult. Sci. 2019, in press. [Google Scholar] [CrossRef]
- Huang, Q.; Xu, W. Protective effects of leucine on redox status and mitochondrial-related gene abundance in the jejunum of intrauterine growth-retarded piglets during early weaning period. Arch. Anim. Nutr. 2017, 71, 93–107. [Google Scholar] [CrossRef]
- Yan, H.L.; Liu, J.B. Production phase affects the bioaerosol microbial composition and functional potential in swine confinement buildings. Animals 2019, 9, 90. [Google Scholar] [CrossRef]
- Zhuo, Y.; Hua, L. Fibroblast growth factor 21 coordinates adiponectin to mediate the beneficial effects of low-protein diet on primordial follicle reserve. EBioMedicine 2019, 41, 623–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.L.; Cao, S.C. Reduced meal frequency alleviates high-fat diet-induced lipid accumulation and inflammation in adipose tissue of pigs under the circumstance of fixed feed allowance. Eur. J. Nutr. 2019, in press. [Google Scholar] [CrossRef]
- Zhang, H.W.; Yin, J.D. Tryptophan enhances ghrelin expression and secretion associated with increased food intake and weight gain in weanling pigs. Domest. Anim. Endocrinol. 2007, 33, 47–61. [Google Scholar] [CrossRef]
- Liang, H.; Dai, Z. Dietary L-tryptophan modulates the structural and functional composition of the intestinal microbiome in weaned piglets. Front. Microbiol. 2018, 9, 1736. [Google Scholar] [CrossRef] [PubMed]
- Capozzalo, M.M.; Kim, J.C. An increased ratio of dietary tryptophan to lysine improves feed efficiency and elevates plasma tryptophan and kynurenine in the absence of antimicrobials and regardless of infection with enterotoxigenic Escherichia coli in weaned pigs. J. Anim. Sci. 2012, 90, 191–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simongiovanni, A.; Corrent, E. Estimation of the tryptophan requirement in piglets by meta-analysis. Animal 2012, 6, 594–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, L.T.; Xiao, Y.P. Molecular characterization of a novel ovodefensin gene in chickens. Gene 2018, 678, 233–240. [Google Scholar]
- Nieto, N.; Torres, M.I. Experimental ulcerative colitis impairs antioxidant defense system in rat intestine. Digest. Dis. Sci. 2000, 45, 1820–1827. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Zheng, P. Benzoic acid beneficially affects growth performance of weaned pigs which was associated with changes in gut bacterial populations, morphology indices and growth factor gene expression. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1137–1146. [Google Scholar] [CrossRef]
- Cao, S.T.; Shen, Z.J. Resveratrol improves intestinal barrier function, alleviates mitochondrial dysfunction and induces mitophagy in diquat challenged piglets. Food Funct. 2019, 10, 344. [Google Scholar] [CrossRef]
- Boudry, G. The Ussing chamber technique to evaluate alternatives to in-feed antibiotics for young pigs. Anim. Res. 2005, 54, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Ji, Y. L-tryptophan activates mammalian target of rapamycin and enhances expression of tight junction proteins in intestinal porcine epithelial cells. J. Nutr. 2015, 145, 1156–1162. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Van Itellie, C.M. Tight junctions and the molecular basis for regulation of paracellular permeability. Am. J. Physiol. 1995, 269, 467–475. [Google Scholar] [CrossRef]
- Wong, V.; Gumbiner, B.M. A synthetic peptide corresponding to the extracellular domain of occludin perturbs the tight junction permeability barrier. J. Cell Biol. 1997, 136, 399–409. [Google Scholar] [CrossRef]
