
Effects of Linseed Supplementation on Milk Production, Composition, Odd- and Branched-Chain Fatty Acids, and on Serum Biochemistry in Cilentana Grazing Goats

 ,
,  ,
,  ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Management
2.2. Milk Collection and Analysis
2.3. Blood Analysis
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAO). Food and Agriculture Organization of the United Nations Statistical Databases. 2019; (accessed on 16 February 2019). [Google Scholar]
- de Asís Ruiz Morales, F.; Castel Genís, J.M.; Mena Guerrero, Y. Current status, challenges and the way forward for dairy goat production in Europe. Asian-Australas. J. Anim. Sci. 2019, 32, 1256–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, B.A.; Lu, C.D. Current status of global dairy goat production: An overview. Asian-Australas. J. Anim. Sci. 2019, 32, 1219–1232. [Google Scholar] [CrossRef] [PubMed]
- Sepe, L.; Argüello, A. Recent advances in dairy goat products. Asian-Australas. J. Anim. Sci. 2019, 32, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Vlaeminck, B.; Fievez, V.; Tamminga, S.; Dewhurst, R.J.; van Vuuren, A.; De Brabander, D.; Demeyer, D. Milk odd- and branched-chain fatty acids in relation to the rumen fermentation pattern. J. Dairy Sci. 2006, 89, 3954–3964. [Google Scholar] [CrossRef] [Green Version]
- Fievez, V.; Colman, E.; Castro-Montoya, J.M.; Stefanov, I.; Vlaeminck, B. Milk odd- and branched-chain fatty acids as biomarkers of rumen function—An update. Anim. Feed Sci. Technol. 2012, 172, 51–65. [Google Scholar] [CrossRef]
- Serment, A.; Schmidely, P.; Giger-Reverdin, S.; Chapoutot, P.; Sauvant, D. Effects of the percentage of concentrate on rumen fermentation, nutrient digestibility, plasma metabolites, and milk composition in mid-lactation goats. J. Dairy Sci. 2011, 94, 3960–3972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, L.; Shingfield, K.J.; Rouel, J.; Ferlay, A.; Chilliard, Y. Effect of plant oils in the diet on performance and milk fatty acid composition in goats fed diets based on grass hay or maize silage. Br. J. Nutr. 2009, 101, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mele, M.; Serra, A.; Buccioni, A.; Conte, G.; Pollicardo, A.; Secchiari, P. Effect of soybean oil supplementation on milk fatty acid composition from Saanen goats fed diets with different forage: Concentrate ratios. Ital. J. Anim. Sci. 2008, 7, 297–311. [Google Scholar] [CrossRef]
- Li, F.; Li, Z.; Li, S.; Ferguson, J.D.; Cao, Y.; Yao, J.; Sun, F.; Wang, X.; Yang, T. Effect of dietary physically effective fiber on ruminal fermentation and the fatty acid profile of milk in dairy goats. J. Dairy Sci. 2014, 97, 2281–2290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todaro, M.; Scatassa, M.L.; Giaccone, P. Multivariate factor analysis of Girgentana goat milk composition. Ital. J. Anim. Sci. 2005, 4, 403–410. [Google Scholar] [CrossRef] [Green Version]
- Infascelli, L.; Tudisco, R.; Iommelli, P.; Capitanio, F. Milk quality and animal welfare as a possible marketing lever for the economic development of rural areas in southern Italy. Animals 2021, 11, 1059. [Google Scholar] [CrossRef] [PubMed]
- Palmquist, D.L.; Jenkins, T.C. Fat in lactation rations: Review. J. Dairy Sci. 1980, 63, 1–14. [Google Scholar] [CrossRef]
- Ferguson, J.D.; Sklan, D.; Chalupa, W.V.; Kronfeld, D.S. Effects of hard fats on in vitro and in vivo rumen fermentation, milk production, and reproduction in dairy cows. J. Dairy Sci. 1990, 73, 2864–2879. [Google Scholar] [CrossRef]
- Davidson, L.A.; Wang, N.; Shah, M.S.; Lupton, J.R.; Ivanov, I.; Chapkin, R.S. n-3 Polyunsaturated fatty acids modulate carcinogen-directed non-coding microRNA signatures in rat colon. Carcinogenesis 2009, 30, 2077–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and non starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3598. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 19th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2012. [Google Scholar]
- INRA. Alimentation des Ruminants; Jarrige, R., Ed.; INRA Editions: Versailles, France, 1978. [Google Scholar]
- Castro, T.; Manso, T.; Jimeno, V.; Del Alamo, M.; Mantecón, A.R. Effects of dietary sources of vegetable fats on performance of dairy ewes and conjugated linoleic acid (CLA) in milk. Small Rumin. Res. 2009, 84, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Tudisco, R.; Morittu, V.M.; Addi, L.; Moniello, G.; Grossi, M.; Musco, N.; Grazioli, R.; Mastellone, V.; Pero, M.E.; Lombardi, P.; et al. Influence of pasture on stearoyl-coa desaturase and mirna 103 expression in goat milk: Preliminary results. Animals 2019, 9, 606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, A.; Radin, N.S. Lipid extraction of tissues with a low-toxicity solvent. Annal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Christie, W.W. Preparation of ester derivatives of fatty acids for chromatographic analysis. In Advances in Lipid Methodology-two; Christie, W.W., Ed.; Oily Press: Dundee, UK, 1993; pp. 69–111. [Google Scholar]
- Nudda, A.; Battacone, G.; Atzori, A.S.; Dimauro, C.; Rassu, S.P.G.; Nicolussi, P.; Bonelli, P.; Pulina, G. Effect of extruded linseed supplementation on blood metabolic profile and milk performance of Saanen goats. Animal 2013, 7, 1464–1471. [Google Scholar] [CrossRef]
- Tudisco, R.; Chiofalo, B.; Lo Presti, V.; Morittu, V.M.; Moniello, G.; Grossi, M.; Musco, N.; Grazioli, R.; Mastellone, V.; Lombardi, P.; et al. Influence of feeding linseed on SCD activity in grazing goat mammary glands. Animals 2019, 9, 786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuentes, M.C.; Calsamiglia, S.; Sánchez, C.; González, A.; Newbold, J.R.; Santos, J.E.P.; Rodríguez-Alcalá, L.M.; Fontecha, J. Effect of extruded linseed on productive and reproductive performance of lactating dairy cows. Livest. Sci. 2008, 113, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Ariza, J.M.; Meignan, T.; Madouasse, A.; Beaudeau, F.; Bareille, N. Effects on milk quantity and composition associated with extruded linseed supplementation to dairy cow diets. Sci. Rep. 2019, 9, 17563. [Google Scholar] [CrossRef] [PubMed]