- Wang, W.; Degroote, J. Effects of intrauterine growth restriction on the intestinal barrier function, redox status and redox-sensitive genes expression during the neonatal period of piglets. FASEB J. 2016, 30, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Messori, S.; Trevisi, P. Effect of susceptibility to enterotoxigenic Escherichia coli F4 and of dietary tryptophan on gut microbiota diversity observed in healthy young pigs. Vet. Microbiol. 2013, 162, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Natividad, J.M.; Agus, A. Impaired aryl hydrocarbon receptor ligand production by the gut microbiota is a key factor in metabolic syndrome. Cell Metab. 2018, 28, 737–742. [Google Scholar] [CrossRef]
- Blasig, I.E.; Bellmann, C. Occludin protein family: Oxidative stress and reducing conditions. Antioxid. Redox Signal. 2011, 15, 1195–1219. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.K.; Rueckert, C. The oligomerization of the coiled coil-domain of occludin is redox sensitive. Ann. N. Y. Acad. Sci. 2009, 1165, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.B.; Xue, P.C. Effects of dietary phosphorus concentration and body weight on postileal phosphorus digestion in pigs. Anim. Feed Sci. Technol. 2018, 242, 86–94. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Song, F. Paeonol and danshensu combination attenuates apoptosis in myocardial infarcted rats by inhibiting oxidative stress: Roles of Nrf2/HO-1 and PI3K/Akt pathway. Sci. Rep. 2016, 6, 23693. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, Y. Medium-chain TAG attenuate hepatic oxidative damage in intra-uterine growth-retarded weanling piglets by improving the metabolic efficiency of the glutathione redox cycle. Br. J. Nutr. 2014, 112, 876–885. [Google Scholar] [CrossRef] [Green Version]
- Christen, S.; Peterhans, E. Antioxidant activities of some tryptophan metabolites: Possible implication for inflammatory diseases. Proc. Natl. Acad. Sci. USA 1990, 87, 2506–2510. [Google Scholar] [CrossRef] [PubMed]
- Sastre, J.; Pallardó, F.V. Mitochondrial oxidative stress plays a key role in aging and apoptosis. IUBMB Life 2000, 49, 427–435. [Google Scholar] [PubMed]
- Zhang, X.N.; Yan, H.J. Cerebral ischemia-reperfusion-induced autophagy protects against neuronal injury by mitochondrial clearance. Autophagy 2013, 9, 1321–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, N.; Wu, Z. L-glutamate enhances barrier and antioxidative functions in intestinal porcine epithelial cells. J. Nutr. 2015, 145, 2258–2264. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Basal Diet | Trp1 Diet 3 | Trp2 Diet 3 |
|---|---|---|---|
| Ingredients composition, % | |||
| Corn | 30.83 | 30.83 | 30.83 |
| Extruded corn | 24.00 | 24.00 | 24.00 |
| Extruded soybean | 6.00 | 6.00 | 6.00 |
| Dehulled soybean meal (44% crude protein) | 10.00 | 10.00 | 10.00 |
| Soybean protein concentrate | 10.00 | 10.00 | 10.00 |
| Whey powder | 5.00 | 5.00 | 5.00 |
| Fish meal | 5.00 | 5.00 | 5.00 |
| Soybean oil | 1.80 | 1.80 | 1.80 |
| Sucrose | 3.00 | 3.00 | 3.00 |
| Glucose | 1.69 | 1.69 | 1.69 |
| Cornstarch | 0.039 | 0.019 | 0 |
| Salt | 0.30 | 0.30 | 0.30 |
| Limestone | 0.72 | 0.72 | 0.72 |
| CaHPO4 | 0.52 | 0.52 | 0.52 |
| Chloride choline | 0.10 | 0.10 | 0.10 |
| Vitamin premix 1 | 0.05 | 0.05 | 0.05 |
| Mineral premix 2 | 0.30 | 0.30 | 0.30 |
| L-Trptophan (98.0%) | 0 | 0.153 | 0.306 |
| L-Alanine (99.8%) | 0.271 | 0.138 | 0.004 |
| DL-Methionine (98.5%) | 0.13 | 0.13 | 0.13 |
| L-Lysine·HCl (78%) | 0.18 | 0.18 | 0.18 |
| L-Threonine (98.5%) | 0.07 | 0.07 | 0.07 |
| Total | 100.00 | 100.00 | 100.00 |
| Nutrient level (Calculated value) | |||
| DE/(MJ/kg) | 15.04 | 15.04 | 15.04 |
| Crude protein (%) | 20.88 | 20.88 | 20.88 |
| Ca (%) | 0.80 | 0.80 | 0.80 |
| Available phosphorous (%) | 0.41 | 0.41 | 0.41 |
| SID Lys (%) | 1.35 | 1.35 | 1.35 |
| SID Met (%) | 0.46 | 0.46 | 0.46 |
| SID Thr (%) | 0.80 | 0.80 | 0.80 |