- Mughetti, L.; Sinesio, F.; Acuti, G.; Antonini, C.; Moneta, E.; Peparaio, M.; Trabalza-Marinucci, M. Integration of extruded linseed into dairy sheep diets: Effects on milk composition and quality and sensorial properties of Pecorino cheese. Anim. Feed Sci. Technol. 2012, 178, 27–39. [Google Scholar] [CrossRef]
- Bernard, L.; Bonnet, M.; Leroux, C.; Shingfield, K.J.; Chilliard, Y. Effect of sunflower-seed oil and linseed oil on tissue lipid metabolism, gene expression, and milk fatty acid secretion in alpine goats fed maizesilage-based diets. J. Dairy Sci. 2009, 92, 6083–6094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chilliard, Y.; Ferlay, A.; Rouel, J.; Lamberet, G. A review of nutritional and physiological factors affecting goat milk lipid synthesis and lipolysis. J. Dairy Sci. 2003, 86, 1751–1770. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Cortés, P.; Cívico, A.; De la Fuente, M.; Núñez Sánchez, N.; Peña Blanco, F.; Martínez Marín, A. Effects of dietary concentrate composition and linseed oil supplementation on the milk fatty acid profile of goats. Animal 2018, 12, 2310–2317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renna, M.; Lussiana, C.; D’Agostino, M.; Mimosi, A.; Fortina, R. Extruded linseed supplementation in dairy goat diet: Effects on productive performance and fatty acid profile of bulk milk, fresh and ripened cheese. J. Anim. Vet. Adv. 2013, 12, 1550–1564. [Google Scholar]
- Loor, J.J.; Ferlay, A.; Ollier, A.; Doreau, M.; Chilliard, Y. Relationship among trans and conjugated fatty acids and bovine milk fat yield due to dietary concentrate and linseed oil. J. Dairy Sci. 2005, 88, 726–740. [Google Scholar] [CrossRef] [Green Version]
- Almeida, O.C.; Ferraz, M.V.C.; Susin, I.; Gentil, R.S.; Polizel, D.M.; Ferreira, E.M.; Barroso, J.P.R.; Pires, A.V. Plasma and milk fatty acid profiles in goats fed diets supplemented with oils from soybean, linseed or fish. Small Rumin. Res. 2019, 170, 125–130. [Google Scholar] [CrossRef]
- Abuelfatah, K.; Zuki, A.B.; Goh, Y.; Sazili, A.Q.; Abubakr, A. Effects of feeding whole linseed on ruminal fatty acid composition and microbial population in goats. Anim. Nutr. 2016, 2, 323–328. [Google Scholar] [CrossRef]
- Chilliard, Y.; Martin, C.; Rouel, J.; Doreau, M. Milk fatty acids in dairy cows fed whole crude linseed, extruded linseed or linseed oil, and their relationship with methane output. J. Dairy Sci. 2009, 92, 5199–5211. [Google Scholar] [CrossRef] [PubMed]
- Dhaka, V.; Gulia, N.; Ahlawat, K.S.; Khatkar, B.S. Trans fats-sources, health risks and alternative approach—A review. J. Food Sci. Technol. 2011, 48, 534–541. [Google Scholar] [CrossRef] [Green Version]
- Sarrazin, P.; Mustafa, A.F.; Chouinard, P.Y.; Raghavan, G.; Sotocinal, S. Performance of dairy cows fed roasted sunflower seed. J. Sci. Food Agric. 2004, 84, 1179–1185. [Google Scholar] [CrossRef]
- Gonthier, C.; Mustafa, A.F.; Ouellet, D.R.; Chouinard, P.Y.; Berthiaume, R.; Petit, H.V. Feeding micronized and extruded flaxseed to dairy cows: Effects on blood parameters and milk fatty acid composition. J. Dairy Sci. 2005, 88, 748–756. [Google Scholar] [CrossRef] [Green Version]
- Chilliard, Y.; Doreau, M.; Gagliostro, G.; Elmeddah, Y. Addition de lipides protégés (encapsulés ou savons de calcium) à la ration de vaches laitières. Effets sur les performances et la composition du lait (Protected (encapsulated or calcium salts) lipids in dairy cow diets. Effects on production and milk composition). INRA Prod. Anim. 1993, 6, 139–150. (In French) [Google Scholar]
- Michalet-Doreau, B.; Martin, C.; Doreau, M. Optimisation de la digestion des parois végétales dans le rumen: Quantification des ineractions digestives (Optimisation of fibre ruminal digestion: Interactions of fibre digestion with other components). Renc. Rech. Rum. 1997, 4, 103–112. (In French) [Google Scholar]
- Dilzer, A.; Park, Y. Implication of conjugated linoleic acid (CLA) in human health, critical reviews. Food Sci. Nutr. 2012, 52, 488–513. [Google Scholar]
- Griinari, J.M.; Corl, B.A.; Lacy, S.H.; Chouinard, P.Y.; Nurmela, K.V.; Bauman, D.E. Conjugated linoleic acid is synthesized endogenously in lactating dairy cows by Delta (9)-desaturase. J. Nutr. 2000, 130, 2285–2291. [Google Scholar] [CrossRef] [PubMed]
- Tudisco, R.; Calabrò, S.; Cutrignelli, M.I.; Moniello, G.; Grossi, M.; Gonzalez, O.J.; Piccolo, V.; Infascelli, F. Influence of organic systems on Stearoyl-CoA desaturase in goat milk. Small Rumin. Res. 2012, 106, 37–42. [Google Scholar] [CrossRef]
- Bu, D.P.; Wang, J.Q.; Dhiman, T.R.; Liu, J. Effectiveness of oils rich in linoleic and linolenic acids to enhance conjugated linoleic acid in milk from dairy cows. J. Dairy Sci. 2007, 90, 998–1007. [Google Scholar] [CrossRef]
- Ferlay, A.; Doreau, M.; Martin, C.; Chilliard, Y. Effects of incremental amounts of extruded linseed on the milk fatty acid composition of dairy cows receiving hay or corn silage. J. Dairy Sci. 2013, 96, 6577–6595. [Google Scholar] [CrossRef]
- Gomez-Cortes, P.; Bach, A.; Luna, P.; Juarez, M.; de la Fuente, M.A. Effects of extruded linseed supplementation on n-3 fatty acids and conjugated linoleic acid in milk and cheese from ewes. J. Dairy Sci. 2009, 92, 4122–4134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kholif, S.M.; Morsy, T.A.; Abedo, A.A.; El-Bordeny, N.; Abdo, M.M. Milk production and composition, milk fatty acid profile, nutrients digestibility and blood composition of dairy buffaloes fed crushed flaxseed in early lactation. Egypt. J. Nutr. Feeds 2011, 14, 385–394. [Google Scholar]
- Roy, A.; Ferlay, A.; Shingfield, K.J.; Chilliard, Y. Examination of the persistency of milk fatty acid composition responses to plant oils in cows given different basal diets, with particular emphasis on trans-C18:1 fatty acids and isomers of conjugated linoleic acid. Anim. Sci. 2006, 82, 479–492. [Google Scholar] [CrossRef]