| SID Trp (%) | 0.22 | 0.37 | 0.52 |
| Gene Symbols | Nucleotide Sequence (5′-3′) | Accession No. |
|---|---|---|
| ACTB | F: CCCCTCCTCTCTTGCCTCTC R: AAAAGTCCTAGGAAAATGGCAGAAG P: (FAM) TGCCACGCCCTTTCTCACTTGTTCT (Eclipse) | DQ452569 |
| mt D-loop | F: GATCGTACATAGCACATATCATGTC R: GGTCCTGAAGTAAGAACCAGATG P: (FAM) CCAGTCAACATGCGTATCACCACCA (Eclipse) | AF276923 |
| Gene Symbols | Nucleotide Sequence of Primers (5′–3′) | Accession No. |
|---|---|---|
| ACTB | F: TCTGGCACCACACCTTCT R: TGATCTGGGTCATCTTCTCAC | XM_003124280.3 |
| TOP2B | F: AACTGGATGATGCTAATGATGCT R: TGGAAAAACTCCGTATCTGTCTC | NM_001258386.1 |
| TBP | F: GATGGACGTTCGGTTTAGG R: AGCAGCACAGTACGAGCAA | DQ178129 |
| ZO1 | F: ATCTCGGAAAAGTGCCAGGA R: CCCCTCAGAAACCCATACCA | XM_003480423.3 |
| ZO2 | F: CCAGGAAGCACAGAATGCAA R: AAGTCTGGCGGGACCTCTCT | XM_005660148.2 |
| OCLN | F: CATGGCTGCCTTCTGCTTCATTGC R: ACCATCACACCCAGGATAGCACTCA | NM_001163647.2 |
| CLDN1 | F: TATGACCCCATGACCCCAGT R: GCAGCAAAGTAGGGCACCTC | NM_001244539.1 |
| CLDN2 | F: TTCCTCCCTGTTCTCCCTGA R: CACTCTTGGCTTTGGGTGGT | NM_001161638.1 |
| SOD1 | F: GGTCCTCACTTCAATCCTGAATCC R: CACACCATCTTTGCCAGCAGT | NM_0011190422 |
| HMOX1 | F: CGCTCCCGAATGAACACTCT R: GCGAGGGTCTCTGGTCCTTA | NM_001004027.1 |
| GPX1 | F: TGCTCATTGAGAACGTAGCGT R CAGGATCTCCCCATTCTTCGC | NM_214201.1 |
| TXNRD | F: GTGCTGAGGAGCTTCCCGAGATGT R: TCCAGGACCATGACCCGCTTGTTAA | NM_214154.3 |
| NRF2 | F: CCCATTGAGGGCTGTGATCT R: GCCTTCAGTGTGCTTCTGGTT | NM_031789.2 |
| KEAP1 | F: GGCTGGGATGCCTTGTAAAG R: GGGCCCATGGATTTCAGTT | NM_057152.2 |
| NRF1 | F: GCCAGTGAGATGAAGAGAAACG R: CTACAGCAGGGACCAAAGTTCAC | AK237171.1 |
| TFAM | F: GGTCCATCACAGGTAAAGCTGAA R: ATAAGATCGTTTCGCCCAACTTC | AY923074.1 |
| PPARGC1A | F: CCCGAAACAGTAGCAGAGACAAG R: CTGGGGTCAGAGGAAGAGATAAAG | NM 213963 |
| SIRT1 | F: TGACTGTGAAGCTGTACGAGGAG R: TGGCTCTATGAAACTGCTCTGG | EU030283.2 |
| SSBP1 | F: CTTTGAGGTAGTGCTGTGTCG R: CTCACCCCTGACGATGAAGAC | AK352341.1 |
| POLRMT | F: CTTTGAGGTTTTCCAGCAGCAG R: GCTCCCAGTTTTGGTTGACAG | XM 001927064.1 |
| Items | Control | Diquat | 0.15% Trp + Diquat | 0.30% Trp + Diquat |
|---|---|---|---|---|
| Body weight (kg) | ||||
| Day 0 | 6.62 ± 0.20 | 6.63 ± 0.21 | 6.62 ± 0.21 | 6.63 ± 0.22 |
| Day 7 | 8.42 ± 0.22 | 8.43 ± 0.20 | 8.66 ± 0.23 | 8.60 ± 0.22 |
| Day 21 | 14.01 ± 0.28 a | 12.11 ± 0.47 b | 13.80 ± 0.49 a | 12.64 ± 0.41 b |
| Average daily weight gain (g) | ||||
| Day 0 to 7 | 256.43 ± 7.10 | 258.22 ± 11.50 | 290.89 ± 5.35 | 281.79 ± 14.90 |
| Day 7 to 21 | 399.64 ± 8.86 a | 262.50 ± 20.5 b | 366.79 ± 19.22a | 288.84 ± 22.36 b |
| Average daily feed intake (g) | ||||
| Day 0 to 7 | 313.42 ± 8.12 b | 311.75 ± 7.87 b | 331.97 ± 5.23 a | 320.45 ± 9.50 b |
| Day 7 to 21 | 566.37 ± 12.53 a | 462.47 ± 14.90c | 560.30 ± 11.18 a | 500.78 ± 11.70 b |
| Feed to gain ratio | ||||
| Day 0 to 7 | 1.23 ± 0.04 | 1.23 ± 0.07 | 1.14 ± 0.02 | 1.16 ± 0.08 |
| Day 7 to 21 | 1.42 ± 0.04 b | 1.81 ± 0.10 a | 1.55 ± 0.06 ab | 1.78 ± 0.11 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhang, Y.; Li, Y.; Yan, H.; Zhang, H. L-Tryptophan Enhances Intestinal Integrity in Diquat-Challenged Piglets Associated with Improvement of Redox Status and Mitochondrial Function. Animals 2019, 9, 266. https://doi.org/10.3390/ani9050266
Liu J, Zhang Y, Li Y, Yan H, Zhang H. L-Tryptophan Enhances Intestinal Integrity in Diquat-Challenged Piglets Associated with Improvement of Redox Status and Mitochondrial Function. Animals. 2019; 9(5):266. https://doi.org/10.3390/ani9050266
Chicago/Turabian StyleLiu, Jingbo, Yong Zhang, Yan Li, Honglin Yan, and Hongfu Zhang. 2019. "L-Tryptophan Enhances Intestinal Integrity in Diquat-Challenged Piglets Associated with Improvement of Redox Status and Mitochondrial Function" Animals 9, no. 5: 266. https://doi.org/10.3390/ani9050266