- Vlaeminck, B.; Fievez, V.; Cabrita, A.R.J.; Fonseca, A.J.M.; Dewhurst, R.J. Factors affecting odd- and branched-chain fatty acids in milk: A review. Anim. Feed Sci. Technol. 2006, 131, 389–417. [Google Scholar] [CrossRef]
- Maia, M.R.G.; Chaudhary, L.C.; Figueres, L.; Wallace, R.J. Metabolism of polyunsaturated fatty acids and their toxicity to the microflora of the rumen. Antonie Van Leeuwenhoek 2007, 91, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Cívico, A.; Núñez Sánchez, N.; Gómez-Cortés, P.; de la Fuente, M.A.; Peña Blanco, F.; Juárez, M.; Schiavone, A.; Martínez Marín, A.L. Odd- and branched-chain fatty acids in goat milk as indicators of the diet composition. Ital. J. Anim. Sci. 2017, 16, 68–74. [Google Scholar] [CrossRef]
- Yang, S.L.; Bu, D.P.; Wang, J.Q.; Hu, Z.Y.; Li, D.; Wei, H.Y.; Zhou, L.Y.; Loor, J.J. Soybean oil and linseed oil supplementation affect profiles of ruminal microorganisms in dairy cows. Animal 2009, 3, 1562–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collomb, M.; Sieber, R.; Bütikofer, U. CLA isomers in milk fat from cows fed diets with high levels of unsaturated fatty acids. Lipids 2004, 39, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Loor, J.J.; Doreau, M.; Chardigny, J.M.; Ollier, A.; Sebedio, J.L.; Chilliard, Y. Effects of ruminal or duodenal supply of fish oil on milk fat secretion and profiles of trans-fatty acids and conjugated linoleic acid isomers in dairy cows fed maize silage. Anim. Feed Sci. Technol. 2005, 119, 227–246. [Google Scholar] [CrossRef]
- Massart-Leën, A.M.; Peeters, G.; Vandeputte-Van Messom, G.; Roets, E.; Burvenich, C. Effects of valerate and isobutyrate on fatty acid secretion by the isolated perfused mammary gland of the lactating goat. Reprod. Nutr. Dev. 1986, 26, 801–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, B.; West, J.A.; Koulman, A. A review of odd-chain fatty acid metabolism and the role of pentadecanoic Acid (c15:0) and heptadecanoic Acid (c17:0) in health and disease. Molecules 2015, 20, 2425–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodge, A.M.; English, D.R.; O’Dea, K.; Sinclair, A.J.; Makrides, M.; Gibson, R.A.; Giles, G.G. Plasma phospholipid and dietary fatty acids as predictors of type 2 diabetes: Interpreting the role of linoleic acid. Am. J. Clin. Nutr. 2007, 86, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venn-Watson, S.; Lumpkin, R.; Dennis, E.A. Efficacy of dietary odd-chain saturated fatty acid pentadecanoic acid parallels broad associated health benefits in humans: Could it be essential? Sci. Rep. 2020, 10, 8161. [Google Scholar] [CrossRef]
- Bas, P.; Archimède, H.; Rouzeau, A.; Sauvant, D. Fatty acid composition of mixed-rumen bacteria: Effect of concentration and type of forage. J. Dairy Sci. 2003, 86, 2940–2948. [Google Scholar] [CrossRef] [Green Version]
- Piccione, G.; Casella, S.; Lutri, L.; Vazzana, I.; Ferrantelli, V.; Caola, G. Reference values for some haematological, haematochemical, and electrophoretic parameters in the Girgentana goat. Turk. J. Vet. Anim. Sci. 2010, 34, 197–204. [Google Scholar]
- Tudisco, R.; Musco, N.; Pero, M.E.; Morittu, V.M.; Grossi, M.; Mastellone, V.; Cavaliere, G.; Wanapat, M.; Infascelli, F.; Lombardi, P. Influence of dietary hydrogenated palm oil supplementation on serum biochemistry and progesterone levels in dairy goats. Anim. Nutr. 2019, 5, 286–289. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Composition g/kg DM | CTR Concentrate | LIN Concentrate | Linseed | Oat Hay | Pasture |
|---|---|---|---|---|---|
| CP | 177.4 ± 4.75 | 178.6 ± 3.69 | 242.3 ± 5.83 | 88.4 ± 5.08 | 160 ± 20.04 |
| EE | 28.70 ± 2.66 | 67.40 ± 1.36 | 350.2 ± 7.45 | 19.4 ± 1.84 | 20.3 ± 1.40 |
| NDF | 262.5 ± 6.13 | 266.9 ± 8.51 | 259.3 ± 7.12 | 594.5 ± 12.35 | 465.9 ± 50.12 |
| ADF | 98.60 ± 6.90 | 101.4 ± 7.04 | 112.1 ± 3.41 | 312.7 ± 9.270 | 338.4 ± 15.25 |
| ADL | 26.70 ± 1.33 | 27.30 ± 1.51 | 54.10 ± 2.11 | 39.6 ± 2.98 | 50.0 ± 7.19 |
| PDIN g/kg DM | 116.0 ± 3.66 | 100.5 ± 2.90 | 168.3 ± 3.86 | 62.3 ± 4.52 | 112.2 ± 12.31 |
| PDIE g/kg DM | 100.8 ± 2.72 | 104.4 ± 3.11 | 133.6 ± 2.98 | 64.5 ± 3.47 | 76.4 ± 9.90 |
| Fatty acid profile % of total FA | |||||
| SFA | 28.4 ± 1.13 | 26.6 ± 1.22 | 10.9 ± 0.31 | . | 17.7 ± 2.22 |
| MUFA | 15.8 ± 0.65 | 14.3 ± 0.25 | 22.2 ± 1.64 | . | 6.3 ± 0.74 |
| PUFA | 55.8 ± 2.61 | 59.12 ± 4.16 | 66.9 ± 2.51 | . | 76.0± 8.36 |
| C18:2 | 48.7 ± 2.34 | 37.9 ± 2.19 | 13.7 ± 0.78 | . | 26.4 ± 6.70 |
| C18:3 | 3.50 ± 0.12 | 18.3 ± 1.43 | 53.2 ± 4.82 | . | 41.9 ± 5.76 |
| Concentrates ingredients % as fed | CTR | LIN | |||
| Soft wheat bran 26.6; Corn meal 15.0; Sunflower meal 14.5; Dried pulp beet 12.0; Fava bean 10.6; Corn gluten feed 7.0; Dried citrus pulp 6.5; Molasses 5.6; Vitamin-mineral premix 2.2 | Soft wheat bran 30.0; Corn meal 23.0; Linseed 20.0; Dried citrus pulp 10.0; Dried pulp beet 8.0; Corn gluten feed 7.0; Vitamin–mineral premix 2.0. |
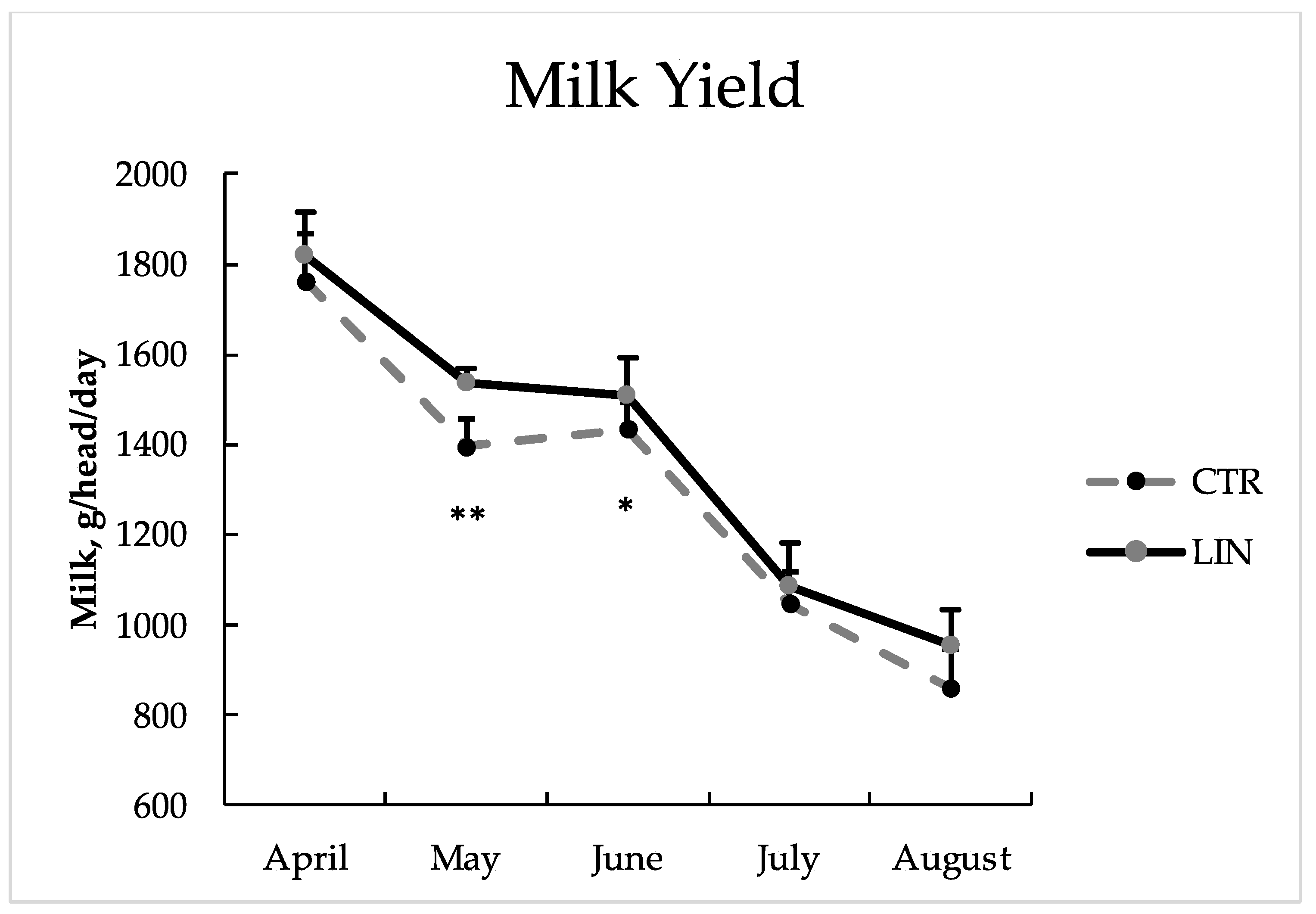
| Milk Production | Fat | Protein | Lactose | |||||
|---|---|---|---|---|---|---|---|---|
| CTR | LIN | CTR | LIN | CTR | LIN | CTR | LIN | |
| Mean value | 1291.38 | 1358.72 | 40.77 | 57.13 | 42.28 | 46.75 | 52.96 | 56.24 |
| April | 1763.33 | 1818.33 | 56.42 | 63.28 | 58.90 | 59.46 | 75.47 | 84.01 |
| May | 1395.20 | 1536.39 | 51.34 | 72.52 | 45.76 | 52.85 | 61.39 | 65.60 |
| June | 1434.17 | 1506.67 | 45.75 | 70.51 | 44.89 | 50.32 | 56.22 | 60.27 |
| July | 1046.67 | 1085.21 | 30.25 | 37.77 | 33.28 | 35.70 | 40.30 | 41.13 |
| August | 858.33 | 953.33 | 23.43 | 41.37 | 28.67 | 34.32 | 33.73 | 35.27 |
| RMSE | 86.004 | 8.14 | 3.74 | 2.81 | ||||
| p-value | ||||||||
| Group | *** | *** | NS | NS | ||||
| Time | *** | *** | NS | *** | ||||
| GxT | NS | NS | NS | * | ||||
| CTR | LIN | RMSE | p-Value | |||
|---|---|---|---|---|---|---|
| Group (G) | Time (T) | GxT | ||||
| SFA | 73.19 | 69.47 | 3.452 | *** | *** | NS |
| MUFA | 23.43 | 26.10 | 2.994 | *** | *** | NS |
| PUFA | 3.32 | 4.20 | 0.454 | *** | *** | *** |
| PUFA n3 | 1.16 | 1.38 | 0.177 | *** | *** | *** |
| PUFA n6 | 1.81 | 2.20 | 0.263 | *** | *** | *** |
| n6/n3 | 1.54 | 1.63 | 0.239 | NS | *** | *** |
| C18:2 (c9, t11) | 0.338 | 0.610 | 0.1648 | *** | ** | ** |
| C18:2 (t10, c12) n6 | 0.091 | 0.108 | 0.1184 | NS | NS | NS |
| CLAs | 0.429 | 0.735 | 0.243 | *** | ** | ** |
| CTR | LIN | RMSE | p-Value | |||
|---|---|---|---|---|---|---|
| Group (G) | Time (T) | GxT | ||||
| iso C13:0 | 0.028 | 0.034 | 0.007 | *** | NS | NS |
| iso C14:0 | 0.088 | 0.094 | 0.013 | NS | NS | NS |
| iso C15:0 | 0.176 | 0.193 | 0.038 | NS | NS | NS |
| iso C16:0 | 0.024 | 0.029 | 0.037 | NS | * | NS |
| iso C17:0 | 0.027 | 0.034 | 0.049 | NS | ** | NS |
| anteiso C:13 | 0.0032 | 0.0040 | 0.004 | NS | NS | NS |
| anteiso C:15 | 0.363 | 0.383 | 0.006 | NS | NS | NS |
| anteiso C:17 | 0.391 | 0.404 | 0.083 | NS | NS | NS |
| C15:0 | 0.823 | 0.609 | 0.053 | *** | *** | *** |
| C17:0 + cis9 C17:1 | 0.924 | 0.676 | 0.091 | *** | *** | *** |
| Total OBCFA | 3.21 | 2.84 | 0.203 | *** | *** | *** |
| AST | ALT | GGT | ALP | GLU | TP | CREA | BUN | CHOL | TRY | |
|---|---|---|---|---|---|---|---|---|---|---|
| U/L | U/L | U/L | U/L | mg/dL | g/dL | mg/dL | mg/dL | mg/dL | mg/dL | |
| CTR | 64.74 | 20.94 | 34.55 | 208.17 | 51.57 | 6.97 | 0.500 | 36.74 | 61.14 | 18.49 |
| LIN | 68.28 | 22.07 | 36.44 | 215.63 | 54.12 | 6.95 | 0.422 | 38.07 | 61.52 | 24.34 |
| RMSE | 7.333 | 2.473 | 5.121 | 39.542 | 5.934 | 0.564 | 0.142 | 5.237 | 8.261 | 4.852 |
| Group (G) | NS | NS | NS | NS | NS | NS | NS | NS | NS | ** |
| Time (T) | NS | *** | NS | NS | NS | NS | ** | ** | NS | NS |
| GxT | NS | NS | NS | NS | NS | NS | * | NS | NS | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musco, N.; Tudisco, R.; Esposito, G.; Iommelli, P.; Totakul, P.; D’Aniello, B.; Lombardi, P.; Amato, R.; Wanapat, M.; Infascelli, F. Effects of Linseed Supplementation on Milk Production, Composition, Odd- and Branched-Chain Fatty Acids, and on Serum Biochemistry in Cilentana Grazing Goats. Animals 2022, 12, 783. https://doi.org/10.3390/ani12060783
Musco N, Tudisco R, Esposito G, Iommelli P, Totakul P, D’Aniello B, Lombardi P, Amato R, Wanapat M, Infascelli F. Effects of Linseed Supplementation on Milk Production, Composition, Odd- and Branched-Chain Fatty Acids, and on Serum Biochemistry in Cilentana Grazing Goats. Animals. 2022; 12(6):783. https://doi.org/10.3390/ani12060783
Chicago/Turabian StyleMusco, Nadia, Raffaella Tudisco, Giulia Esposito, Piera Iommelli, Pajaree Totakul, Biagio D’Aniello, Pietro Lombardi, Ruggero Amato, Metha Wanapat, and Federico Infascelli. 2022. "Effects of Linseed Supplementation on Milk Production, Composition, Odd- and Branched-Chain Fatty Acids, and on Serum Biochemistry in Cilentana Grazing Goats" Animals 12, no. 6: 783. https://doi.org/10.3390/ani12060783